
Abstract
Crigler-Najjar syndrome is an autosomal recessive disorder with severe unconjugated hyperbilirubinemia due to deficiency of bilirubin UDP-glucuronosyltransferase isozyme 1A1 (UGT1A1) encoded by the UGT1A1 gene. Current therapy relies on phototherapy to prevent life-threatening elevations of serum bilirubin levels, but liver transplantation is the only permanent treatment. Muscle-directed gene therapy has several advantages, including easy and safe access through simple intramuscular injections, and has been investigated in human clinical trials. In this study, we have investigated the efficacy of adeno-associated viral (AAV) vector-mediated muscle-directed gene therapy in the preclinical animal model of Crigler-Najjar syndrome, that is the Gunn rat. Serotype 1 AAV vector expressing rat UGT1A1 under the control of muscle-specific creatine kinase promoter was injected at a dose of 3×1012 genome copies/kg into the muscles of Gunn rats and resulted in expression of UGT1A1 protein and functionally active enzyme in injected muscles. AAV-injected Gunn rats showed an approximately 50% reduction in serum bilirubin levels as compared with saline-treated controls, and this reduction was sustained for at least 1 year postinjection. Increased excretion of alkali-labile metabolites of bilirubin in bile and urine was detected in AAV-injected animals. High-performance liquid chromatography analysis of bile from AAV-injected Gunn rats showed a metabolite with retention time close to that of bilirubin diglucuronide. Taken together, these data show that clinically relevant and sustained reduction of serum bilirubin levels can be achieved by simple and safe intramuscular injections in Gunn rats. AAV-mediated muscle directed gene therapy has potential for the treatment of patients with Crigler-Najjar syndrome type 1.
Introduction
Although correction of the deficient enzymatic activity in the affected organ, that is, the liver, would be the most straightforward, expression within an ectopic tissue, different from the natural production site, is an attractive option for clearance of toxic metabolites from the circulation. Interestingly, transplantation of small bowel and kidney, which also express UDP-glucuronosyltransferase (UDPGT), was effective at reducing hyperbilirubinemia in Gunn rats, thus suggesting that expression of the enzyme in nonhepatic sites is sufficient for metabolic correction of the disease (Kokudo et al., 1999).
Muscle has been the preferred target for gene transfer because of its simple access by intramuscular injections and safety. Previous studies have shown reduction of hyperbilirubinemia in the Gunn rat by pDNA delivery into skeletal muscle via limb perfusion (Danko et al., 2004; Jia and Danko, 2005a). In those studies injections of pDNA expressing human UGT1A1 under the control of the cytomegalovirus (CMV) promoter resulted in excretion of bilirubin glucuronosides in bile and short-term reduction of serum bilirubin lasting for 2 to 4 weeks. Loss of correction was associated with a decrease in UGT1A1 protein in muscle, while pDNA and transcript were detectable 4 weeks after gene delivery. Longer correction required repeated pDNA delivery achieved by a relatively invasive procedure and immunosuppression (Danko et al., 2004; Jia and Danko, 2005a).
AAV vectors are ideal candidates for muscle-directed gene therapy and have shown encouraging preclinical results in various disease models. In one trial, testing the safety and efficacy of intramuscular administration of AAV2/2 in patients with severe hemophilia B, there was no evidence of local or systemic toxicity up to 40 months after injection (Kay et al., 2000; Manno et al., 2003). AAV vector sequences and local factor IX (F.IX) expression was found in muscle biopsies by PCR/Southern blot and immunohistochemical analyses, respectively, up to 3.7 years after vector administration (Jiang et al., 2006). However, despite evidence of gene transfer and expression, circulating levels of F.IX were less than 2% in all subjects and mostly less than 1%, thus lacking clinical benefit. A clinical trial testing the safety of intramuscular administration of AAV2/1 vectors in patients with α1-antitrypsin deficiency showed long-term gene expression despite evidence of an immune response to AAV1 capsids (Brantly et al., 2009), suggesting the presence of a cytotoxic T lymphocyte (CTL) response to AAV capsids that does not result in the elimination of transduced cells (Brantly et al., 2006). T cell immune responses against the AAV1 capsid were detected in half of the subjects with lipoprotein lipase (LPL) deficiency receiving intramuscular administration of AAV2/1 vector during the first months after vector administration (Mingozzi et al., 2009). This immune response was transient and associated with an increase in the muscle enzyme creatinine phosphokinase (CPK) about 4 weeks after gene transfer. The presence of AAV vector dose-dependent activation of capsid-specific CD4+ and CD8+ T cells suggests that CTL responses against the capsid may have damaged or destroyed the transduced muscle fibers, similarly to the immune-mediated destruction of hepatocytes transduced by intravascular administration of AAV2/2 that occurred in the hemophilia B liver clinical trial (Manno et al., 2006). Therefore, an immunosuppresive regimen was started in LPL-deficient patients. Nevertheless, the overall results of the trial are encouraging and showed a reduction of circulating trygliceride levels and of episodes of pancreatitis (Gaudet et al., 2010). In summary, the multiple human clinical trials using AAV vectors by intramuscular injections have demonstrated excellent safety data, evidence of gene transfer, and in one case therapeutic efficacy.
In this study, we investigated the preclinical safety and efficacy of muscle-directed gene transfer mediated by AAV vectors for the therapy of Crigler-Najjar syndrome type 1.
