
Abstract
Antisense oligonucleotide (AON)-mediated exon skipping to restore dystrophin expression in Duchenne muscular dystrophy (DMD) therapy shown promise in a number of human clinical trials. Current AON design methods are semi-empirical, involving either trial-and-error and/or preliminary experimentations. Therefore, a rational approach to design efficient AONs to address the wide spectrum of patients' mutations is desirable. Retrospective studies have extracted many AON design variables, but they were not tested prospectively to design AONs for skipping DMD exons. Not only did the variables differ among the various studies, no numerical cutoff for each variable was inferred, which makes their use in AON design difficult. The challenge is to thus select a minimal set of key independent variables that can consistently design efficient AONs. In this prospective study, a novel set of design variables with respective cutoff values was used to design 23 novel AONs, each to skip one of nine DMD exons. Nineteen AONs were found to be efficacious in inducing specific exon skipping (83% of total), of which 14 were considered efficient (61% of total), i.e., they induced exon skipping in >25% of total transcripts. Notably, the satisfactory success rates were achieved by using only three design variables; namely, co-transcriptional binding accessibility of target site, presence of exonic splicing enhancers, and target length. Retrospective analyses revealed that the most efficient AON in every exon targeted has the lowest average cumulative position (ACP) score. Taking the prospective and retrospective studies together, we propose that design guidelines recommend using the ACP score to select the most efficient AON for each exon.
Introduction
The prerequisite for rational AON design is a prior knowledge of the design variables and their effects on AON efficiency. Owing to the lack of such knowledge, prior approaches used in designing AONs for DMD exon skipping are semi-empirical (Aartsma-Rus et al., 2005, 2009b; Popplewell et al., 2009; Wilton et al., 2007), entailing a mix of trial-and-error and/or preliminary experimentation (Supplementary Table S1; Supplementary Data are available online at
In this prospective study, we proposed and experimentally validated a novel set of three design variables, namely co-transcriptional binding accessibility of target site, presence of exonic splicing enhancers (ESEs), and AON target length, to design efficient AONs targeting exon-internal sequences. Respective cutoff values were specified for the design variables that were subsequently used as criteria to select AON target sites. This combination of design variables and the associated cutoff criteria have not previously been applied in AON design, and the rationale for their selection is explained as follows.
The first two design variables are associated with the two respective key biological processes during AON-mediated exon skipping—(1) specific binding of an AON to an accessible target site, and (2) the binding thereafter prevents the spliceosome from identifying the target as an exon, and it will be removed along with the introns during splicing. As exon recognition and splicing is co-transcriptional (Maniatis and Reed, 2002; Tennyson et al., 1995), AONs need to compete with splicing factors for binding to the target exon during transcription. Throughout this process, however, the target exon secondary structure is dynamic as newly transcribed nucleotides create new combinations of complementary Watson-Crick base-pairings within the pre-mRNA. As a result, the target exon accessibility is dynamic and is accounted for by the first design variable, co-transcriptional binding accessibility of target site. To prompt target exon removal, AONs must bind to target sites possessing ESE motifs, as considered in the second design variable. These ESEs motifs are exon-internal sequences to which splicing factors bind for exon recognition. ESE-dependent exon recognition is particularly important in the DMD gene due to the presence of long introns (Graveley et al., 1999; Lam and Hertel, 2002). The third design variable, AON target length, is required for practical considerations because many short AON target sites (<10 bases) with high co-transcriptional binding accessibilities and ESE motifs are generated. Besides the issue of nonspecific binding to other sites, they may not anneal stably to the target due to low annealing temperatures. Generally, AON target length between 20 to 30 nt correlates with efficient exon skipping using AONs with either 2′-O-methyl phosphorothioate (2OMePS) or phosphorodiamidate morpholino (PMO) chemistries (Aartsma-Rus et al., 2010; Arechavala-Gomeza et al., 2007; Harding et al., 2007; Heemskerk et al., 2009; Popplewell et al., 2009, 2010). For 2OMePS AONs, Aartsma-Rus et al. (2009b) concluded that the optimal length is about 20 nt, while Harding et al. (2007) reported an optimal range of 25 to 30 nt. Whereas Popplewell et al. (2009, 2010) reported an optimal length of 30 nt for PMO AONs.
A total of 23 novel 2OMePS AONs, each targeting exon-internal sequences of one of nine exons, were designed based on the proposed set of design variables. Of these, 19 AONs (83% of total) were validated to be efficacious in inducing specific skipping of their targeted exons. Fourteen of the efficacious AONs (61% of total) were deemed efficient, inducing specific skipping of 25% or more transcripts; this standard classification criteria of AON efficiencies is used by prior studies (Aartsma-Rus et al., 2009b; Popplewell et al., 2009; Wilton et al., 2007). At least one efficient AON was obtained per targeted exon. The satisfactory success rates validated the selected set of design variables as major factors influencing AON efficiencies, and hence supported the plausibility of rational AON design. Retrospective analyses revealed that the most efficient AON in every exon targeted had the lowest average cumulative position (ACP) score. Taking the prospective and retrospective studies together, the proposed design guidelines will expedite the development of efficient AONs for therapy.
Materials and Methods
Computational prediction of the dynamical pre-mRNA secondary structure
The methodology for quantifying and analyzing the dynamics of the pre-mRNA structures in the progression of transcription did not depend on the choice of the prediction tool as long as co-transcriptional structures were obtainable. The mfold algorithm was eventually chosen because of its relative efficiency in computing long RNAs. It also has the advantage of being used in most published experimental results on AONs that target the DMD gene, and therefore results of this work can be compared with others on a common basis. Each window of analysis sequence was fed into mfold version 3.1 (Mitrpant et al., 2009; Zuker, 2003), which was executed on a Dell PowerEdge SC1420 server running Red Hat Enterprise Linux 4.0 OS (Fig. 1a). Since it was highly probable that the nascent pre-mRNA may not have the chance to assume optimal structures, we accepted suboptimal structures whose energies lie within 5% of the optimum.
