
Abstract
Newcastle disease virus (NDV) is a naturally oncolytic virus that has been shown to be safe and effective for cancer therapy. Tumor virotherapy using NDV emerged in the 1950s and has advanced more recently by the increased availability of reverse genetics technology. In this study, we constructed a reverse genetics system based on the virulent and oncolytic NDV Italien strain, and generated two recombinant NDVs carrying a gene encoding either enhanced green fluorescent protein or firefly luciferase. We evaluated the replication and antitumor characteristics of these viruses in vitro and in vivo. Our data showed that the insertion of exogenous reporter genes did not affect NDV replication and sensitivity to type I interferon. The recombinant NDVs kept the property of tumor-selective replication both in vitro and in vivo and strongly induced syncytium formation leading to cell death. Moreover, the recombinant NDVs significantly prolonged the survival of tumor-bearing athymic mice (p=0.017) and suppressed the loss of body weight after intratumoral injection. Taken together, our study provides a novel platform to develop recombinant oncolytic viruses based on the NDV Italien strain and shows the efficiency of recombinant NDV Italien for oncolytic virotherapy of tumors.
Introduction
Newcastle disease virus (NDV), a member of the paramyxoviruses, is a negative-sense single-stranded RNA virus. NDV is also known as a naturally oncolytic virus that has been used in the clinic since the 1950s. Nowadays, three strains of NDV, that is, MTH-68/H, NDV-HUJ, and PV701, are being employed in phase I/II clinical trials for oncolytic tumor therapy (Csatary et al., 2004; Freeman et al., 2006; Hotte et al., 2007; Lorence et al., 2007). All data demonstrate that NDV is a safe agent for oncolytic virotherapy in human body, with the most severe side effects being mild flulike symptoms and conjunctivitis (Pecora et al., 2002; Laurie et al., 2006). However, the maximization of NDV therapeutic efficacy is still a challenge.
NDV strains are categorized as velogenic, mesogenic, or lentogenic, based on the pathogenicity for avian species. The virulence of NDV is primarily determined by the F0 cleavage site, which is involved in the maturation of the fusion (F) protein (Peeters et al., 1999). Usually, the amino acid sequence of the cleavage site is 112R/K-R-Q-R/K-R-F117 for velogenic and 112G/E-K/R-Q-G/E-R-L117 for lentogenic strains (Collins et al., 1993). The F0 cleavage site of velogenic strains can be cleaved by various host proteases in contrast to that of lentogenic strains, which are cleaved only by trypsin-like proteases. As a result, velogenic strains have a higher in vivo replication rate and an extended tissue distribution compared with lentogenic and mesogenic strains (de Leeuw et al., 2003). This property of velogenic strains, we presume, will be a benefit to its antitumor activity in cancer therapy. It has been shown that a recombinant NDV modified to contain a highly fusogenic F protein with a multibasic cleavage site enhances its oncolytic properties (Vigil et al., 2007; Altomonte et al., 2010). In previous experiments we cloned the complete genomic cDNA of NDV strain Italien and obtained the sequence of the F0 cleavage site, 112R-R-Q-R-R-F117, which indicates that NDV Italien should be categorized as a velogenic strain (GenBank accession number: EU293914) (Wei et al., 2008). The oncolytic potential of NDV Italien was previously evaluated by retargeting virions to tumors in the presence of a bispecific protein to enhance the tumor specificity and reduce the side effects, because intravenous injection of a high dose of NDV Italien caused acute cytopathic side effects including small-sized pale spots on the liver, enlarged spleen, and alimentary tract hemorrhage 24 hr postinjection (Bian et al., 2006). However, there remains a clear need to produce recombinant NDV Italien with improved oncolytic properties.
In this study, we developed a reverse genetics system based on the NDV Italien strain. Using this system, we generated two recombinant NDV strains expressing either firefly luciferase or the enhanced green fluorescent protein (EGFP). We investigated the characteristics of the recombinant NDVs, including replication rate, sensitivity to type I interferon, and antitumor activity in vitro and in vivo. Our results describe the first recombinant NDV based on the Italien strain and provide a platform and rationale for further work to develop agents for cancer virotherapy.
Materials and Methods
Cell lines and virus
The BSR-T7/5 cell line was kindly donated by K.-K. Conzelmann (Ludwig Maximilian University, Munich, Germany). HeLa, HuH-7, HepG2, SMMC-7721, BHK-21, and MRC-5 cells were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). All cells were grown in appropriate media and were maintained at 37°C in a humidified incubator supplied with 5% CO2. The NDV Italien strain was prepared as reported previously (Wei et al., 2008).
Infection
Virus was diluted with serum-free medium appropriately based on multiplicity of infection (MOI) and cell number. Cells were washed twice with serum-free media and then incubated with virus for 1 hr in 37°C incubator. After infection, cells were washed and placed in culture medium.
Reverse genetic system
The genomic RNA of NDV Italien was isolated with a MiniBEST viral RNA/DNA extraction kit (TaKaRa Bio, Otsu, Japan). cDNA fragments covering the complete genome and the NP, P, and L genes were amplified, using a PrimeScript RT reagents kit (TaKaRa Bio) and PrimeSTAR HS high fidelity DNA polymerase (TaKaRa Bio). All primers for reverse transcription and PCR amplification are listed in Supplementary Tables S1 and S2 (supplementary data are available online at
To clone the nucleocapsid protein (NP) gene, primers NP-F and NP-R were used for high-fidelity RT-PCR. The PCR product was purified with a gel extraction kit (TaKaRa Bio). The phosphoprotein (P) and large polymerase protein (L) genes were cloned with primers P-F/R and L-F/R, respectively.
To construct the three helper plasmids T7-NP, T7-P, and T7-L, the internal ribosome entry site (IRES) sequence was recovered from plasmid pIRES2-EGFP (Clontech/TaKaRa Bio, Mountain View, CA) with primers IRES-F and IRES-R in which the T7 promoter was added at the 5′ terminal of the fragment. The T7-IRES fragment was fused to the NP, P, and L genes, respectively, using overlap PCR. Finally, the PCR products were cloned into pMD-19T T-vector (TaKaRa Bio) and the resulting plasmids were named T7-NP, T7-P, and T7-L, respectively. The correct function of the helper plasmids was determined with a minigenome system as described previously (Feng et al., 2011).