Materials and Methods
Construction and production of AAV vectors
The rat UGT1A1 (rUGT1A1) coding sequence was obtained from Wistar rat liver mRNA and cloned into the pAAV-MCK-EGFP plasmid (Tessitore et al., 2008) by replacing the enhanced green fluorescent protein (EGFP) sequence. The cloned rUGT1A1 sequence was entirely verified by direct DNA sequencing.
The AAV vectors were produced and characterized by the Telethon Institute of Genetics and Medicine (Naples, Italy) AAV Vector Core. pAAV2.1-MCK-rUGT1A1 and pAAV2.1-MCK-EGFP were triple-transfected in subconfluent 293 cells along with pAd-Helper and pack2/1 packaging plasmids as described previously (Xiao et al., 1999). Recombinant AAV2.1-MCK-rUGT1A1 and AAV2.1-MCK-EGFP vectors were purified by two rounds of CsCl gradient centrifugation, as described previously (Xiao et al., 1999). Vector titers, expressed as genome copies per milliliter (GC/ml), were assessed by both PCR quantification (TaqMan; PerkinElmer, Life and Analytical Sciences, Waltham, MA) and dot-blot analysis.
Animal experiments
Animal procedures were performed in accordance with the regulations of the Italian Ministry of Health. Breeding pairs of Gunn rats were obtained from the Rat Resource and Research Center (RRRC, Columbia, MO) and a colony of Gunn rats was established at the Institute of Genetics and Biophysics-Telethon Institute of Genetics and Medicine (IGB-TIGEM) animal facility (Naples, Italy). For AAV vector or saline administrations, three intramuscular injections (three injections of 30 μl each) were performed in the gastrocnemius of 4- to 6-week-old male Gunn rats (75–150 g), for a total vector dose of 3×1012 genome copies (GC)/kg of AAV2.1-MCK-rUGT1A1 (n=9) or saline (n=5), using a 100-μl Hamilton syringe.
After vector injections, blood samples were collected by retro-orbital venipuncture. Bile was collected through a 26-gauge angiocatheter (Delta Med, Milan, Italy) inserted into the bile duct over 15-min periods, protected from light, frozen, and store at −80°C until analyses. Random urine spots were collected for measurement of alkali-labile bilirubin concentrations. Urinary creatinine was measured by colorimetric assay based on the Jaffe method (Clarke, 1961). Creatine phosphokinase (CPK) was measured in serum samples in the first week and 3, 15, 24, and 52 weeks postinjection (Gentaur, Milan, Italy).
To determine UGT1A1 protein and activity, muscle and liver tissues were collected 4 and 12 months postinjection. Tissues for real-time PCR were harvested from Gunn rats injected intramuscularly with AAV2.1-MCK-rUGT1A1 vector.
Bilirubin determinations
Blood was centrifuged at 1500×g for 20 min, and the serum was used for colorimetric measurement of total bilirubin by a diazo-based assay (Gentaur). The average serum bilirubin measured in 40 wild-type Wistar rats, 3–4 weeks of age, was 0.91±0.51 mg/dl.
Biliary, serum, and urinary unconjugated bilirubin and alkali-labile pigment concentrations were determined by high-performance liquid chromatography (HPLC) as previously described (Zelenka et al., 2008). Unconjugated bilirubin was measured before and after the addition of 1 M NaOH to the sample for 10 min, and the concentration of alkali-labile bilirubin pigments, expressed as bilirubin equivalents, was calculated from the difference between the two measurements. Qualitative analysis of bilirubin pigments in bile was performed by direct HPLC of undiluted bile by the McDonagh method (Toietta et al., 2005).
Western blot analysis, enzyme assay, and qPCR on tissues
Muscle and liver samples harvested 4 and 12 months postinjection were homogenized in 0.5 ml of phosphate-buffered saline (pH 7.4), using a TissueLyser homogenizer (Qiagen, Milan, Italy). The tissue homogenate was mixed with 4 ml of microsome buffer (2.62 mM KH2PO4, 1.38 mM K2HPO4, 2% glycerol, and 0.5 mM dithiothreitol) and first centrifuged at 12,000×g for 20 min at 4°C. The supernatant was then recentrifuged at 105,000×g for 60 min at 4°C. The pellet was resuspended in microsome buffer and the protein concentration was determined by the Bradford method.
Microsomal extracts were characterized by detection of a calnexin band by Western blotting using anti-calnexin antibody (Assay Designs, Ann Arbor, MI). These extracts were used to measure UGT1A1 activity. Approximately 10–20 μg of microsomal proteins from Wistar and Gunn rat liver and muscle was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and blotted onto PVDF membrane. Goat anti-rat UGT1A1 antiserum (diluted 1:1000; Santa Cruz Biotechnology, Santa Cruz, CA) with rabbit anti-goat IgG (diluted 1:3000; BioVision, Mountain View, CA) were used for immunodetection. Membranes were developed with an enhanced chemiluminescence kit (Thermo Scientific, Milan, Italy) and detected with ChemiDoc (Bio-Rad, Hercules, CA).
UGT1A1 enzyme activity in muscle microsomes was measured according to a previously published assay, using bilirubin as substrate (Heirwegh et al., 1972).