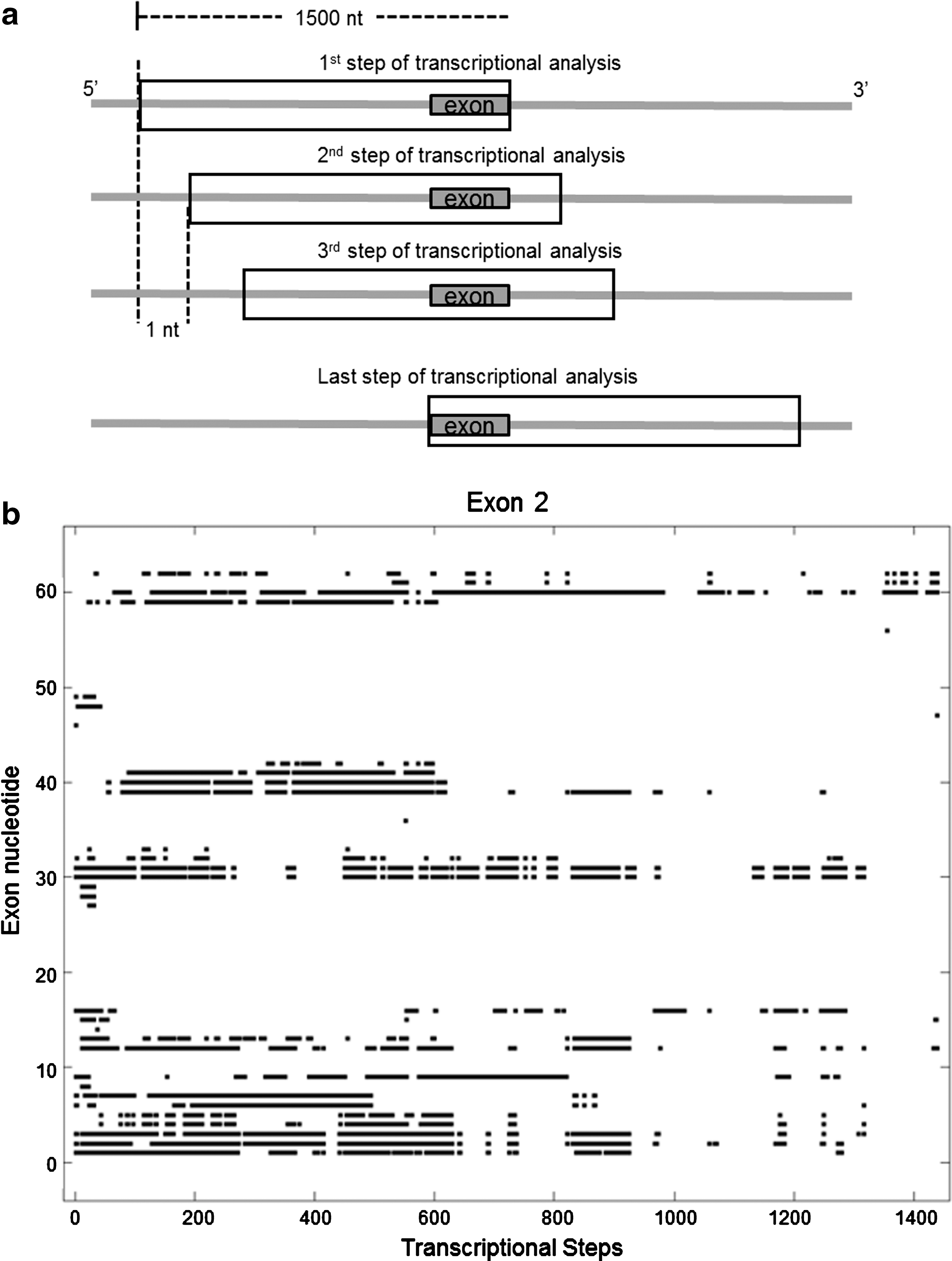
Co-transcriptional binding accessibility of target exon.
Using the predicted secondary structures at each step of transcription, a particular nucleotide of the exon is termed as an engaged nucleotide when all the structures indicate that it is paired to another complementary base. Figure 1b depicts the occurrences of all engaged nucleotides (shown as black spots) of exon 2. The L3 score of an AON target site is then computed from the engaged nucleotides plot as follows: the sum of total number of engaged nucleotides (black spots) divided by the product of the target length and total number of transcriptional steps of analysis.
Definition of ACP
The ACP score is the average of the sum of each target nucleotide position from exon 5′.
where Pi is relative position of the ith target nucleotide from exon 5′
L AON is AON target length
L Exon is target exon length
Cell cultures
Primary myoblast cells from DMD patients (kindly provided by Prof. Francesco Muntoni, UK) and primary human fibroblast cells from normal individual (Coriell, USA) were used. Fibroblast cells were used to test the AONs for their effect in restoring dystrophin mRNA reading frame when suitable patient myoblast cells are not available. To obtain a measurable amount from the low dystrophin mRNA in fibroblast cells, a higher amount of template was used in the nested PCR amplification of harvested dystrophin transcript.