To construct the full-length genomic cDNA plasmid, an oligonucleotide (named Adaptor) containing multiple restriction sites was synthesized (GenScript, Piscataway, NJ) and introduced into plasmid pBR322 (named pBR322-M). The viral genome was divided into 10 subgenomic cDNA fragments and assembled into pBR322-M, using unique restriction sites. To facilitate the exogenous gene insertion, 36 extra nucleotides including an NDV consensus gene-start (GS) and gene-end (GE) transcription unit and an AgeI restriction site were introduced into the noncoding region between the hemagglutinin–neuraminidase (HN) and F genes. The size of the recombinant genome obeyed the “rule of six.” The final plasmid was named pBR-rNDV. The genome plasmid was verified by sequencing (Beijing Genomics Institute, Beijing, China).
Recombinant NDV
To construct recombinant NDV carrying exogenous genes, the firefly luciferase and EGFP genes were cloned from plasmid pGL3-Basic (Promega, Madison, WI) and pIRES2-EGFP with primer pairs Luc-F/R and EGFP-F/R, respectively. The fragments were provided with a translation initiation (Kozak) sequence and flanked on either side with an AgeI restriction site. The PCR products were purified with a gel extraction kit and cloned into pBR-rNDV, using the AgeI site. The resulting plasmids were named pBR-rNDV-Green and pBR-rNDV-Luci.
To generate recombinant NDV, BSR-T7/5 cells were cultured overnight to 80% confluence in a six-well plate and transfected with 2.5 μg of pBR-rNDV (or pBR-rNDV-Green and pBR-rNDV-Luci), 1 μg of T7-NP, 1 μg of T7-P, and 0.5 μg of T7-L, using Lipofectamine 2000 (Life Technologies, Carlsbad, CA). In the control, pBR-rNDV was replaced with pBR322.
To identify recombinant NDV, virus was collected from the cotransfected BSR-T7/5 cells and isolated by plaque purification. Primers VT-F (5′-GACGGGATAACTCTGAGG-3′) and VT-R (5′-GTATGCCAACAAGGTCGC-3′) were used to amplify the noncoding region between the HN and F genes. The PCR product was cloned into pMD-18T vector (TaKaRa Bio) and verified by sequencing.
Immunofluorescence
HeLa cells were cultured in a 24-well plate to 80% confluence and infected with rNDV-Green or NDV Italien at an MOI of 0.01. At 36 hr postinfection, cells were fixed with 4% paraformaldehyde and stained with rabbit anti-NDV polyclonal antibody and cyanine 3 (Cy3)-labeled goat anti-rabbit IgG (Zymed/Life Technologies). Samples were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Life Technologies) and visualized with a BX60 fluorescence microscope (Olympus, Tokyo, Japan).
Luminescence
HeLa cells were infected with rNDV-Luci (MOI, 0.01). The supernatant and cell lysate were collected 6, 12, 24, 36, 48 and 72 hr postinfection to determine the luminescence intensity, using a Bright-Glo luciferase assay system (Promega) with GENios Plus (Tecan Group, Maennedorf, Switzerland).
Viral titer and MTT assay
Viral titer was measured by end-point dilution assay (50% tissue culture infective dose [TCID50]/ml), using BHK-21 cells with eight replicates for each dilution. The TCID50 per milliliter was calculated according to the method of Reed and Muench (Reed and Muench, 1938). Each assay was repeated three times.
The viability of cells was assessed with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) cell proliferation and cytotoxicity assay kit (Beyotime Institute of Biotechnology, Haimen, China) with six replicates for each sample. Absorbance was determined with ELISA microplate readers (BioTek Instruments, Winooski, VT).
Live cell imaging
HeLa cells were plated on a 35-mm glass bottom dish and infected with rNDV-Green (MOI, 0.1). Cells were cultured with an INU incubation system (Tokai Hit, Shizuoka, Japan) and observed under an A1 confocal laser microscope system (Nikon Instruments, Melville, NY). Live cell imaging was recorded every 15 min and analyzed with NIS-Elements microscope imaging software (Nikon Instruments).
Syncytial cell death
To identify the fate of syncytia, HeLa cells were infected with rNDV-Luci (MOI, 0.01) for 24 hr. Cells were stained with a DeadEnd fluorometric TUNEL (terminal deoxynucleotidyltransferase dUTP nick end labeling) system (Promega) and PI solution and visualized under a BX60 fluorescence microscope (Olympus).
Transmission electron microscopy
HeLa cells were infected with rNDV-Luci (MOI, 0.01) for 24 hr. Cells were collected at 300 g, fixed with 2.5% glutaraldehyde in sodium cacodylate (0.1 mol/liter) and CaCl2 (1 mmol/liter) (pH 7.4), and postfixed in 1% osmium tetroxide. The samples were then dehydrated in graded ethanol, from 50 to 100%, and embedded in Spurr's resin. Ultrathin sections (90 nm) were cut and placed on 150 mesh copper grids. After staining with lead citrate and uranyl acetate (2% in 50% ethanol), the samples were examined with a transmission electron microscope (JEM-2000EX; JEOL, Tokyo, Japan).
Animal study
To establish a xenograft tumor model, 5×106 HeLa cells were injected subcutaneously into the middle flank of 6-week-old athymic mice (Fourth Military Medical University, Xi'an, China). Therapy started when the short diameter of the tumor reached 5 mm. Mice were randomly divided into three groups, that is, an intravenous injection group (n=10), an intratumoral injection group (n=9) and a control group (n=9). The animals in the intravenous group were injected via the caudal vein with 5×108 plaque-forming units (PFU) of rNDV-Luci suspended in 100 μl of phosphate-buffered saline (PBS). Virus was administered to the animals of the intratumoral group, by multipoint intratumoral injection of 5×108 PFU of rNDV-Luci suspended in 10 μl of PBS. The animals in the control group were injected with an equal volume of PBS both intravenously and intratumorally. For all groups the treatment was repeated on days 3 and 5 after the first injection.
Tumor diameter was measured by image measurement and quantified with ImageJ software (National Institutes of Health, Bethesda, MD). Tumor volume was calculated according to the following the formula: volume=(width)2×length/2. Both tumor volume and body weight were measured every 3 days.
