
Abstract
Airway infiltration by eosinophils is a major characteristic of chronic asthma. CCL11 (eotaxin-1) is secreted by lung epithelial cells and functions as the major chemokine for eosinophil recruitment. Pseudotyped adeno-associated virus (AAV) 2/9, composed by the AAV2 rep and AAV9 cap genes, can efficiently target lung epithelial cells and might carry gene sequences with therapeutic potential for asthma. This study aimed to determine whether pseudotyped AAV2/9 virus carrying the small hairpin RNA targeting CCL11 and expressed by CMV/U6 promoter could reduce eosinophilia and asthmatic responses in mite allergen-sensitized mice. Mice were sensitized by intraperitoneal and challenged by intratracheal injection with recombinant Dermatophagoides pteronyssinus group 2 allergen (rDp2). AAV2/9 viral vectors were intratracheally injected three days before the first challenge. AAV2/9 sh47 virus significantly reduced airway hyperresponsiveness, airway resistance, CCL11 levels, and eosinophilia in the lungs of sensitized mice. Th2 cytokines, including interleukins (IL)-4, IL-5, and IL-10, were also significantly reduced in the bronchoalveolar lavage fluid of AAV2/9 sh47 virus-treated mice. Th2 cytokine levels were also reduced in rDp2-stimulated mediastinal lymphocytes in treated mice. However, serum levels of rDp2-specific IgG1 and IgE, as well as Th2 cytokine levels in rDp2-stimulated splenocyte culture supernatants, were comparable to the sensitized control group. The results suggest that AAV2/9 sh47 virus relieved local instead of systemic inflammatory responses. Therefore, the CMV/U6 promoter with AAV2/9 viral vector, which is preferable to target lung epithelia cells, might be applied as a novel therapeutic approach for asthma.
Introduction
With the importance of small interfering RNA and microRNA in eukaryotic mRNA regulation, RNA interference has been proposed as a new therapeutic strategy for human diseases (Dykxhoorn and Lieberman, 2006). Small hairpin RNA (shRNA) can be efficiently transcribed into small interfering RNA in mammalian cells via the U6 or hybrid CMV/U6 promoter. The hybrid promoters, CMV/U6 or CMV/H1, showed better shRNA expression efficiency than U6 or H1 promoter (Hassani et al., 2007; Su et al., 2008). Adeno-associated virus (AAV), a single-stranded DNA virus that belongs to the Parvoviridae family, is characterized by its nonpathogenic nature, nonintegrating episomal vector genome, and ability to confer stable gene expression (Daya and Berns, 2008). Additionally, AAV has the advantage of infecting both dividing and nondividing cells (Alexander et al., 1996). With new AAV strains being continuously isolated and characterized (Rutledge et al., 1998; Auricchio et al., 2002; Gao et al., 2004; Mori et al., 2004), pseudotyped recombinant AAVs can be derived with rep and cap genes from different serotypes. Since pseudotyped AAVs gain the ability to infect cells that are more susceptible to other serotypes, they had been applied to enhance gene expression and direct tissue tropism (Rabinowitz et al., 2002). AAV5 (Zabner et al., Auricchio et al., 2002), AAV6 (Halbert et al., 2001), and AAV9 (Gao et al., 2004) can efficiently infect epithelial cells. Preferential alveolar cell infections by AAV9 were reported in the last few years (Limberis and Wilson, 2006; Limberis et al., 2009). Therefore, pseudotyped AAVs containing CAP proteins of these serotypes may deliver therapeutic genes and achieve long-term gene expression in the lungs of asthmatic model animals.
In this study, we generated an IRES/hrGFP reporter gene system at the downstream of hybrid CMV/U6 promoter of shRNA expression cassette. The hybrid CMV/U6 promoter construct had better gene knockdown efficacy than that of either U6 or CMV promoter construct. Notable, hrGFP positive cells represented the shRNA expressing cells. We subsequently applied pseudotyped AAV2/9 viral vector carrying shRNA, which was driven by CMV/U6 promoter, targeting the CCL11 gene to the lungs of rDp2-sensitized mice. Data demonstrate significant reduction of AHR, eosinophilia, and local Th2 cytokine levels, but the vector did not affect systemic immune responses. These results suggest that CMV/U6 promoter has greater gene silencing efficacy and pseudotyped AAV2/9 efficiently delivered the shRNA sequence targeting CCL11 to alleviate airway inflammation.
Materials and Methods
shRNA construction
The shRNA sequences (Supplementary Table S1; Supplementary Material available online at

Reduction of CCL11 gene expression after infection with adeno-associated virus (AAV)-small hairpin RNA (shRNA) vector viruses in vitro. CCL11-NIH 3T3 cells that constitutively expressed CCL11 were generated by transfecting pcDNA3.1-CCL11 into NIH 3T3 cells with G418 selection.
Preparation of recombinant adeno-associated viruses
The recombinant pseudotyped adeno-associated virus (rAAV) was generated with the tri-plasmids system (Limberis and Wilson, 2006), including plasmids containing hrGFP, and each of different promoters with shRNA, pHelper, and pAAV2/9 (containing AAV2 rep and AAV9 cap genes). The plasmid/turbofect complex was cultured with 293T cells for 72 hrs, according to the manufacturer's instructions. The cells were then lysed using the freeze/thaw method. The virions were purified from cell lysates by caesium chloride (CsCl) gradient centrifugation, concentrated, and dialyzed against phosphate buffered saline (PBS) with Amicon Ultra-15 centrifugal filters (Millipore, Billerica, MA). The titer, presented as genome copies (GC), of rAAV viruses was determined by real-time polymerase chain reaction (PCR) analysis, based on the hrGFP-specific primer sequences.
Real-time PCR for viral genome copies (GC) determination
Viral GCs were determined by real-time PCR. Primers for hrGFP were used (forward primer, 5′-TGAACAGCGGCAAGTTCTACAG-3′; reverse primer, 5′-GTGCTGGATG AAGTGGTACTCG-3′). Virions were pre-treated with protease K (Roche Applied Science, Mannheim, Germany) to release the viral genome. The viral genome was then mixed with SYBR green qPCR Master mix (Fermentas, Ontario, Canada) for analysis. The real-time PCR steps were as follows: initial denaturation at 95°C for 10 min, followed by 40 cycles of 95°C for 10 s, 60°C for 10 s, and 72°C for 10 s.
