
Abstract
Asthma is a chronic, inflammatory, respiratory disease caused by an abnormal reactivity against allergens. The most promising treatments for asthma are based on specific immunotherapies, but they lack efficiency and can induce deleterious side effects. Among new modalities of immunotherapy currently in development, DNA vaccination presents a promising approach, as it enables targeted immunotherapy in association with reduced allergenicity. We have developed an innovative, DNA-based vaccine against Dermatophagoides farinae 1 allergen (Der f 1), one of the allergens most commonly encountered by asthma patients in Europe. Intramuscular administration of a Der f 1-encoding plasmid formulated with the block copolymer 704 in healthy mice induced a strong humoral and cellular response with a pro-helper T cell type 1 bias. Administration of the same formulation in asthmatic mice, according to an early vaccination protocol, led to a reduction of airway hyperresponsiveness and a significant decrease in the level of inflammatory cytokines in the bronchoalveolar lavage of Der f 1-vaccinated mice.
Introduction
Patients with asthma are usually treated with corticosteroids, which, however, only suspend the disease and are associated with deleterious side effects (Barnes, 2010). An alternative treatment for allergic asthma is based on a specific immunotherapy protocol: the repeated administration of increasing doses of allergen to induce hyposensitivity, and hence reduced symptoms when another subsequent exposure to this allergen occurs. Nevertheless, the efficacy of immunotherapy remains limited (Pipet et al., 2009), and its efficacy is variable between patients. Moreover, immunotherapy may cause an anaphylactic crisis when administered subcutaneously.
Thus, new modalities for immunotherapy are under development, notably based on DNA vaccination, which presents a promising approach. Here, DNA encoding immunodominant peptide sequences of the allergen are administered instead of allergen extracts, leading to a targeted immunotherapy associated with reduced allergenicity. Some studies have shown the therapeutic potential of this strategy. Jarman and Lamb reported that intramuscular administration to asthmatic mice of a naked plasmid encoding an immunodominant peptide of Der p 1, which is one of the major allergens in allergic asthma, led to a decrease in type 2 cytokines in bronchoalveolar lavages compared with untreated mice (Jarman and Lamb, 2004). The antigen-encoding plasmids may also be formulated with adjuvants that are able to modulate the immune response toward a helper T cell type 1 (Th1) or type 2 (Th2) bias as reported by Kim and colleagues, who used bacillus Calmette-Guérin, known to induce a Th1 bias, in a DNA vaccination protocol against Der p 2 (Kim et al., 2006). Nevertheless, the amounts of DNA used in these studies were too large to consider a future application in humans. Moreover, no study has demonstrated that DNA vaccination against the specific allergen, Der f 1, could prevent or reduce the development of asthmatic symptoms in a relevant animal model.
Thus, the future application of DNA vaccines in humans requires the development of a new, efficient, and safe adjuvant that is capable of inducing a strong humoral and cellular response with a low dose of injected DNA. Previous studies by our group showed that formulations comprising plasmid DNA in association with a new class of synthetic vector, a tetrafunctional block copolymer, were able to safely increase the transfection efficiency of reporter or therapeutic genes in lung, skeletal muscle, and cardiac muscle in healthy subjects and animal models of disease compared with results achieved with naked DNA (Pitard et al., 2004; Richard et al., 2005; Jespersen et al., 2007; Piron et al., 2008; Richard-Fiardo et al., 2008; Biliczki et al., 2009). It has also been shown that the tetrafunctional block copolymer 704 is able to promote low-dose DNA vaccination efficiency (McIlroy et al., 2009). This novel class of vector has also been used to treat hepatocellular carcinoma in a highly autochthonous, relevant mouse model (Cany et al., 2011).
In the present study, we developed an innovative synthetic DNA vaccine, based on block copolymer against the allergen Der f 1, to modulate the allergic response in asthmatic mice. The formulation of the Der f 1 vaccine has been optimized in a model of healthy mice, and allowed to elicit a strong humoral and cellular response with a Th1 and CD8+ T cell type 1 (Tc1) bias. Administration of this formulation in a highly relevant new model of asthmatic mice, according to an early vaccination approach, led to a significant reduction in airway hyperresponsiveness, and of levels of inflammatory cytokines in the bronchoalveolar lavage (BAL), which were, in contrast, present at higher levels in the lungs of nonvaccinated asthmatic mice.
Materials and Methods
Animal procedures
BALB/c mice (Elevage Janvier, Le Genest, France) were housed under conventional conditions according to INSERM (Institut National de la Santé et de la Recherche Médicale) guidelines. Mice, 8 weeks of age, were used for all experiments. For intramuscular DNA vaccination, mice were anesthetized by isoflurane inhalation, and then various DNA–polymer formulations were injected into both tibialis anterior muscles, using an Insumed Pic Indolore 30-gauge syringe (Artsana, Grandate, Italy). Two sites were injected per animal, and DNA doses were given via the tibialis anterior. In all cases, the injection volume was 50 μl per injection site. After vaccination, sera and spleens were recovered for analysis of the immune response at the end of each experiment.
Plasmid preparation and formulation
The pCMV-βgal plasmid (Clontech, Saint-Germain-en-Laye, France) encoding β-galactosidase, and the pVAX-Derf1 plasmid encoding Der f 1, under the control of the human cytomegalovirus immediate promoter, were used as antigen. The pVAX-Derf1 plasmid was obtained by cloning the Der f 1 gene sequence into pVAX (Invitrogen, Courtaboeuf, France) with HindIII and XhoI restriction enzymes. pQE30 (Qiagen, Courtaboeuf, France) was used as carrier DNA. All plasmids were purified by passage through EndoFree plasmid purification columns (Qiagen). The tetrafunctional block copolymers 304, 704, and 904 were kindly supplied by In-Cell-Art (Nantes, France). Plasmid DNA was formulated immediately before intramuscular injection, as previously described (McIlroy et al., 2009).
Der f 1 expression
For Der f 1 expression experiments, tibialis anterior muscles were dissected and immediately frozen in liquid nitrogen. Der f 1 expression was then quantified in muscle extracts, using a Der f 1 ELISA kit (Indoor Biotechnologies, Wiltshire, UK) according to the manufacturer's protocol.