Transfection into human fibroblast cells
AONs were synthesized by Sigma (Singapore) and modified with 2OMePS. Human fibroblast cells were cultured in six-well plates containing a growth medium of Dulbecco's modified Eagle's medium supplemented with 2 mmol/L
Transfection into primary cultures of DMD patients' myogenic cells
Two primary cell cultures derived from skeletal muscle of DMD patients carrying deletion of exons 45–50 and deletion of exons 48–52 were used to test the efficacy and efficiency of the AONs targeting exons 51 and 53, respectively. AONs were synthesized with identical chemistry as described above. The cells were seeded in six-well cell culture plates coated with collagen (Becton Dickinson, USA), and grown in supplemented muscle cell growth medium. Once confluent, the cultures were switched to supplemented muscle cell differentiation medium. Both media were purchased from Promocell (Germany). Transfection of AON was then performed when myoblasts fused to form visible myotubes (elongated cells containing multiple nuclei and myofibrils). Transfection was performed using protocols identical to those used in fibroblast cells except for the following. After 5-hr incubation, the transfection medium containing AON-Lipofectamine2000 was replaced with differentiation medium to maintain the cells. The cells were harvested 24 hr after transfection. Total RNA was isolated using Trizol (Invitrogen, Singapore) followed by first-strand cDNA synthesis using 1 μg of the total RNA, oligo random hexamer primers and the SMART™ cDNA Synthesis Kit (Clontech, USA) in a total reaction volume of 20 μl. First PCR was performed using 4 μl of cDNA in a total volume of 20 μl with 30 cycles. One microliter from the 20 μl of PCR products was then used for nested PCR of 32 cycles in a total 20-μl volume reaction. Exon skipping efficiency was also estimated by densitometry.
Immunocytochemistry
To show the restoration of dystrophin expression, immunohistochemistry staining was performed on the patients' muscle cells after treatment with AONs targeting exons 51 and 53. To analyze the effect of AON therapy, the expression of dystrophin was detected by immunochemistry staining of both AON-treated and untreated muscle cells. Primary antibody anti C-terminal dystrophin was used to assess the restoration of the dystrophin expression. Since DMD patients carry exon deletions that lead to an mRNA reading frame shift with a premature stop codon/truncation of the protein, the antibody against C-terminal (3′) of dystrophin should not be able to detect any dystrophin in their muscle cells. The success of the AON in restoring dystrophin expression should be marked by the detection of the dystrophin in the patients' muscle cells. The efficiency of the AON was reflected by the number of dystrophin-positive cells compared to dystrophin-negative myogenic cells.
On the third day after transfection, the muscle cells on the cover slips were fixed with methanol for 5 min followed by acetone treatment for 1 min. A double-staining immunocytochemistry method was used to detect dystrophin and desmin. Mouse monoclonal anti-dystrophin antibody (Novocastra Laboratories Ltd., UK) and rabbit polyclonal anti-desmin antibody (Sigma, Singapore) were used as the respective primary antibodies. Following the primary antibody, Alexa Fluor 594 goat anti-mouse lgG (Invitrogen, Singapore) and Alexa Fluor 488 goat anti-rabbit lgG (Invitrogen, Singapore) were applied to stain dystrophin and desmin, respectively. 6′-Diamino-2-phenylindole was used to visualize the nucleus.
Results
Rational AON design
Twenty-three novel AONs targeting exon-internal sequences of exons 2, 43, 45, 46, 47, 50, 51, 53, and 57 of the DMD gene were designed based on the three design variables—co-transcriptional binding accessibility of target site (L3 < 0.23), presence of ESEs (RESE > 0.44) and AON target length (20–35 nt). The target exons were chosen based on the therapeutic potential of their skipping on known DMD mutations (Aartsma-Rus et al., 2009a). Table 1 lists the AON target sequences and the numerical values of each of the respective design variables.
In the “Third” column, 1, 2, and 3 indicate that the first nucleotide of the AON target site is located at the first, second, and last third of the exon 5′, respectively.
ACP (last column) refers to average cumulative position that is defined in Materials and Methods .
AON efficiency obtained using patient myoblast cells.
The co-transcriptional binding accessibility of each target exon was determined as previously described (Wee et al., 2008) and is outlined in Fig. 1a (see Materials and Methods ). As an illustrative example, the co-transcriptional binding accessibility of exon 2 is depicted in Fig. 1b. A black spot on the figure indicates that the nucleotide of a target exon (as indicated by the vertical axis) is paired with a complementary nucleotide in all predictions of secondary structure at the transcriptional step (as indicated by the horizontal axis). Such a nucleotide is termed an engaged nucleotide at that transcriptional step. Thus, the black spots indicate the occurrences of potentially inaccessible nucleotides during transcription. The L3 score (Wee et al., 2008), which denotes the average number of engaged nucleotides over all the transcriptional steps manifested by the nucleotides of an AON target site, was used to identify target sites with high co-transcriptional binding accessibility; a low score indicates an AON target site is highly accessible for AON binding during transcription.
Next, target exon sequences were predicted for ESE motifs using ESE-finder (Cartegni et al., 2003) and Rescue-ESE (Fairbrother et al., 2002). The ESE prediction tools were chosen because they were widely used in prior AON studies (Aartsma-Rus et al., 2005, 2009b; Harding et al., 2007; Popplewell et al., 2009; Wilton et al., 2007), which facilitates the comparisons of our results with previously reported AON datasets. RESE score, which denotes the fraction of nucleotides within a target sequence with ESE motifs, was computed from merging the predicted motifs from the two sources with equal weight. The predicted ESE motifs were then merged with the co-transcriptional binding accessibility plot; see Fig. 2 for the case of exon 45 (see Supplementary Fig. S1 for the other target exons).