Determining the efficacy of shRNA in vitro
A murine CCL11 cDNA fragment was cloned into a pcDNA3.1 plasmid (Invitrogen, Carlsbad, CA); the resultant plasmid was designated as pcDNA3.1-CCL11 and transfected into NIH 3T3 cells. After G418 selection, 1×105 cells/well were infected with AAV2/9 shRNA and shRNA control viruses (5×1010 GC) for 48 hrs. Next, the supernatant was collected and subjected to CCL11 enzyme-linked immunosorbent assay (ELISA).
Animals and sensitization protocol
The recombinant Dermatophagoides pteronyssinus group II allergen (rDp2) was purified from a yeast clone (Tsai et al., 2000). Female BALB/c mice (8 to 10 weeks old) were sensitized by intraperitoneal injection of 40 μg rDp2 or normal saline mixed with aluminum hydroxide on experimental days 1, 7, 14, and 21. Virus-treated mice received 5×1011 GC in 50 μl saline on day 25. Mice were challenged with intratracheal injection of 40 μg rDp2 or normal saline on days 28 and 34. AHR and airway resistance was determined on day 35, and mice were sacrificed on day 36. Care and handling of mice were according to the guidelines of the Animal Care Committee of Chang Gung University and the National Institutes of Health (NIH) Guidelines for the Care and Use of Laboratory Animals.
Measurement of airway hyperresponsiveness
Mice inhaled cumulatively increased concentrations of methacholine (10, 20, 30, and 40 mg/ml) (Sigma, St. Louis, MO) for 3 minutes each to induce airway contraction. Airway hyperresponsiveness (AHR) was measured 24 hrs after the final challenge. Mice were placed into the chamber and the enhanced pause (Penh) was analyzed with a whole-body plethysmograph and BioSystem XA software (Buxco Electronics, Troy, NY).
Airway resistance
Mice inhaled cumulatively increased concentrations of methacholine (1.1, 3.3, 10, and 30 mg/ml) for 30 sec each and then were immediately placed into the chamber to record the resistance index (RI) for 3 min. Airway resistance was measured using Buxco FinePointe™ RC.
Detection of rDp2-specific IgG1 and IgE antibodies
Serum rDp2-specific IgG1 and IgE levels were determined by ELISA. Briefly, the rDp2-coated plates (10 μg/ml; Corning, Corning, NY) were first incubated with diluted serum samples. Next, biotinylated rat anti-mouse IgG1 or IgE monoclonal antibodies (BD PharMingen, San Diego, CA) and streptavidin-conjugated horseradish peroxidase (HRP) (Sigma) were added sequentially. The plates were incubated with TMB substrate (R&D Systems, Minneapolis, MN), and the absorbance was determined with an ELISA reader at 450 nm.
Lung histology and immunohistochemistry
Ten-micrometer-thick lung sections were fixed with 4% formaldehyde and subjected to hematoxylin and eosin, Trichrome, and periodic acid-Schiff staining. The hrGFP expression in lung tissues was determined by immunohistochemistry with rabbit anti-hrGFP antibody (Stratagene), as previously described (Yang et al., 2009). The percentages of eosinophil infiltration (defined as inflammatory index), goblet cell hyperplasia, and collagen deposition were analyzed by MetaMorph microscopy automation and image analysis software (Molecular Devices, Sunnyvale, CA). Data were analyzed from three sections of each mouse lung and five selected areas with 100 μm square size in each section. The data were calculated using the mean values of sensitized control mice as 100%.
Assay of Th2 cytokines and CCL11
The concentrations of interleukin (IL)-4, IL-10, IL-13, and CCL11 were determined with Duoset ELISA kits (R&D Systems, Minneapolis, MN). IL-5 was analyzed by BD Pharmingen ELISA set (BD PharMingen). Assays were performed according to the manufacturers' instructions.
Statistical analysis
Results are presented as mean±SEM. Data were analyzed with one-way ANOVA Tukey test for multiple-group comparisons. The P values less than 0.05 were considered statistically significant.
Results
Reduction of CCL11 level by AAV2/9 shRNA viral vectors in CCL11-NIH 3T3 cells
Three CCL11-targeting shRNA expression promoter constructs were generated containing CMV/U6, CMV, or U6 promoters (Fig. 1A). The results showed that AAV2/9 CMV/U6 and CMV, but not U6, promoter-based virus induced hrGFP expression in virus-infected NIH 3T3 cells (Fig. 1B). The inhibitory efficacy of pseudotyped AAV2/9 vector viruses containing CCL11 shRNA sequences derived from each different promoter construct was examined in CCL11-transformed NIH 3T3 cells (Fig. 1C and E). A dose-dependent CCL11 reduction was observed with different CCL11 shRNA expressing AAV2/9 viruses. High and medium doses of AAV2/9 CMV/U6 and AAV2/9 U6 sh47 viruses significantly reduced CCL11 level compared to that of AAV2/9 CMV sh47 virus (CMV/U6 and U6 vs. CMV, both P<0.001). Compare to the AAV2/9 U6 sh47 virus, medium-dose treatment of AAV2/9 CMV/U6 sh47 virus had better gene inhibitory efficacy. The different doses of AAV2/9 shLuc control (with shRNA sequence against luciferase) viruses did not affect CCL11 levels. The results suggest that hybrid CMV/U6 promoter construct not only to be more efficient for gene knockdown than either U6 or CMV promoter construct, but also to have the advantage to trace targeted tissues with the expression of reporter genes. Therefore, this construct with CMV/U6 promoter was applied to drive shRNA in the following studies.
