
Abstract
This study was aimed at examining whether rapid portal vein infusion (RPVI) of a small volume of naked oligodeoxynucleotides (ODNs) could be used to transfect sufficient amounts of nuclear factor-κB (NF-κB) decoy ODN into the liver to suppress NF-κB activation during liver ischemia–reperfusion (I/R) injury, in which NF-κB plays a central role in regulating the production of inflammatory cytokines. One milliliter of naked NF-κB decoy ODN solution was administered into the portal vein for a few seconds. Transfection efficacy was examined by labeling the ODN with a fluorescent tag. Activation of NF-κB was investigated by electrophoretic mobility shift assay. Levels of serum liver enzymes and cytokines were measured during liver I/R injury. NF-κB decoy ODN was preferentially incorporated into Kupffer cells and sinusoidal endothelial cells, but not hepatocytes, in the rat liver. Transfected NF-κB decoy ODN suppressed the function of NF-κB in both Kupffer cells and sinusoidal endothelial cells during liver I/R injury, causing significant decreases in serum tumor necrosis factor-α and interleukin-6 levels 3 hr after reperfusion. Although the decrease in serum liver enzymes was not significant, naked NF-κB decoy ODN was successfully incorporated into Kupffer cells and sinusoidal endothelial cells by rapid portal vein infusion, inhibited NF-κB activation in both cells, and suppressed the production of inflammatory cytokines during the early phase of liver I/R injury.
Introduction
Liver ischemia–reperfusion (I/R) injury after liver transplantation and liver surgery with prolonged vascular occlusion remains a crucial clinical problem (Clavien et al., 1992; Liu et al., 1996). During ischemia, liver is insulted by necrotic injury because of the shortage of oxygen and energy. On reperfusion, reactive oxygen species (ROSs) are released by hepatocytes and Kupffer cells, and make Kupffer cells and sinusoidal endothelial cells activated during the early phase of I/R (Rauen et al., 1994; Bautista, 2002). These cells play a central role in the production of several inflammatory cytokines, such as tumor necrosis factor (TNF)-α and interleukin (IL)-1β (Suzuki and Toledo-Pereyra, 1994). Secreted TNF-α in turn initiates a cascade of events leading to liver injury (Wanner et al., 1996; Lentsch et al., 2000; Serracino-Inglott et al., 2001; Jaeschke, 2003; Montalvo-Jave et al., 2008).
Nuclear factor (NF)-κB is a transcription factor that predominantly regulates gene expression of some inflammatory cytokines, chemokines, and adhesion molecules, which are known to be strongly associated with liver I/R injury (Tran-Thi et al., 1995; Lentsch et al., 1998; Zwacka et al., 1998; Pahl, 1999). In the steady state, NF-κB is located in the cytoplasm and is prevented from translocation into the nucleus by forming a complex with inhibitor-κB (I-κB) (Baeuerle and Baltimore, 1988; Regnier et al., 1997). On cell activation, I-κB is dissociated from the complex and allows NF-κB to translocate into the nucleus, where it binds to κB sites in the regulatory region of the relevant genes (Lenardo et al., 1987) and activates their transcription.
NF-κB decoy oligodeoxynucleotide (ODN) with the same sequence as the NF-κB-binding site (κB site) reportedly inhibited binding of NF-κB to its target genes. The organ-protective action of NF-κB decoy ODN was first discovered in myocardial infarction (Morishita et al., 1997) and was suggested to suppress NF-κB function in endothelial cells. Hepatic injury induced by endotoxin (Ogushi et al., 2003) or carbon tetrachloride (Son et al., 2007) can be ameliorated by NF-κB decoy ODN-mediated suppression of Kupffer cell activation. However, it is not yet clear whether NF-κB decoy ODN can attenuate liver I/R injury.
The present study was performed to examine whether rapid portal vein infusion (RPVI) of naked NF-κB decoy ODN could be used to transfect the ODN into liver, and investigated whether this strategy feasibly worked in vivo during liver I/R injury as a model of NF-κB activation in liver.
Materials and Methods
Sequences and preparation of ODNs
Sequences of the synthetic phosphorothionate ODNs (Greiner, Tokyo, Japan) were 5′-CCTTGAA
Animals and transfection of ODNs
The Animal Committee at Akita University Graduate School of Medicine (Akita, Japan) approved the protocols for animal experimentation described in this paper, and animal experiments were performed in strict adherence to our institutional guidelines for animal experimentation.
Eight-week-old male Sprague-Dawley rats (SLC, Hamamatsu, Japan) were anesthetized, and either naked NF-κB decoy ODN (NF-κB decoy) or scrambled decoy ODN (SC decoy) solution (50 nmol/rat), diluted in 1 ml of Ringer's solution (Otsuka Pharmaceutical, Tokyo, Japan), was infused manually into the portal vein over a period of a few seconds (2–3 sec). Vehicle without any ODN (1 ml of Ringer's solution) was administered to rats in the control group (Nil decoy). There were no deaths or signs of toxicity in any of the groups.
Twenty-four hours after rapid portal vein infusion (RPVI), the portal triad was clamped for 45 min with a microvascular clip (Aesculap, Tuttlingen, Germany), and the rats were killed 1, 3, and 6 hr after removal of the clip. Lipopolysaccharide (LPS)-induced liver injury models were prepared as positive controls for NF-κB activation (Muller et al., 1993). Samples were collected 6 hr after intraperitoneal injection of LPS (Sigma-Aldrich, St. Louis, MO) at a dose of 20 mg/kg body weight (Mercer et al., 1996) in the absence of any ODN or vehicle infusion into the portal vein.
Measurement of fluorescence intensity in liver and immunofluorescence staining
FITC–ODN was infused by RPVI and livers were removed 24 hr after infusion. To measure fluorescence intensity in liver, liver tissues were homogenized and centrifuged at 1000×g for 10 min, and the supernatants were examined with a fluorescence meter with excitation and emission wavelengths of 494 and 518 nm, respectively (Hitachi, Tokyo, Japan). Protein concentrations in the supernatants were measured, and the results are shown as fluorescence intensity per milligram of protein. For immunofluorescence staining, frozen liver sections were incubated with anti-CD68 antibody (Serotec, Oxford, UK), anti-hepatic sinusoidal endothelial cell (HSEC) antibody (American Research Products, Belmont, MA), or anti-albumin antibody (Cappel, Aurora, OH), followed by incubation with Alexa Fluor 594-conjugated anti-mouse or anti-rabbit IgG antibody (Molecular Probes/Life Technologies, Eugene, OR). The distribution of fluorochrome was analyzed by confocal laser microscopy (Carl Zeiss, Jena, Germany).