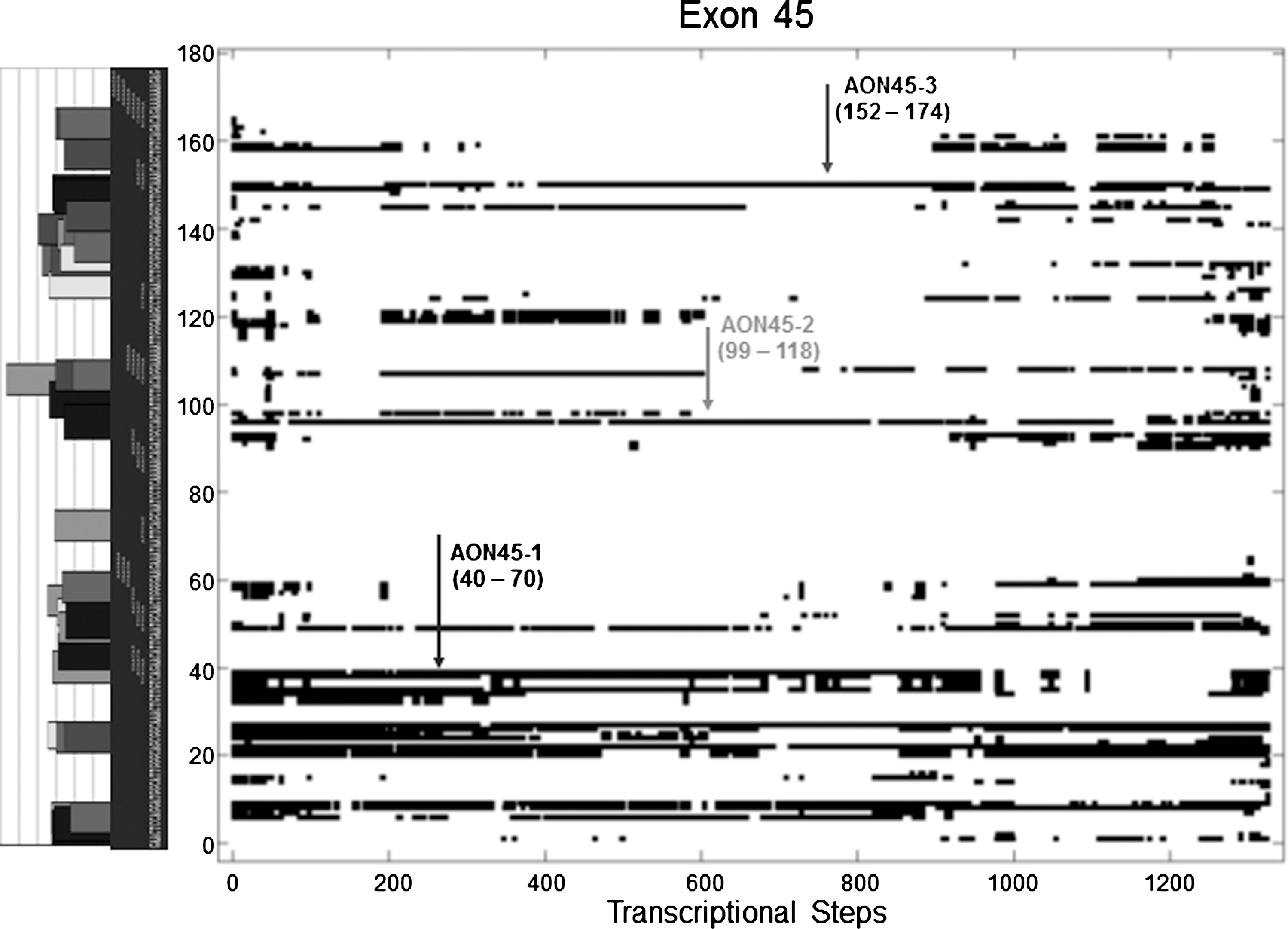
Rational identification of antisense oligonucleotide (AON) target sites for the case of exon 45. AON target sequences were determined by first identifying the sequences with the lowest possible L3 score, followed by selecting those among them with the highest possible RESE score. A low L3 score means that there are minimal engaged nucleotides (or black spots on the plot). Segments of the target exon sequence that are predicted to contain exonic splicing enhancer (ESE) motifs are indicated on the vertical axis—bar charts (ESE-finder) and short sequences (Rescue-ESE). The three AON target sites selected are indicated in the plot. See Supplementary Fig. S1 for the remaining target exons. RESE score denotes the fraction of nucleotides within a target sequence with ESE motifs.
Lastly, AON target sequences with lengths between 20 and 35 nucleotides long were subsequently identified first by having the lowest possible L3 score (<0.23) and then by the highest possible RESE score (>0.44). The range of AON target length, upper limit of L3 score, and lower limit of RESE score used were inferred from prior studies wherein efficient AON target sequences lie between this range (Aartsma-Rus et al., 2005, 2009b; Popplewell et al., 2009; Wilton et al., 2007).
Experimental validation of AON efficiency in inducing exon skipping
Twenty-one AONs were tested for their efficiency in exon skipping in primary cultures of human fibroblast cells, while five AONs each targeting exon 51 or 53 were tested in primary cultures of patient muscle cells (see Materials and Methods ). Their efficacy was determined by RT-PCR analysis, followed by sequencing analysis, to show the target exon skipping induced in DMD transcripts. Their efficiency was measured by densitometry of the gel images that semiquantified the amplicons with exon skipping and the amplicons of native transcripts. AON efficiency is presented as a percentage of the amplicons with exon skipping relative to total amplicons and are classified as ++ (>25%), + (<25%), and − (0%); these standard classification criteria of AON efficiencies were used in prior studies (Aartsma-Rus et al., 2005, 2009b; Popplewell et al., 2009; Wilton et al., 2007). An efficient or “++” AON is defined as one that can consistently induce transcription of more than 25% of total dystrophin mRNAs based upon the observation that 22% of human and 20% of mice native dystrophin transcripts are sufficient to either avoid or ameliorate DMD symptoms (Faulkner et al., 1997; Neri et al., 2007; Sharp et al., 2011). The agarose gel electrophoresis of nested PCR products from the cDNA derived from DMD patients' muscle cells, which were treated with AONs targeting exons 51 and 53, showed a band with a molecular weight between the native-transcript amplicons and the exon-skipping-carrying-transcript amplicons (Figs. 3c and 3d). Bidirectional sequencing confirmed that this band is a hetero-duplex formed between the native-transcript amplicons with the exon-skipping-carrying-transcript amplicons. In such hetero-duplex, the contributions from native transcripts and transcripts with exon skipping to the band were considered to be 50%/50%. Table 1 lists the average efficiency of each AON; the standard deviation of the average efficiency is given in Supplementary Table S2.