The inhibitory efficacy of pseudotyped AAV2/9 vector viruses containing CCL11 shRNA sequence (sh47, sh124, or sh137) driven by CMV/U6 promoter was examined in CCL11-NIH 3T3 cells (Fig. 1F). Significant reduction of CCL11 levels was observed in AAV2/9 sh47 and sh137 virus-infected cells, compared with non-infected shLuc control, or sh124-infected cells (sh47 and sh137 vs. sh124, non-infected and shLuc control, all P<0.001). Based on these results, we used AAV2/9 sh47 and sh137 viral vectors for further experiments.
Significant reduction of eosinophilia and CCL11 level in bronchoalveolar lavage fluid from AAV2/9 sh47 AAV vector virus-treated mice
To test whether the pseudotyped AAV2/9 vector viruses were able to reduce CCL11 expression and the recruitment of eosinophils in vivo, the AAV vector viruses were introduced into rDp2-sensitized mice (Fig. 2A). The results showed that total cell number in bronchoalveolar lavage fluid (BALF) was dramatically increased in rDp2-sensitized mice compared with normal control mice (rDp2-sensitized groups vs. normal control, P<0.01; Fig. 2B). Although the total cell number in AAV2/9 sh47 and sh137 virus-treated mice was slightly decreased compared with sensitized control and shLuc control mice, a significant reduction of eosinophilic infiltration was detected in AAV2/9 sh47 virus-treated mice, compared with sensitized control mice (P<0.05; Fig. 2C).
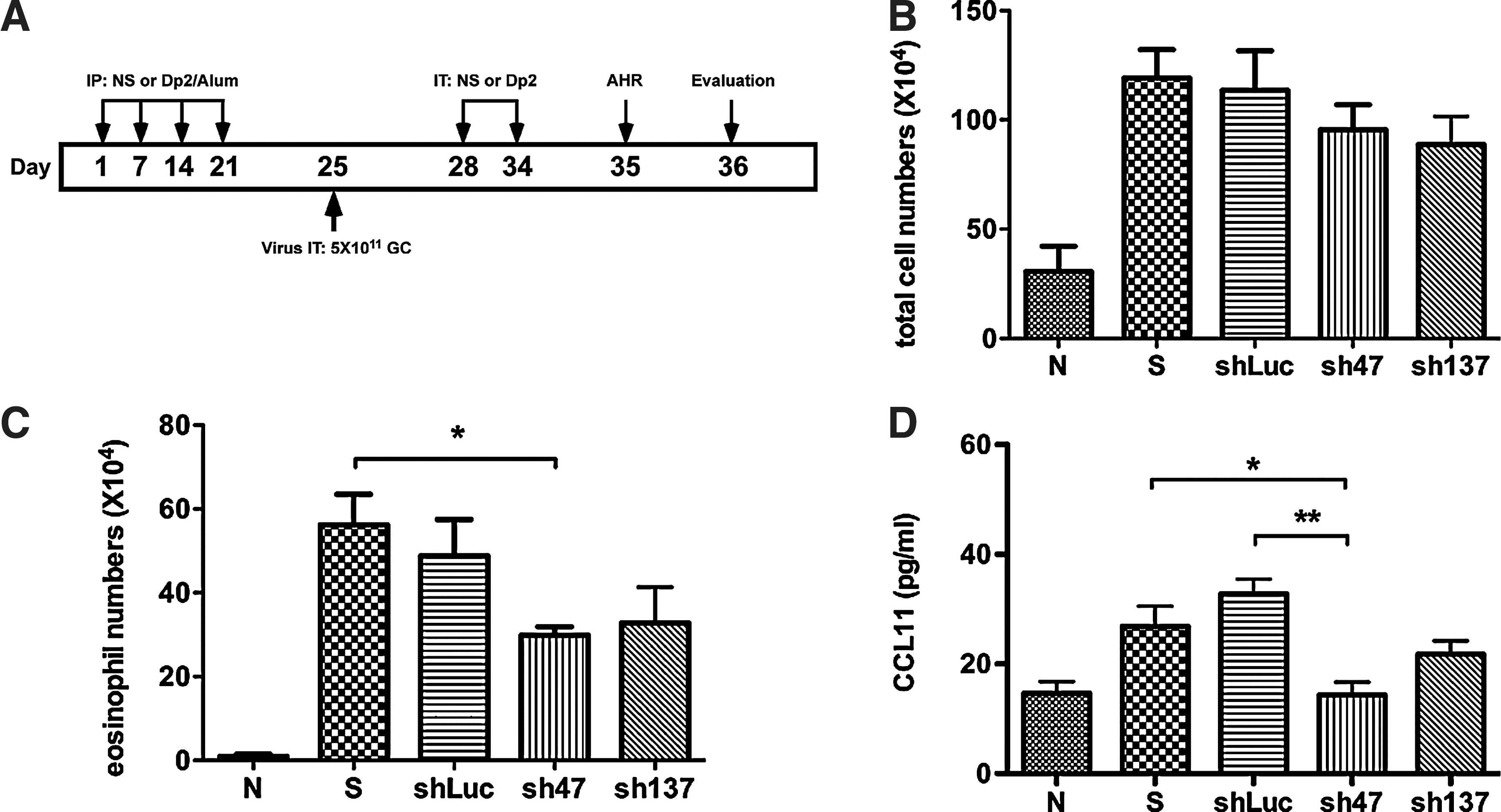
AAV2/9 sh47 virus reduced total cell numbers, eosinophilia, and CCL11 in bronchoalveolar lavage fluid (BALF) from rDp2-sensitized mice.
Compared with the sensitized control group, the mice treated with AAV2/9 sh137 virus experienced a slight, statistically insignificant reduction of eosinophils. In addition, a significantly reduced CCL11 level was detected in BALF from AAV2/9 sh47 virus-treated mice compared with sensitized and shLuc control mice (P<0.05 and P<0.01, respectively; Fig. 2D). The BALF CCL11 level in sh47 virus-treated mice was comparable to that of normal control mice. A slightly reduction, but no significant difference of CCL11, was detected in AAV2/9 sh137 virus-treated mice, compared with sensitized and shLuc control mice. Taken together, these data indicate that the AAV2/9 sh47 virus was more effective than the AAV2/9 sh137 virus at reducing eosinophilic infiltration and BALF CCL11 levels in rDp2-sensitized mice.