Total DNA was extracted from tissue samples, using phenol–chloroform extraction, and quantitated by absorbance at 260 nm. Quantitative real-time PCR was performed with LightCycler SYBR green master mix I (Roche, Indianapolis, IN) in a total volume of 20 μl with 100 ng of template DNA and a 1 μM concentration each of AAV-specific primers (5′-TCTAGTTGCCAGCCATCTGTTGT-3′ and 5′-TGGGAGTGGCACCTTCCA-3′). Cycling conditions consisted of 95°C for 10 min followed by 45 cycles at 95°C for 10 sec, 60°C for 7 sec, and 72°C for 20 sec. Serial dilutions of a plasmid bearing the PCR target sequence were used as a control to determine the amounts of AAV and results were analyzed with LightCycler 480 system (Roche).
GFP expression and RT-PCR after intramuscular injection of AAV
We harvested the following tissues from injected Gunn rats: muscles, liver, kidney, spleen, and heart. Tissues were fixed in 4% paraformaldehyde (PFA) for 24 hr. Muscle and liver specimens were embedded into paraffin blocks and sectioned into 10-μm serial sections, using a microtome, and GFP fluorescence was visualized with a Zeiss microscope.
Total RNA was extracted from liver, muscle, spleen, kidney, and heart in TRIzol reagent (Invitrogen, Monza, Italy), using an RNeasy kit (Qiagen, Italy). RNA was reverse transcribed, using a first-strand complementary deoxyribonucleic acid kit with random primers according to the manufacturer's protocol (Applied Biosystems, Monza, Italy). Primers for amplification of a 95-bp fragment of GFP were as follows: forward, 5′-ACGACGGCAACTACAAGACC-3′; and reverse, 5′-GTCCTCCTTGAAGTCGATGC-3′. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as loading control. A 136-bp segment of GAPDH was amplified with a forward primer (5′-ATGACTCTACCCACGGCAAG-3′) and a reverse primer (5′-TACTCAGCACCAGCATCACC-3′). Reaction conditions were as follows: Reverse transcription (RT) products were subjected to 39 cycles of amplification with 2.5 U of Taq DNA polymerase in 50 μl. After an initial denaturing cycle at 95°C for 7 min, subsequent cycles consisted of denaturation, 1 min at 95°C; annealing, 1 min at 60°C; and extension, 1 min at 72°C.
Statistical analysis
All data are expressed as means±SD. The statistical significance of differences between the means from two independent samples was tested by t test.
Results
Sustained reduction of hyperbilirubinemia and excretion of conjugated bilirubin in bile and urine from Gunn rats