Measurement of immune response
Humoral immune responses were measured by ELISA. For anti-Der f 1 IgG, IgG1, and IgG2a antibody dosage, 96-well plates (MaxiSorp; Nunc/Thermo Fisher Scientific, Roskilde, Denmark) were coated overnight at 4°C with purified Der f 1 (2.5 μg/ml) (Indoor Biotechnologies) in NaHCO3 (50 mmol/liter), pH 9.5, and then blocked for 1 hr at room temperature with phosphate-buffered saline (PBS)–0.05% Tween 20–1% bovine serum albumin, before distributing diluted sera in duplicate. Plates were incubated at 37°C for 90 min, and then Der f 1-specific IgG, IgG1, and IgG2a were detected with peroxidase-conjugated goat anti-mouse IgG (Jackson ImmunoResearch, Newmarket, UK) diluted 1:5000, goat anti-mouse IgG1 (AbD Serotec, Oxford, UK) diluted 1:10,000, and goat anti-mouse IgG2a (AbD Serotec) diluted 1:10,000 in PBS–0.05% Tween 20, respectively. Peroxidase activity was revealed with o-phenylenediamine (1 mg/ml) in pH 5 citrate buffer. Reactions were stopped by addition of H2SO4 (1 mol/liter), and optical density was measured at 490 nm. Anti-Der f 1 IgG titers were calculated from the regression curve of a standard Der f 1-specific mouse serum included in each ELISA plate. The absolute titer of the standard was 204,800 and was diluted from 1:1000 to 1:64,000 to build a calibration curve. Each tested sample was diluted two times and dilutions with optical density (OD) included in the linear part of the calibration curve were conserved for antibody titer determination. Other dilutions were excluded. Similarly, for anti-β-galactosidase (β-Gal) IgG antibody dosage, 96-well plates (MaxiSorp; Nunc/Thermo Fisher Scientific) were coated overnight at 4°C with purified β-Gal (5 μg/ml) (Roche, Rosny-sous-Bois, France) in NaHCO3 (50 mmol/liter), pH 9.5, and then blocked for 1 hr at room temperature with PBS–0.05% Tween 20–1% bovine serum albumin, before distributing diluted sera in duplicate. Plates were incubated at 37°C for 90 min, and then β-Gal-specific IgG was detected with peroxidase-conjugated goat anti-mouse IgG (Jackson ImmunoResearch) diluted 1:5000 in PBS–0.05% Tween 20. Peroxidase activity was revealed with o-phenylenediamine (1 mg/ml) in pH 5 citrate buffer. Reactions were stopped by addition of H2SO4 (1 mol/liter), and optical density was measured at 490 nm. Anti-β-Gal IgG titers were calculated from the regression curve of a standard β-Gal-specific mouse serum included in each ELISA plate. The titer of the standard was arbitrarily fixed at 5000 and was diluted from 1:1000 to 1:64,000 to build a calibration curve. Each tested sample was diluted two times and dilutions with OD included in the linear part of the calibration curve were conserved for antibody titer determination. Other dilutions were excluded.
As a paradigm readout of the cellular response to block copolymer 704 vaccine formulation, the Der f 1-specific CD8+ response was tested by class I-restricted interferon (IFN)-γ secretion determined by enzyme-linked immunospot (ELISPOT) (AbCys, Paris, France). Sixteen 8-mer Der f 1-derived peptides predicted to bind with H-2Kb were selected on the basis of the binding score, as calculated by BIMAS and NetMHC software (Supplementary Table S1; supplementary data are available online at
Allergic asthma mouse model
BALB/c mice were sensitized and challenged with total extract of Dermatophagoides farinae, kindly supplied by Stallergenes (Antony, France), according to the protocol described in Fig. 5. Mice were sensitized percutaneously in each ear with 250 μg total extract of HDM diluted in 10 μl of dimethyl sulfoxide, and challenged by intranasal inhalation of 250 μg total extract of HDM diluted in 40 μl of PBS.
Measurement of airway responsiveness
Airway responsiveness was measured in conscious, unrestrained mice using barometric, whole-body plethysmography by recording respiratory pressure curves (EMKA Technologies, Paris, France) in response to inhaled methacholine (Sigma-Aldrich, St. Louis, MO) at concentrations of 5–20 mg/ml for 1 min. Airway responsiveness was expressed in enhanced pause (P enh) units, a calculated value, which correlates with the measurement of airway resistance, impedance, and intrapleural pressure in the same mouse: P enh=(t e/t r−1)×PEF/PIF (where t e is the expiration time, t r is the relaxation time, PEF is the peak expiratory flow, and PIF is the peak inspiratory flow).
Bronchoalveolar lavage analysis
Mice were tracheotomized and administered 1 ml of sterile PBS intratracheally through a flexible catheter. Cells and supernatants from recovered fluid were separated by centrifugation. Total cell number was determined by optical microscopy and after cell samples were spotted on Cytospin slides using a cytocentrifuge (Cytospin; Thermo Fisher Scientific). Cell proportions were established after May-Grünwald Giemsa staining of about 300 cells.
Cytokine analysis
Expression levels of cytokines interleukin (IL)-4, IL-5, IL-13, and IFN-γ were determined in BAL supernatant by Luminex technology. A bead-based Bio-Plex kit (Bio-Rad Laboratories, Munich, Germany) was used to measure IL-4, IL-5, IL-13, and IFN-γ. The assays were performed according to the manufacturer's specifications. Cytokines were quantified on the basis of an eight-point calibration curve constructed from the standard provided. Data analysis was performed with Bio-Plex Manager software version 4.0.
Statistical analysis
Statistical analysis was conducted with SigmaStat 2.03 software (Systat, San Jose, CA). All data are expressed as means±standard deviation. Statistical analysis was performed by Student–Newman test. p<0.05 was considered significant.
Results
Optimization of formulation containing various amounts of DNA encoding Der f 1
We previously showed that a formulation containing β-galactosidase-encoding plasmid with the block copolymer 704 led to high antigen expression in vivo, correlating with a strong humoral and cellular response against β-Gal (McIlroy et al., 2009). Here, mice were injected intramuscularly with plasmid encoding the Der f 1 antigen formulated with block copolymer 704. Five days after injection, muscles were collected and evaluated for Der f 1 expression levels by ELISA. As shown in Fig. 1, injection of naked plasmid DNA resulted in no significant expression of the Der f 1 protein in muscle, irrespective of the amount of injected DNA. Conversely, injection of plasmid DNA formulated with the block copolymer 704 led to a significant increase in Der f 1 expression in muscle when the amount of injected DNA encoding Der f 1 was increased.