Live imaging of athymic mice
Xenograft tumor model mice were randomly divided into an intravenous group (n=3), intratumoral group (n=3), and control group (n=3), and treated with rNDV-Luci (5×108 PFU) on days 1, 4 and 7. On the day after each injection, mice were anesthetized with isoflurane mixed with oxygen and then injected intraperitoneally with
Statistical analysis
Statistical data were obtained with SigmaPlot (Systat Software, San Jose, CA). A two-tailed Student t test was applied for comparison of two individual data. One-way analysis of variance (ANOVA) was used to determine the significance of differences among the various groups. Survival analysis was determined by Kaplan–Meier estimation and log-rank test. A p value less than 0.05 was considered statistically significant. Data are presented as means plus the standard deviation (SD).
Results
Construction of a reverse genetics system for NDV Italien
The reverse genetics rescue system consisted of helper plasmids (Fig. 1a), a minigenome plasmid, and a genome plasmid containing the complete antigenomic cDNA (Fig. 1b). All plasmids were constructed as described in Materials and Methods and verified by DNA sequencing. To verify the correct function of helper plasmids, T7-NP, T7-P, and T7-L were cotransfected with the minigenome MG-L (Feng et al., 2011) into BSR-T7/5 cells. Results showed that the luminescence of samples of cotransfected cells was significantly greater than that of the negative controls (p<0.001) (Fig. 1c), indicating that the helper plasmids could express the viral NP, P and L proteins, facilitating assembly of ribonucleoprotein complexes in BSR-T7/5 cells.
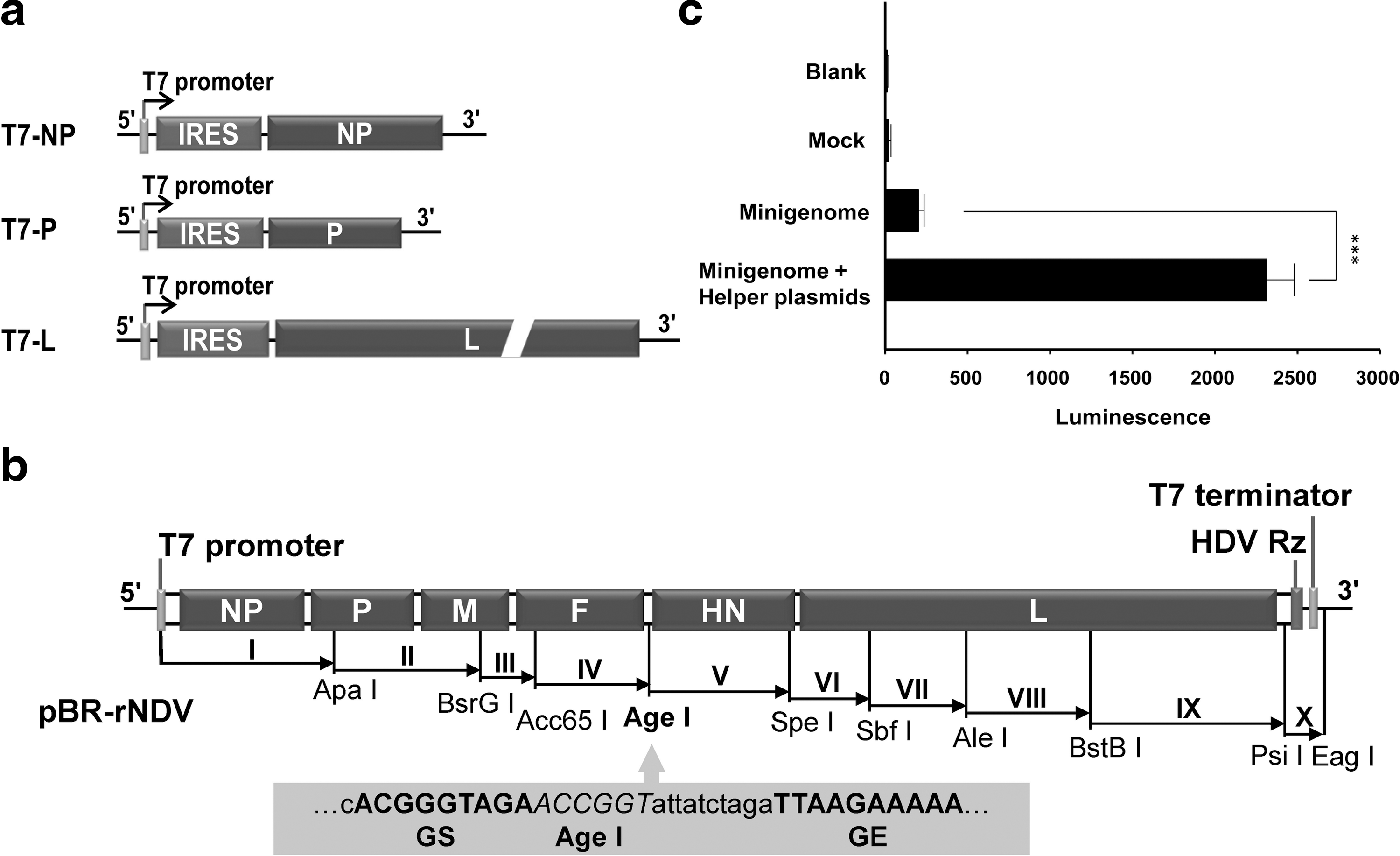
Construction of reverse genetics system.
To rescue recombinant NDV Italien, pBR-rNDV and the three helper plasmids were cotransfected into BSR-T7/5 cells. On day 4 posttransfection, several plaques were observed in the cotransfected cells, indicating that recombinant virus was produced successfully. Sequence analysis of the noncoding region between the HN and F genes of the rescued virus confirmed the presence of the exogenous nucleotides as we designed. These results demonstrate that recombinant NDV Italien was generated successfully using the reverse genetics system.
Generation of recombinant NDV carrying reporter genes
Plasmids pBR-rNDV-Green (carrying EGFP) and pBR-rNDV-Luci (carrying firefly luciferase) (Fig. 2a) were constructed as described in Materials and Methods and verified by DNA sequencing. To generate recombinant virus, pBR-rNDV-Green and pBR-rNDV-Luci were cotransfected with the three helper plasmids into BSR-T7/5 cells. The rescued viruses, rNDV-Green and rNDV-Luci, were collected and purified. To examine expression of the exogenous gene, HeLa cells were infected with rNDV-Green. At 36 hr postinfection, EGFP expression was detected in the rNDV-Green-infected cells and the recombinant NDV induced the extensive formation of syncytia (Fig. 2b). The supernatant and cell lysate of HeLa cells infected with rNDV-Luci were collected and subjected to detection of luciferase activity. Results showed that the luminescence in cell lysate and supernatant of rNDV-Luci-infected cells was significantly higher than that of NDV Italien-infected control cells (Fig. 2c). Although luciferase is not a secreted protein and no signal peptide was introduced in the cloned gene, we presume that luciferase was released into the supernatant as a result of cell lysis induced by virus replication.