Fractionation of liver cells
After 45 min of ischemia and 1 hr of reperfusion, livers were fractionated into Kupffer cell, sinusoidal endothelial cell, and hepatocyte fractions by collagenase digestion and elutriation (Kono et al., 2002). Briefly, livers were perfused with Ca2+, Mg2+-free Hanks' balanced salt solution (Sigma-Aldrich) containing 0.5 mM EDTA, and then digested in type I collagenase solution (Wako, Osaka, Japan). Liver cells were centrifuged at 50×g for 1 min at 4°C, and the pellet was collected as the hepatocyte fraction. The supernatant was again centrifuged at 350×g for 10 min, and the cell pellet was incubated in 1% actinase E (Kaken, Tokyo, Japan), 0.25% trypsin (Becton Dickinson, Sparks, MD), and 0.05% DNase I (Worthington, Lakewood, NJ) for 10 min at 37°C. The cell suspension was layered on top of 50% Percoll (Amersham, Uppsala, Sweden) solution and centrifuged at 900×g for 20 min. The cells above the 50% Percoll solution were collected and applied to an elutriator. Counterflow centrifugal elutriation (CR21G centrifuge and R5E elutriation rotor; Hitachi) at a constant speed (3250 rpm) and different flow rates was used for separation of sinusoidal endothelial cell and Kupffer cell fractions, which were obtained at flow rates of 22–28 and 40–70 ml/min, respectively. The temperature during elutriation was kept at 4°C. Cell viability as determined by trypan blue exclusion test was >85% in the Kupffer cell fraction and >70% in the sinusoidal endothelial cell fraction. The purity of the cell fractions was determined by immunofluorescence staining, and was ≥75% in both fractions.
Electrophoretic mobility shift assay
The probe of the same sequences as NF-κB decoy ODN was labeled with [γ-32P]ATP, using T4 polynucleotide kinase (Takara, Otsu, Japan). Nuclear protein (3 μg) prepared from the cell fractions was reacted with 50 fmol of radiolabeled probe for 30 min. Reaction mixtures were incubated with an additional 1 μg of anti-NF-κB p65 antibody (Santa Cruz Biotechnology, Santa Cruz, CA) for the supershift assay, or with a 200-fold molar excess of the unlabeled probe for the competition assay. The samples were electrophoresed on polyacrylamide gels and visualized by autoradiography.
Measurements of serum liver enzymes and cytokines
We initially planned to evaluate the effects of NF-κB decoy ODN after 45 min of ischemia, and 3 or 6 hr of reperfusion. However, the 45 min of ischemia was too harsh and most rats did not survive longer than 2 hr. Therefore, the effects of NF-κB decoy ODN on liver function were examined in a 30-min ischemia model. Aspartate aminotransferase (AST), alanine aminotransferase (ALT), and lactate dehydrogenase (LDH) were measured in serum samples, using commercial kits (Wako). Serum cytokines, TNF-α (BioSource/Life Technologies, Camarillo, CA), IL-1β (R&D Systems, Minneapolis, MN), IL-6 (R&D Systems), and growth-related oncogene/cytokine-induced neutrophil chemoattractant (GRO/CINC)-1 (rat IL-8 homolog; Amersham) were quantified with ELISA kits.
Specific esterase histochemical staining
Liver specimens were stained with a kit (Sigma-Aldrich) containing α-naphthyl acetate and Fast Blue RR salt. Stained cells in the sinusoidal space were counted as granulocytes in 50 randomly chosen high-power fields (HPFs) ( ×400) per specimen. Results are shown as the total granulocyte number in 50 HPFs.
Statistical analysis
All data are presented as means±SD. An unpaired t test and analysis of variance (ANOVA) were used for statistical analysis. In all analyses, p<0.05 was taken to indicate statistical significance.
Results
RPVI can be used to transfect naked NF-κB decoy ODN into Kupffer cells and sinusoidal endothelial cells
We first examined whether naked FITC–ODN could be incorporated into liver by RPVI. Fluorescence microscopy showed a wide distribution of fluorochrome in liver 24 hr after RPVI (Fig. 1A) compared with the control (Fig. 1B). FITC–ODN incorporation was also confirmed by measuring the fluorescence intensity in liver homogenates (Fig. 1C). The fluorescence intensity of the FITC–ODN RPVI group (83.7±10.6) was significantly higher than that of the control (34.4±4.2).