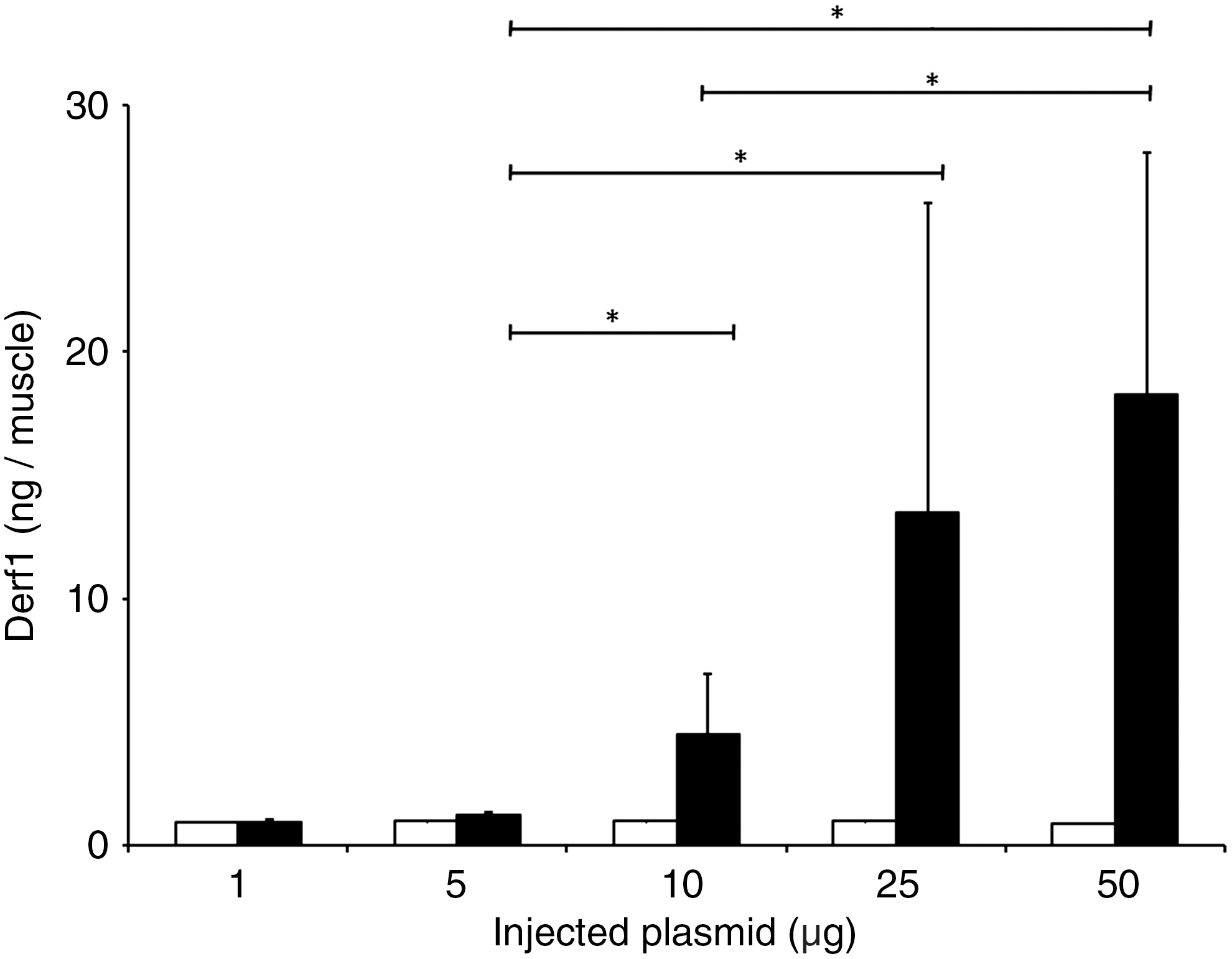
Formulation of pVAX-Derf1 with block copolymer 704 increases the Dermatophagoides farinae 1 allergen (Der f 1) expression level after intramuscular injection. Mice were injected intramuscularly with Der f 1-encoding plasmid DNA, either alone (open columns) or formulated with block copolymer 704 (solid columns). Mice were killed 5 days later and Der f 1 expression levels were analyzed by ELISA. *p<0.05 by Student Newman–Keuls test.
We then evaluated the ability of this synthetic formulation to elicit specific humoral and cellular immune responses against Der f 1. Mice were injected intramuscularly on days 0 and 21 with various amounts of pVAX-Derf1 formulated with block copolymer 704. Cellular and humoral responses were analyzed on days 21 and 42 after the first injection. Dose–response experiments showed a maximal humoral response for 10 μg of pVAX-Derf1, which then remained at this level for higher doses (Fig. 2a). To analyze the cellular response, we synthesized sixteen 8-mer immunodominant peptides from Der f 1, predicted to bind to the H-2Kb-restricted MHC-I molecule (Supplementary Table S1). These peptides were used in IFN-γ ELISPOT assays to evaluate the specific CD8+ T cell response induced by Der f 1 DNA/704 immunizations in mice. Results showed that the cellular response increased with increasing amounts of injected pVAX-Derf1 formulated with block copolymer 704 and reached a plateau at 10 μg of pVAX-Derf1 (Fig. 2b). To measure the orientation of the immune response, we measured different, specific anti-Der f 1 antibody isotypes for various amounts of injected, formulated plasmid DNA. Formulation with block copolymer 704 led to the induction of a Th1 bias, as indicated by the predominance of Der f 1-specific IgG2a compared with Der f 1-specific IgG1 (Fig. 2c).
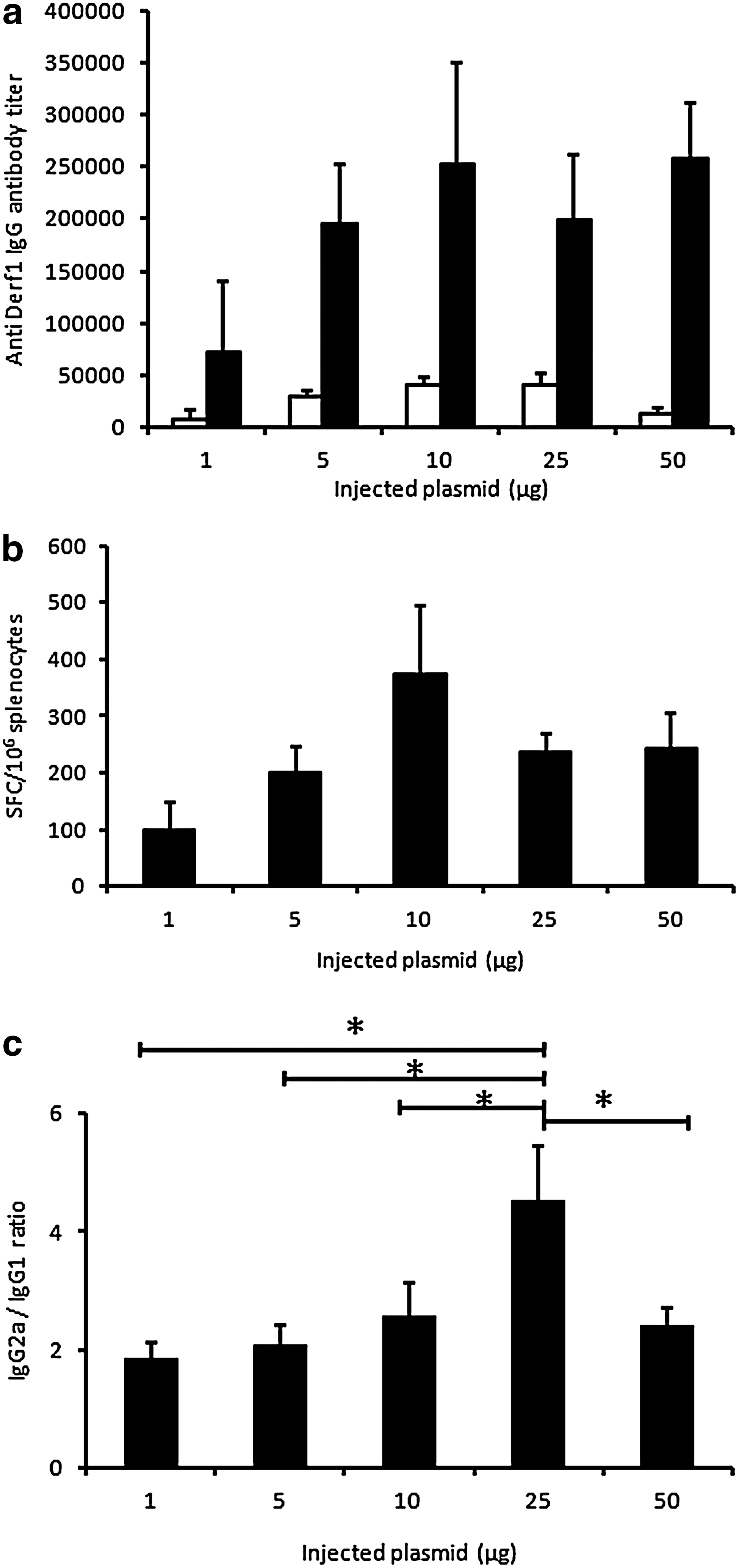
Influence of the amount of pVAX-Derf1 plasmid on humoral and cellular immune responses. Groups of mice (n=5) were injected intramuscularly with various amounts of pVAX-Derf1, ranging from 1 to 50 μg, formulated with block copolymer 704, on days 0 and 21.