Significant reduction of AHR and airway resistance in AAV2/9 sh47 virus-treated mice
Next we evaluated the efficacy of pseudotyped AAV vector viruses on AHR and airway resistance, which are the key parameters used to assess asthma severity. After sensitization and challenging mice with rDp2, AHR was measured by methacholine challenge using non-invasive whole body plethysmography (Fig. 3A). When mice were challenged with 40 mg/ml methacholine, the Penh value was significantly lower in AAV2/9 sh47 virus-treated mice than in sensitized control mice (P<0.05), but was reduced only slightly compared with AAV2/9 shLuc control mice. Airway resistance was measured by methacholine challenge using FinePointe™ RC, an invasive measurement (Fig. 3B). The virus-treated mice also had significantly lower RI values than sensitized control (P<0.01) and AAV2/9 shLuc control (P<0.05) mice when responded to 10 mg/ml methacholine. The data show that AAV2/9 sh47 virus is able to reduce AHR and airway resistance in the airways of rDp2-sensitized mice.
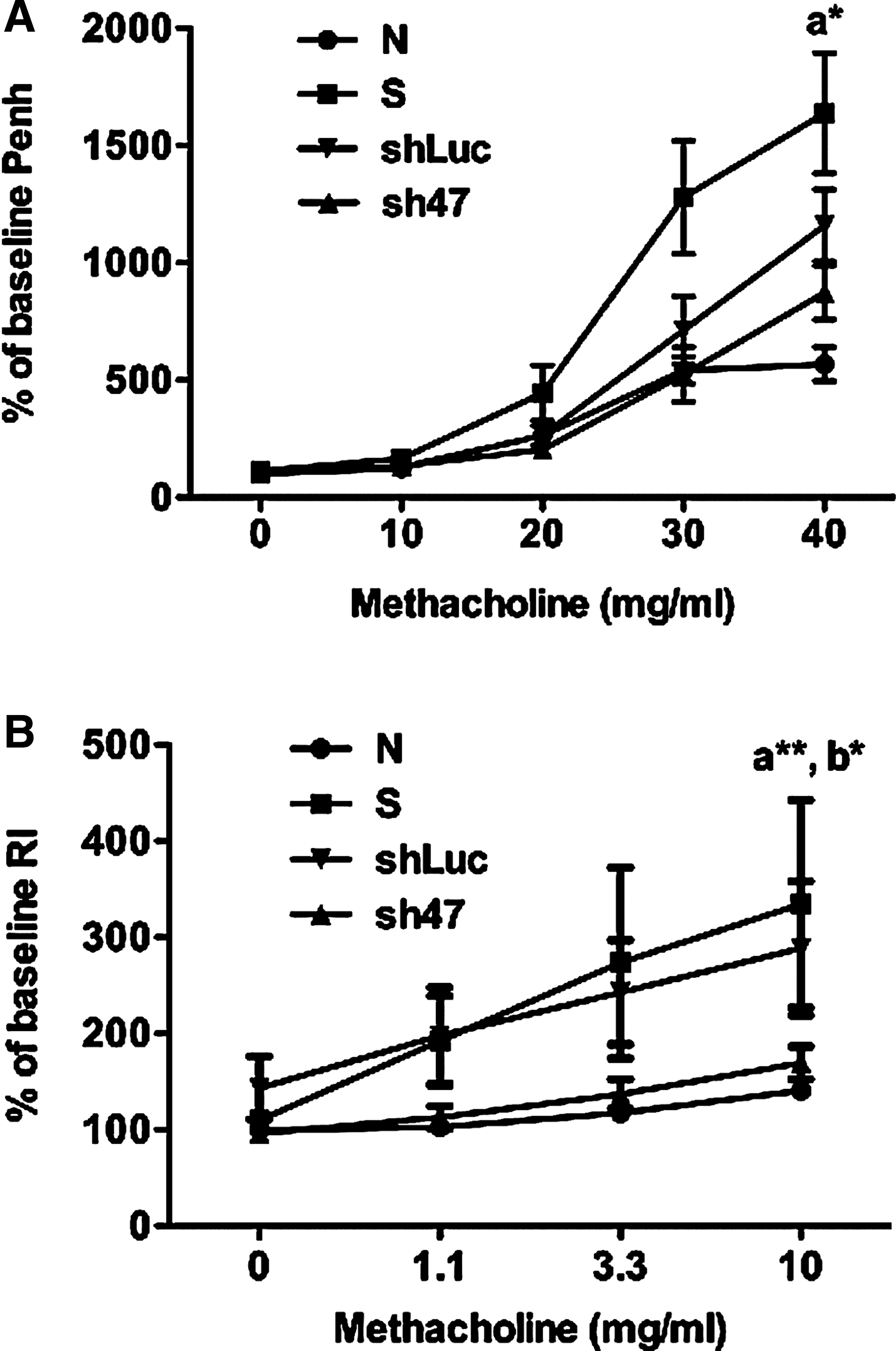
Reduction of airway hyperresponsiveness and airway resistance with CCL11-specific AAV2/9 shRNA.
rDp2-induced tissue remodeling was rescued in the lungs of AAV2/9 sh47 virus-treated mice
We also examined whether AAV2/9 sh47 virus could affect lung tissue remodeling. Formaldehyde-fixed, paraffin-embedded lung samples were sliced and subjected to hematoxylin/eosin (HE), Trichrome, and periodic acid-Schiff staining (Fig. 4). More eosinophils were infiltrated into the lungs of sensitized and shLuc control mice than normal control mice (Fig. 4A). The quantitative results indicate significant reduction of eosinophils infiltrated into the lungs (presented as inflammatory index) of AAV2/9 sh47 virus-treated mice, compared with sensitized and shLuc control mice (Fig. 4E). Additionally, significant reduction of collagen deposition (Fig. 4B and F) and goblet cell hyperplasia (Fig. 4C and G) was also observed in the lungs of AAV2/9 sh47 virus-treated mice. These results demonstrate that AAV2/9 sh47 viruses reduced eosinophilia, collagen deposition, and goblet cell hyperplasia in the lungs of rDp2-sensitized mice. Immunohistochemical staining specific for hrGFP indicated that alveolar epithelial cells were targeted by AAV2/9 viral vectors (Fig. 4D).