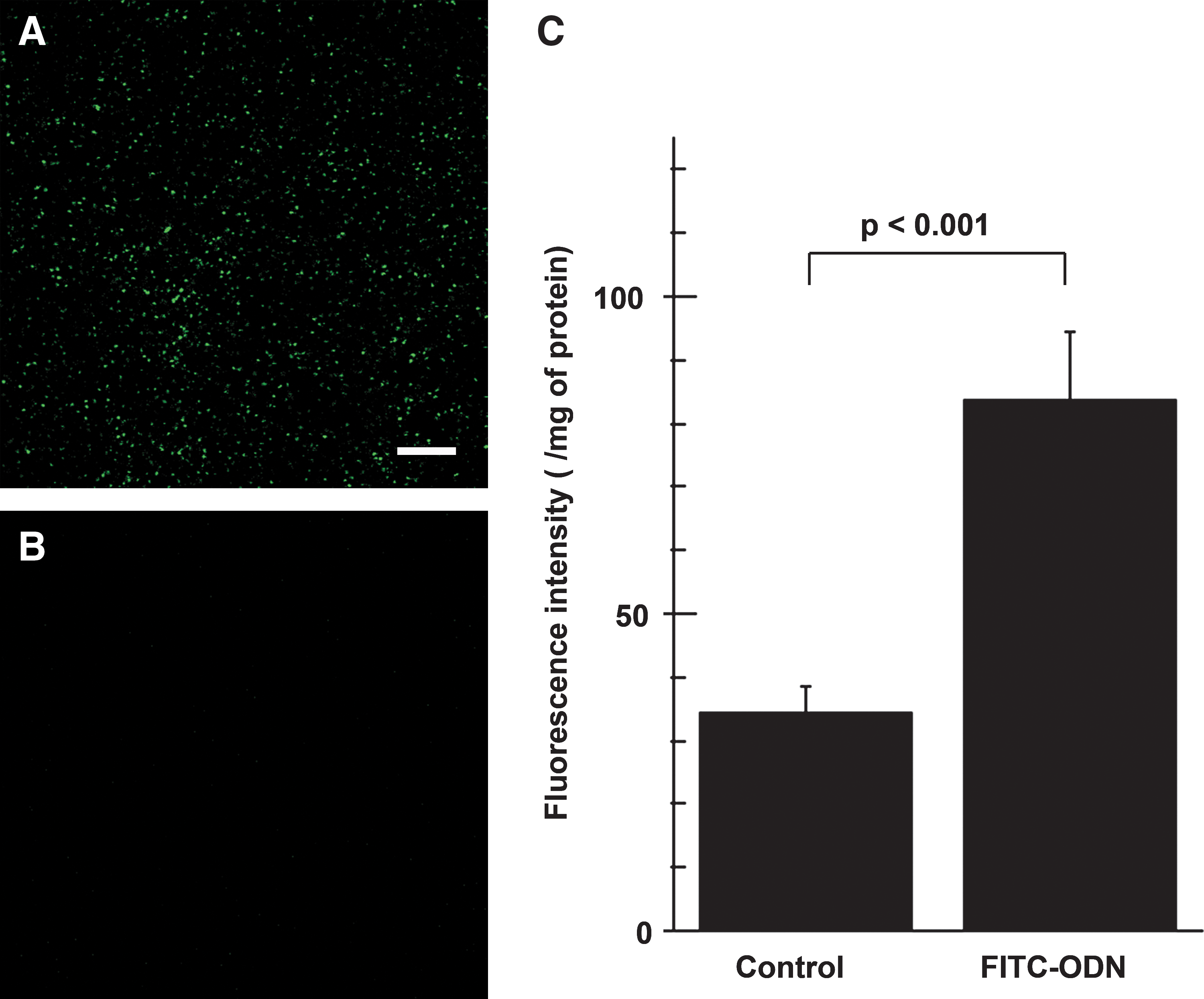
Naked FITC–ODN could be incorporated into liver by rapid portal vein infusion (RPVI). Frozen liver sections 24 hr after infusion of FITC–ODN
Immunofluorescence staining showed that FITC–ODN was localized in the cells stained positively for CD68 or HSECs (Fig. 2, top and middle). In contrast, there was no FITC fluorescence in hepatocyte cytoplasm (Fig. 2, bottom). These results indicated that the ODN infused by RPVI was incorporated into Kupffer cells and sinusoidal endothelial cells, but not hepatocytes.
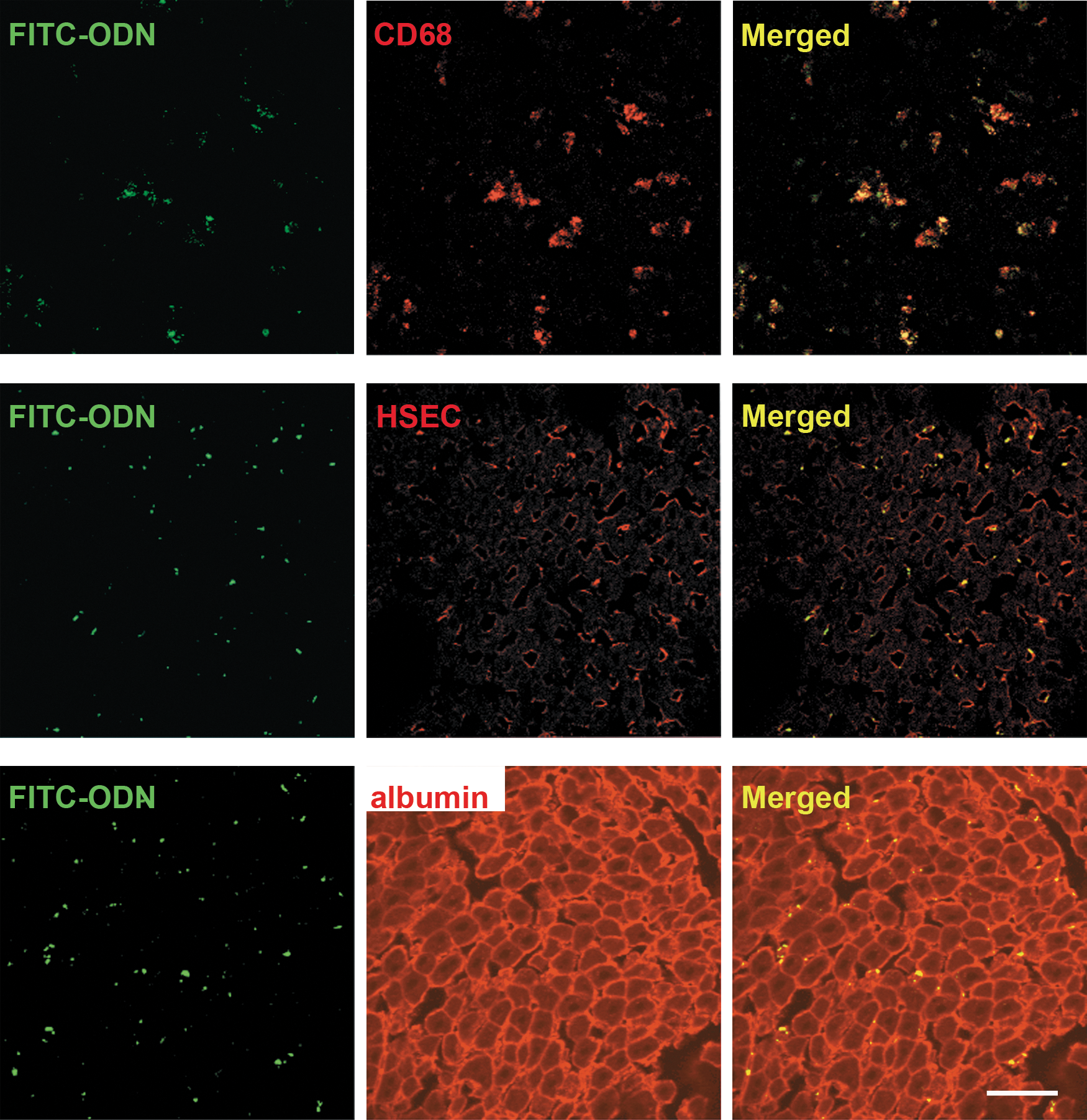
Naked FITC–ODN could be incorporated into Kupffer cells and sinusoidal endothelial cells. Frozen liver sections 24 hr after infusion of FITC–ODN by the RPVI method were immunostained with Alexa Fluor 594 labeling for CD68 (top), hepatic sinusoidal endothelial cells (HSECs; middle), or albumin (bottom). Distributions of FITC–ODN in liver were analyzed by confocal laser microscopy (original magnification, ×400). Scale bar: 50 μm.