In a previous study we also reported the adjuvant property of noncoding DNA when added to DNA vaccination (McIlroy et al., 2009). Thus, we tested the effect of noncoding DNA on Der f 1 vaccine formulation efficiency. Mice were injected on days 0 and 21 with Der f 1-encoding plasmid only, or in the presence of noncoding plasmid pQE30, formulated with block copolymer 704. As previously reported, decreasing the amount of coding plasmid, while maintaining the total amount of DNA, led to similar humoral and cellular immune response levels (Fig. 3a and b). Figure 3a also shows that with only 3 μg of pVAX-Derf1 formulated with block copolymer 704, the same anti-Der f 1 antibody titer was obtained as with 5 or 10 μg of formulated plasmid DNA encoding Der f 1. Results also show that 0.5 μg of plasmid encoding Der f 1, in the presence of pQE30, was sufficient to lead to a strong humoral response against Der f 1, which was similar to that obtained with a 20-fold higher amount of formulated plasmid encoding Der f 1. In the same experimental group of mice, the class I-restricted cellular response indicated that plasmid DNA encoding Der f 1 formulated with block copolymer 704 led to the same high ELISPOT response irrespective of the amount of formulated pVAX-Derf1, in the presence or absence of pQE30.
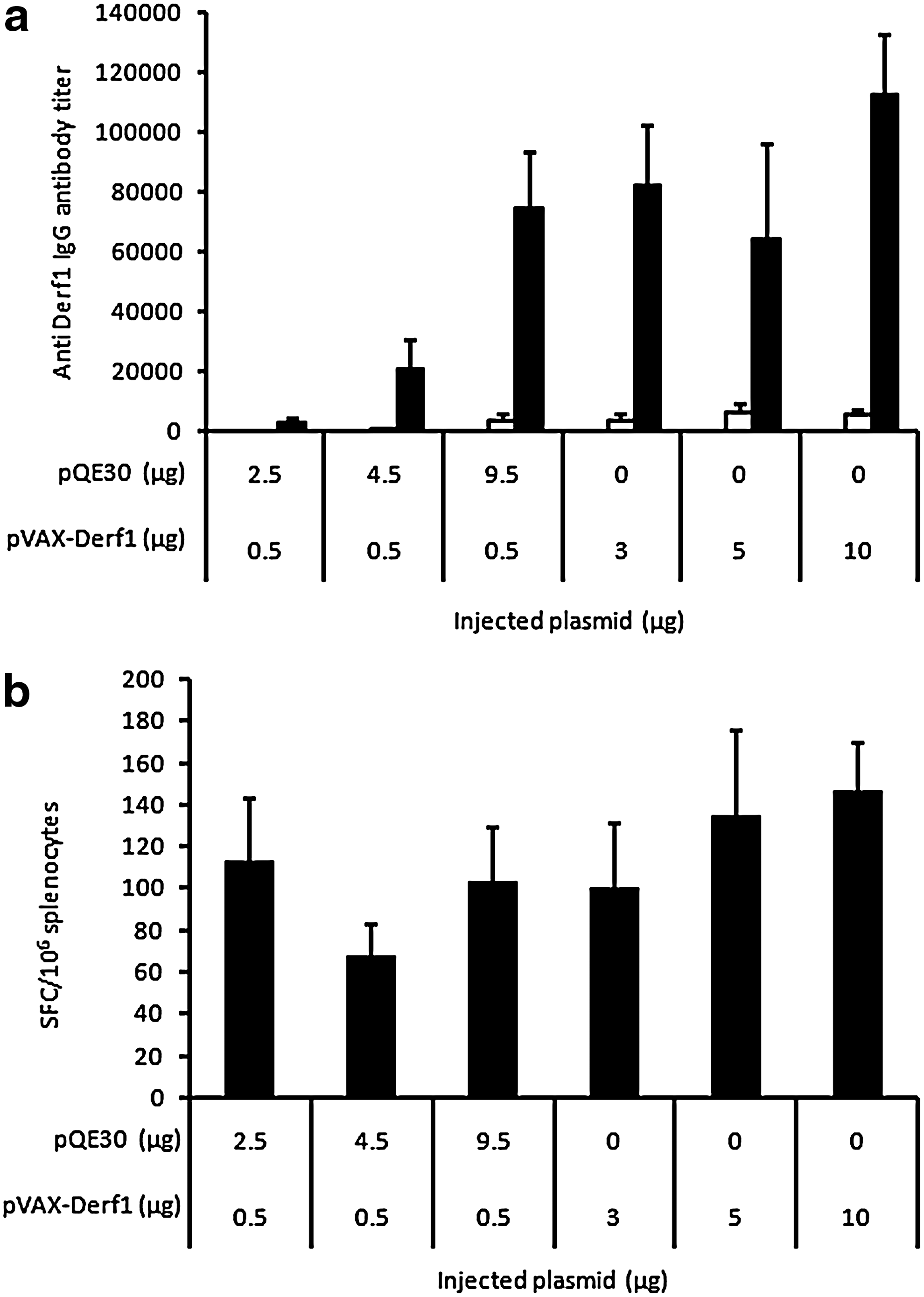
Conservation of vaccination efficiency with a low dose of Der f 1-encoding plasmid and carrier DNA associated with block copolymer 704. Groups of mice (n=5) were injected intramuscularly on days 0 and 21 with various amounts of pVAX-Derf1, in the presence or absence of plasmid carrier DNA, formulated with block copolymer 704.
Optimization of vaccine formulation and protocol
To optimize the anti-Der f 1 vaccine formulation, we tested the effect of three different block copolymers that were previously shown to enhance reporter gene expression in vivo and vaccination efficiency. Mice were injected intramuscularly on days 0 and 21 with 10 μg of pVAX-Derf1 plasmid formulated at two doses of block copolymer 304, 704, or 904. Each formulation led to a significant and specific humoral immune response on day 42, whereas the maximal humoral response was obtained with block copolymer 704 at a concentration of 0.15% (Supplementary Fig. S1a). All formulations tested were able to elicit a cellular response. Interestingly, block copolymer 704 at 0.15% promoted a high cellular response, as measured by INF-γ ELISPOT assay (Supplementary Fig. S1b). Moreover, the various polymers tested provided a Th1 bias, as attested by the predominance of Der f 1-specific IgG2a compared with Der f 1-specific IgG1 (Supplementary Fig. S1c). We subsequently investigated the influence of mouse strain on the immune response. Mice from two different strains, BALB/c and C57BL/6, were injected intramuscularly on days 0 and 21 with 10 μg of pVAX-Derf1 plasmid formulated with block copolymer 704. On day 42, BALB/c and C57BL/6 inbred mice, which are typically used for immunological studies, had similar anti-Der f 1 antibody titers (Supplementary Fig. S2), indicating that mouse strain did not influence vaccination efficiency against Der f 1.
