
Abstract
Although clinical benefits have been reported in several human hematopoietic gene therapy trials, a remaining important goal is the transition to nonmyeloablative pretransplantation conditioning to decrease toxicity. Previous attempts at reduced intensity conditioning in nonhuman primates have resulted in only temporary vector marking of autologous blood cells or their persistence at low levels, well below the thresholds for clinical efficacy. In addition, we reasoned that lentiviral vector particles displaying cytokines at their surface have the potential to preserve stem cell fitness better than current ex vivo transduction protocols, which involve exposure to cytokine overstimulation. Here we show that the classically nonmyeloablative agent fludarabine (30 mg/m2/day for 3 days) together with low-level total body irradiation (2 Gy) and the use of a stem cell factor-displaying simian immunodeficiency virus-based vector, resulted in sustained, single-copy vector marking of autologous blood cells in two macaques over 3 years posttransplantation at levels averaging 1% of all lineages. This percentage is within the range of anticipated efficacy levels for hemophilia and related diseases and forms a basis for further improvement.
Introduction
Before hematopoietic gene therapy can be broadly available to patients, key obstacles must be addressed. It would, for instance, be highly desirable (1) to lower the doses in the preconditioning regimen in an effort to decrease their toxicity while maintaining clinical efficacy, and (2) to decrease HSC exposure to cytokines during the ex vivo transduction process to preserve the integrity of the HSC pool.
Although classical vesicular stomatitis virus glycoprotein G (VSV-G)-pseudotyped HIV-derived LVs can transduce nondividing cells, fully quiescent G0 cells are poorly transduced (Korin and Zack, 1998; Sutton et al., 1999; Frecha et al., 2008). Hence, human HSCs are poorly permissive to classical VSV-G-LV transduction because ∼75% of them are in G0 (Sutton et al., 1999). Only cytokine cocktails that induce cell cycle entry result in efficient HSC transduction (Horn et al., 2002; Hanawa, 2004; Verhoeyen et al., 2005; Trobridge et al., 2008), although there are concerns that intense exposure to cytokines may ultimately impair HSC function, homing, or viability (Verhoeyen et al., 2005). In an effort to achieve efficient HSC transduction with only mild and transient cytokine stimulation, we previously engineered LVs displaying “early-activating cytokine” at their surface (Verhoeyen et al., 2005). Although we demonstrated effective and selective transduction of mouse HSCs and human cord blood CD34+ cells, evidence that these cytokine-presenting LVs are effective at transducing HSCs capable of long-term repopulation in humans or nonhuman primates in transplant settings was lacking.
Nonhuman primate models have been invaluable for developing hematopoietic gene therapy strategies in humans because they closely mimic human biology. Only three published studies have reported results of RV or LV gene transfer to nonhuman primates after reduced intensity conditioning (RIC), with either low-dose intravenous busulfan (Kahl et al., 2006; Kang et al., 2006) or reduced-level total body irradiation (TBI) (Kang et al., 2001). These attempts have resulted in only temporary vector marking of autologous blood cells or their persistence at low levels, well below clinical efficacy thresholds (Kung et al., 2003; Zhang et al., 2010).
In the present study, we thus explored the possibility of transplanting macaques with autologous CD34+ cells transduced with an LV displaying “early-activating cytokines” together with a novel RIC regimen. Because HIV-based LV gene transfer into certain macaque and baboon cells is inefficient (Besnier et al., 2002; Horn et al., 2002; Verhoeyen et al., 2005; Trobridge et al., 2008; Hayakawa et al., 2009) due to an early postentry block in transduction caused by host restriction factors such as TRIM5α (tripartite motif 5α) (Besnier et al., 2002; Stoye, 2002), we transferred the early-acting-cytokine displaying HIV-LV approach to an SIVmac251-LV system, which we had developed previously (Negre et al., 2000; Derdouch et al., 2008). Although fludarabine is classically exploited for its dominant immunosuppressive property in allogeneic transplantation, fludarabine is also known to be a radiosensitizer (Plunkett et al., 1993; Gregoire et al., 1994, 1998) and to offer an adjunct myelosuppressive effect in the treatment of hematological malignancies based on purine analog chemotherapy (Grever et al., 1990; Von Hoff, 1990; Keating et al., 1994). We thus decided to evaluate a regimen that combines fludarabine with 2-Gy TBI, a level of irradiation inadequate to achieve, in and of itself, autologous engraftment in the macaque (Kang et al., 2001).
Here we show that the association of this RIC regimen with a stem cell factor (SCF)-displaying SIV vector was well tolerated and resulted in a level of sustained (over 3 years), pan-lineage hematopoietic vector marking that is substantially higher than in previous studies (Kang et al., 2001, 2006; Kahl et al., 2006). This level of chimerism remains modest but is now within the range of anticipated efficacy for hemophilia-related diseases and forms a platform for further improvement.
Materials and Methods
Envelope construction
The TPO171HA chimeric envelope glycoprotein (TPOHA) has been previously described (Verhoeyen et al., 2005). The SfiI–NotI fragment from SCFSU (Verhoeyen et al., 2005) was fused to the hemagglutinin (HA) env gene, using the SfiI–NotI backbone fragment of TPOHA, resulting in the SCFHA glycoprotein (GP). All chimeric Env GPs were expressed in the phCMV-G expression vector backbone (Maurice, 2002). The plasmid SIV4+ encodes Gag/Pol, Rev, and Tat and is devoid of accessory proteins (Mangeot et al., 2000), and the self-inactivating SIV (SIN-SIV) vectors (SIN-SIV-EF1α-eGFP, SIN-SIV-SFFV-eGFP, SIN-SIV-EF1α-eGFP; Fig. 1a) are derived from the vector GAE-SIV (Mangeot, 2002).
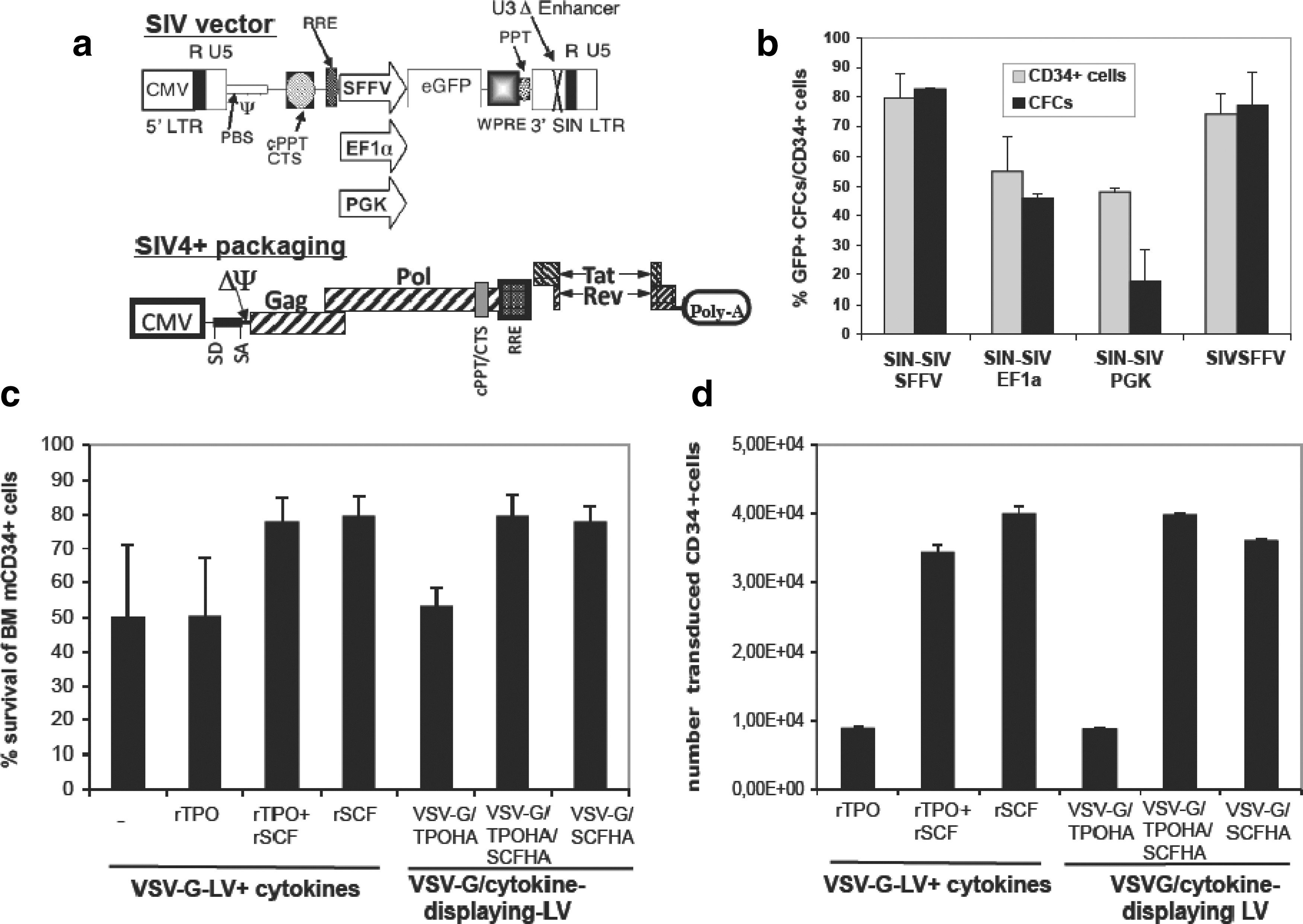
Stem cell factor (SCF) surface-engineered SIV vectors induce high levels of gene transfer in macaque bone marrow (BM) CD34+ cells.