Blood samples were collected 7 weeks after intramuscular injections of AAV2.1-MCK-rUGT1A1 vector at 3×1012 GC/kg, and monthly thereafter for measurement of total serum bilirubin levels. The vector dose was chosen on the basis of previous studies applying a similar dose in animal models and humans (Manno et al., 2003; Arruda et al., 2004; Toromanoff et al., 2008). Baseline levels of total serum bilirubin in Gunn rats injected with AAV vector or with saline were higher (7.95±1.8 mg/dl) than in wild-type Wistar rats (0.91±0.51 mg/dl). We observed an average 51% reduction of serum bilirubin in Gunn rats injected with AAV vector. This reduction was sustained for at least 51 weeks (Fig. 1). Reduction in total serum bilirubin levels was not observed in saline-injected rats (Fig. 1). The difference between total serum bilirubin in the AAV-injected group and the saline-injected group was statistically significant (p<0.05) throughout the period of study (Fig. 1).
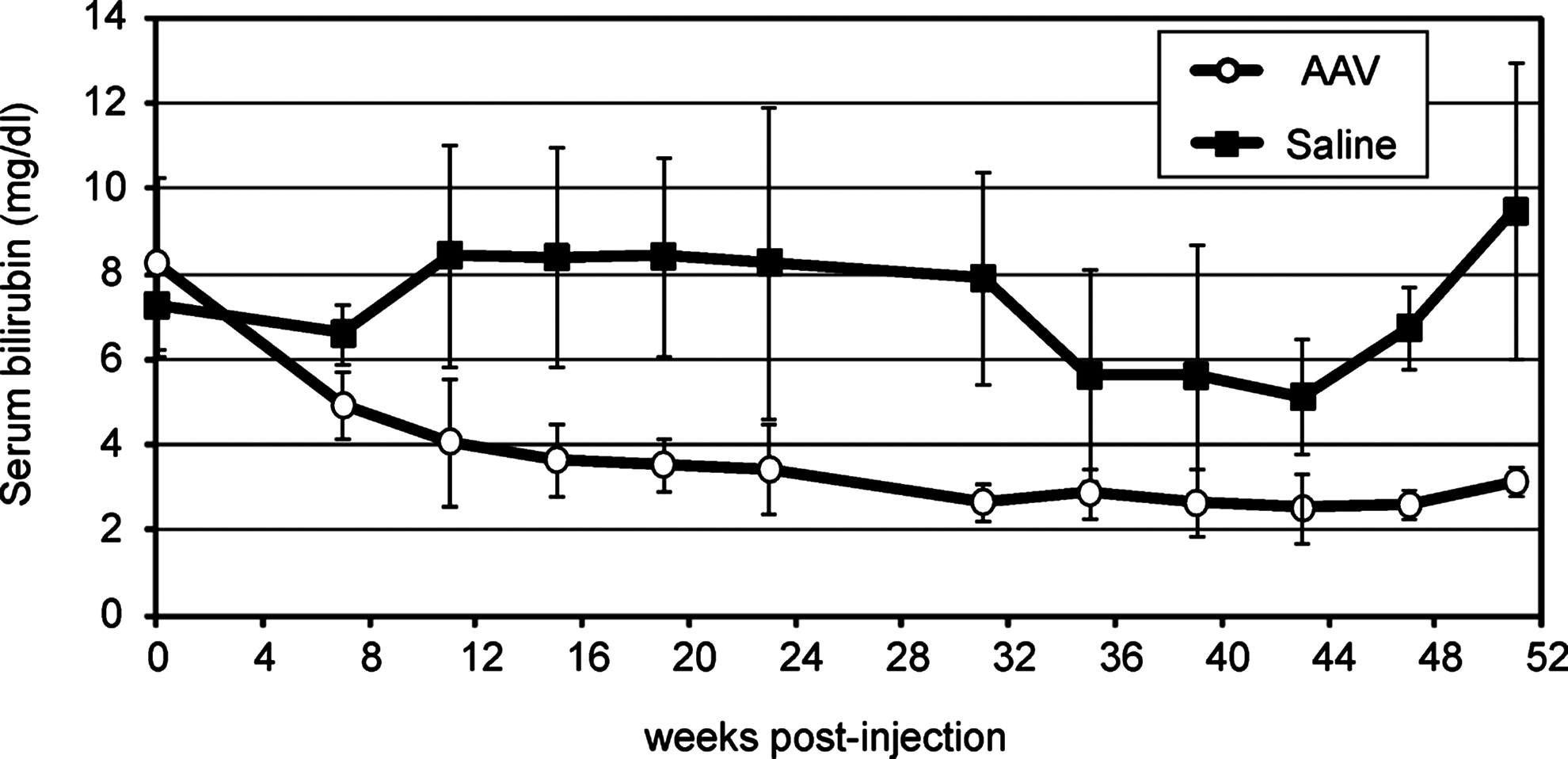
Intramuscular delivery of AAV results in sustained reduction of hyperbilirubinemia in Gunn rats. Serum bilirubin levels in Gunn rats after intramuscular injection of AAV2/1-MCK-rUGT1A1 (n=9) or saline (n=5). The normal range in wild-type rats (n=40) is 0.91±0.51 mg/dl. After AAV vector injection, long-term reduction of serum bilirubin in Gunn rats, lasting for at least 51 weeks, was observed. This reduction was on average approximately 51% of the saline-injected control levels (p<0.05).
To monitor muscle damage after intramuscular injection of the AAV vector, serum CPK levels were measured at various time points (24 and 48 hr, and 1, 3, 15, 24, and 52 weeks) after vector administration and found to be not increased as compared with saline-injected animals (data not shown).
To determine whether the reduction in serum bilirubin in vector-injected animals was due to enhanced biliary excretion of bilirubin metabolites, bile was collected by cannulation of the common bile duct from AAV- and saline-injected animals 51 weeks postinjection. Analysis by the method of Zelenka and colleagues (2008) showed an increase in alkali-labile bilirubin pigments in the bile of AAV-injected animals as compared with saline-treated controls (p<0.05) (Fig. 2A). Similar analysis showed that AAV-injected animals also had increased urinary excretion of bilirubin pigments compared with saline-injected controls (p<0.05) (Fig. 2B) and a reduction in serum unconjugated bilirubin (p<0.05) (Fig. 2C).
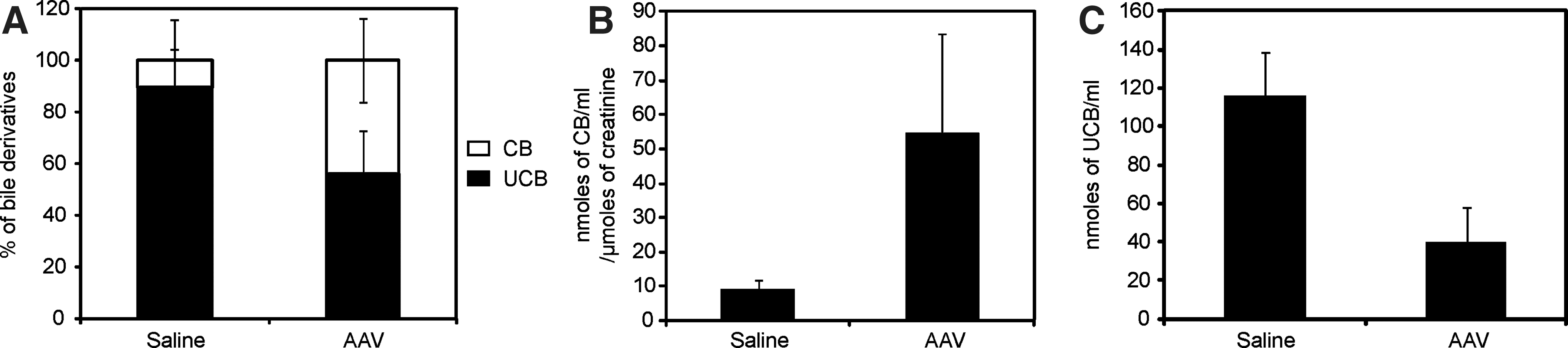
Bilirubin pigments in bile and urine samples. The amount of alkali-labile bilirubin pigments in the
Direct HPLC of bile samples showed only trace amounts, if any, of bilirubin glucuronides, but it revealed the presence of a polar metabolite peak eluting close to bilirubin diglucuronide (Fig. 3B). The absorbance spectrum of this metabolite was similar to, but still different from, that of bilirubin diglucuronide (Fig. 3A).