Given that DNA vaccination usually requires a repeated injection scheme to elicit the development of a strong immune response characterized by high antibody titers and memory B cell activation, we investigated the influence of the prime–boost injection protocol on the immune response. Mice were injected intramuscularly on day 0 with 10 μg of pVAX-Derf1 plasmid formulated with block copolymer 704, and then boosted 1, 2, 3, or 4 weeks later. Antibody titers were analyzed each week during 10 weeks (Fig. 4). It was clear that the mice that received a boost 1 or 2 weeks after the primary injection presented low antibody titers, comparable with those obtained using only one injection. On the other hand, mice receiving a boost injection 3 or 4 weeks after the primary injection showed a robust increase in their antibody titers, occurring 2 weeks or 1 week after the boost injection, respectively. Seventy days after the primary injection, antibody titers were still detectable for all conditions. As boosting at 21 days after the primary injection led to the sustained presence of antibodies in serum, we selected this injection scheme for Der f 1 vaccination in an allergic mouse model.
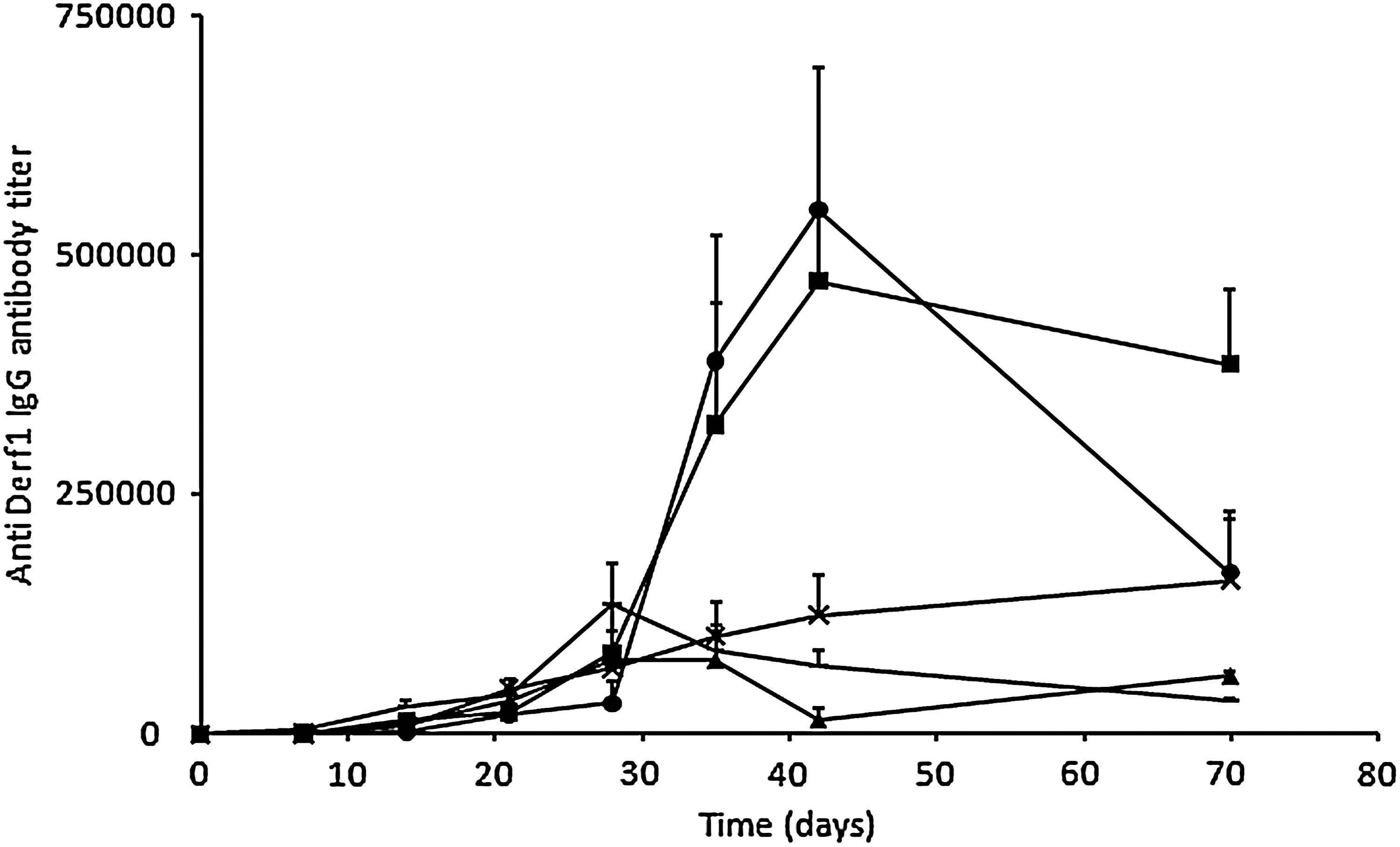
Influence of the number of boost injections and time of boost injection on the amplitude and kinetics of the humoral response. Groups of mice (n=5) were injected with 10 μg of pVAX-Derf1 formulated with block copolymer 704 and the anti-Der f 1 antibody IgG titer was determined at various time points. The mean titer and standard deviation are shown for each group. Doses given are per tibialis anterior. Groups of mice were injected on day 0 only (triangles), on days 0 and 7 (diamonds), on days 0 and 14 (mult symbols), on days 0 and 21 (squares), or on days 0 and 28 (circles). Doses given are per tibialis anterior.
Der f 1 vaccination in an allergic asthma mouse model
The efficiency of our vaccination formulation against Der f 1 was tested in a model of asthmatic mice. The allergic asthma phenotype was induced with a total house dust mite (HDM) extract in BALB/c mice. First, mice were sensitized by percutaneous administration of 250 μg of total HDM extract, once per week for 4 weeks (days 0, 7, 14, and 21). They were then challenged three times by intranasal inhalation of 250 μg of total HDM extract, on days 28, 42, and 49. Two different pVAX-Derf1/704 vaccination schemes of asthmatic mice were used to study the impact of anti-Der f 1 vaccination in these mice.