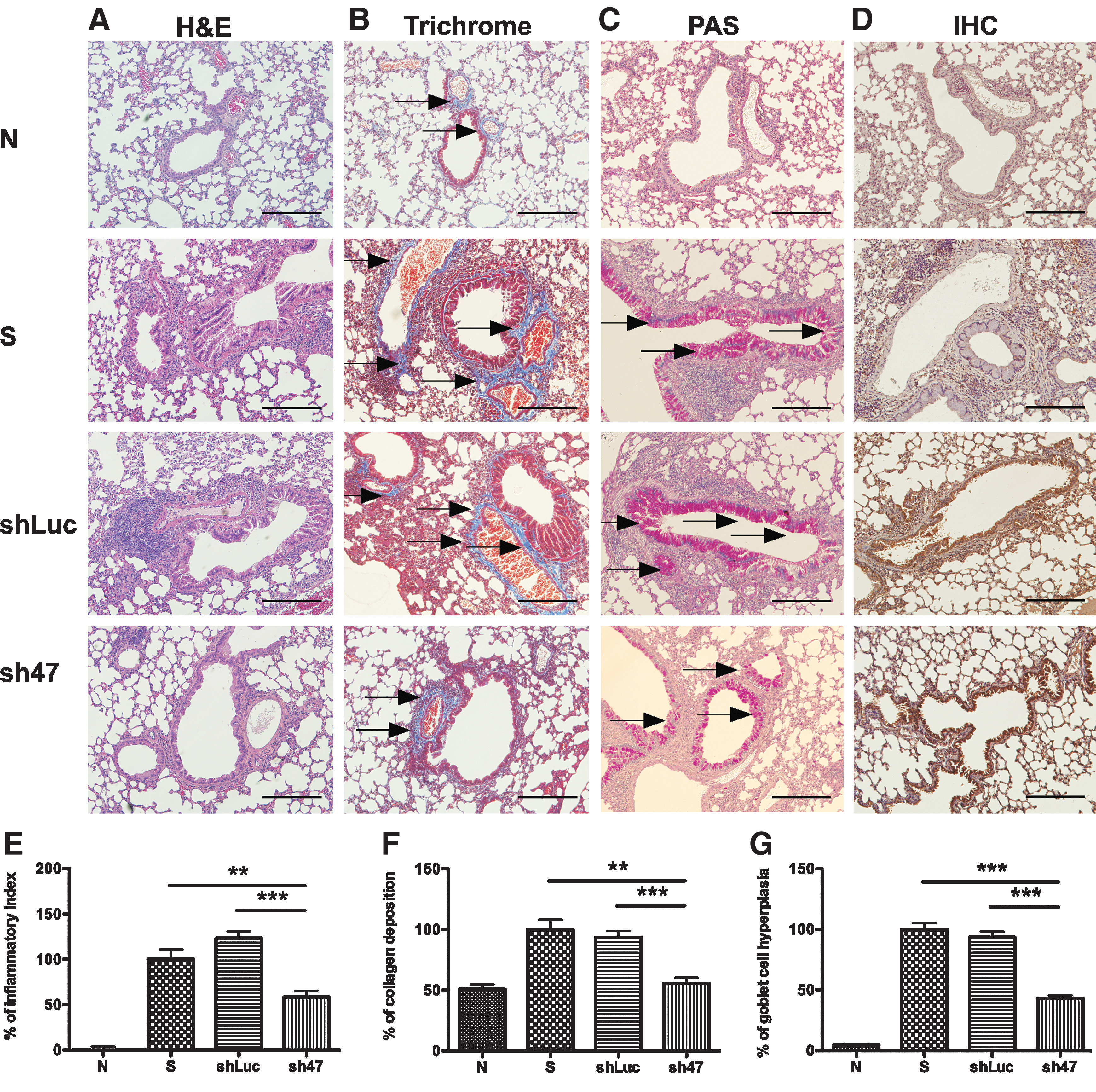
Histopathology of lung tissues in mice treated with AAV2/9 shRNA vector viruses. Formaldehyde-fixed, paraffin-embedded lung samples were sliced and subjected to
Reduction of local instead of systemic immune responses in AAV2/9 sh47 virus-treated mice
We also explored whether the AAV2/9 sh47 virus affected Th2 cytokine levels in BALF and rDp2-stimulated lymphocytes in mediastinal lymph nodes (mLN) (Fig. 5). The levels of IL-4, IL-5, IL-10, and IL-13 were significantly lower in BALF from AAV2/9 sh47-treated mice than from sensitized control mice (P<0.05). Compared to shLuc control mice, significantly low levels of IL-4, IL-10, and IL-13 in BALF from AAV2/9 sh47 virus-treated mice were also observed. The rDp2-stimulated mLN lymphocytes of AAV2/9 sh47 virus-treated mice had significantly lower Th2 cytokine production than those of sensitized control mice (IL-4, P<0.01; IL-5, IL-10, and IL-13, P<0.05) and shLuc control mice (all P<0.05). The cell populations in mLNs were also analyzed by flow cytometry. The results showed no cell distribution differences among normal, sensitized control and virus-treated mice (data not shown). To examine the effects of the AAV2/9 sh47 virus on systemic immune responses to rDp2 allergen, the serum levels of rDp2-specific antibodies (IgG1 and IgE) and the cytokine levels of rDp2-activated splenocytes were also measured. Compared with the sensitized control mice, virus-treated mice had slightly reduced rDp2-specific IgE levels (P=0.11) and similar rDp2-specific IgG1 levels were observed (Fig. 6). The cytokine concentrations in the supernatants of rDp2-stimulated splenocytes were comparable between AAV2/9 sh47 virus and control mice (data not shown). The data suggest that AAV2/9 sh47 virus modulated local instead of systemic immune responses in rDp2-sensitized mice.