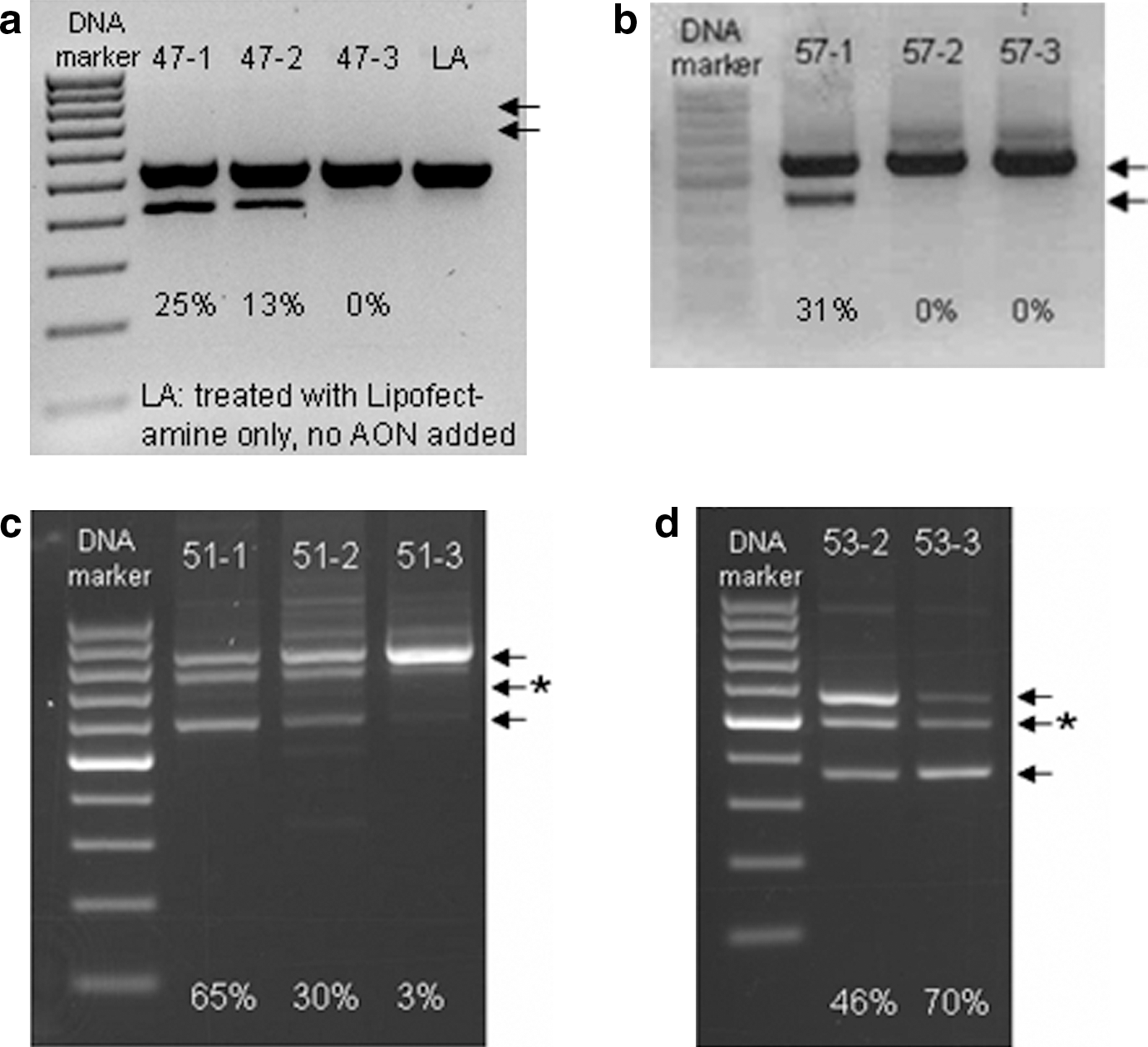
Efficiency of novel AONs targeting exons 47, 51, 53, and 57. AON at 100 nM concentration was used to treat cells to induce specific exon skipping of DMD transcripts. AON efficiency was assessed by comparing amplicons of DMD transcripts harboring exon skipping (lower arrow) with amplicons of native DMD transcript (upper arrow). AONs were able to induce exon skipping efficiently in human primary fibroblast cells for exons
Of the 23 validated AONs, 19 AONs were efficacious in inducing skipping of their specific target exon, among which 14 AONs induced specific skipping of 25% or more transcripts (efficiency score ++) and were considered efficient AONs. Notably, each target exon could be skipped by at least one efficient AON. Skipping of exons 47 and 57, which have been deemed unskippable by single target AONs (Aartsma-Rus and Van Ommen, 2007), was induced by at least one of our efficient AONs (Figs. 3a and 3b). Figures 3c and 3d show the RT-PCR products gel images demonstrating specific exon skipping induced by AONs targeting exons 51 and 53, respectively, in DMD patients' muscle cells. Immunocytochemical staining showing the restoration of dystrophin expression in DMD patients' muscle cells induced by AONs targeting exons 51 and 53, respectively, are shown in Fig. 4.