Production of retroviral vectors
Self-inactivating SIV-derived vectors were generated as previously described by transient calcium phosphate-based transfection of 293T cells. Briefly, for codisplay of VSV-G with SCFHA and TPOHA, 2 μg of each envelope plasmid was cotransfected with the Gag/Pol packaging construct SIV4+ (8.6 μg) and an enhanced green fluorescent protein (eGFP)-encoding SIN-SIV lentiviral vector (8.6 μg). At 16 hr posttransfection, the Dulbecco's modified Eagle's medium (DMEM) was replaced by serum-free Cellgro medium (CellGenix, Freiburg im Breisgau, Germany) and at 36 hr after transfection, vectors were harvested, filtrated through a 0.45-μm pore-sized membrane, and concentrated with a filter concentration system (Sartorius, Aubagne, France).
Determination of activity of cytokine-displaying vectors
The functional activity of each preparation of SCFHA-displaying vector was evaluated on Ba/F3-c-Kit cells expressing the SCF receptor c-Kit, whereas Ba/F3-Mpl cells, expressing the thrombopoietin (TPO) receptor c-Mpl, were used to evaluate the activity of TPOHA-displaying vectors as described in Verhoeyen and colleagues (2005).
Animals
Adult male cynomolgus macaques (Macaca fascicularis) weighing 3 to 4.5 kg were imported from Mauritius and housed in single cages within level 2 biosafety facilities, in accordance with national guidelines. All experimental procedures were conducted in accordance with the European guidelines for animal care (Journal Officiel des Communautés Européennes, L358, 18 Décembre 1986—animal facility agreement no. A 92 032 02). Studies were conducted under protocols approved by the regional committee for animal experimentation (05025; January 7, 2006). All procedures and blood samplings were performed after the animals had been anesthetized with ketamine (10 mg/kg) (Imalgène 1000; Rhône Mérieux, Toulouse, France).
Isolation of macaque bone marrow CD34+ cells
Bone marrow mononuclear cells were obtained from the humeri and iliac crest by aspiration and were isolated by standard Ficoll density-gradient centrifugation. Cells were washed twice in phosphate-buffered saline (PBS) and were resuspended in PBS–1% fetal calf serum (FCS). The fraction was then enriched for CD34+ cells by positive immunomagnetic selection (clone 561; Dynabeads M-450 CD34; Dynal, Compiègne, France). After immunoselection, the CD34+ cell-purified fraction was stained with an anti-CD34 monoclonal antibody (clone 563; BD Pharmingen, Le Pont de Claix, France). The purity of the CD34+ selected cells ranged from 90 to 95%.
Transduction assays
To determine the infectious titers of SIV-derived vectors, serial dilutions of vector preparations were added to 293T cells. Multiplicities of infection (MOIs) were determined on proliferating 293T cells and are indicated in all transduction experiments. For comparison of the transduction efficiency in CD34+ cells with various SIV vectors (SIN-SIV-PGK-eGFP, SIN-SIV-SFFV-eGFP, SIN-SIV-EF1α-eGFP; Fig. 1a), bone marrow (BM) macaque CD34+ cells (mCD34+ cells) were prestimulated for 24 hr with TPO (20 ng/ml), SCF (200 ng/ml), and Flt-3 (200 ng/ml). BM mCD34+ cells (5×104) were transduced in 48-well plates with concentrated LV supernatant at the indicated MOIs in Cellgro serum-free medium (CellGenix) in the presence of the same cytokine cocktail. For the cytokine-displaying vectors, 5×104 BM mCD34+ cells were transduced in 48-well plates with concentrated LVs at the indicated MOIs in Cellgro serum-free medium without prestimulation or addition of exogenous cytokines. For control transductions with VSV-G-LVs, the cytokines were added at the time of transduction. Transduction efficiencies were determined on days 3 and 6 posttransduction.
Colony-forming cell essay
The colony-forming cell (CFC) assay was performed as described previously (Derdouch et al., 2008).
Conditioning protocol and follow-up
Before conditioning, several blood samples were taken to establish a baseline of the macaques' blood profile. As outlined in Fig. 2, the macaques were injected with fludarabine by intravenous injection (Fludara, 30 mg/m2; Schering-Plough, Paris, France) into the saphenous vein on three consecutive days (days −3 to −1). On the last day of fludarabine injection, the macaques were uniformly exposed in a field of 60Co γ rays with an anterior unilateral direction. A total midline tissue dose of 2 Gy was applied (0.250 Gy/min). At the indicated time points the weight of the macaques was determined; blood samples were taken to measure blood cell counts and hemoglobin; and a phenotypic follow-up of various myeloid blood cell lineages (erythrocytes, platelets, granulocytes, and monocytes) and lymphoid lineages (T and B cells) was performed by fluorescence-activated cell-sorting (FACS) analysis. At the indicated time points, BM biopsies were taken to evaluate the BM compartment for the presence of CFCs. For the CFC essay, 2×105 total BM cells were seeded in methylcellulose (H4434; StemCell Technologies, Grenoble, France) culture sustaining myeloid differentiation.
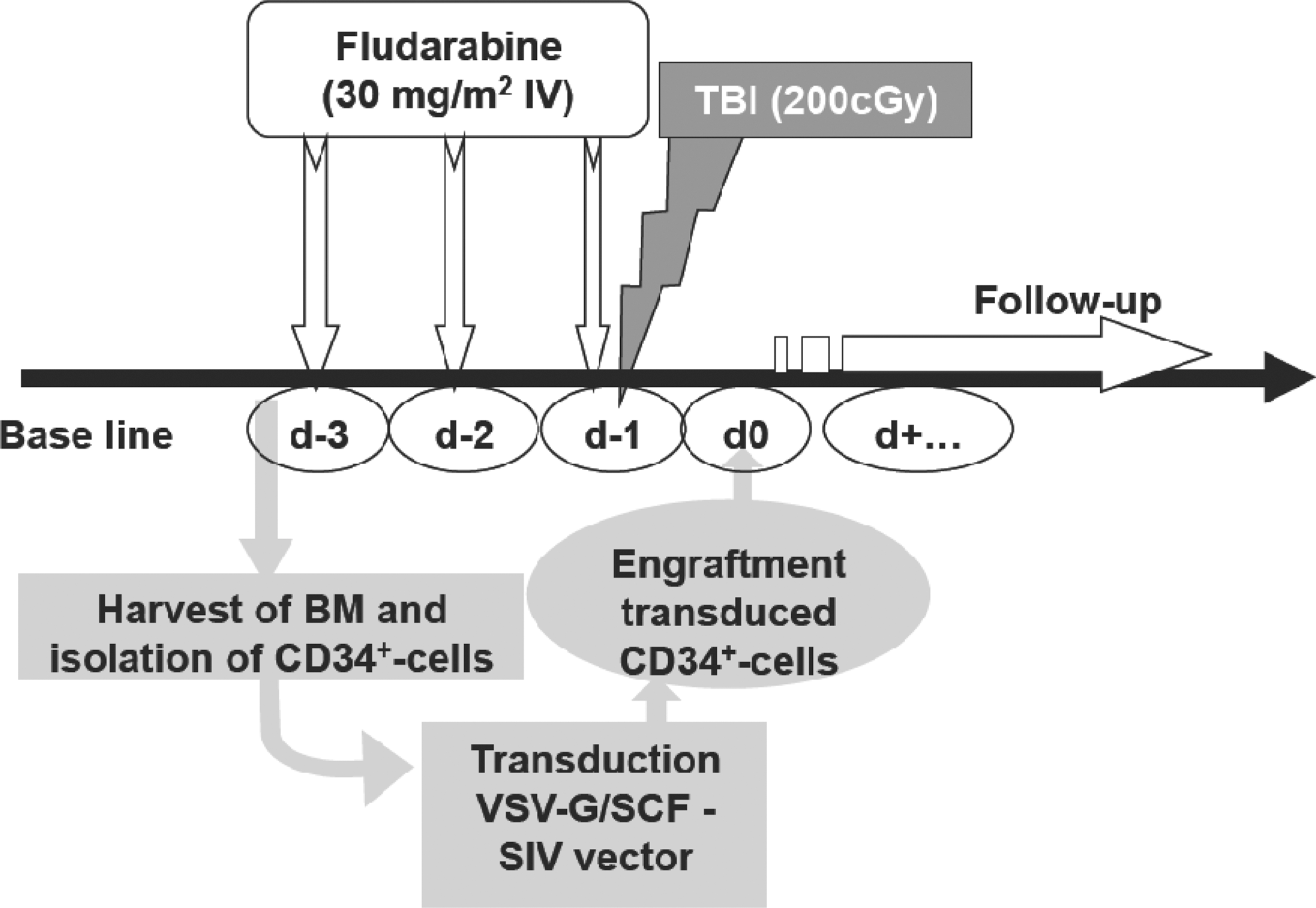
Layout of the reduced intensity conditioning (RIC) protocol and autologous transplantation. One month before enrolling the animals in the protocol, a baseline was established for the cell counts of all blood lineages. RIC conditioning consists of three intravenous injections of fludarabine (30 mg/m2) per animal per day (day −3, day −2, and day −1). On day −1 after fludarabine injection the macaques were subjected to total body irradiation (TBI). For the animals receiving autologous engraftment, BM was harvested on day −4 and mCD34+ cells were immediately isolated and processed for transduction. Meanwhile, these animals received the RIC protocol as described previously. One day after conditioning was finished, the transduced mCD34+ cells were injected intravenously. A detailed follow-up consisted of (1) weighing the animals, (2) measurement of hemoglobin concentration, (3) determination of blood cell counts in the various lineages, (4) detection of eGFP-expressing cells in various lineages of the peripheral blood, and (5) less frequent biopsies of the BM to determine eGFP+ cells and BM CFC capacity.