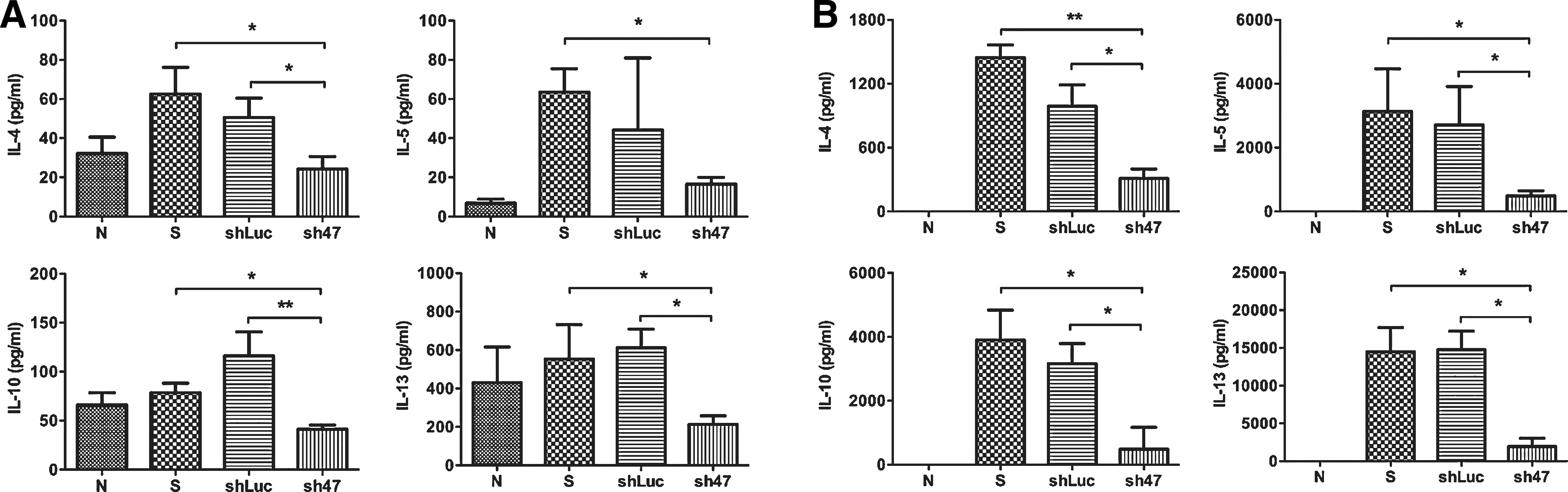
AAV2/9 sh47 virus reduced Th2 cytokine levels in bronchoalveolar fluids and the supernatants of lymphocytes from rDp2-stimulated mediastinal lymph nodes in rDp2-sensitized mice.
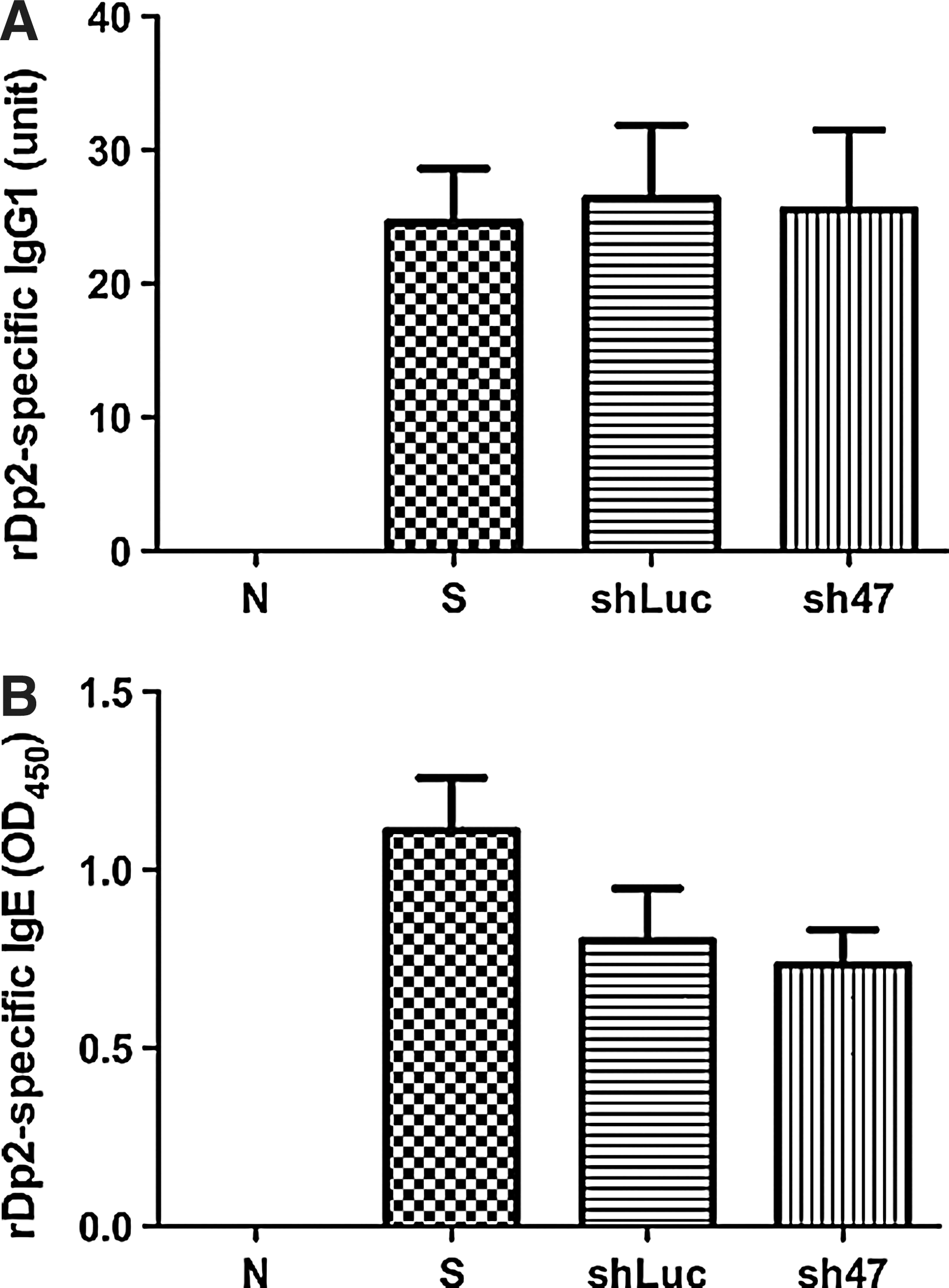
Serum
Discussion
The results in this study demonstrate for the first time that gene transfer using pseudotyped AAV2/9 with shRNA driven by CMV/U6 promoter-reduced specific gene expression in lung tissues alleviate airway inflammation in allergen-sensitized mice. In addition, the constructs containing hrGFP reporter gene system allowed the detection of shRNA-expressing cells or tissues. With the reduction of CCL11 expression in the lungs, most asthmatic parameters were significantly reduced in AAV shRNA virus-treated mice; while Dp2-specific serum IgE and IgG1 levels and Dp2-stimulated Th2 cytokines were not affected. This result provides support for the notion that the pseudotyped AAV2/9 virus is a potential therapeutic vector for the treatment of asthma, without targeting systemic immune responses.