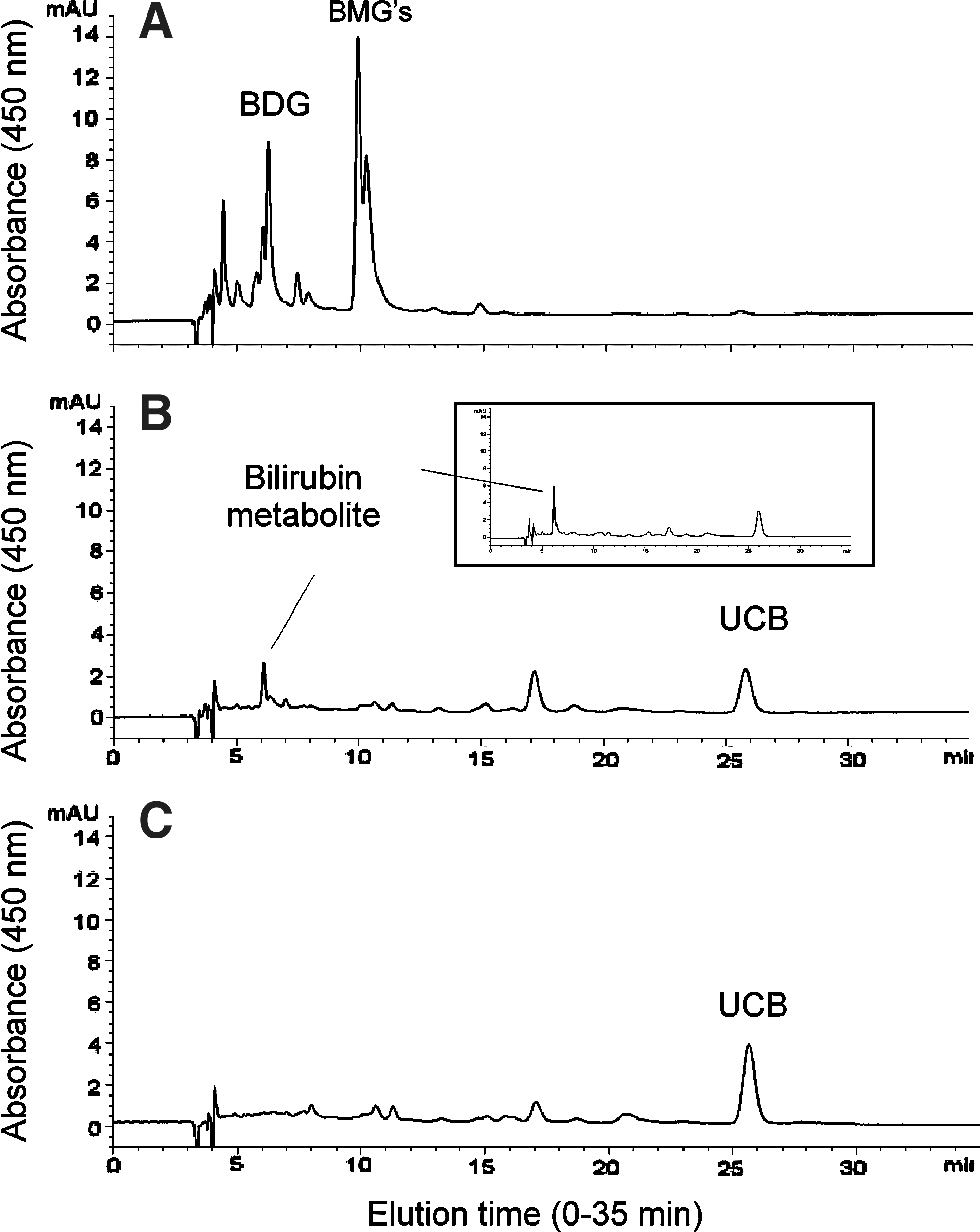
Bile HPLC: HPLC chromatograms of bile from wild-type rats, AAV-treated Gunn rats, and saline-injected control Gunn rats. Bile from
Expression of functionally active UGT1A1 protein in Gunn rat muscles
A subgroup of Gunn rats injected with either saline (n=3) or AAV (n=4) was sacrificed 4 months postinjection to harvest muscle microsomes for determination of enzyme activity and UGT1A1 Western blotting. As controls, microsomes were prepared from muscle and liver of wild-type Wistar rats. Western blot using an anti-rUGT1A1 antibody showed a band of 55 kDa, corresponding to rUGT1A1 in normal rat liver microsomes and in AAV-injected muscle microsomes but not in muscle microsomes from saline-injected controls (Fig. 4A). The presence of a band corresponding to calnexin confirmed that the purified muscle fractions corresponded to microsomes (Fig. 4A).