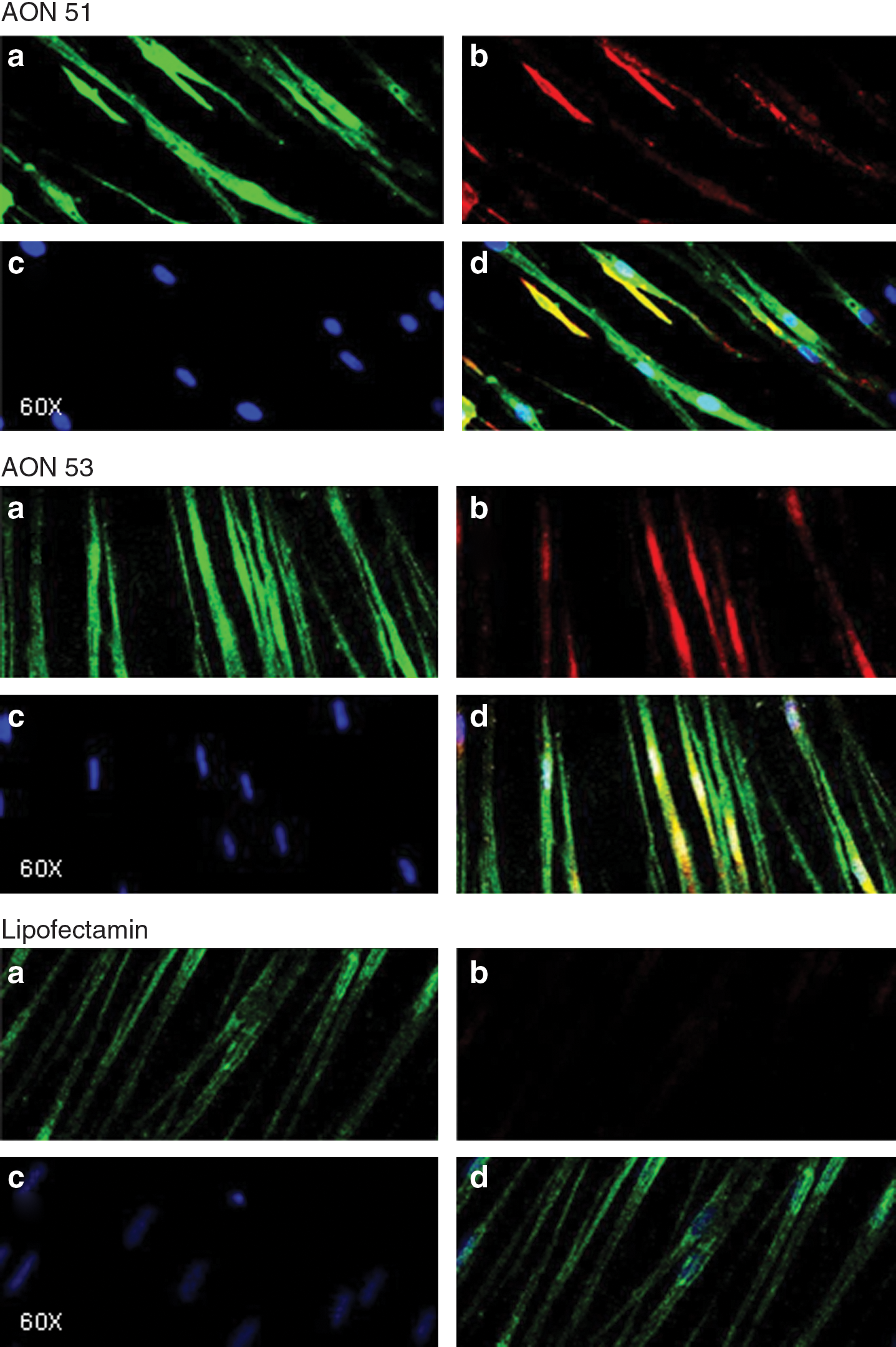
Immunocytochemistry of dystrophin protein in DMD patient skeletal muscle cells. AON induced restoration of dystrophin expression in DMD patient skeletal muscle cells. Immunocytochemistry were performed to stain desmin that mark
It is possible that the efficiency of AON57-1 reported here is underestimated. This is because the skipping of exon 57 generates an out-of-frame transcript that could be subjected to nonsense-mediated decay (NMD), possibly reducing the amount of skipped transcripts for amplification. Possible future work to offset the NMD effect is to transfect AON57-1 into DMD patient myoblasts. Although the reading frame of the intact transcript cannot be restored, the NMD effect of the skipped transcript can be offset by the NMD of the intact transcript as long as both transcripts undergo NMD at similar rates. However, because the locations and number of premature termination codons, which could influence the initiation and/or rate of NMD (McGlincy and Smith, 2008), differ in the intact and skipped transcripts, it is possible that they may not undergo NMD at similar rates.
Retrospective analysis
Retrospective analysis of the AON efficiencies obtained was performed to uncover additional design variables that may explain the basis for our inefficient and/or ineffective AONs. As expected, since all the AONs designed have low L3 scores, high RESE scores, and target length within the range, there is no statistically significant difference in L3 score, RESE, and AON length between efficient (++) and both inefficient and ineffective (+ and −) AONs (Supplementary Fig. S2). However, correlation between AON efficiency and the starting position of the AON target site was discerned. AONs whose first target nucleotide lies in the first third of the exon (from 5′ to 3′) are all efficient whereas for inefficient (+) and nonefficacious (−) AONs, the first target nucleotide lies in the middle and last third of the exon, respectively (Table 1). The ACP score (see Materials and Methods for its mathematical definition) is the average of the sum of each target nucleotide position from exon 5′ (Table 1). Statistical analysis revealed that efficient AONs have the lowest ACP scores (p=2E-5, see Fig. 5); i.e., they target sites nearest to the exon 5′ end. This finding is not due to lower L3 score or presence of more RESE motifs near the 5′ of the target exon (Supplementary Fig. S3).