Eosinophils release several inflammatory mediators and are considered the key effector cells in the progression of lung inflammation. Eotaxins, especially CCL11 (expressed primarily by epithelial cells), is the primary chemoattractant ligand that binds to the CCR3 receptor on eosinophils and activated T lymphocytes. Significant reduction of eosinophilia has been demonstrated in the BALF of ovalbumin (OVA)-sensitized CCL11-knockout (Rothenberg et al., 1997) or CCL11-knockdown mice (Huang et al., 2009). However, comparable eosinophil numbers and AHR in CCL11 deficient mice (Tomkinson et al., 2001) argued the role of CCL11 in the development of allergen-induced eosinophilic airway inflammation. Moreover, the findings that the administration of anti-CCL11 antibody at the provocation stage was ineffective at reducing eosinophil infiltration (Campbell et al., 1998), but was effective in reducing eosinophil infiltration throughout the challenge period (Gonzalo et al., 1998), suggest that CCL11 has a transient effect on eosinophil infiltration. Therefore, a stable knockdown of the CCL11 gene seems required to consistently suppress eosinophil infiltration and AHR in response to allergen challenges.
Although shRNA expression is usually directed by polymerase III promoters, such as U6 or H1, better gene inhibitory efficacy has been demonstrated with the hybrid CMV/U6 or CMV/H1 promoters, compared to single U6 or H1 promoter (Hassani et al., 2007; Su et al., 2008). Similar results were also observed in the current study. Moreover, the IRES-hrGFP DNA sequence at the downstream of the CMV/U6 shRNA expression cassette assisted the detection of shRNA expressing cells or tissues. The shRNA sequences, including sequences for IL-4 and IL-13 (Zavorotinskaya et al., 2003), IL-5 (Huang et al., 2008), IL-1 receptor antagonist (Wang et al., 2006), and chitinase (Yang et al., 2009), have been employed in some studies to reduce inflammation responses in asthmatic animals. Most experimental animal models used OVA as an allergen to provoke airway inflammatory responses. However, the current study applied rDp2 allergen derived from the house dust mite, the most common trigger factor for asthmatic patients (Wills-Karp, 1999). Although a few other reports have also applied rDp2 to induce allergic responses in mice, these studies did not characterize eosinophil infiltration or CCL11 levels in BALF (Tsai et al., 2002), or the severity of AHR (Li et al., 2006; Ou-Yang et al., 2009). Our data suggest a comprehensive animal model in which airborne allergen successfully induced AHR, eosinophil infiltration, and Th2-related immune responses.
In addition to the allergen usage in animal models, significant reductions of AHR and lung eosinophils have been demonstrated with shRNA against CCL11, which was delivered by lentivirus vector (Huang et al., 2009). However, the changes that Th2-type cytokines undergo in BALF or in OVA-stimulated lymphocytes of mLNs were not addressed. In addition, the use of lentivirus makes it difficult to control specific tissue targeting and can never exclude the possibility of mutagenesis (Heilbronn and Weger, 2010). A human clinical trial directly applied antisense oligonucleotide against CCR3, the receptor for CCL11, to patients with mild atopic asthma (Gauvreau et al., 2008). The results indicated that only early-phase response (not late-phase response) was reduced. Moreover, antisense therapy only reduced CCR3 mRNA, without affecting cell surface CCR3 protein expression. More efficient gene transfer approaches or optimal doses of antisense oligonucleotides remain to be determined to achieve effective treatment for asthma.
Greater systemic than lung effects were demonstrated with the intra-tracheally delivered antisense RNA oligonucleotides (ASOs) (Moschos et al., 2011). The relative high amount of ASOs used, and the potentially sequestered ASOs in circulation, might lead to more systemic effects. Although we cannot absolutely rule out the possibility that the shRNA in our study might reach other tissues, and more studies are required to clarify if there is any unanticipated off-target effects of shRNA used in the current study, we believe that pseudotyped AAV2/9-delivered shRNA specifically targeted CCL11 in lung epithelial cells. Our data provide a stable and tissue-specific gene transfer system that employs shRNA against CCL11 that mainly expressed in the epithelial cells.
In addition to the reduction of airway resistance and local Th2 responses, collagen deposition and goblet cell hyperplasia in the lungs were also significantly relieved. Serum Dp2-specific IgE, the systemic immune response, was not affected after AAV2/9 virus treatment. Similar results were also observed in mice treated with intranasal (IN) administration of lentivirus-delivered shRNA against CCL11 (Huang et al., 2009). In contrast, total or antigen-specific serum IgE levels were significantly reduced in animals treated with the intramuscular (IM) injection of AAV-delivered IL-10 (Mueller et al., 2009) or gene sequences targeting the soluble receptors of IL-13 and IL-17e (Mueller et al., 2010). These controversial results of serum IgE levels may be due to the different routes of AAV administration or the cytokine vs. chemokine genes, which have more local effects.