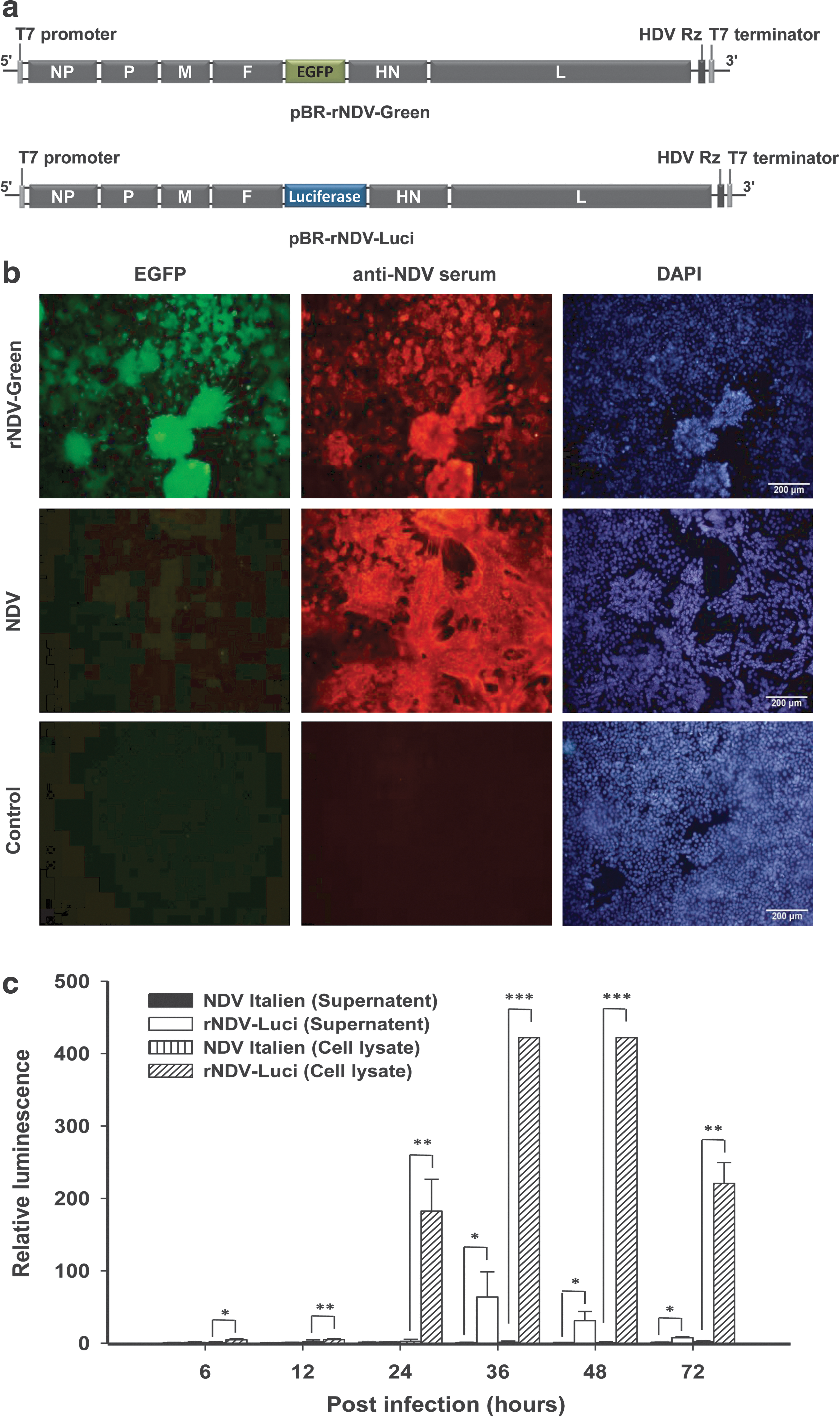
Construction and identification of recombinant NDV.
Replication characteristics of recombinant NDV
To evaluate the growth characteristics of the recombinant NDV strains, viral titers of the supernatant of infected BHK-21 cells were determined by TCID50. As show in Fig. 3, the insertion of an exogenous gene, either EGFP or luciferase, into the NDV genome did not significantly influence viral replication during 72 hr postinfection when compared with the parental NDV Italien strain.