Clinical support
All animals received clinical support in the form of antibiotics and freshly irradiated whole blood and/or platelet-rich plasma (PRP), as required. A prophylactic antibiotic regimen was initiated when the leukocyte count fell below 1000/μl and continued daily until it exceeded 1000/μl for three consecutive days: gentamicin (Bi-Gental, 1 ml/10 kg/day; Schering-Plough Santé Animale, Paris, France) and oxytetracycline (Terramycin, 1 ml/10 kg; Pfizer, Berlin, Germany). Fresh, irradiated (25 Gy; 137Cs γ radiation) whole blood from a random donor pool was administered if the hemoglobin concentration was less than 8 g/dl and PRP was infused when the platelet count fell below 10,000/μl.
Macaque BM cell harvesting and autologous transplantation
BM was aspirated from the left and right femurs of macaques, using a “trocart de Mallarmé” (1.5 mm/17 gauge; length, 20 mm; Thiebaud Biomedical Devices, Thonon-les-Bains, France), 1 day before starting conditioning. mCD34+ cells were isolated from this BM directly and the fresh cells were transduced immediately with the SCF/VSV-G-LVs as described previously. The macaques (nos. 20672 and 20703) that underwent autologous transplantation were subjected to the same conditioning protocol as those without autologous transplantation (nos. 13586 and 13696). Transduced autologous cells were reinfused (saphenous vein) 20 hr after the last injection of fludarabine (Fig. 2).
Quantitative PCR
DNA extracts of mononuclear cells derived from peripheral blood (PB) or BM were obtained with a High Pure DNA purification kit (Boehringer-Mannheim, Mannheim, Germany) according to the manufacturer's instructions. Samples were quantified by the optical density method (SpectraMax 190; Molecular Devices, Sunnyvale, CA), controlled and normalized using primers specific to the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) housekeeping gene (5′ GAPDH F, 5′-GTATCGTGGAAGGACTCATGACC-3′; 3′ GAPDH R, 5′-GTTCAGCTCAGGGATGACCTTG-3′; and TaqMan probe, 5′-AGTCCATGCCATCACTGCCACCCA-3′). Both GAPDH and SIV vector sequences were amplified by PCR on a CFX96 real-time thermocycler (Bio-Rad, Hercules, CA) performed on 250 ng of DNA with Platinum quantitative PCR SuperMix-UDG (Life Technologies, Cergy-Pontoise, France). The primers used allow the specific amplification of a 92-bp fragment of the SIV vectors (5′SIV 2061 F, 5′-TACGGCTGAGTGAAGGCAGTAAG-3′; and 3′SIV 2134 R, 5′-CTCCTCACGCCGTCTGGTA-3′). The TaqMan probe sequence was 5′-AGGAACCAACCACGACGGAGTGCTC-3′. Samples were run in duplicate, and the values reported correspond to the means for replicate wells.
Flow cytometric analysis
eGFP expression was monitored by FACS in PB and BM mononuclear cells. Samples were incubated for 30 min at 4°C with monoclonal antibodies for single- or triple-color staining. The following antibodies were used: allophycocyanin (APC)–cyanine 7 (Cy7)-conjugated anti-CD3 (clone SP34-2; BD Pharmingen), phycoerythrin (PE)–Cy7-conjugated anti-CD8 (clone RPA-T8; BD Pharmingen) and peridinin chlorophyll protein (PerCP)–Cy5.5-conjugated anti-CD4 (clone L200; BD Pharmingen), PerCP-conjugated anti-CD20 (clone B9E9; Immunotech, Marseille, France), APC–Cy7-conjugated anti-CD11b (clone ICRF44; BD Pharmingen), Alexa Fluor 647-conjugated anti-CD16 (clone 3G8; BD Pharmingen), PerCP–Cy5.5-conjugated anti-CD14 (clone M5E2; BD Pharmingen), APC-conjugated anti-CD41a (clone HIP8; BD Pharmingen), and PE-conjugated anti-CD34 (clone 563; BD Pharmingen). For all analyses, isotype control antibodies were used in parallel. Cells were sorted with a FACSAria I and analyzed with a BD FACSCanto II flow cytometry apparatus equipped with FACSDiva software (Becton Dickinson). Negative controls from normal macaques were run with every experimental sample and were used to establish gates for eGFP quantification.
Results
Efficient transduction of macaque bone marrow CD34+ cells with a further optimized SIV lentiviral vector
HIV-based lentiviral gene transfer into cynomolgus macaque cells is extremely inefficient because of the presence of restriction factors in simian cells, causing an intracellular block (Horn et al., 2002; Stoye, 2002; Trobridge et al., 2008). Because SIV-derived vectors are not subjected to this restriction (Sandrin, 2002; Hanawa, 2004; Swainson et al., 2006; Derdouch et al., 2008), we developed a series of latest-generation SIN-SIV vectors (Fig. 1a), carrying the woodchuck posttranscriptional regulatory element (WPRE). These vectors were produced by transient transfection of the SIV vector, the Gag/Pol-encoding construct (SIV4+ in Fig. 1a [Mangeot et al., 2000]), and a VSV-G-encoding plasmid into 293T cells.
To identify the best performing SIV vector, mCD34+ cells were prestimulated for 24 hr with a cocktail of cytokines containing recombinant TPO (rTPO), rSCF, recombinant Flt3 ligand (rFlt3L), and recombinant interleukin-3 (rIL-3). Cells were subsequently transduced with three different SIN-SIV vectors carrying the internal promoter for spleen focus-forming virus (SFFV), elongation factor-1α (EF-1α), or phosphoglycerate kinase (PGK) (Fig. 1a) and an SIV-LV without deletion in the U3 region carrying the SFFV internal promoter. The SIV-SFFV-eGFP vectors were markedly superior to the two others for mCD34+ cell transduction and easily reached an 80–90% transduction level after a single vector hit (Fig. 1b).
To exclude that pseudotransduction of eGFP protein contributed to this high transduction, transduced mCD34+ cells were induced to myeloid differentiation in methylcellulose and eGFP+ CFCs were scored on day 14. SIV-LVs carrying the EF-1α or PGK promoter tended to have lower percentages of eGFP+ CFCs compared with the mCD34+ cells from which they were derived. For both SIV-SFFV-eGFP vectors with (SIN) and without U3 enhancer deletion, the percentage of eGFP+ CFCs was equivalent to that of the eGFP+mCD34+ cells. We therefore considered that the SIN-SIV-SFFV-GFP vector was the best and safest candidate to be used in further investigations.
SCF-displaying SIV vectors are highly efficient for the transduction of macaque CD34+ cells
We previously developed VSV-G/thrombopoietin (TPO)- and VSV-G/stem cell factor (SCF)-codisplaying HIV vectors), which outperformed conventional VSV-G-HIV-LVs for ex vivo gene delivery into the most immature human HSCs (Verhoeyen et al., 2005). Because of the restrictions of HIV vectors in macaque cells mentioned previously, we transferred this cytokine-displaying LV technology from HIV to SIV vectors. TPO was fused to the N terminus of the influenza HA GP (named TPOHA), as was SCF (named SCFHA), because this resulted in efficient coincorporation with VSV-G on SIV-LVs (Supplementary Fig. S1b; supplementary data are available online at
To evaluate a potential synergistic effect between SCF and TPO, we produced TPO/SCF-codisplaying vectors using the combination of VSV-G/TPOHA/SCFHA GPs. Clearly, the TPO/SCF-displaying vectors maintained survival of BAF3-Mpl and BAF3-c-Kit cells to the same extent as their corresponding single cytokine-displaying vectors. As expected, control incubations with VSV-G-displaying vectors in the absence of cytokines resulted in rapid cell death of both BaF3 cell lines (Supplementary Fig. S1c and d).