With the capacity for large inserted DNA segments, the use of adenovirus to carry therapeutic genes has been commonly proposed for the treatment of airway diseases (Barnett et al., 2002). Most studies aimed to apply adenovirus to transfer immune modulatory genes, such as IL-10, IL-12 (Hsu et al., 2010), or uteroglobin-related protein 1 (Chiba et al., 2006), in the control of allergic inflammation in mice. The adenoviral vectors carrying therapeutic shRNA to control asthmatic responses have only been described targeting IL-5 (Huang et al., 2008) or IL-5 receptor-α (Mao et al., 2006). Moreover, the reduction of lung inflammation was achieved with the adenovirus transducing shRNA against C5aR (Sun et al., 2009). Similar to lentiviral vectors, broad tissue tropism becomes a limitation to reduce the off-target effect of adenovirus vectors. The strong response of the host immune system to capsid proteins is another potential disadvantage of using adenovirus vectors (Douglas, 2007).
A successful gene-delivery system must efficiently infect target tissue and establish long-term gene expression. Because it is more resistant to extreme conditions and ordered genomic structure, the AAV vector readily emerges as a tool for gene transfer into animals. Recombinant AAV2 vectors have been applied to preclinical studies for a variety of diseases, such as hemophilia, α1 anti-trypsin deficiency, cystic fibrosis, and rheumatoid arthritis (Warrington and Herzog, 2006). Stable gene expression has also been detected for longer than 1.5 years (Song et al., 2001). The ability to infect both dividing and nondividing cells (Alexander et al., 1996) makes AAV a powerful vector to transduce exogenous gene sequences into epithelial cells that are slow dividing cells (Leigh et al., 1995).
Zavorotinskaya et al. showed that intratracheal injection of AAV2 vector carrying IL-4 receptor antagonist significantly inhibited the development of airway eosinophilia and Th2-type cytokines in BALF. The transfer of T-bet by AAV2 vector balanced the Th1/Th2 cytokine profile in OVA-induced allergic mice (Wang et al., 2008). AAV2-mediated expression vector containing shRNA vector specific to chitinase has been applied to the treatment of asthmatic mice (Yang et al., 2009). Although AAV2 has been the most commonly used AAV prototype (Goncalves, 2005), pseudotyped recombinant AAVs that are composed with rep and cap genes from different serotypes have been applied to enhance gene expression and controlled tissue tropism (Halbert et al., 2000). Halbert et al. demonstrated the higher level and longer gene expression of both AAV2/5 and AAV2/9 compared with AAV2/2 in lung tissues (Halbert et al., 2001). In addition, AAV9 vector has been applied to the treatment of human disease models and recently has been suggested to be beneficial for liver and brain gene delivery (Chen et al., 2009; Xue et al., 2010).
Since transcytosis of some AAV serotypes has been shown in airway epithelial cells (Di Pasquale and Chiorini, 2006), the AAV2/9 vector used in this study may also have the potential to cross epithelial cells or capillary barriers to deliver the vector to other tissues. It would be interesting to evaluate and to compare the effect of AAV9 with local or system delivery routes in the future. With the limited cell types for CCL11 expression, we believe that the AAV2/9-delivered shCCL11 specifically reduced CCL11 RNA levels in the lung tissues of rDp2-sensitized mice. In a bacterial infection study of mouse lungs, the AAV2/9 vector showed more sufficient gene expression and resistance to second administration than AAV2/5 for the transduction of mouse lung (Limberis and Wilson, 2006). The authors also suggested that AAV2/9 is better at preventing the activation of mucosal immunity.
Similarly, we observed no effective gene transfer efficiency when the AAV2/5 vector was used in rDp2-asthmatic animals (data not shown). Our data demonstrated that AAV2/9-delivered hrGFP expression was observed in alveolar epithelial cells (Fig. 4D). The preferential alveolar epithelium gene delivery by AAV2/9 viral vectors has been described (Limberis and Wilson, 2006; Limberis et al., 2009 ; Bell et al., 2011). Since type I alveolar epithelial cells cover>98% of internal surface area of the lung, they are the major source of eotaxin after antigen stimulation (Gonzalo et al., 1996; Gonzalez et al., 2005 ). Thus, we believe that pseudotyped AAV2/9 sh47 viruses infected type I alveolar epithelial cells and effectively reduced CCL11 expression in rDp2-sensitized mice.
In conclusion, the CMV/U6 promoter shRNA expression cassette showed improved gene inhibitory efficacy and hrGFP reporter gene expression. A single treatment of AAV2/9 shRNA against CCL11 significantly reduced asthmatic responses and airway remodeling in rDp2-sensitized mice. That serum rDp2-specific immunoglobulin concentration and cytokines from rDp2-stimulated splenocytes were unaffected suggests the local modulation of immune responses to allergen challenge. This experimental study suggests the use of pseudotyped AAV2/9 vector with CMV/U6 promoter can be further applied to long-term shRNA expression for the management of asthma.
Footnotes
Acknowledgments
We deeply appreciate Dr. James M. Wilson at the University of Pennsylvania and Dr. Kaw-Yan Chua at The National University of Singapore for generously providing pAAV2/9 vector and Dp2-yeast clone, respectively. The authors also thank Dr. Ching-Ping Tseng, Department of Medical Biotechnology and Laboratory Science for his assistance in the generation of shRNA constructs; Dr. Min-Chi Chen, Dept. of Public Health, for her assistance of the statistic analyses; and Dr. Tse-Ching Chen at Chang Gung Memorial Hospital for his assistance in reviewing cell types in histology analyses. This study was supported, in part, by grants from National Health Research Institute NHRI-EX(91-95)-9110SC and NHRI-EX96-9629SI); Department of Education (EMRPD190081); National Science Council (NSC98-2320-B-182-030-MY3 and NSC95-2320-B-182-043-MY3) of Republic of China; and Chang Gung Memorial Hospital (CMRPD190511).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.