Naked NF-κB decoy ODN suppresses NF-κB activation
Nuclear protein prepared from liver cell fractions after 45 min of ischemia and 1 hr of reperfusion was analyzed by electrophoretic mobility shift assay (EMSA). The bands indicated by arrows in Fig. 3 correspond to NF-κB because these bands were supershifted in the presence of anti-NF-κB p65 antibodies and competitively inhibited binding to the radiolabeled probes by an excess of unlabeled probe.
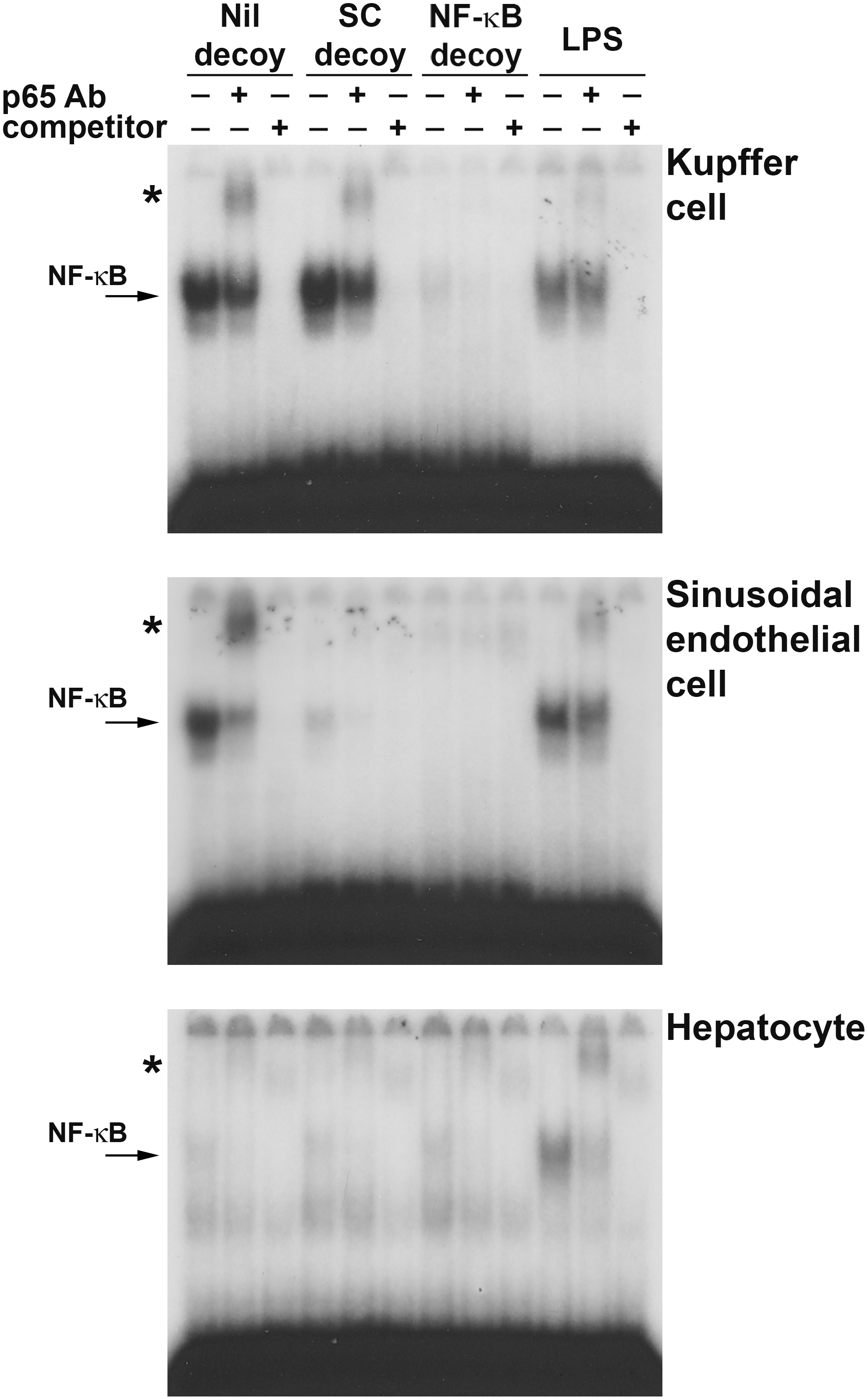
NF-κB decoy ODN could suppress NF-κB activation during liver I/R injury. Rats infused with NF-κB decoy ODN (NF-κB decoy), scrambled decoy ODN (SC decoy), or vehicle without decoy ODN (Nil decoy) by the RPVI method were injured by 45 min of ischemia and 1 hr of reperfusion. Livers were fractionated into Kupffer cell, sinusoidal endothelial cell, and hepatocyte fractions, and the nuclear protein of each fraction was subjected to EMSA. NF-κB (arrow) was confirmed by supershift (*) assay with anti-NF-κB p65 antibody (p65 Ab) and by competitive assay with an excess of competitor. Livers injured by lipopolysaccharide administration (LPS) were used as positive controls of activated NF-κB.