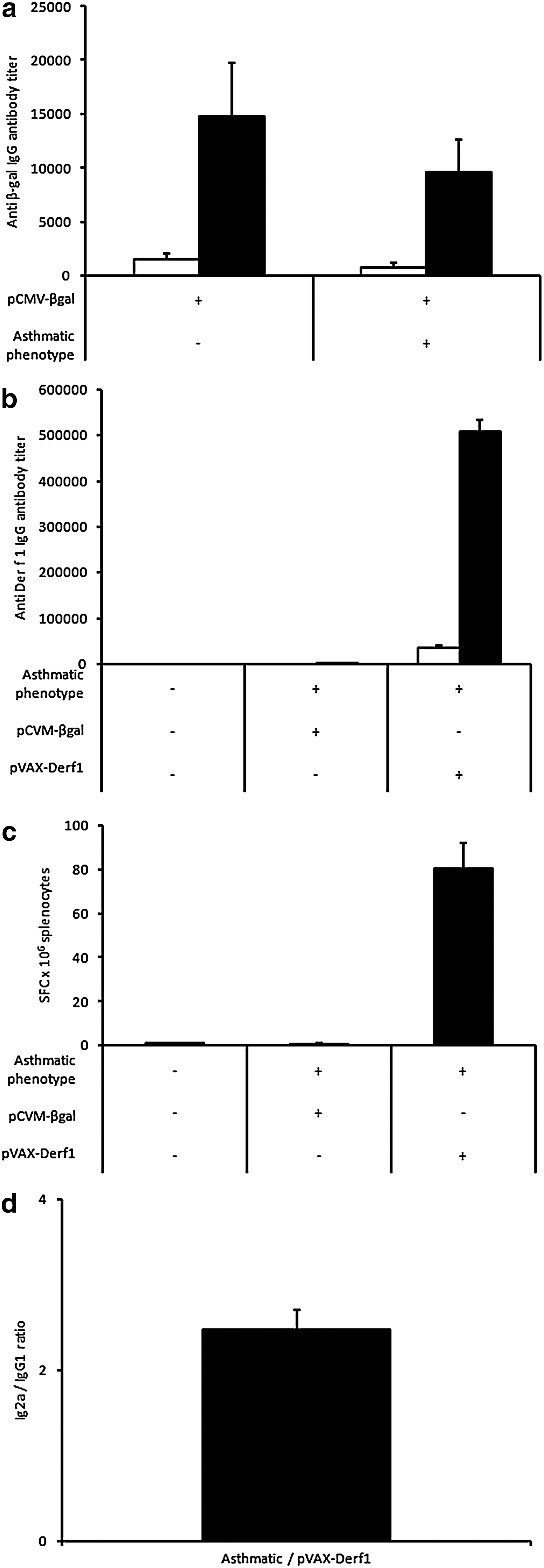
In the first setting, BALB/c mice were vaccinated at the onset of HDM sensitization (Fig. 5a). First, we evaluated the influence of the mouse model on block copolymer 704-mediated vaccination efficiency. To this end, mice were immunized with the β-galactosidase antigen at the onset of HDM sensitization. Mice were immunized twice, according to the protocol described in Fig. 5, using pCMV-βgal reporter plasmid formulated with block copolymer 704. The humoral response was assessed on day 52, after the last HDM challenge. The dosage of specific, anti-β-galactosidase IgG antibodies indicated that no significant differences between healthy or asthmatic mice were obtained. This result shows that the induction of an asthmatic phenotype does not modify the immune response against an antigen expressed by gene transfer with block copolymer 704 (Fig. 6a). As a control, we also investigated whether the immunization protocol with block copolymer 704 against Der f 1 could induce, by itself, an asthmatic phenotype. Analysis of respiratory parameters of healthy mice immunized with the synthetic vector pVAX-Derf1/704 showed no differences compared with untreated healthy mice, indicating that the immunization protocol did not promote an asthmatic phenotype (data not shown). Then, at the same time as the HDM sensitization, mice were immunized on days 0 and 21, using pCMV-βgal plasmid as control plasmid or using pVAX-Derf1 plasmid formulated with block copolymer 704. On day 52, after the last HDM challenge, various parameters were assessed. Results showed that asthmatic mice vaccinated against Der f 1 present high levels of Der f 1-specific IgG antibodies (Fig. 6b), associated with a strong Th1 bias, as attested by the predominance of Der f 1-specific IgG2a compared with Der f 1-specific IgG1 (Fig. 6d). As a control, the dosage of the specific Der f 1 IgG antibodies in nonvaccinated healthy mice, and in β-galactosidase-vaccinated asthmatic mice, did not show the presence of Der f 1-specific IgG antibodies. This indicates that production of the asthmatic mouse model did not induce a specific immune response against Der f 1. Under the same experimental conditions, healthy and asthmatic mice vaccinated against β-galactosidase did not present a significant cellular response, whereas asthmatic mice vaccinated against Der f 1 presented a high level of IFN-γ-secreting splenocytes stimulated by a cocktail of Der f 1 peptides (Fig. 6c).
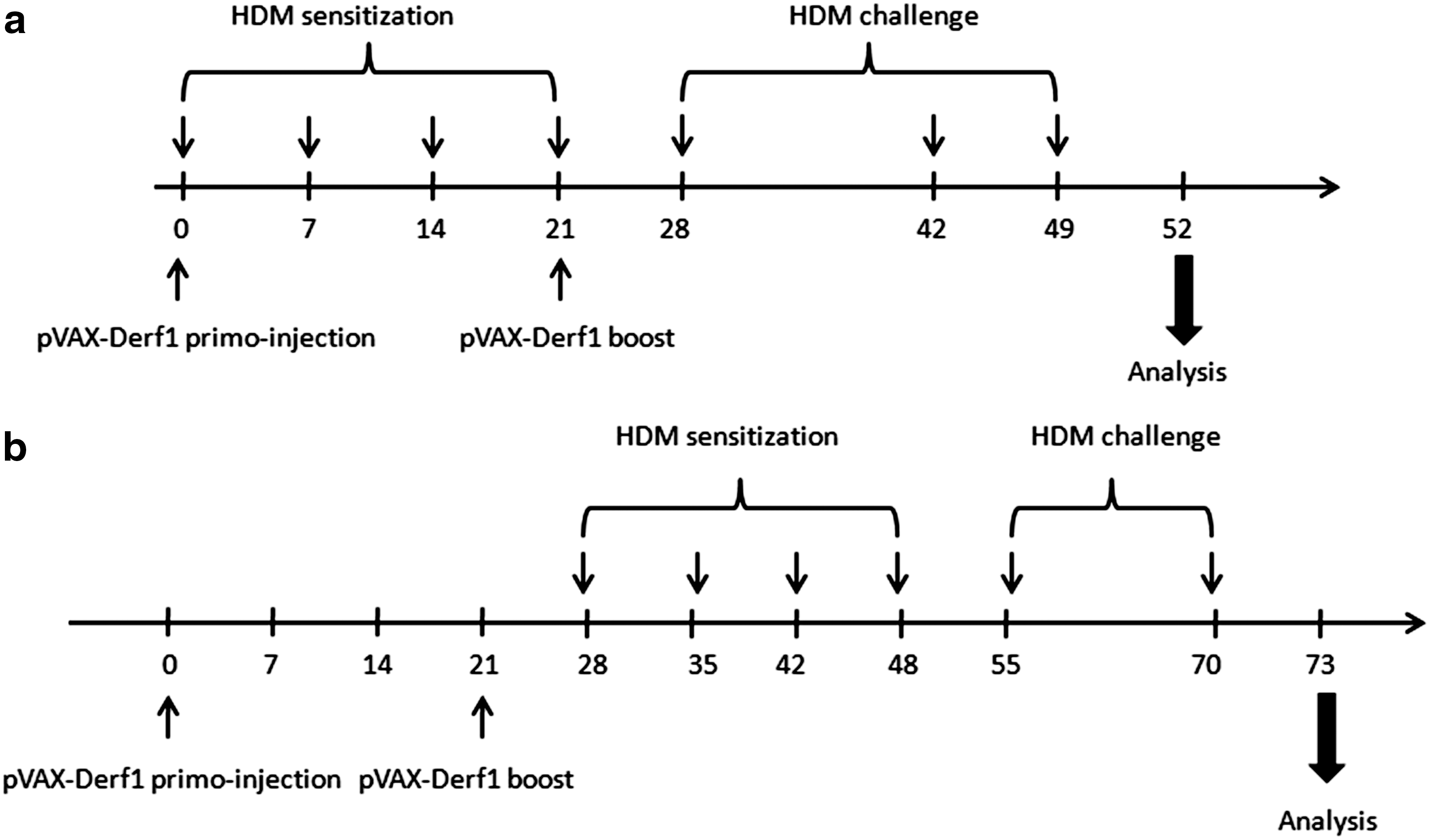
Immunization protocols against Der f 1 in asthmatic mice.