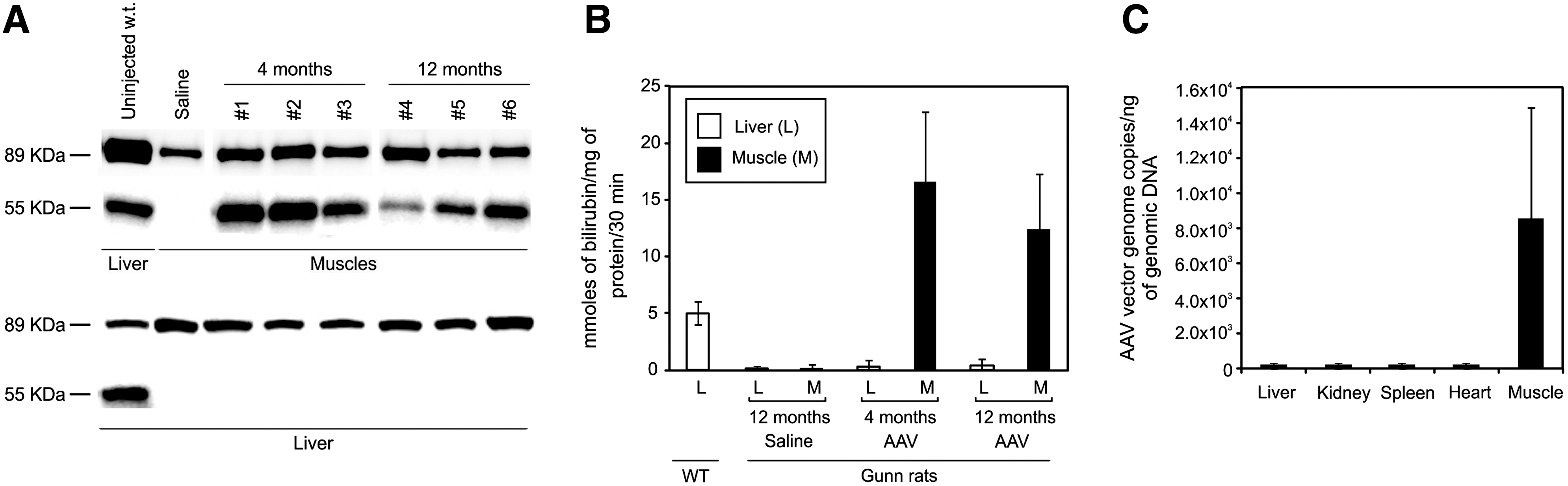
UGT1A1 expression in Gunn rat muscle.
In the same extracts used for Western blotting, UGT1A1 activity was measured and found to be increased in AAV-injected Gunn rat muscle microsomes relative to saline-injected muscles (p<0.05), whereas no UGT1A1 activity was detected in liver of the same animals (Fig. 4B). Taken together, these results indicate that AAV-transduced muscles can express long-term functional UGT1A1 protein.
Vector biodistribution in intramuscularly injected Gunn rats
Biodistribution of AAV2.1-MCK-rUGT1A1 vector by real-time PCR showed detectable vector genomes only in muscle, whereas the amounts of vector genomes detected in other organs (liver, kidney, spleen, and heart) were below the limit of detection (<1×102 copies of vector genome) (Fig. 4C).
To determine tissue specificity of transgene expression after intramuscular injection and possible hepatic expression from the MCK promoter, we injected Gunn rats (n=3) with AAV-MCK-GFP (3×1012 GC/kg), an AAV vector expressing the reported GFP protein under the control of the MCK promoter, and sacrificed the rats 1 month postinjection to determine tissue GFP expression and vector genome copy distribution. Injected muscles showed extensive GFP staining (Fig. 5A), whereas no staining was detected in liver (Fig. 5B). Expression of the GFP transgene driven by the MCK promoter was detected by RT-PCR in muscle, but not in liver or other tissues of the injected animals (Fig. 5C). Together with the absence of detectable hepatic UGT1A1 protein by Western blot analysis, this finding confirms that intramuscular injections of AAV2.1-MCK-rUGT1A1 result in muscle-specific expression (Fig. 5A).
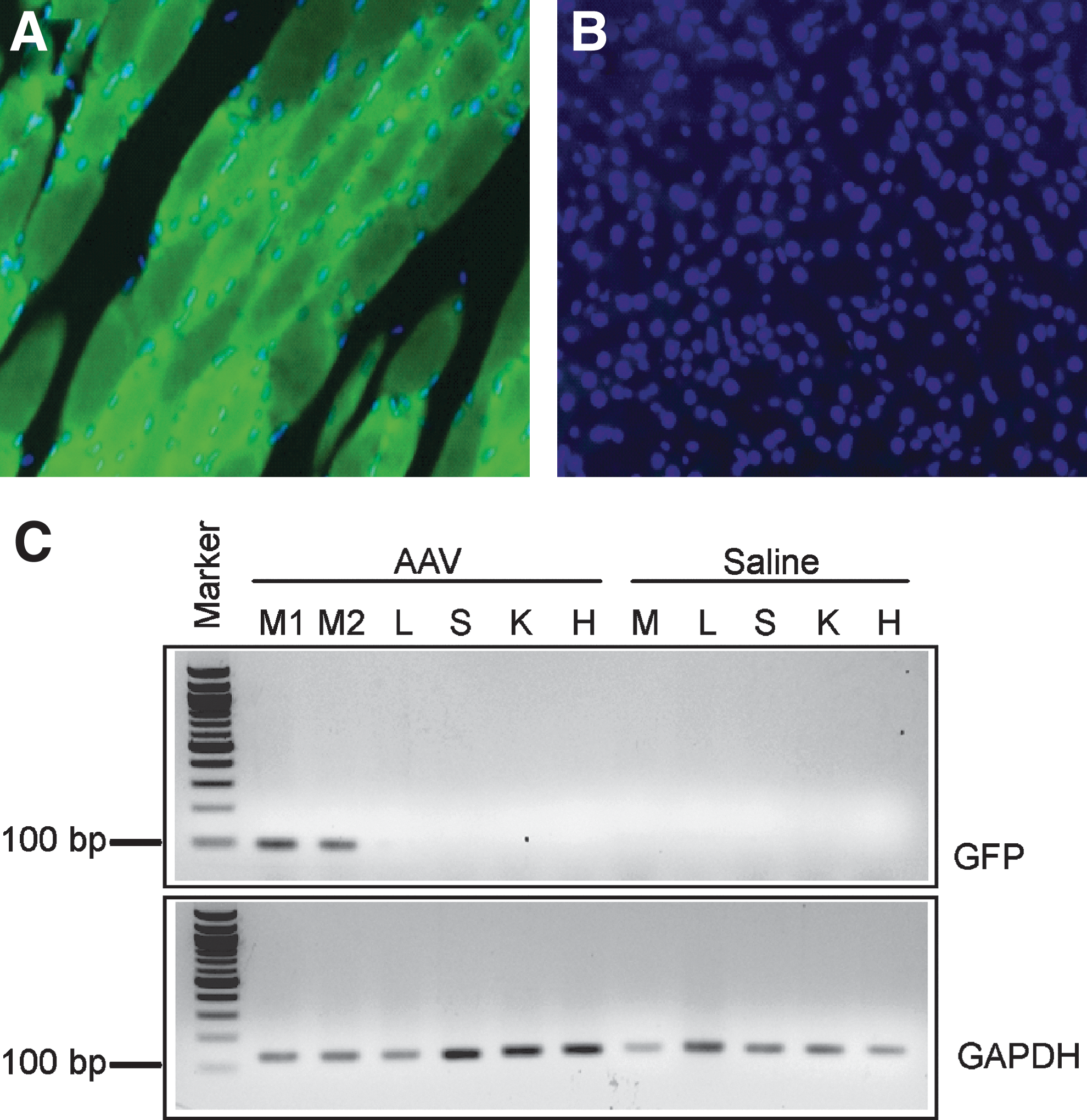
AAV vector muscle-specific transduction. GFP-positive cells were detected only in Gunn rat muscle
Discussion
Patients with Crigler-Najjar syndrome type 1 have life-threatening elevations of serum bilirubin and are currently managed with phototherapy throughout childhood and adolescence. Although effective, phototherapy is cumbersome and inconvenient, and its efficacy may diminish with age because of increased skin thickness and decreased surface-to-mass ratio (Strauss et al., 2006). Moreover, despite this treatment, patients remain at risk of brain damage when intercurrent infections may increase production of bilirubin above those levels that can be controlled by phototherapy (Strauss et al., 2006). Therefore, patients with Crigler-Najjar type 1 are often advised to consider liver transplantation, most frequently in the range of 18–25 years of age.