We then evaluated specific ligand–receptor interactions on the primary targets, the immature macaque CD34+ bone marrow (BM) progenitor cells. Of utmost importance was that SCF- or TPO/SCF-displaying LVs protected mCD34+ cells against apoptosis and to the same extent as that observed after incubation with high levels of rSCF (Fig. 1c). In contrast, TPO-displaying vectors were unable to maintain BM mCD34+ cell survival (the same was observed for rTPO incubation), although this vector showed functional display of TPO (Fig. 1c and Supplementary Fig. S1c). This is most likely due to a reduced effect of TPO on human and macaque BM CD34+ cells, as we have previously observed that TPO-displaying vectors efficiently sustained survival of human cord blood (CB) CD34+ cells (Verhoeyen et al., 2005). Indeed, TPO-induced survival and expansion are much lower for human BM CD34+ cells compared with human CB CD34+ cells (Liu et al., 1999; van den Oudenrijn et al., 2000) (and data not shown). This is consistent with the limited mCD34+ cell expansion after 3 days of incubation with TPOHA-LVs or rTPO versus SCFHA-LVs or rSCF, which allowed a 3-fold expansion (Supplementary Fig. S2a).
A single exposure to TPO-, SCF-, or TPO/SCF-displaying vectors in the absence of serum or other stimuli promoted high transduction levels in mCD34+ cells (Supplementary Fig. S2b). Thus, SCF- or TPO/SCF-displaying vectors not only improved survival of mCD34+ cells and allowed slight expansion but they also promoted high transduction levels in these cells, in a manner similar to VSV-G-pseudotyped SIV-LVs in the presence of recombinant cytokines (Fig. 1c and Supplementary Fig. S2a and b).
We then analyzed the absolute numbers of transduced mCD34+ cells (defined as initial number of cells×transduction [%]×cell survival [%]×expansion; Fig. 1d) after incubation with the LVs at a multiplicity of infection of 20. TPO-, SCF-, or TPO/SCF-displaying vectors transduced equivalent numbers of mCD34+ cells as unmodified LVs in the presence of their counterpart recombinant cytokines, rTPO and/or rSCF (Fig. 1d). The significantly lower number of transduced cells obtained with VSV-G/TPO-LVs was due to a combination of lower cell survival and lack of expansion of mCD34+ cells for TPO-displaying LVs (Fig. 1c and Supplementary Fig. S2a), whereas VSV-G/SCF-LVs induced high cell survival and modest expansion. Indeed, alternative transduction protocols using extensive stimulation by cytokines (rTPO, rSCF, rFlk-3, and rIL-3) led to more than 10-fold expansion (data not shown). In contrast SCF- or TPO/SCF-displaying LVs resulted in only 2.5- to 3.5-fold expansions, which was similar to incubations with low-dose rTPO and/or rSCF (Supplementary Fig. S2a).
We hypothesized that the VSV-G/SCF-displaying vector, due to its mild stimulation of mCD34+ cells by SCF, would result in a lower number of integrated copies per cell as compared with VSV-G-LV transduction of mCD34+ cells, prestimulated with a strong cytokine cocktail (rTPO, rSCF, rFlk-3, and rIL-3). Indeed, we found for VSV-G/SCF-LVs a lower integrated copy number than in the latter condition (VSV-G/SCF-LV: 1.40±0.49 copies/cell vs. VSV-G-LV+TPO/SCF/FLk-3/IL-3: 2.40±1.11 copies/cell, respectively; p=0.042; MOI=50; data not shown). This result implies that genotoxicity may be reduced by the use of VSV-G/SCF-LVs for HSC transduction.
We thus concluded that the SCF-displaying SIV-LV was far superior to the TPO-displaying SIV-LV, and it was therefore the candidate of choice for ex vivo transduction of macaque CD34+ cells destined for autologous engraftment.
Novel nonmyeloablative conditioning of macaques results in efficient white blood cell depletion, although with low toxicity
Most conditioning protocols for autologous engraftment with RV- or LV-transduced hematopoietic cells in macaques and baboons use a myeloablative conditioning protocol based on TBI (Hanawa, 2004; Derdouch et al., 2008; Trobridge et al., 2008; Kim et al., 2009; Stoddart et al., 2009). Only three published studies attempted to apply milder conditioning regimens that included 2-Gy TBI alone (Kang et al., 2001) or low-dose intravenous busulfan (Kahl et al., 2006; Kang et al., 2006), with a maximal level of chimerism for vector-bearing cells less than 0.1% 6 months posttransplantation (PTx). We thus explored the possibility of complementing 2-Gy TBI with the mildly myeloablative radiosensitizer fludarabine (Plunkett et al., 1993; Gregoire et al., 1994, 1998), an intentional use thus distinct from its classical administration as an immunosuppressive agent in allogeneic transplantation. This RIC protocol, hereafter referred to as XF-RIC, has been extensively applied in human patients for the purpose of allogeneic hematopoietic stem cell transplantation, with substantial engraftment success. However, the ability of this regimen to induce sufficient myelosuppression for long-term persistence of an autologous engraftment has not been documented. CD34+ BM progenitor cells are normally less sensitive to fludarabine compared with T and B cells because the drug works efficiently on cycling cells by inhibition of DNA/RNA synthesis (Goodman et al., 1996). Nevertheless, phase I/II clinical trials (Grever et al., 1990; Von Hoff, 1990) and reports of hematological malignancies (Keating et al., 1994) have observed that myelosuppression is a severe dose-limiting toxic effect of fludarabine. In addition, fludarabine appears to be a radiation sensitizer with synergistic activity (Plunkett et al., 1993; Gregoire et al., 1994, 1998).
To evaluate the degree of myelosuppression and its tolerance, we first subjected two macaques (nos. 13586 and 13696; Table 1), in the absence of gene transfer, to the XF-RIC regimen that consisted of fludarabine injections (30 mg/m2/day) for three consecutive days together with the addition of 2-Gy TBI on day 3 of fludarabine (Fig. 2). Several BM biopsies after XF-RIC confirmed transitory myelosuppression with undetectable mCD34+ cell clonogenic capacity (CFCs) in both animals from days 1 to 34 after conditioning (Fig. 3). BM CFCs were detected again on day 42 after RIC (Fig. 3).
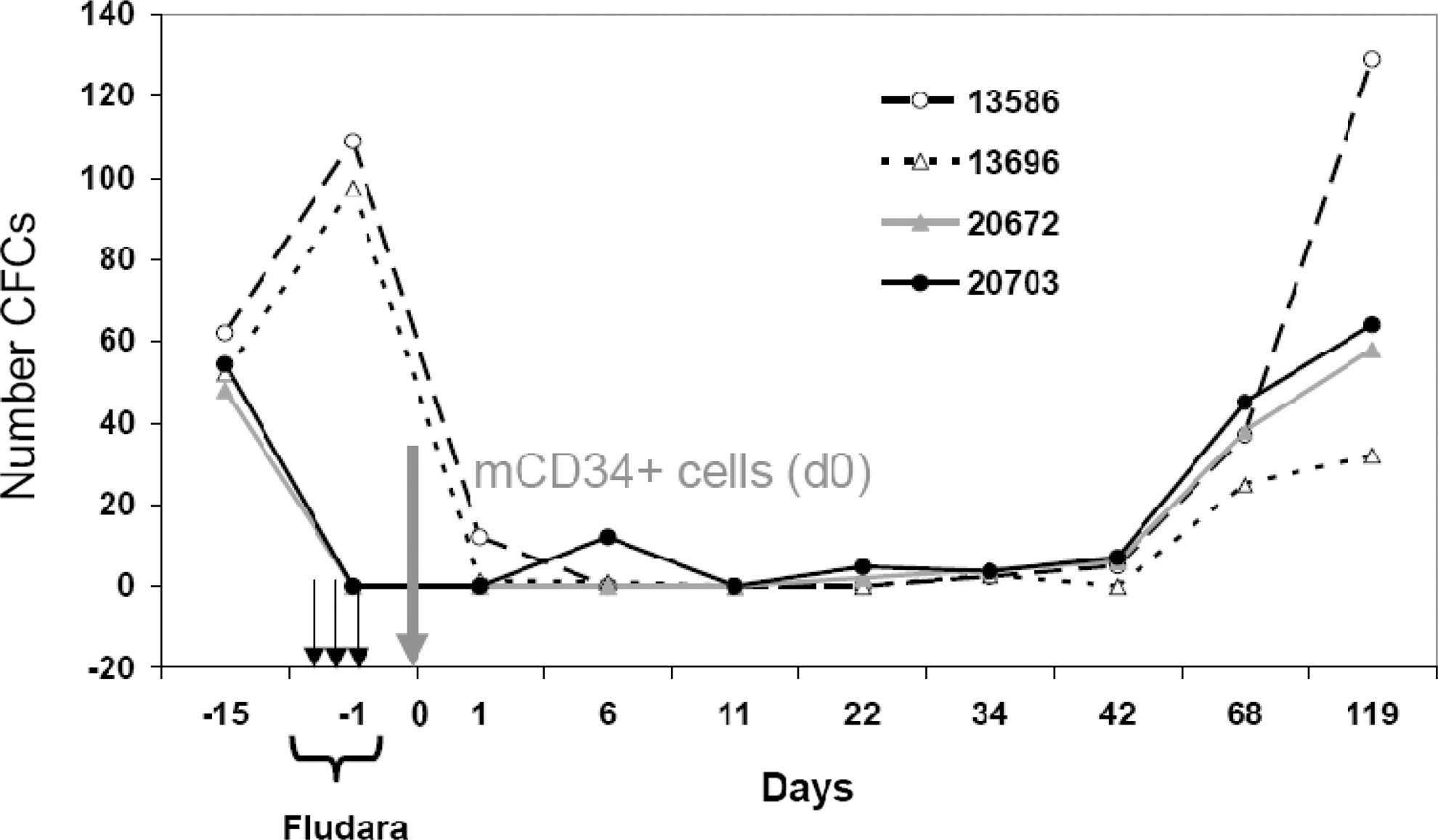
Follow-up of BM myeloid colony-forming cell (CFC) capacity in animals subjected to conditioning alone or combined with autologous engraftment. Four animals (13586, 13696, 20703, and 20672) were subjected to the conditioning protocol outlined in Fig. 2. Animals 20703 and 20672 received an autologous graft of SIV-LV-transduced mCD34+ cells. Starting from day −1, BM biopsies were further taken on days 1, 6, 11, 22, 34, 42, 68, and 119 after injection of transduced autologous mCD34+ cells. Total BM cells (2×105 cells) were seeded in methylcellulose supplemented with a cytokine cocktail to induce myeloid differentiation into erythrocytes, granulocytes, and dendritic cells. All assays were performed in triplicate. The total number of colonies (CFCs) was scored on day 14 of culture.