In Kupffer cell fractions (Fig. 3, top), NF-κB was strongly activated by I/R in the Nil decoy group and the scrambled decoy group (SC decoy). However, NF-κB was almost suppressed in the presence of NF-κB decoy ODN (NF-κB decoy). The scrambled decoy ODN had no inhibitory effect on NF-κB activity.
In sinusoidal endothelial cells (Fig. 3, middle), the results of the Nil decoy group and the NF-κB decoy group were similar to those of the Kupffer cell fraction. However, scrambled decoy ODN also suppressed NF-κB activity. This was unexpected, but was confirmed in experiments using another set of liver cell fractions and another scrambled decoy ODN sequence. These reproducible results suggested that the naked ODN itself suppressed NF-κB activity in sinusoidal endothelial cells regardless of its sequence.
In hepatocytes (Fig. 3, bottom), as the activation of NF-κB per se was not strong even in the Nil decoy group, NF-κB activity was equivalent in all groups.
Naked NF-κB decoy ODN suppresses cytokine production
Serum liver enzyme and cytokine measurements were performed, using blood samples from rats insulted by 45 min of ischemia and 1 hr of reperfusion (Fig. 4). At this time point, elevated serum liver enzyme levels showed no statistically significant difference, although they did show a tendency to be lower in the NF-κB decoy group. Production of TNF-α was also better suppressed in the NF-κB decoy group, but the difference was not significant. Serum IL-6 concentrations were almost similar among the three groups. All liver enzyme levels rose further in the Nil decoy group and the SC decoy group 3 hr postreperfusion, but serum enzyme levels in the NF-κB decoy group remained relatively low although the differences were not statistically significant (Fig. 5). Serum concentrations of TNF-α and IL-6 increased further in the Nil decoy group and the SC decoy group. On the other hand, those of the NF-κB decoy group decreased. As a result, serum concentrations of TNF-α and IL-6 were significantly lower in the NF-κB decoy group (1.5±3.2 and 329.0±307.4, respectively) than in the Nil decoy group (18.8±10.4 and 1567.4±886.5, respectively). The serum IL-1β level was also lower in the NF-κB decoy group than in the Nil decoy group. Six hours after reperfusion, all liver enzyme levels decreased and the effect of NF-κB decoy ODN was no longer evident (Fig. 6). Regarding serum GRO/CINC-1 and the numbers of accumulated granulocytes in the sinusoidal space, there were no differences in serum GRO/CINC-1, but the granulocyte number was slightly reduced in the NF-κB decoy group compared with the Nil decoy group. These results indicated that naked NF-κB decoy ODN introduced by RPVI attenuated NF-κB-mediated inflammatory cytokine production at 3 hr after reperfusion.
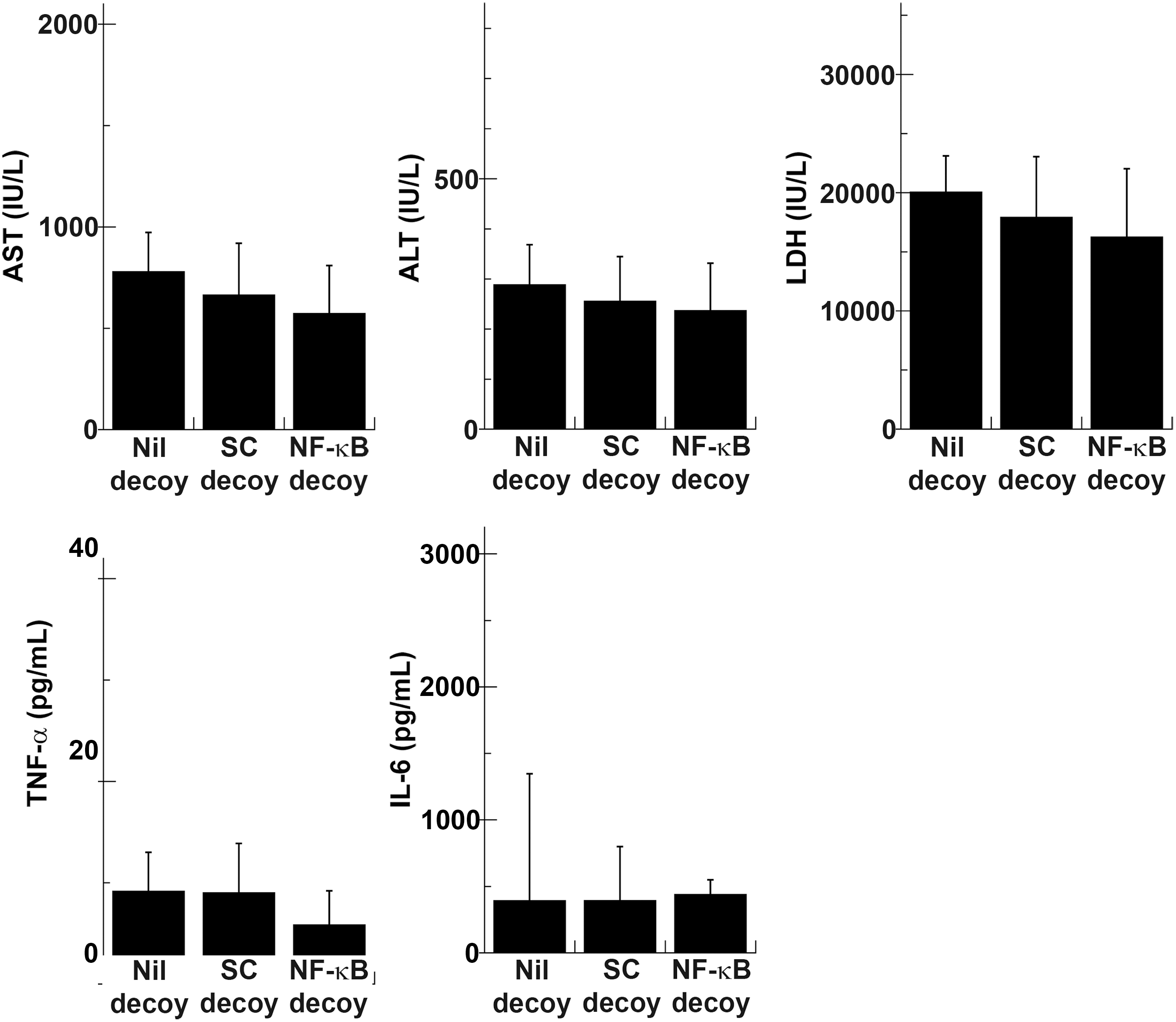
NF-κB decoy ODN did not significantly affect serum liver enzyme levels or cytokine production 1 hr postreperfusion after 45 min of ischemia. Rats infused with NF-κB decoy ODN (NF-κB decoy), scrambled decoy ODN (SC decoy), or vehicle without decoy ODN (Nil decoy) by the RPVI method were injured by 45 min of ischemia and 1 hr of reperfusion. Serum liver enzyme and cytokine levels were measured (n=10). ALT, alanine aminotransferase; AST, aspartate aminotransferase; LDH, lactate dehydrogenase; TNF-α, tumor necrosis factor-α.