Muscle-directed gene therapy is attractive for Crigler-Najjar syndrome type 1 because skeletal muscle is easily and safely accessible by intramuscular injections and contains both dividing and nondividing cells with long half-lives resulting in stable episome expression (Jiang et al., 2006; Koo et al., 2011). For this strategy to be effective in treating hyperbilirubinemia in Crigler-Najjar syndrome, the enzyme produced in the ectopic site must be functional and expressed long term. In this study, we have shown that expression of the defective enzyme in muscle (Fig. 4) results in long-term reduction of hyperbilirubinemia (Fig. 1) without toxicity as demonstrated by serial CPK measurement (data not shown). This finding is supported by the Western blot (Fig. 4A) and in vitro UGT1A1 enzyme assay (Fig. 4B), showing that muscle expresses, long term, a functionally active UGT1A1 protein. Bile and urine of AAV-injected animals showed increased excretion of alkali-labile bilirubin derivatives (Fig. 2), thus suggesting that bilirubin esterified by AAV-transduced muscles is excreted in bile and urine. However, bilirubin mono- or diglucuronides were not detectable in bile by HPLC (Fig. 3); only an unidentified metabolite was detected (Fig. 3B). This metabolite is reminiscent of the metabolite observed by Seppen and colleagues, who showed correction of hyperbilirubinemia in Gunn rats receiving transplantation of cells that do not normally express UGT1A1 but were genetically modified ex vivo to express UGT1A1 (Seppen et al., 1997). It is possible that UGT1A1 expressed in muscle may result in a protein that is not identical to the physiological liver-expressed protein, as previously found in the case of erythropoietin expressed at ectopic sites (Lasne et al., 2004; Menzel et al., 2009). Whether posttranslational modifications of muscle-expressed UGT1A1 play a role in the formation of this unidentified metabolite is unclear at this time.
The negligible formation of bilirubin glucuronides in the present work despite the in vitro detection of UGT1A1 activity in muscle microsomes may reflect inadequate formation of the necessary uridine 5′-diphosphoglucuronic acid (UDPGA) cofactor (Wong, 1977). Coexpression of UDP-glucose dehydrogenase (UGDH), which generates UDPGA, may further increase the therapeutic efficacy of muscle-directed gene therapy, as in the case of muscle-directed gene therapy of phenylketonuria, which requires the expression of the complete phenylalanine hydroxylase (PAH) system (PAH and BH4-biosynthetic enzymes) to effectively clear phenylalanine from the blood (Ding et al., 2008). Nevertheless, UGDH catalyzes the conversion of UDP-glucose to UDP-glucuronic acid that is used for production of proteoglycans, which are involved in promoting normal cellular growth and migration (Auvinen et al., 2000; Wang et al., 2010). Therefore, overexpression of UGDH raises concern for risks of malignant transformation, which may impede clinical translation.