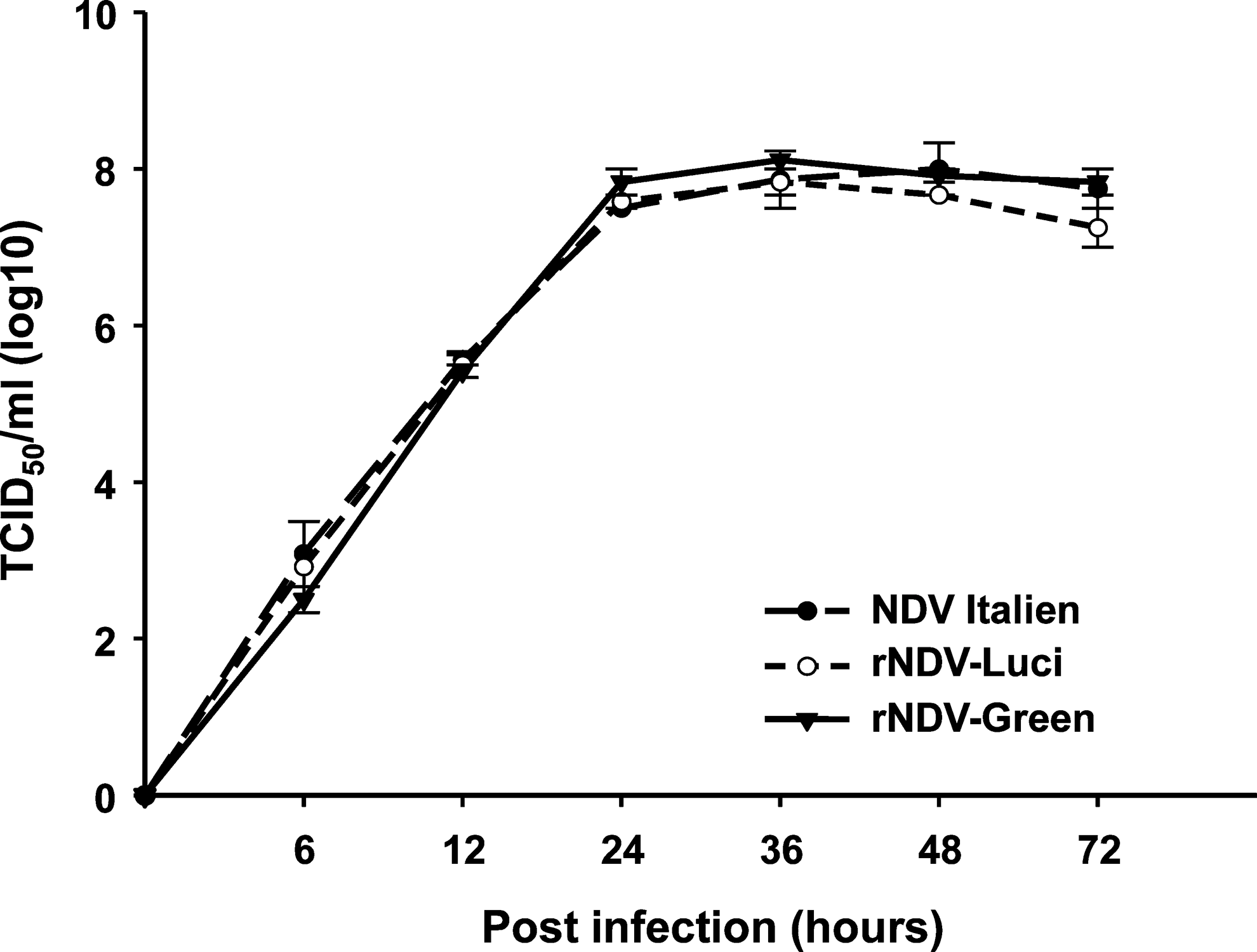
Proliferation characteristics of recombinant NDV. BHK-21 cells were infected with recombinant NDV or NDV Italien at an MOI of 0.01. The culture supernatant was collected 6, 12, 24, 36, 48, and 72 hr postinfection and the viral titer (TCID50/ml) was determined.
Sensitivity of recombinant NDV to type I interferon
It has been reported that tumor-selective replication of NDV is due to defects in interferon (IFN)-mediated antiviral responses. Therefore, the safety of NDV in humans depends on the sensitivity of NDV to type I IFN. We wondered whether, similar to its parental strain, the recombinant NDV strains retained the sensitivity to IFN. To this end, cells were pretreated with human IFN-α and IFN-β followed by infection with rNDV-Luci or NDV Italien. We first demonstrated that the cell viability of NDV-infected cells was inversely related to the viral titer. Therefore, the MTT assay of cell viability could be used to determine viral replication (Supplementary Fig. S1a and b). Infected HepG2 cells showed no differences in cell viability after infection with recombinant NDV or NDV Italien regardless of IFN-α or IFN-β pretreatment, and cell viability was dose dependent with IFN-α and IFN-β, indicating that the exogenous gene did not interfere with the sensitivity of NDV to type I IFN. Thus, the recombinant NDV strain maintained the safety for humans of the parental NDV strain (Fig. 4a).
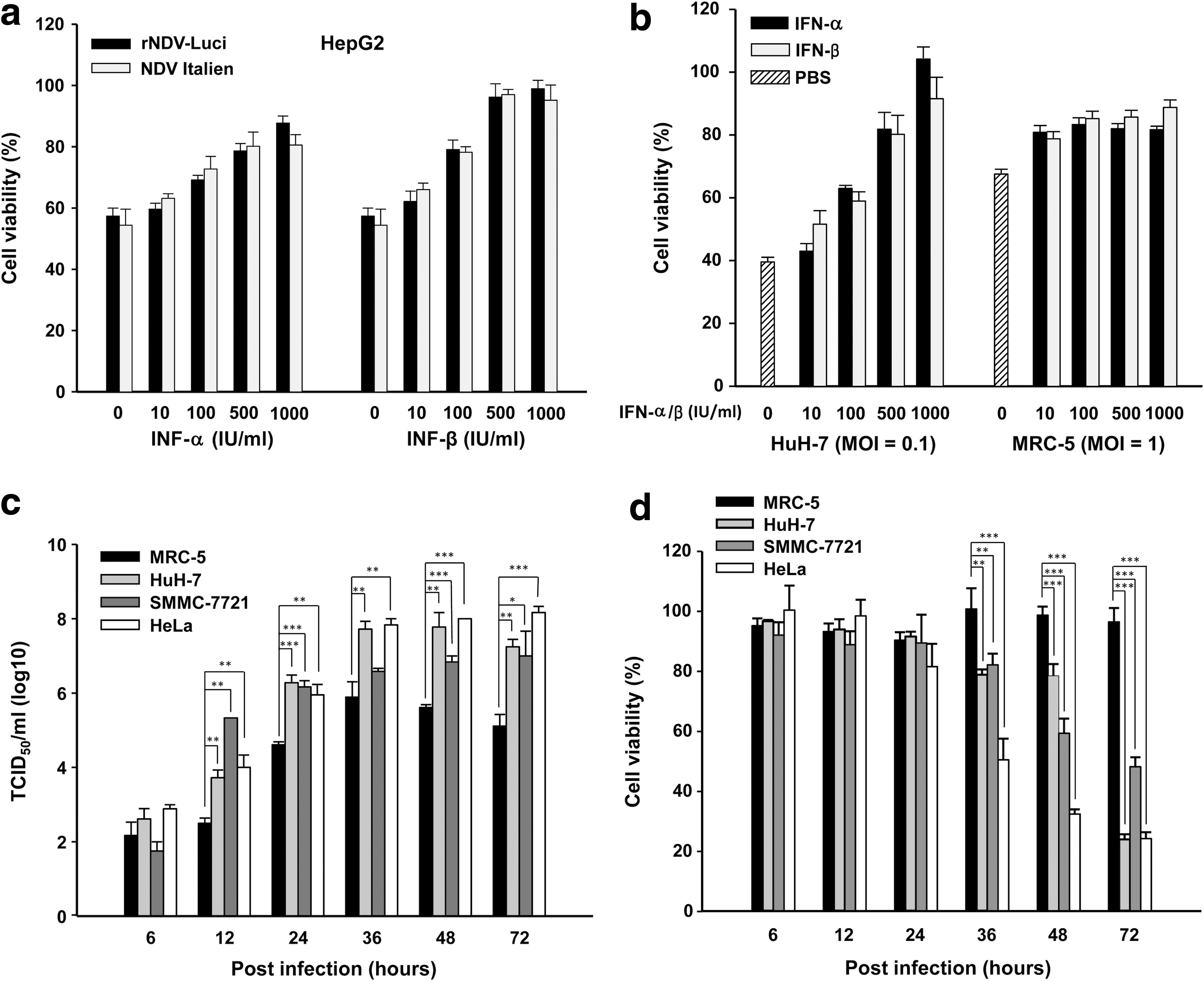
Characteristics of recombinant NDV.