eGFP, enhanced green fluorescent protein; MOI, multiplicity of infection.
Overall, XF-RIC was well tolerated with no mucositis or other severe toxicities and no significant weight loss (Fig. 4a). As frequently observed in patients, macaques suffered from diarrhea during the fludarabine treatment, which stopped 1 day after the last injection. One month before starting this protocol, a baseline was established for the various blood lineages. During the first 50 days of follow-up, blood samples were taken every 3–4 days. The XF-RIC regimen induced severe leukopenia in these animals with a significant decrease in white blood cell counts on day 5 after the last fludarabine injection (Fig. 4c). Overall recovery was detected 40 days after XF-RIC (Fig. 4c). As expected, more detailed analysis showed that monocytes were less affected than lymphocytes (Fig. 5c), which showed a significant decrease over at least 3 weeks (days 4–32 in Fig. 5d and e). T cells were slightly more resistant than B cells, whereas in the myeloid compartment granulocytes and platelets were most affected by the treatment (Fig. 5b–e). A significant drop in platelet count was observed in both animals from day 10 onward. Severe thrombocytopenia (platelet counts <20,000/μl) was detected in one of the two macaques (no. 13696; days 15–26). Five platelet infusions were required to stabilize the platelet level in this animal. The observed decrease in granulocytes and platelets was due to a combination of the short half-life of these cells and the failure of bone marrow hematopoietic cell production, as revealed by the CFC assay (Fig. 3). Erythrocyte counts and hemoglobin levels were only transiently affected (Fig. 4b, macaques 13586 and 13696; and data not shown). Red blood cells are not directly targeted by XF-RIC, and their decrease could thus be due to multiple microbleeding episodes related to thrombocytopenia. This hypothesis is supported by the petechial rash observed between days 19 and 25. Both animals received red blood cell transfusions when their hemoglobin concentration dropped below 8 g/dl.
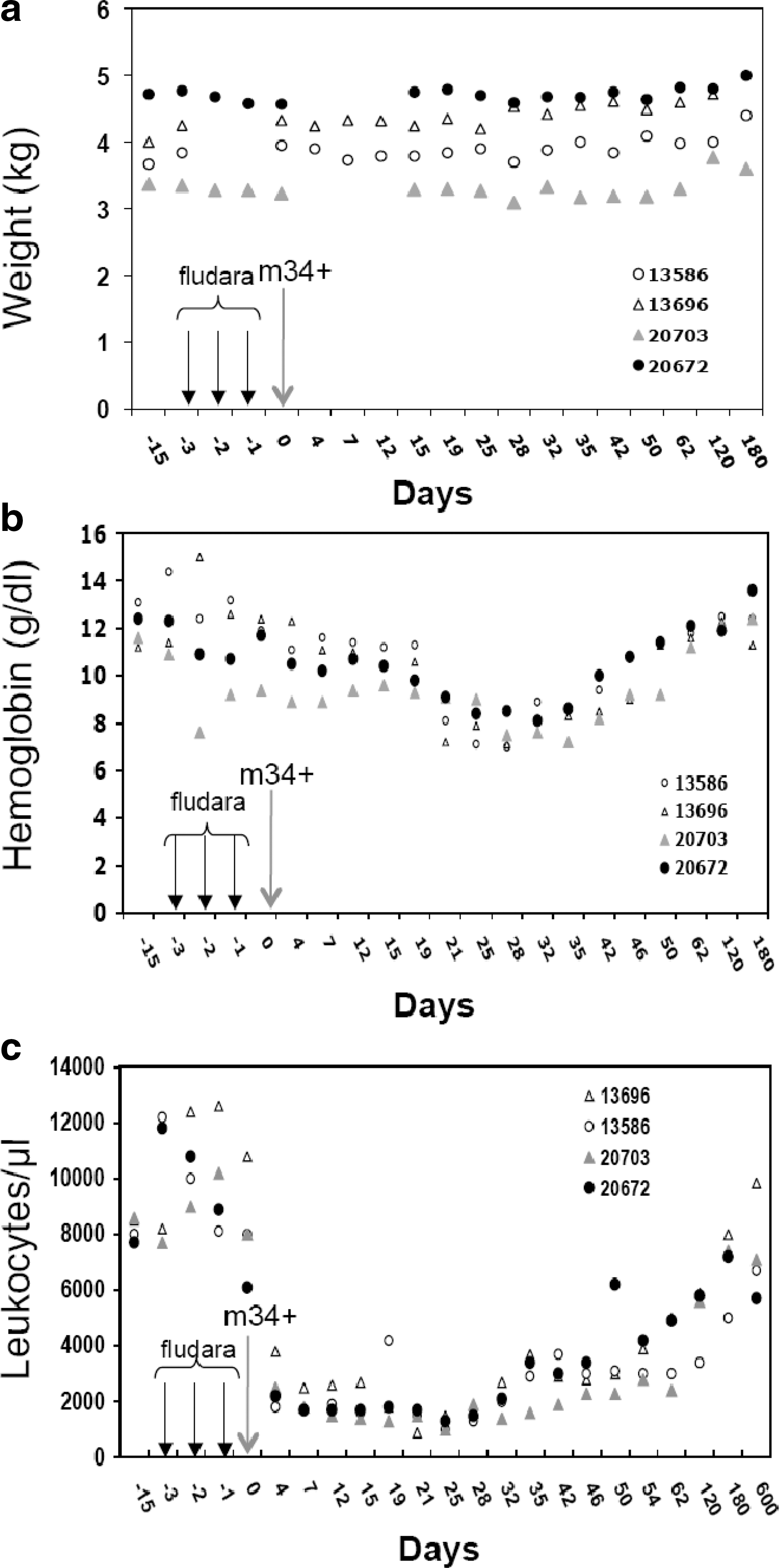
Follow-up of animals receiving conditioning alone or combined with autologous engraftment. Four animals (13586, 13696, 20703, and 20672) were subjected to the conditioning protocol outlined in Fig. 2. Only animals 20703 and 20672 received an autologous graft of SIV-LV-transduced mCD34+ cells (large arrow). The three smaller arrows indicate the time points of fludarabine injection, and the autologous mCD34+ cells were reinfused on day 0. The weight
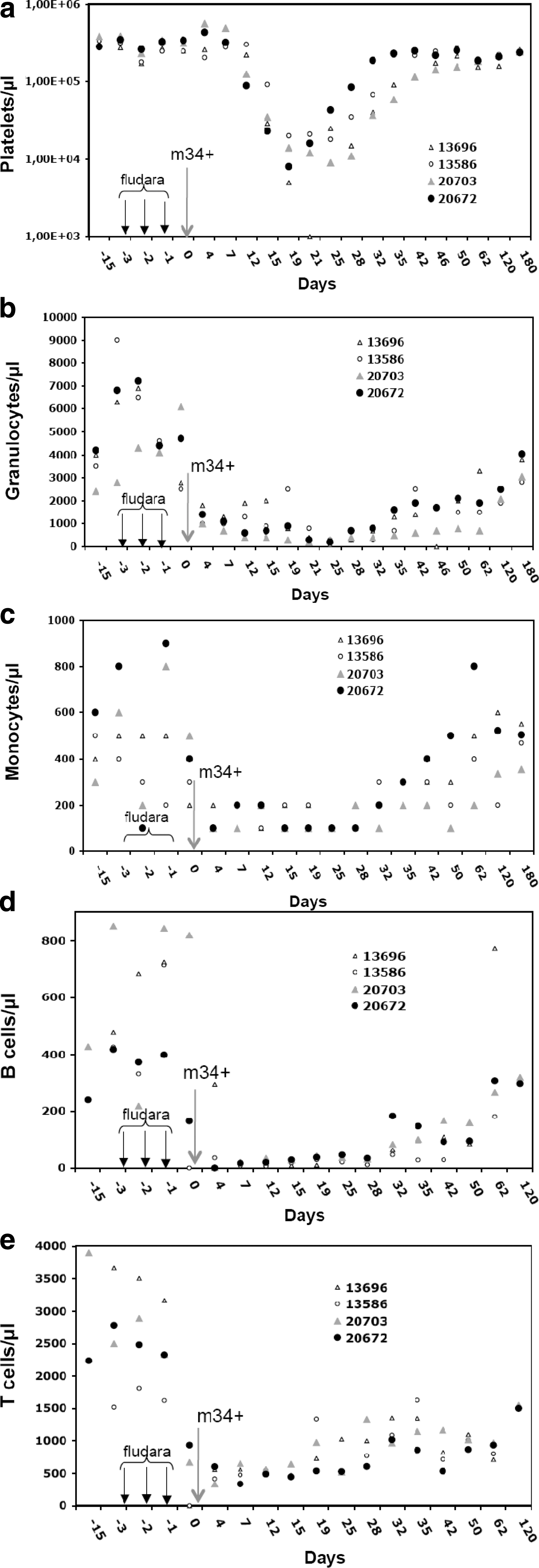
Detailed follow-up of myeloid and lymphoid cell counts in animals receiving conditioning alone or combined with autologous engraftment. Four animals (13586, 13696, 20703, and 20672) were subjected to the conditioning protocol outlined in Fig. 2. Only animals 20703 and 20672 received an autologous graft of SIV-LV-transduced mCD34+ cells (larger arrow). The three smaller arrows indicate the time points of fludarabine injection, and the autologous mCD34+ cells were reinfused on day 0. The myeloid cell counts (