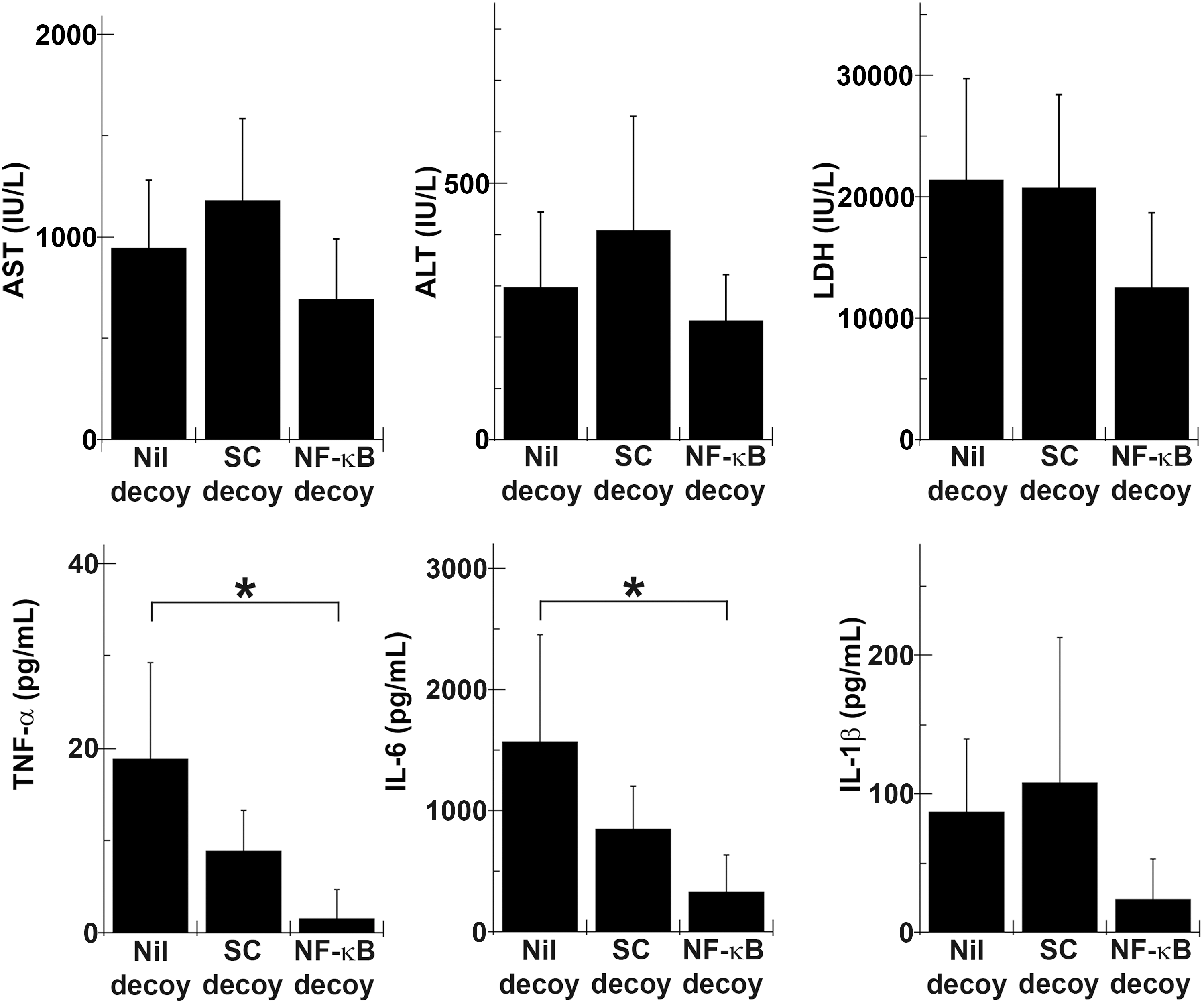
NF-κB decoy ODN suppressed serum cytokine production 3 hr postreperfusion after 30 min of ischemia. Rats infused with NF-κB decoy ODN (NF-κB decoy), scrambled decoy ODN (SC decoy), or vehicle without decoy ODN (Nil decoy) by the RPVI method were injured by 30 min of ischemia and 3 hr of reperfusion. Serum liver enzyme and cytokine levels were measured (n=5). *p<0.05.