We compared the various responses to IFN-α/β pretreatment of normal and tumor cell lines that had been infected with recombinant rNDV-Luci. As shown in Fig. 4b, the viability of HuH-7 tumor cells after infection with virus (MOI, 0.1) was dependent on IFN-α/β and increased with higher IFN doses. In contrast, after infection of normal MRC-5 cells (MOI, 1) the maximal responses to IFN were already achieved at a low dose, suggesting that viral replication in healthy cells could be inhibited by type I IFN at early stages of infection.
Selective replication of recombinant NDV in tumor cells
One normal cell line (MRC-5) and three tumor cell lines (HeLa, SMMC-7721, and HuH-7) were infected with rNDV-Luci at an MOI of 0.01, and the viral titer and cell viability were assayed at various time points after infection. At early time points after infection (6 hr), neither viral titers nor cell viability differed between the normal and tumor cell lines. However, on extension of the incubation time, viral titers in all tumor cell lines were significantly higher than those in MRC-5 cells, showing an elevation exceeding 200-fold at 72 hr postinfection (Fig. 4c). Meanwhile, the cell viability of MRC-5 cells remained greater than 95% until 72 hr postinfection, whereas the survival rates of the three tumor cell lines decreased significantly by viral infection from 36 hr postinfection onward (Fig. 4d). These results indicate that the recombinant virus still retains the intrinsic tumor-selective and oncolytic properties of NDV.
Induction of syncytial cell death by recombinant NDV
The formation of syncytia by infected cells is one of the characteristics of paramyxoviruses. To examine the kinetics of syncytium formation by an NDV recombinant that expresses an exogenous gene, HeLa cells were infected with rNDV-Green (MOI, 0.1) and directly observed under a time-lapse confocal microscope. As shown in Fig. 5a and Supplementary Video S1, rNDV-Green induced cell fusion and expressed EGFP as early as 7 hr postinfection, and expression was maximal at 14 hr postinfection. Thereafter the syncytia began to shrink, finally leading to detachment of the fused cells. Concurrently, the intensity of EGFP in syncytia decreased from 14 hr postinfection onward. More than 100 nuclei were observed to aggregate in the cytoplasm of fused cells, appearing as typical rosette-like structures at the periphery. Viral replication contributed to this structure formation as shown by EGFP reporter gene expression (Fig. 5b). To examine the fate of syncytia, HeLa cells infected with rNDV-Luci were examined with a TUNEL kit and propidium iodide (PI) staining at 24 hr postinfection. As can be seen in Fig. 5c and d, syncytial cells were strongly positive by TUNEL assay and PI staining demonstrating that NDV-fused cells suffered severe DNA damage, finally followed by cell death. When examined by electron microscopy the dead syncytia presented as compact chromatin and electron-dense structures underlying the nuclear envelope, which is distinct from typical apoptosis features (Fig. 5e).
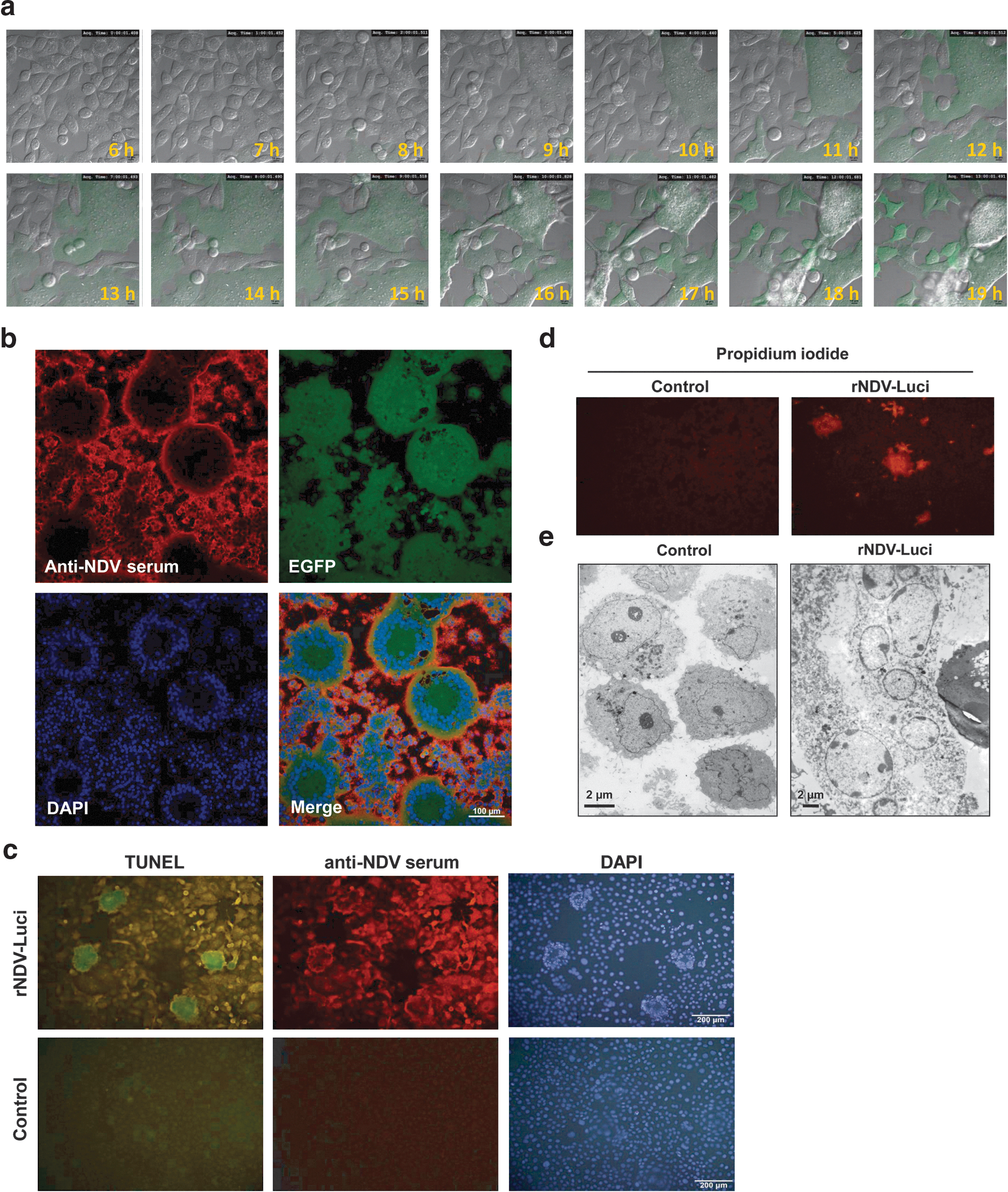
Recombinant NDV induced syncytium formation and cell death.