XF-RIC and SCF-displaying SIV vectors result in improved long-term multilineage autologous engraftment with vector-bearing cells
Two other macaques (20672 and 20703) were then subjected to the same XF-RIC protocol used for the previous animals (13586 and 13696), except that gene transfer with SCF-displaying LV was also applied. One day before starting fludarabine injections, BM aspirates were taken from both the left and right humerus of the animals. Macaque CD34+ cells were isolated by immunobead selection from the BM and purity was determined (Table 1). The cells of both animals were subsequently transduced, each with a different preparation of SCF/VSV-G-SIV-LV at an MOI of 20 without addition of any exogenous cytokines or other additives. This resulted in high transduction efficiencies of 73% (20672) and 49% (20703) eGFP+mCD34+ cells on the day of cell injection (Table 1). Transduced cells were infused 1 day after the last fludarabine injection (Fig. 2). The animal showing the highest preinfusion transduction received 2.7×106 cells/kg, whereas the other animal received 3.7×106 cells/kg (Table 1).
Both animals recovered without adverse events. Close follow-up of these transplanted animals showed similar myeloid and lymphoid cell recovery compared with those that did not receive a graft (Fig. 5b–e). None of the autologous engrafted animals needed platelet or blood cell infusion because their counts did not drop under critical levels (<20,000 platelets/μl or <8 g/dl hemoglobin).
The proportion of genetically modified cells was then scored by both qPCR for the presence of the vector provirus and flow cytometry for eGFP expression. Both nucleated PB and BM cells were analyzed and the kinetics recorded up to the time of sacrifice: 156 weeks (∼3 years) and 93 weeks (∼2 years) PTx for animals 20672 and 20703, respectively (Fig. 6a–d).
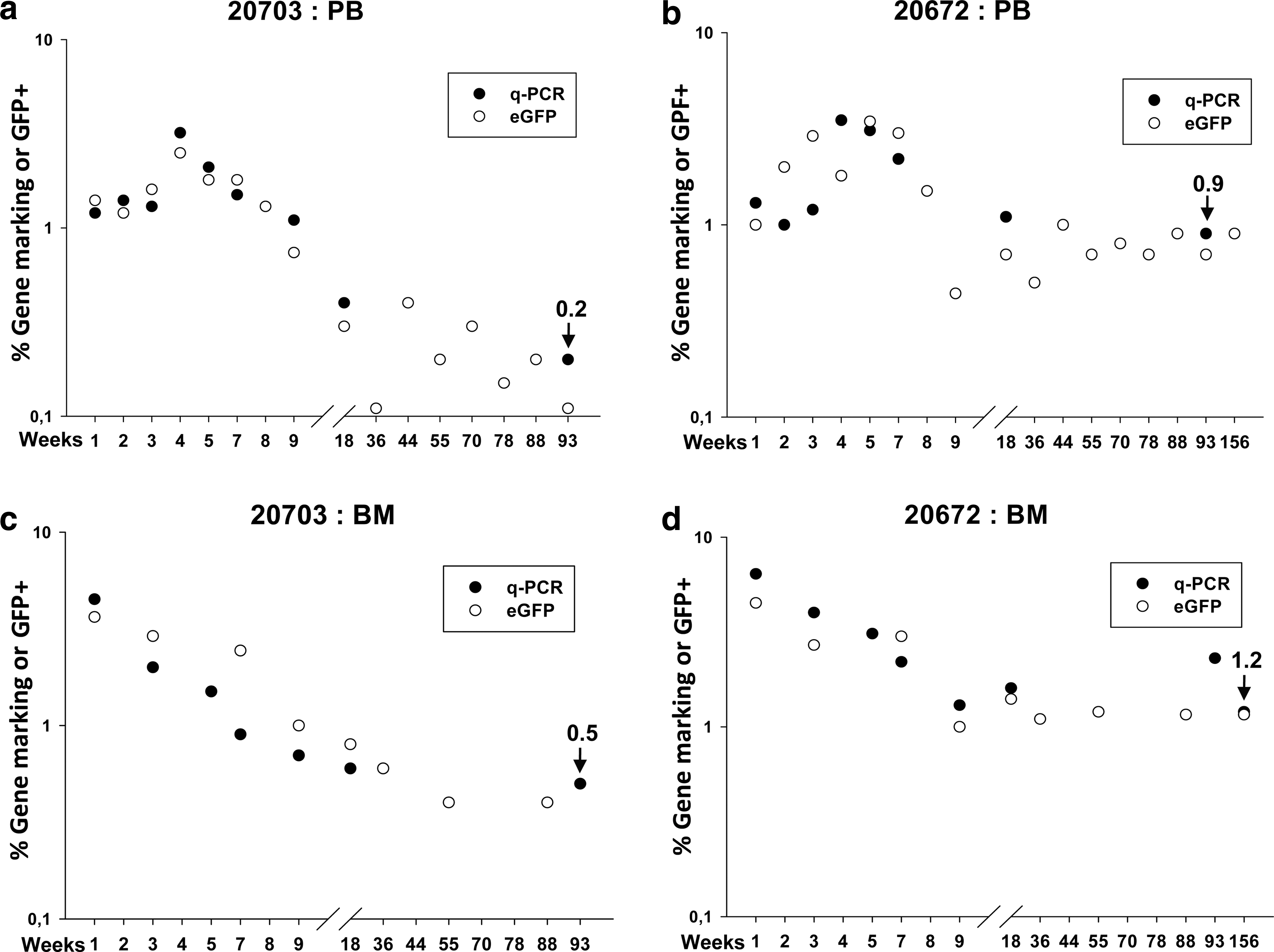
Engraftment of gene-marked macaque CD34+ cells after a combination of XF-RIC and autologous transplantation. Complete follow-up of eGFP-expressing nucleated cells and copy numbers of integrated SIV provirus per cell in the peripheral blood
By qPCR, 1 to 3.5% of the nucleated PB cells were marked in both animals up to ∼day 50 posttransplantation. As expected from other animal and human studies, levels subsequently decreased, but they remained at about 1% (animal 20672) and 0.2% (animal 20703), approximately 3 and 2 years PTx, respectively. These long-term results compare favorably with data from the previous three published reports (Table 2). The proportion of PB cells positive for eGFP expression was similar to qPCR data with a slight lower trend (Fig. 6a–d).