Consistent with our results, Bortolussi and colleagues have shown reduction of hyperbilirubinemia due to UGT1A1 muscle expression in Ugt1a1 –/– mice injected systemically with an AAV2/9 vector expressing human UGT1A1 under the control of the ubiquitous CMV promoter (Bortolussi et al., 2011). However, in those studies bilirubin glucuronide formation was demonstrated in vitro in muscle microsomes incubated with bilirubin and a large excess of UDPGA.
After vector intramuscular injection, AAV vector genome was detected primarily in muscle tissues with undetectable distribution to other tissues (Fig. 4C), consistent with the conclusion that reduction of serum bilirubin was due to muscle expression of functional UGT1A1.
The main goal of gene therapy for Crigler-Najjar syndrome is prevention of brain damage due to hyperbilirubinemia; complete normalization of serum bilirubin is not required to achieve this goal. In the present study, intramuscular delivery of the AAV vector did not result in complete normalization of serum bilirubin levels. On average, a 51% reduction of bilirubin levels was observed as compared with saline-injected control rats (Fig. 1). Such reduction is clinically highly relevant because a 30–60% decrease would result in serum bilirubin levels below 20 mg/dl in most patients (Strauss et al., 2006). Patients with Crigler-Najjar syndrome type 2, with levels below this threshold, are not at risk for brain damage (Arias et al., 1969). Therefore, such a reduction would result in important clinical benefit in patients with Crigler-Najjar syndrome type 1. Given the limited invasiveness of AAV-mediated muscle-directed gene therapy, this approach may be useful also for patients with Crigler-Najjar syndrome type 2, thus avoiding long-term treatment with phenobarbital.
Muscle-directed gene therapy based on pDNA delivery was previously investigated in Gunn rats (Jia and Danko, 2005a). In contrast with the pDNA study, we have demonstrated sustained correction and persistence of transgene expression after a single procedure of intramuscular injections. We also observed long-term correction of hyperbilirubinemia, whereas the study using pDNA detected immune elimination of transfected cells caused by the presence of anti-hUGT1A1 antibodies and lymphocytic inflammation (Jia and Danko, 2005a). Possible explanations for this difference include different vectors (AAV vs. pDNA), different routes of administration (intramuscular vs. limb perfusion), different promoters (MCK vs. CMV), different transgenes (rUGT1A1 vs. hUGT1A1), or a combination thereof.
A limitation of muscle-directed gene therapy is the small number of muscle fibers that are transduced after an intramuscular injection. However, efficient delivery methods based on limb perfusion for AAV vector delivery have been developed to allow distribution of the vector to a larger muscle mass (Arruda et al., 2010; Haurigot et al., 2010). This approach may potentially be applicable also for Crigler-Najjar syndrome type 1 to permit improved phenotypic correction by transduction of a larger number of muscle cells.
Peripheral vein infusion of self-complementary AAV2/8 vector encoding F.IX under the control of a liver-specific promoter resulted in F.IX transgene expression at levels sufficient to improve the bleeding phenotype in patients with hemophilia B (Nathwani et al., 2011). Although immune-mediated clearance of AAV-transduced hepatocytes remains a concern and was eventually controlled with a short course of glucocorticoids (Nathwani et al., 2011), this clinical trial may pave the way towards applications of liver-directed gene therapy for a wide range of inborn errors of liver metabolism, including Crigler-Najjar syndrome type 1. Nevertheless, AAV vectors are nonintegrating vectors and loss of transgene expression will occur during cell division in liver of growing patients. The onset of Crigler-Najjar syndrome type 1 is in the neonatal period and, thus, liver-directed gene therapy performed at that time will likely results in loss of therapeutic effect. In contrast, as shown by both small and large animal studies (Sabatino et al., 2007; Yue et al., 2008), intramuscular injections of AAV vectors in newborns result in long-term transgene expression in muscle, a tissue that can grow by cell fusion or by increase in protein content (Otto and Patel, 2010). Therefore, muscle remains attractive as a target organ for gene therapy for Crigler-Najjar syndrome type 1. Moreover, newborns may not mount a vigorous immune response because of their immature immune system and thus muscle-directed gene therapy in the neonatal period may be an option for patients carrying disease mutations (e.g., nonsense mutations), who are more prone to develop anti-UGT1A1 antibodies.
In summary, intramuscular injection of AAV resulted in reduction of serum bilirubin levels for at least 1 year. The reduction of serum bilirubin was associated with increased excretion of bilirubin species in bile and urine. On the basis of the HPLC spectrum, as previously shown with ectopic expression of UGT1A1 (Seppen et al., 1997), nonphysiological bilirubin species were observed in Gunn rats receiving intramuscular injections of AAV vector, but their identities remain to be determined. Nevertheless, intramuscular injections of AAV proved an effective, simple, and safe gene delivery method that maintained long-term functional UGT1A1 in Gunn rat muscle, and resulted in clinically relevant, long-term reduction of serum bilirubin levels.
Footnotes
Acknowledgments
The authors are grateful to Amit Nathwani for comments on the manuscript. The authors thank Renato Minopoli for assistance with animal procedures, and Antonella Ferrara and Monica Doria from the Telethon Institute of Genetics and Medicine Vector Core for AAV vector production. This work was supported by Fondazione Telethon (Rome, Italy) (TGM06C07) to N.B.P.
Author Disclosure Statement
The authors have no conflicts of interest to disclose.