Tumor-targeted gene expression by recombinant NDV in vivo
We proceeded to determine whether the recombinant NDV maintained tumor-selective replication in vivo. Subcutaneous tumor-bearing athymic mice were infected with rNDV-Luci by intravenous or intratumoral administration. Luciferase gene expression was visualized by luminescence, using an in vivo image system (IVIS). As show in Fig. 6, in intratumorally inoculated animals luminescence was detected only in tumor, not in other tissues, on day 2 postinfection and could be detected until day 8 postinfection. In intravenously inoculated animals, the luminescence was exclusively detected in tumor on day 5 postinfection, and the intensity of the signal had increased by day 8 postinfection. These results indicate that the recombinant NDV replicates selectively in tumor tissue in vivo, leading to exogenous gene expression that is limited to tumor tissue either after locoregional or systemic administration.

Recombinant NDV replicates selectively in tumor tissue of athymic mice. Mice containing subcutaneous tumors derived from HeLa cells were injected with rNDV-Luci (5×108 PFU) intratumorally or intravenously on days 1, 4, and 7. On the next day after each treatment (days 2, 5, and 8), in vivo images were captured with the IVIS imaging system. The exposure time was set to 3 min for intravenously injected animals and 30 sec for intratumorally injected animals.
Prolongation of survival of tumor-bearing mice by recombinant NDV
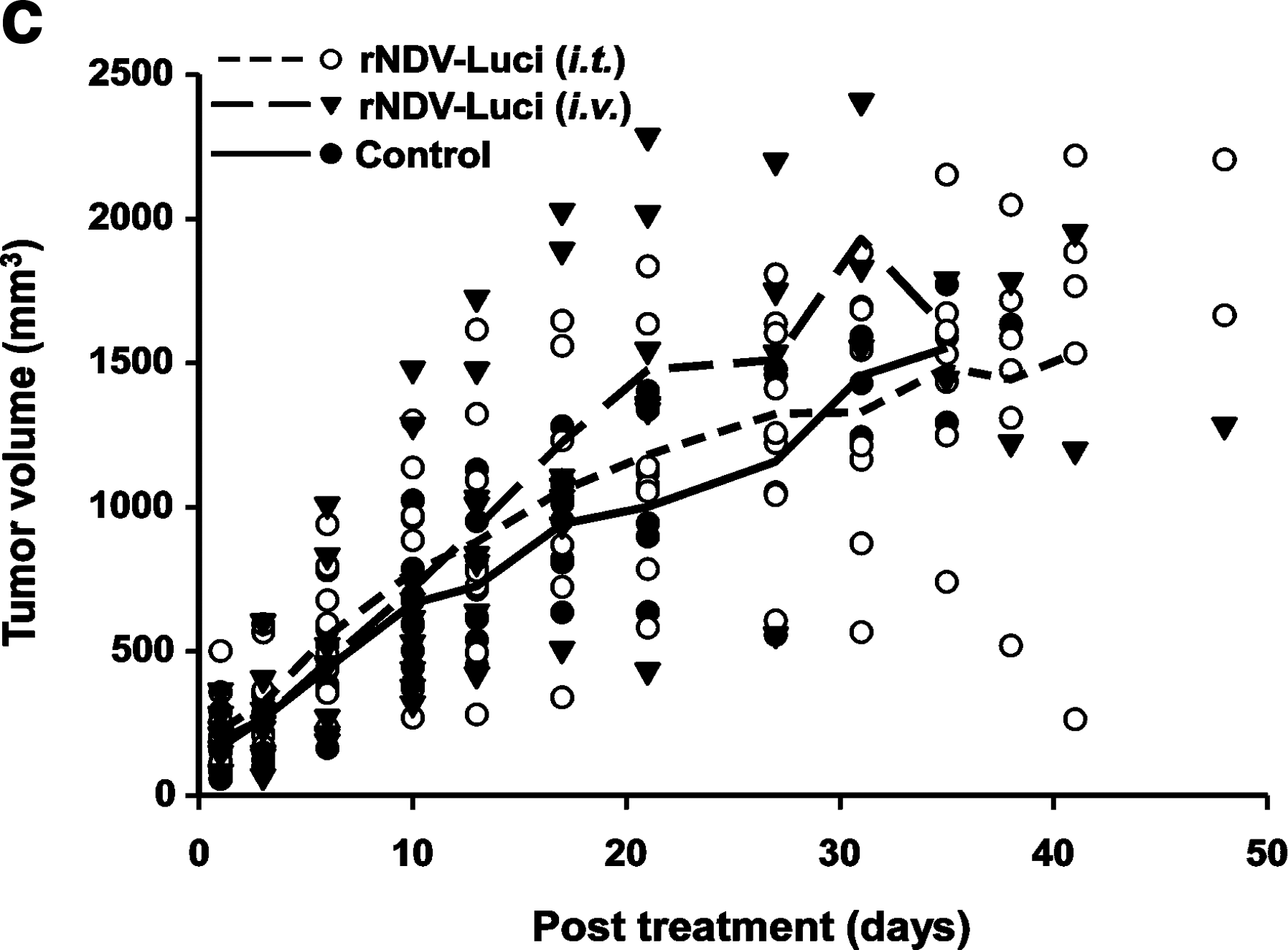
To assess whether the in vitro cytolytic effect of NDV would translate into effective antitumor activity in vivo, tumor-bearing athymic mice were randomly divided into three groups followed by treatment with rNDV-Luci by intravenous or intratumoral administration. One group of mice was injected with PBS as control. As can be seen in Fig. 7a and b, treatment with rNDV-Luci through intratumoral injection prolonged survival of the mice significantly (p=0.017) and markedly suppressed the loss of body weight compared with the control group. Although there appeared to be no difference in tumor volume between the intratumorally treated group and control group (Fig. 7c), tumors of surviving mice in the intratumoral group showed more severe skin ulceration and necrosis than the control group (Supplementary Fig. S2). We believe the ulcers were induced by the oncolytic virus, which might interfere with the determination of tumor sizes. However, there were no differences in survival, body weight, and tumor volume between the intravenously treated animals and the control animals (Fig. 7a–c).