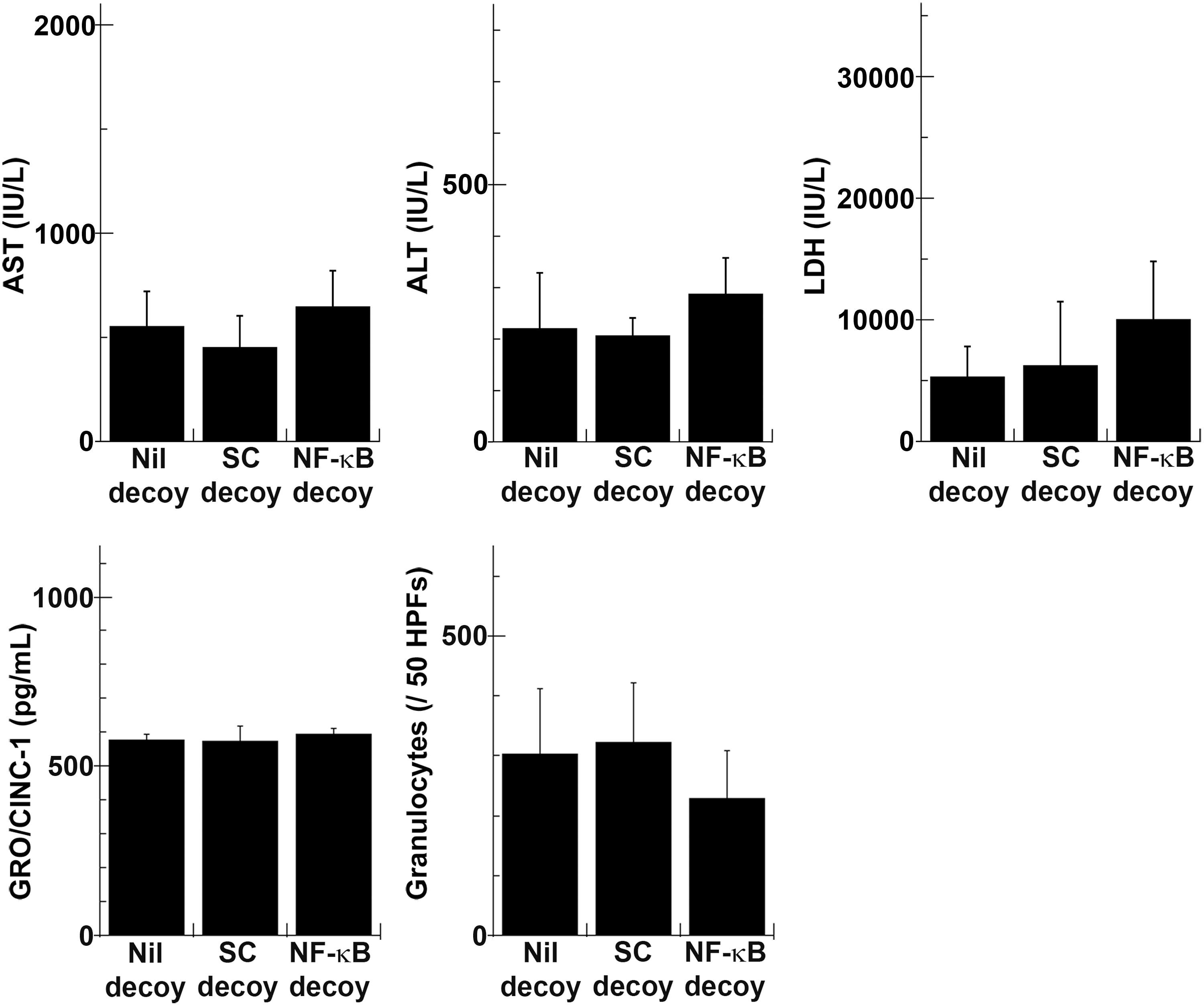
NF-κB decoy ODN augmented liver injury, and did not affect the serum growth-related oncogene/cytokine-induced neutrophil chemoattractant (GRO/CINC)-1 level or the number of granulocytes in the liver sinusoidal space 6 hr postreperfusion after 30 min of ischemia. Rats infused with NF-κB decoy ODN (NF-κB decoy), scrambled decoy ODN (SC decoy), or vehicle without decoy ODN (Nil decoy) by the RPVI method were injured by 30 min of ischemia and 6 hr of reperfusion. Serum liver enzyme levels and GRO/CINC-1 concentration were measured (n=5). Liver specimens were subjected to specific esterase histochemical staining, and positively stained cells in the liver sinusoidal space were counted. Results are shown as the total granulocyte number in 50 randomly chosen high-power fields (HPFs; original magnification, ×400) (n=5).
The degree of liver injury was also evaluated on hematoxylin–eosin (H&E) staining; however, no particular difference was detected throughout the time course (Supplementary Fig. S1; supplementary data are available online at
Discussion
The results of this study indicated that the RPVI method can be used to transfect naked NF-κB decoy ODN into Kupffer cells and sinusoidal endothelial cells, and that incorporated NF-κB decoy ODN inhibited NF-κB activation. It was reported previously that transfection of NF-κB decoy ODN into liver, using cationic liposomes, ameliorated endotoxin-induced liver failure (Ogushi et al., 2003) and carbon tetrachloride-induced liver injury (Son et al., 2007). However, even cationic liposomes, which are regarded as a safe nonviral vector, induced the innate immune response and the production of proinflammatory cytokines (Sellins et al., 2005). Therefore, delivery of naked NF-κB decoy ODN into liver is potentially useful.
In comparison with the delivery of naked ODN, the procedures used to deliver naked genes larger than ODN into liver have been studied in some detail. Budker and colleagues reported the expression of various gene products in mouse hepatocytes after intraportal injection of 1 ml or ∼4% body weight of naked plasmid DNA solution at a flow rate of 2 ml/min with hypertonic solution and hepatic outflow clamping (Budker et al., 1996). This group also reported that rats tolerated the intraportal injection of a volume corresponding to 5–7.5% of body weight (Zhang et al., 1997). Liu and colleagues (1999) and Zhang and colleagues (1999) reported hydrodynamic gene delivery by systemic injection of a large volume of naked DNA solution (∼10% body weight) via the tail vein of mice at a rate of 6–20 ml/min; they showed high levels of gene delivery into hepatocytes. In rats, the efficacy was reported to be highest at 10% body weight at 100 ml/min (Maruyama et al., 2002). However, this method does not seem to be optimal for in vivo use. Sawyer and colleagues reported that hydrodynamic gene delivery was too harsh, with even a volume of 1% body weight at 100 ml/min causing severe decreases in both blood pressure and heart rate. Although the effects on vital parameters were transient when the injection volume was 1–2% body weight, injection of a volume exceeding 4% body weight resulted in prolonged reduction of blood pressure and arrhythmia (Sawyer et al., 2007). In comparison with the intraportal or systemic injection method used in these previous experiments, the volume reported here was small, corresponding to only ∼0.5% body weight, and was infused by the RPVI method at a rate of 20–30 ml/min. The effect on cardiac function would be much lower with our method, compared with the previous ones, because of the much lower volume used. In addition, despite rapid infusion, perfusion resistance at the liver sinusoid may compensate for the volume load on cardiac function.