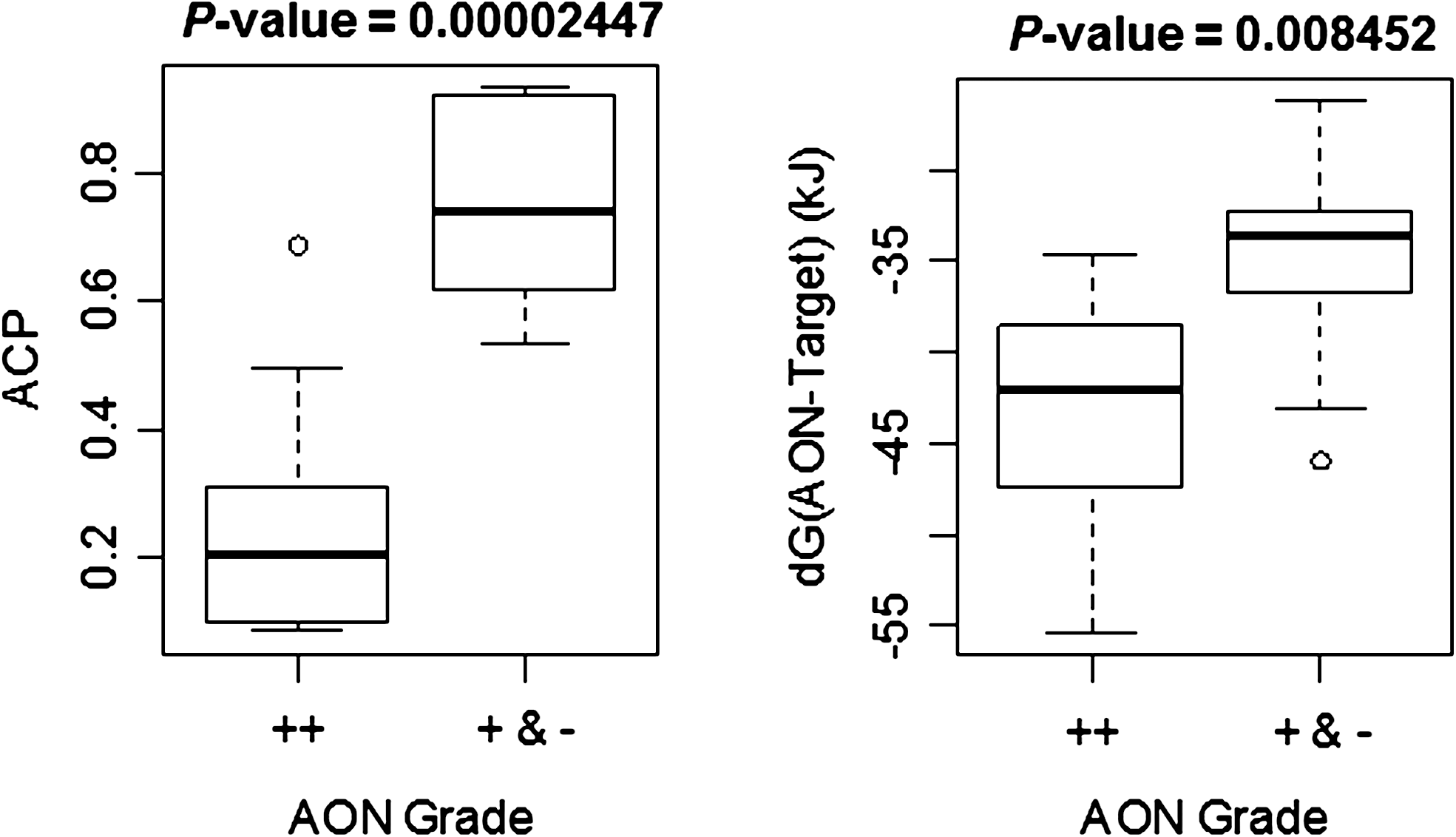
Boxplots of average cumulative position (ACP) score and AON-target Gibbs energy. ACP and AON-target Gibbs energy of 14 efficient (++) and 9 inefficient and nonefficacious (+ and −) AONs were compared. Kolmogorov-Smirnov (K-S) tests were performed for each score and the p-values are given on the respective boxplots.
Other variables were used to test for statistical significance between efficient and nonefficient AONs. With the exception of AON-target Gibbs energy (Fig. 5), no statistical significance was detected (Supplementary Fig. S4).
To determine possible variables that can be used to discern the most efficient AON per target exon, we sorted all AONs that target the same exon by their measured average efficiencies to examine whether their rankings correlate with any of the variables tested above. The AON with the lowest ACP score consistently correlated with the efficiency rankings in all exons targeted. That is, AON efficiency increases when ACP score decreases among AONs targeting the same exon (Supplementary Fig. S5). This suggests that the most efficient AON is one with the lowest ACP score.
Discussion
In this prospective study, a new combination of three design variables—co-transcriptional binding accessibility of target site, presence of ESE motifs, and AON length, were used to design 23 novel 2OMePS AONs each targeting one of nine exons of DMD pre-mRNA. Upon experimental validation, 14 AONs were found to be efficient among 19 efficacious AONs, with at least one efficient AON obtained per target exon. This translates to success rates of 83% and 61% for designing efficacious and efficient AONs, respectively. This study demonstrates the plausibility of attaining satisfactory success rates in designing AONs through an effective AON design guideline. Notably, this is achieved with only three design variables, which underscores the importance of selecting independent design variables appropriately and defining a specific numerical boundary for each variable. Retrospective analyses of the validated AON efficiencies revealed that the most efficient AON in every exon targeted had the lowest ACP score. This suggests that the most efficient AON could be selected from the pool of AONs by satisfying the design guideline of having the lowest ACP score. In contrast, reported design guidelines, which were inferred from retrospective analyses of published AONs (Aartsma-Rus et al., 2009b; Popplewell et al., 2009), have not been validated in a prospective study to design new AONs for exon skipping in the DMD gene. Furthermore, because these guidelines do not define numerical cutoffs of each design variable to limit the number of predicted AON target sites, some form of trial-and-error is inevitable in selecting AON candidates for validation.