Antitumor effect of recombinant NDV. Female athymic mice bearing HeLa cell tumors were treated with rNDV-Luci (5×108 PFU) by intravenous or intratumoral injection. PBS-treated mice were taken as controls.
Discussion
For nonsegmented negative-strand RNA viruses, there is a polar attenuation in transcription that is important for viral replication (Huang et al., 2001). The transcriptional gradient also determines the relative level of expression of foreign genes by the site of insertion within the viral genome (Huang et al., 2003). The replication of recombinant NDV is reported to be delayed by insertion of a foreign gene, which is more prominent when the insertion site is closer to the 3′ proximal end (Zhao and Peeters, 2003). Therefore, construction of a recombinant NDV should strike a balance between exogenous gene expression and viral protein production. We here introduced an extra transcription unit in the noncoding region between the HN and F genes to generate two recombinant NDV strains, rNDV-EGFP and rNDV-Luci. Both recombinants expressed high levels of exogenous protein and showed the same replication features as the parental strain, demonstrating that the insertion of an exogenous gene between HN and F does not disturb the replication cycle of NDV.
As a novel therapeutic strategy to fight tumors, oncolytic virotherapy has developed rapidly. At present, the oncolytic properties of these viruses need improvement to achieve (1) high specificity for tumor tissue, (2) maximal cytolytic effect, and (3) induction of a specific antitumor immune response. The sensitivity of NDV to IFN-α and IFN-β, coupled with the defective IFN signal transduction pathway in tumor cells, provides the specificity of NDV replication in neoplastic tissue (Krishnamurthy et al., 2006; Elankumaran et al., 2010). Furthermore, it has been reported that the oncolytic specificity of NDV for tumor cells is mediated by selectivity for apoptosis-resistant tumor cells (Mansour et al., 2011).
In this study, we showed that the NDV Italien strain and its derived recombinants were still sensitive to cellular antiviral responses that are induced by type I IFN in normal cells, suggesting that they can be safely used in clinical trials. In particular, the defective IFN induction and response pathways, especially through the interferon regulatory factor pathways in hepatocellular (Ghosh et al., 2003; Wang et al., 2009) and cervical carcinoma cell lines (Cheng et al., 2006), endow NDV with the property of tumor-selective replication, which to our knowledge was demonstrated for the first time in a tumor-bearing mouse model by in vivo imaging technology.
Oncolytic virotherapy represents a potent approach to cancer immunotherapy, combining the enhanced release of TAAs via tumor cell death and tumor-associated danger signals, which are a benefit to antitumor immunity (Prestwich et al., 2008). TAA-directed immunotherapy by oncolytic recombinant NDV has been shown to enhance therapeutic efficacy (Vigil et al., 2008). However, the development of specific antiviral immunity is considered to inhibit the spread of virus in tumor tissue, thereby limiting the therapeutic efficiency of NDV, and finally eliminating the virus (Boisgerault et al., 2010). We detected syncytial cell death induced by recombinant NDV Italien, which expresses a foreign protein, as early as 7 hr postinfection, suggesting that the fast cytolytic activity of NDV is able to avoid the specific antiviral immune response that is induced after NDV inoculation.
Our animal study showed a prolonged survival period for intratumorally injected animals, but not for intravenously injected animals. Although tumor volume was not reduced by NDV, it should be noted that animals in both the intratumoral and intravenous groups showed severe, wide areas of ulceration, as shown in Supplementary Fig. S2. We believe that ulceration interfered with the validity of tumor volume and eliminated the potential differences in tumor volume between treated mice and control mice. The ulceration and necrosis induced by NDV in tumor tissues could reduce the tumor burden and might be of significant benefit to mouse survival.
As an effective treatment modality for metastatic tumors, intravenous administration has been proven to be well tolerated and to induce the regression of solid tumors in phase I clinical trials of NDV PV701 (Lorence et al., 2003). However, NDV recognizes target cells through sialic acid, which is present on the surface of a wide range of cells, leading to NDV adsorption by normal tissues after systemic injection. Although in vivo imaging in our study indicated that the remaining virus selectively replicated in tumor tissue, we presume that the amount of virus was inadequate to suppress tumor growth and improve survival. Therefore, construction of tumor-specific recombinant NDV or encapsulation of NDV within microparticles may represent ways to increase the amount of virus in tumor.
In conclusion, we have established a reverse genetics system based on the NDV Italien strain, and we have generated two recombinant NDVs expressing either the EGFP or firefly luciferase reporter gene. Furthermore, we demonstrated the therapeutic efficiency of oncolytic NDV Italien. Therefore, this study provides a rationale and strategy for further studies on recombinant NDV Italien as a potential oncolytic viral vector for human tumor therapy.
Footnotes
Acknowledgments
The authors acknowledge Prof. Volker Schirrmacher (German Cancer Research Center, Germany) for the gift of NDV Italien and rabbit anti-NDV polyclonal antibody. The authors thank Prof. Karl-Klaus Conzelmann (Ludwig Maximilian University, Munich, Germany) for providing BSR-T7/5 cells. This work was funded by the National Natural Science Foundation of China (nos. 30870944 and 81172144), the Hi-Tech Research and Development Program of China (no. 2009CB521706), and the National Science and Technology Major Project (no. 2012AA020806).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.