Interestingly, with the RPVI method, Kupffer cells and sinusoidal endothelial cells were preferentially transfected; few hepatocytes were transfected. This transfection pattern was different from that of hydrodynamic gene delivery or intraportal injection with hepatic outflow clamping. The exact cellular trafficking of ODN is still under debate, but Liu and colleagues reported that naked DNA was taken up primarily by liver endothelial cells, not by Kupffer cells, during a single pass after simple plasmid DNA intravenous injection (Liu et al., 2007). Efficient incorporation of naked ODN into Kupffer cells requires two clearly distinct anatomical conditions (Jacobs et al., 2010): migration to the space of Disse through sinusoidal fenestrae and contact with Kupffer cells in the space of Disse. We supposed that the pressure caused by the RPVI method might allow naked ODN to pass through sinusoidal fenestrae. However, it remained unclear why naked ODN in the space of Disse was not incorporated into hepatocytes. One possible reason is the difference in phagocytic activity between hepatocytes and Kupffer cells.
Another interesting, rather curious, finding of the present study was that scrambled decoy ODN inhibited NF-κB function in sinusoidal endothelial cells. This phenomenon was limited to sinusoidal endothelial cells and was not observed in Kupffer cells. These observations were reproducible in other sets of rats and with another scrambled decoy ODN sequence (5′-GCTTCGACCTCGATTCCGAT-3′, 3′-CGAAGCTGGAGCTAAGGCTA-5′). A similar, unexpected phenomenon has been described in skin keratinocytes. Dorn and colleagues showed that 16- to 20-mer ODN was incorporated into cultured skin keratinocytes and caused the suppression of IL-8 production in a sequence-independent manner, suggesting posttranscriptional regulation by ODN (Dorn et al., 2007). However, the results of the present study cannot be explained by their suggestion that any ODN of this size regulates gene expression at the posttranscriptional level. In Kupffer cells, NF-κB decoy ODN used in this study undoubtedly caused the suppression of NF-κB activation through transcriptional regulation. Does this strange finding arise from the peculiarity of sinusoidal endothelial cells or for other reasons? Further studies are required to resolve this issue.
Previous studies indicated that NF-κB decoy ODN ameliorated several types of liver injury (Ogushi et al., 2003; Xu et al., 2005; Son et al., 2007; Hoffmann et al., 2009). However, with respect to inhibition of NF-κB activation, these previous reports provided only immunohistochemical images showing a decreased number of cell nuclei stained for NF-κB. Here, we directly demonstrated that the level of nuclear NF-κB with DNA-binding activity was certainly decreased in the isolated Kupffer cells and sinusoidal endothelial cells (Fig. 3). For methodological reasons, that is, because cells were isolated from liver after I/R injury in the present study, the cell purity (≥75%) and viability (>70%) of both Kupffer and sinusoidal endothelial cell fractions were limited. However, they were acceptably high for checking NF-κB activity by EMSA.
As well as the effects at the molecular level, incorporated NF-κB decoy ODN partially attenuated warm liver I/R injury at the physiological level. A similar previous study indicated the effectiveness of NF-κB decoy ODN against liver I/R injury during liver transplantation (Xu et al., 2005). That report showed that transfection of NF-κB decoy ODN into grafted livers attenuated injury through decreases in TNF-α and interferon (IFN)-γ levels. In their study, only the decreased NF-κB activation in the entire liver was shown, and the details regarding what types of cells were inhibited by NF-κB decoy ODN were not described. Our study indicated that NF-κB decoy ODN suppressed NF-κB activation in nonparenchymal cells, but not in hepatocytes. In addition, 3 hr after reperfusion, NF-κB decoy ODN significantly inhibited the production of cytokines such as TNF-α and IL-6, which are usually increased during the early phase of reperfusion. On the other hand, there were no apparent effects on GRO/CINC-1 six hours after reperfusion. This distinctly different effect of NF-κB decoy ODN on cytokines that are produced in the different time frames may explain why the amelioration of liver injury was limited in I/R injury despite the fact that NF-κB decoy ODN was so effective in suppressing liver injury caused by endotoxin or carbon tetrachloride. The types of cytokines involved during different phases of liver I/R injury and in different cell types may also be concerned in this context. TNF-α is produced mainly by Kupffer cells in the early phase of I/R injury (Suzuki and Toledo-Pereyra, 1994). In contrast, GRO/CINC-1 is produced not only from nonparenchymal cells but also from hepatocytes during the late phase (Farhood et al., 1995; Mawet et al., 1996). The effects of NF-κB decoy ODN administered by the RPVI method may be limited to the early phase of injury because the decoy was transfected into few hepatocytes.
A similar study was reported by Hoffmann and colleagues (2009). NF-κB decoy ODN transfected with gelatin nanoparticles was selectively taken up by Kupffer cells and showed an inhibitory effect on NF-κB activation 2 hr after reperfusion. However, in their study, NF-κB decoy ODN augmented reperfusion injury 24 hr after reperfusion, although they did not evaluate the degree of liver I/R injury in the earlier phase as we did. Taking the difference in observation points between our study and theirs into account, NF-κB inhibition in Kupffer cells by NF-κB decoy ODN may be limited to a short time frame. To overcome this limitation, it might be necessary to transfect NF-κB decoy ODN sequentially in order to suppress NF-κB activation for a longer period of time after liver I/R injury. Of course, a mechanism of warm liver I/R injury may be also involved in the limitation of amelioration. Warm liver I/R injury is composed of (1) warm ischemic injury due to energy shortage and (2) reperfusion injury caused by the production of several inflammatory cytokines. This mechanism is totally different from endotoxin-induced or carbon tetrachloride-induced liver injury, in which inflammatory cytokines are solely responsible for the damage. In the present study, although NF-κB decoy ODN significantly inhibited the production of cytokines during the reperfusion period, NF-κB decoy ODN had presumably no effects on warm ischemic injury.
Here, we showed that RPVI could be used to transfect NF-κB decoy ODN into Kupffer cells and sinusoidal endothelial cells to suppress NF-κB activation in these cells. This transfection markedly decreased serum inflammatory cytokine levels during the early phase of liver I/R injury.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.