Effect of a prophylactic immunization protocol on the immune response of asthmatic mice. Groups of mice (n=5) were injected intramuscularly on days 0 and 21 according to a prophylactic protocol (see Fig. 5) with 50 μg of plasmid formulated with block copolymer 704.
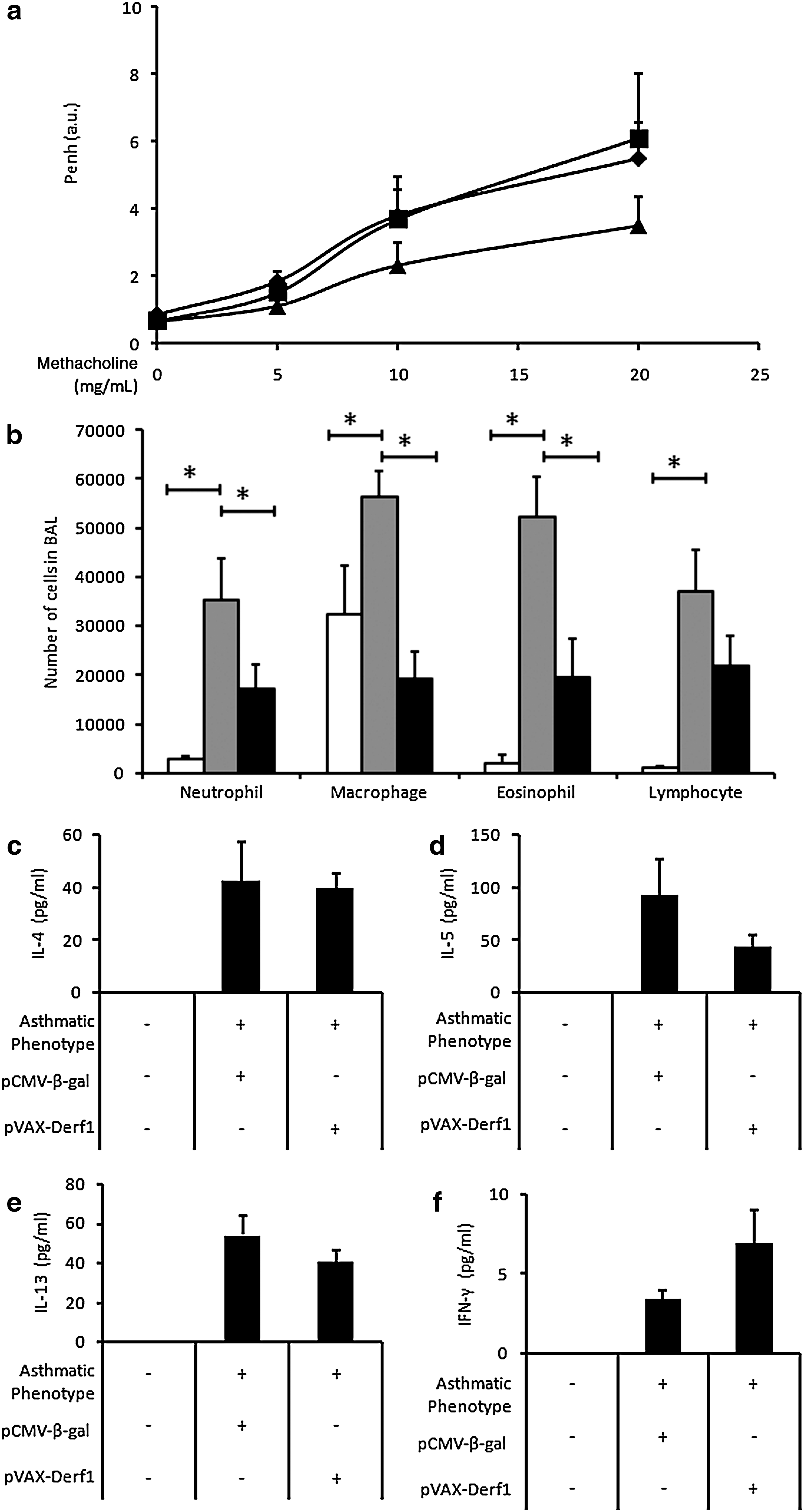
Asthma is usually characterized by airway hyperresponsiveness and by an influx of inflammatory cells and cytokines into the lungs. Airway responsiveness was measured after inhalation of methacholine (5 to 20 mg/ml), which induces bronchoconstriction, and expressed as enhanced pause (P enh). Results show that asthmatic mice presented a high P enh value compared with healthy mice (Fig. 7a). Asthmatic mice immunized against β-galactosidase or Der f 1 antigen presented no significant variation of P enh compared with nonvaccinated, asthmatic mice. The influx of inflammatory cells into the lungs was assessed by cell counts in bronchoalveolar lavage (BAL) and by histological analysis of lung tissues. The BAL of healthy mice contained essentially macrophages, whereas the BAL of asthmatic mice contained macrophages and also numerous neutrophils, eosinophils, and lymphocytes. Interestingly, the asthmatic, Der f 1-vaccinated mice presented significantly reduced amounts of macrophages and eosinophils compared with asthmatic mice vaccinated against β-galactosidase antigen (Fig. 7b). The numbers of neutrophils and lymphocytes were also reduced. We also determined the expression of inflammatory cytokines in the BAL, using Luminex technology. The BAL of asthmatic mice immunized against the control antigen, β-galactosidase, presented high levels of IL-4, IL-5, and IL-13, which are proinflammatory cytokines, and reduced levels of IFN-γ, which is a pro-Th1 cytokine. Conversely, asthmatic mice presented reduced levels of IL-5 and IL-13 and an increased level of IFN-γ in BAL, compared with asthmatic mice vaccinated against the β-galactosidase antigen (Fig. 7c–f). Altogether these data show that, in this first setting, vaccination of asthmatic mice against the major allergen, Der f 1, promoted a significant reduction in inflammatory cells and cytokines observed in BAL, as compared with asthmatic mice vaccinated against a control antigen.
Effect of immunization protocol on asthmatic phenotype of asthmatic mice. Groups of asthmatic mice (n=5) were injected intramuscularly on days 0 and 21 with pVAX-Derf1 plasmid formulated with block copolymer 704. Doses given are per tibialis anterior.