BM-MNCs, bone marrow mononuclear cells; PB-MNCs, peripheral blood mononuclear cells; RIC, reduced intensity conditioning; TBI, total body irradiation.
Cell marking, scored by qPCR and eGFP, in BM followed exactly the same profile as in PB cells, with a slight increase in overall values when compared with PB (Fig. 6a–d).
Of note is that the animal that showed the highest transduction of mCD34+ cells pretransplantation (20672; Table 1) also showed the highest level of long-term gene-marked cells in both PB and BM cells. We also wanted to rule out a possible residual toxic effect of the last fludarabine injection on the engrafted cells. In our protocol, we injected the transduced CD34+ cells by the intravenous route, 20 hr after the last fludarabine injection. We thus subjected two other macaques to the same XF-RIC and harvested sera at the time of infusion of the transduced cell population, as per the established XF-RIC protocol (Fig. 2). Freshly isolated macaque CD34+ cells were incubated with these sera and then seeded into a CFC assay. No significant reduction in CFC numbers was obtained for the sera of the fludarabine-treated animals versus the control sera (Supplementary Fig. S5).
After the sacrifice of both animals, gene expression was assessed in the various hematopoietic lineages. Although results were similar for both animals, only data for the longest-kept animal (20672), 156 weeks (∼3 years) PTx, are presented (Fig. 7). There were equivalent levels of eGFP in BM CD34+ progenitor cells, CD20+ B and pro-B cells, and CD11b+ granulocytes (Fig. 7a). On enrichment of the BM population for eGFP+ cells, CD34+ cells, B cells, granulocytes, and platelets were detected in the sorted population (Supplementary Fig. S3). Because granulocytes and platelets have short life spans, this suggests that HSCs capable of long-term reconstitution were transduced and engrafted in this animal. In the thymus, eGFP marking was found in all thymic subpopulations (CD4+ and CD8+ single and double populations) (Fig. 7b). Last, eGFP+ cells were revealed in peripheral CD4+CD8+ T cells, CD20+ B cells, and CD14+ monocytes (Fig. 7c). In agreement with the theory of central tolerance induced by mixed chimerism, no anti-eGFP-specific antibodies were detected in the serum of the animals (Supplementary Fig. S4).
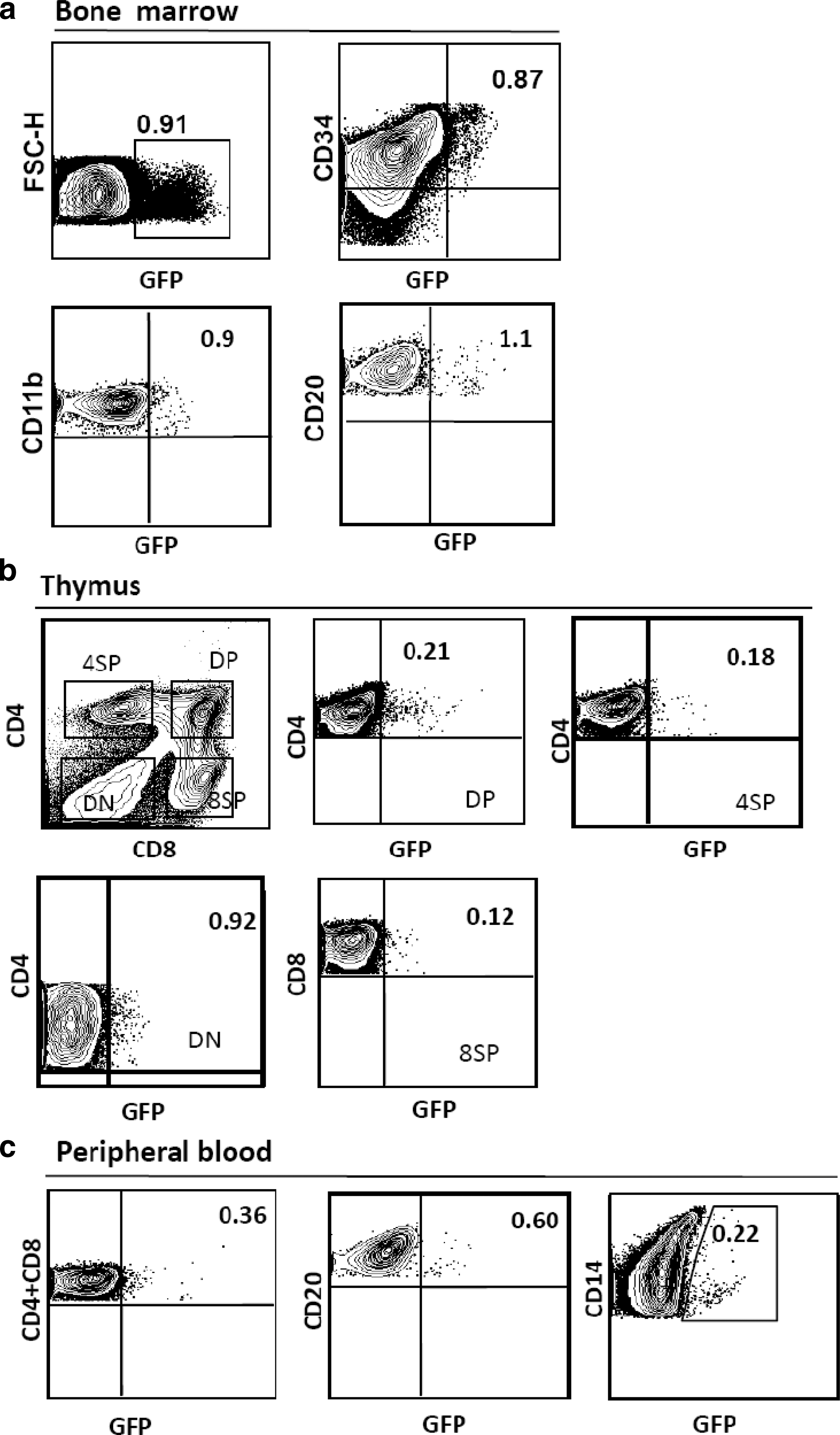
Long-term multilineage reconstitution with genetically modified cells. Macaques receiving autologous transplants were killed 3.1 years postengraftment and all hematopoietic tissues were collected (thymus, peripheral blood, and bone marrow). The percentage of eGFP+ cells in the various hematopoietic lineages was determined by FACS analysis using phenotype-specific antibodies. In
During the entire 3-year follow-up period, neither biological signs of myelo- or lymphoproliferation nor clonal expansion was detected. No replication-competent lentivirus (RCL) emerged during the same period, as assessed, for instance, by detection of the p27 SIV capsid protein (data not shown).
Discussion
These data show that the SCF-displaying SIV vector allowed for ex vivo gene transfer into autologous HSCs capable of long-term repopulation in the macaque and their sustained persistence, using the XF-RIC regimen, at levels unprecedented with previously reported lentiviral vector and RIC combinations.
A major restriction for the use of LVs in nonhuman primate transplant models is that HIV-derived lentiviral vectors transduce CD34+ cells from Old World monkeys (baboons, rhesus, and macaques) poorly, in contrast to their effective use for human cells. This is attributed to restriction factors that interfere at several postentry steps of HIV in simian cells (Horn et al., 2002; Stoye, 2002; Trobridge et al., 2008). It has been shown that HIV-LVs can efficiently transduce pigtail macaque CD34+ cells because of the absence of functional TRIM5α restriction factors (Trobridge et al., 2008). However, to develop LVs available for all nonhuman primate models, we switched to an SIV-based LV system we previously developed (Mangeot et al., 2000; Mangeot, 2002). These late-generation SIN-SIV-SFFV-GFP-LVs allowed for stable transduction of up to 90% of macaque CD34+ cells. This is in agreement with the transduction levels of nonhuman primate CD34+ cells reported by Hanawa (2004).
Although LVs transduce dividing cells efficiently, they do not allow for efficient gene transfer into a subpopulation of quiescent (G0) HSCs (Sutton et al., 1999), and this hampers their application for HSC gene therapy. To overcome these limitations, current protocols for human or nonhuman primate LV transduction of CD34+ cell employ high vector input together with cytokine cocktails (TPO, SCF, Flt-3, IL-6, IL-3) at high dosage to induce HSC cell cycle entry (Horn et al., 2002; Hanawa, 2004; Derdouch et al., 2008; Trobridge et al., 2008; Hayakawa et al., 2009). RetroNectin, a fragment of fibronectin, is often included to colocalize vector particles and target cells. Other additives such as protamine sulfate are often added or multiple administrations of vector are applied to achieve high gene transfer rates in HSCs (Horn et al., 2002; Hanawa, 2004; Derdouch et al., 2008; Trobridge et al., 2008; Hayakawa et al., 2009). However, the undesirable effects of extended cytokine stimulation and the aforementioned additives include a decrease in the multipotentiality and long-term engraftment of human as well as nonhuman primate HSCs (Tisdale et al., 1998; Glimm et al., 2000; Jetmore et al., 2002). Moreover, a dose of vector that is too high poses the risk of multiple vector copy integration with the possible untoward consequence of insertional mutagenesis (Woods, 2002; Hematti et al., 2004; Seggewiss et al., 2006). In an effort to resolve this conundrum (the requirement for cytokine stimulation with its deleterious consequences), we had previously reported the design of novel LVs that are surface-engineered to display “early-acting cytokines” to target and promote gene transfer in resting HSCs while preserving their stem cell potential (Verhoeyen et al., 2005). Although effective at transducing early human hematopoietic progenitors, their usefulness in nonhuman primate transplant models and their ability to transduce HSCs capable of long-term repopulation in a large animal closely related to humans were unknown before the present study.