Statistical analysis of the validated AONs efficiencies reveals that efficient AONs have the lowest ACP scores (p=2E-5) and the lowest AON-target Gibbs energy (p=0.008) (see Fig. 5). The ACP score quantifies the position of each target nucleotide from the target exon 5′, and our results show that AONs whose target sites had their first starting (5′) nucleotide in the first third of the exon (5′ to 3′) were efficient, whereas those that lay in the last third were all nonefficacious. In fact, several labs have reported the correlation of AON efficiency with position of target site from target exon 5′ end in their retrospective studies (Aartsma-Rus et al., 2005, 2009b; Popplewell et al., 2009; Wilton et al., 2007). The success rate for designing efficient AONs can be improved with ACP score as the fourth design variable; i.e., among candidate AON target sites with similar L3 score, AON length, and RESE, the one nearest to the exon 5′ should be selected. It is an independent variable since it is not correlated with the first three design variables (Supplementary Fig. S3). However, the effect of AON-target Gibbs energy is less clear because it is associated with both AON-target binding energy and GC content. In fact, the three variables have been reported to correlate with efficiency of mRNA down-regulation via AON-induced RNaseH degradation (Kinali et al., 2009). However, no statistical significance was detected for the latter two associated variables in our dataset, whereas only AON-target binding energy has been reported to be significant in two datasets (Aartsma-Rus et al., 2009b; Popplewell et al., 2009). Without future work to determine the relative importance and extent of interdependence among the variables, it is unclear whether the inclusion of AON-target Gibbs energy will lead to substantial improvement in the success rates. Furthermore, since the p-value of AON-target Gibbs energy is 345 times larger than the p-value of ACP score, ACP exerts more influence on AON efficiency.
Nevertheless, the approach used in this study is not without limitations. The effectiveness of the design guidelines depends on the accuracies of the computational tools used for predicting the pre-mRNA secondary structures and the presence of ESE motifs. Secondly, transcription, exon recognition, and exon splicing are complex regulated processes that involve myriad cellular components and not all mechanisms have been elucidated. Thus, the possibility of omitting a critical design variable cannot be ruled out. These limitations do not seem to be involved in this study as all the ineffective AONs can be explained by the fourth design variable.
To evaluate our AON design guidelines against reported AONs targeting exon-internal sequences, the following two assessments were performed: comparison of success rates and classification of AONs.
Comparison of success rates
Reported success rates for designing efficient (++) and efficacious (++ and +) AONs range from 18% to 38% and from 56% to 79%, respectively (Aartsma-Rus et al., 2005, 2009b; Harding et al., 2007; Popplewell et al., 2009; Wilton et al., 2007) (Supplementary Table S1). If only AONs targeting the same exons as in this study were considered, the reported range of success rates are lower; i.e., from 21% to 27% and from 44% to 73%, respectively (Supplementary Table S1). By comparison, with a success rate of 61%, our design guidelines confer a more substantial improvement in designing efficient AONs. Nevertheless, although the comparison of success rates is relatively less sensitive to the AON efficiencies than a direct comparison of AON efficiencies, differences in success rates detected between studies could be partially attributed to differences in AON efficiencies, which result from experimentation differences among the studies including, but not limited to transfection method, cell density, incubation time and PCR condition.
Classification of AONs
Our design guidelines were tested for the ability to correctly classify observed AON efficiencies in three reported datasets. Our design guidelines can classify the first dataset (Wilton et al., 2007) reasonably well, whereas only efficient and efficacious AONs in the second dataset (Popplewell et al., 2009) and only ineffective AONs in the third dataset (Aartsma-Rus et al., 2005) were classified well. The respective classification rates of efficient, efficacious, and ineffective AONs for the three dataset are (Supplementary Table S3): {97%, 96%, and 57%}, {80%, 88%, and 21%}, and {22%, 28%, and 68%}. On the other hand, Aartsma-Rus et al. (2009b) used their design guidelines encompassing four design variables inferred from retrospective analysis to classify efficacious and ineffective AONs from their dataset and those reported by Wilton et al. (2007). The respective classification rates for their and the Wilton et al. datasets were {94% and 48%} (Aartsma-Rus et al., 2009b) and {100%, 0%}(Aartsma-Rus et al., 2010). Aartsma-Rus et al. (2010) then went on to infer six-variables design guidelines from the Wilton et al. (2007) dataset and use it to reclassify the two datasets. The respective classification rates were {19% and 87%} and {92%, 63%}.
In conclusion, we have proposed and validated new design guidelines in this first prospective study to design AONs for efficient exon skipping in the DMD gene. The satisfactory success rates obtained through the application of the design guidelines support the use of a rational approach in designing AONs to determine efficient candidates for clinical trials. Since exon skipping has already been shown to be a viable therapy for DMD in clinical trials (Goemans et al., 2011; Kinali et al., 2009; van Deutekom et al., 2007), the determination of a set of validated design variables is timely given that different customized AONs are needed to address specific individual mutations in DMD patients (Aartsma-Rus et al., 2009a). Therefore, by eliminating trial-and-error and/or preliminary experimentations, our approach confers the advantage of reducing cost and resources while speeding up the AON design process. Besides DMD, a rational approach for designing efficient AONs could be applied to other genetic diseases which are amenable to AON therapy (Dominski and Kole, 1993; Gorman et al., 2000; Mercatante et al., 2001; Suwanmanee et al., 2002; Vetrini et al., 2006).
Footnotes
Acknowledgments
This article is dedicated to Dr. Woon Chee Yee, who passed away on August 31, 2011, for his intense involvement and contribution in this work. The authors wish to thank Professor Francesco Muntoni of the Dubowitz Neuromuscular Centre, UCL Institute of Child Health, London, UK, for kindly providing samples of DMD patient skeletal muscle cells used in this study. This study was funded by National Medical Research Council Singapore (grant no. NRN05/007).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.