Here, we demonstrate that SIV-LVs engineered to display a single cytokine, SCF, allowed for efficient gene transfer to autologous CD34+ cells capable of long-term reconstitution in the macaque. Surface display of TPO, alone or in combination with surface display of SCF, did not enhance gene transfer beyond levels achieved with SCF display alone. These sustained results obtained after long-term (up to 3 years) transplantation in a large animal closely related to humans validate LV surface display of SCF as an effective means of gene transfer to long-term HSCs without additives or major cytokine stimulation. It is all the more convincing that an RIC regimen was used, as discussed below, to achieve this result.
In the present study, we also attempted to leverage the adjunct radiosensitizing and mildly myelosuppressive properties of fludarabine to supplement 2-Gy TBI, referred to as XF-RIC, in an autologous setting in the macaque. This regimen is compatible with immunocompromised fragile patients or patients suffering from malignancies (Hentschke et al., 2003; Kelly et al., 2007; Mitsuyasu et al., 2009; Shimoni et al., 2010; Thakar et al., 2010). As expected, this protocol induced extensive cytopenia in all blood lineages. However, XF-RIC was well tolerated in the macaque, without severe toxic side effects, similar to what has been reported with human patients (Goodman et al., 1996).
After autologous transplantation in macaques by XF-RIC, transgene marking and expression were still detected in lymphoid and myeloid lineages of the blood, thymus, and BM up to the time of sacrifice for each of the two macaques, approximately 2 and 3 years PTx, respectively. The three previous studies evaluating RIC regimens in nonhuman primates in autologous settings after ex vivo LV transduction have made use of a decreased dosage of the myeloablative agent busulfan, administered intravenously (Brenner et al., 2006; Kahl et al., 2006), or TBI at 2 Gy or less (Kang et al., 2001). Kang and colleagues (2001) reported that the degree of chimerism for vector-bearing PB cells rapidly dropped to levels below 0.1% after 2-Gy TBI. Kang and colleagues achieved PB marking levels below 0.01 and 0.05%, depending on the dose of busulfan used (Kang et al., 2006). Kahl and colleagues (2006) reported that gene marking over a 6-month PTx period ranged from undetectable (no busulfan) to below 0.1% in PB cells from animals achieving the highest busulfan area under the curve (Kahl et al., 2006; Stein et al., 2010). Long-term results of the present study are substantially higher than those reported, although they remain modest. One should, nevertheless, emphasize that the degree of chimerism obtained for vector-bearing and expressing cells is now within the range of efficacy for certain diseases, which include the hemophilias (Roth et al., 2001), or for the establishment of sustained central tolerance to alloantigens (Kung et al., 2003).
The fact that the proportion of PB cells positive for eGFP expression was similar to qPCR data with a slightly lower trend is likely explained by the possibility that only a small number of cells express eGFP below the detection threshold and/or that the mean vector copy number is slightly above 1. The internal SFFV promoter used in this study is primed to silencing due to methylation of the promoter itself over the long term, as became clear during the GCD gene therapy trial and during in vitro and in vivo experiments in mice (Ott et al., 2006; Zhang et al., 2010). In contrast, Stoddart and colleagues (2009) used an LV carrying an SFFV internal promoter for transduction of macaque CD34+ cells that were reinfused into the animals. After several months of engraftment, T cells isolated from the periphery still expressed the gene under the control of the SFFV promoter, indicating that no silencing occurred. In the present study, this silencing phenomenon is unlikely because there was a strong correlation between gene marking detected by qPCR and eGFP expression levels.
Hence, several reasonable hypotheses can be considered to explain the relatively low degree of long-term in vivo gene marking. The most obvious explanations are that gene transfer to macaque HSCs remains partial within the graft and that RIC protocols do not eradicate all endogenous HSCs that cannot be exposed to the vector. Additional possibilities exist, as follows.
For instance, there might be an immune response against cells expressing eGFP. Some studies have described the induction of an anti-eGFP response or the appearance of cytotoxic T cells directed against eGFP after myeloablative or nonmyeloablative conditioning and autologous engraftment of transduced CD34+ cells (Rosenzweig, 2001; Morris, 2004). However, eGFP-specific central immunological tolerance in myeloablated primates engrafted with autologous LV-transduced CD34+ cells has also been reported (Kung et al., 2003). Morris (2004) reported that animals with an eGFP-specific immune response completely lost gene marking. This was not the case in our study because eGFP+ cells persisted for 2 years after engraftment, and this observation implies that no substantial immune response against eGFP was induced. Indeed, no anti-eGFP-antibody response was detected in the serum of either of the engrafted animals in our study (Supplementary Fig. S4).
A possible immune response against VSV-G or SCFHA might also have been induced because of VSV-G/SCFHA-LV vector particles that were still attached to the surface of the macaque CD34+ cells at the moment of reinfusion. It is true although that in the gene therapy trial by Cavazzana-Calvo and colleagues (2010), using VSV-G-pseudotyped lentiviral vectors for hCD34+ transduction, no signs of anti-VSV-G immune response were reported. In our study, the transduced cells were washed thoroughly before reinfusion into the animals to avoid transfer of vector particles. Moreover, the vector doses that we applied were not as high as reported in that clinical trial and a strong immunosuppressive response was induced by the RIC at the time of autologous cell reinfusion. Nevertheless, we cannot formally exclude the possibility that some vector particles were still attached to the cell surfaces and induced an immune response, possibly resulting in a sharp decline in gene-modified cells on autologous engraftment.
Another point to consider is the correlation between the number of genetically modified CD34+ cells infused and the level of engraftment obtained in vivo, as revealed by multiple human clinical trials (Hacein-Bey-Abina et al., 2002; Cartier et al., 2009; Cavazzana-Calvo et al., 2010). In the present study, we infused 1.9×106 and 1.8×106 eGFP+ cells/kg, which is low compared with other studies using a myeloablative conditioning regimen that resulted in efficient engraftment (Hanawa, 2004; Kim et al., 2009). However, these authors used strong cytokine cocktails leading to cell expansion. In contrast, SCF displayed on the vector surface in the present study induced only slight expansion and is therefore thought to preserve HSC potential.
The last possibility we have envisaged is the residual toxic effect of fludarabine metabolites on the injected transduced cells. This has been ruled out by experiments we conducted (Supplementary Fig. S5).
The risk of leukemia due to vector integration remains a concern for HSC gene therapy. It is obvious that the higher the vector copy number per cell, the higher the risk to induce genotoxicity, all other parameters considered. In our study, VSV-G/SCF-LV transduction of mCD34+ cells resulted in a low mean copy number of integrated provirus per cell (close to 1; data not shown). In comparison, current CD34+ transduction protocols, which use cytokine cocktails, result in a mean of several copies per cell (data not shown). Therefore, we might speculate that VSV-G/SCF-LVs have a reduced risk of genotoxicity while sustaining high transduction levels. The detailed follow-up of our animals did not show clonal outgrowth or signs of malignancy at any time point, indicating that no obvious genotoxic adverse events occurred during 3 years of engraftment.
Although this study focused on the combination of XF-RIC with VSV-G/SCF-LVs, it would be of interest to compare the engraftment of autologous macaque CD34+ cells transduced with SCF-displaying vector with current methods employing exogenous cytokine prestimulation in the context of myeloablative conditioning. In addition, long-term gene marking may ultimately be improved by means of vector built-in selection systems.
Footnotes
Acknowledgments
The authors thank Benoit Delache, Thibault Andrieu, and Christophe Jouy for excellent technical assistance. The authors acknowledge the support given by the following grants: the Agence Nationale pour la Recherche contre le SIDA et les Hépatites Virales (ANRS), the Association Française contre les Myopathies (AFM no. 13787), and the European Community (FP7-HEALTH-2007-B/222878, PERSIST; E-RARE-06-01, GETHERTHAL), ANR-05-MRAR006-01, PRES UniverSud 2010-16, and NIH grant HL090921 to P.L.
Author Disclosure Statement
E.V., F.R., M.C., C.C., D.N., F.L., C.J., R.L.G., F.L.C., A.D.-K., and S.P. have nothing to declare. P.L. is involved with Bluebird Bio (Cambridge, MA).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.