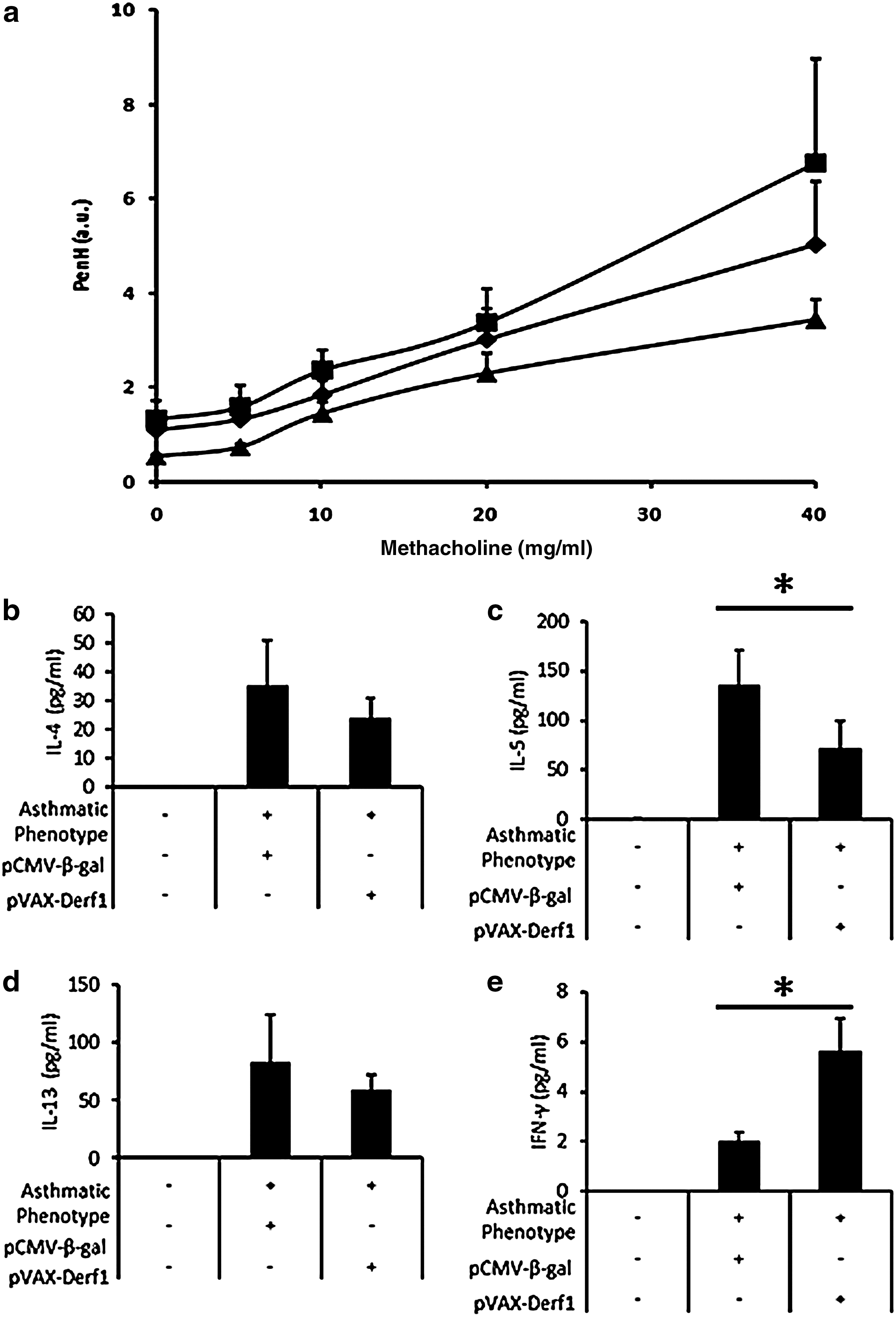
In the second setting, mice were given two vaccination injections on days 0 and 21. On day 28, they were HDM-sensitized followed by HDM challenges (see Fig. 5b). Mouse airway hyperreactivity was measured on day 73. Asthmatic mice immunized with the control antigen, β-galactosidase, presented a high P enh value compared with healthy mice, whereas asthmatic mice vaccinated with pVAX-Derf1 formulated with block copolymer 704 presented a reduction of P enh for all amounts of methacholine used, indicating a reduction of airway hyperresponsiveness (Fig. 8a). Measurements of inflammatory cytokine expression levels in the BAL, using Luminex technology, showed that the BAL of asthmatic mice immunized against the control antigen, β-galactosidase, presented high levels of IL-4, IL-5, and IL-13, which are proinflammatory cytokines, and reduced levels of IFN-γ, which is a pro-Th1 cytokine. Conversely, asthmatic mice presented reduced levels of IL-4, IL-5, and IL-13 and increased levels of IFN-γ in BAL, compared with asthmatic mice vaccinated against β-galactosidase antigen (Fig. 8b–e).
Effect of prophylactic immunization protocol in asthmatic mice on asthmatic phenotype. Groups of asthmatic mice (n=5) were injected intramuscularly on day 0 and day 21 with pVAX-Derf1 plasmid formulated with 704. Doses given are per tibialis anterior.
Taken altogether, these data show that early vaccination of asthmatic mice against the major allergen Der f 1 promotes a significant reduction of airway hyperresponsiveness and of cytokines observed in the BAL, as compared with asthmatic mice vaccinated with a control antigen. Thus, early vaccination of asthmatic mice may confer a protective effect on the development of asthmatic symptoms.
Discussion
The treatment and prevention of allergic asthma represent a major public health issue, and specific immunotherapy based on DNA vaccination seems to be a promising approach to reduce the incidence of this disease. Preclinical evaluation of treatments against allergic asthma requires adequate models. The most relevant mouse models of asthma rely on repeated sensitizations of mice with house dust mite (HDM) total extract followed by repeated intranasal exposures to HDM to induce an asthmatic phenotype that mimics the human disease (Cheng et al., 1998). We performed initial experiments that verified the kinetics of the HDM-induced asthma phenotype in mice. We observed that the development of the asthma phenotype in this model parallels the pattern observed in a clinical setting. Mice sensitized with HDM present bronchial hyperreactivity and important infiltration of inflammatory cells and cytokines in the lung and the BAL when challenged with HDM.
Der f 1 protein, from the mite Dermatophagoides farinae, is reported to be one of the major allergens encountered by patients with asthma. More than 50% of the house dust mite-allergic population are carriers of Der f 1 IgE-specific antibodies (Zock et al., 2006). A previous study showed that intramuscular administration of a plasmid encoding Der p 1, another allergen encountered in patients with asthma, leads to the reduction of proinflammatory cytokines in the BAL of treated mice (Jarman and Lamb, 2004).
However, Der f 1-specific immunization has never been tested in mice presenting an asthmatic phenotype. Because Der f 1 is a major allergen implicated in the development of the asthmatic phenotype, we sought to test Der f 1-specific immunotherapy in a mouse model of asthma induced by HDM sensitization. Immunization was performed in two different settings, before or simultaneously with HDM sensitizations, to mimic a clinical situation of prophylactic treatment before exacerbation of asthmatic symptoms.
In the present study, two injections, 3 weeks apart, with the synthetic vector pVAX-Derf1/704 led to a significant reduction of inflammatory cell infiltration and cytokines in the BAL of asthmatic mice, 52 days after the first immunization, compared with mice immunized with a control antigen. Further, early immunization against Der f 1, before induction of the asthmatic phenotype, led to a reduction of airway hyperresponsiveness and of inflammatory cytokine levels in the BAL of asthmatic mice, 73 days after the first immunization.
To our knowledge, this is the first demonstration that Der f 1 is an effective target, and well adapted for specific immunotherapy in allergic asthma. These results show that vaccination with a block copolymer 704-based vaccine induces a strong and specific humoral and cellular immune response, characterized by a strong Th1 bias (increase in IgG2a/IgG1 ratio) and Tc1 (increase in IFN-γ produced by CD8+ cells by stimulation of splenocytes of vaccinated mice with Der f 1 class I-restricted peptides). ELISA analysis revealed the presence of Der f 1-specific IgG induced by vaccination with a plasmid-encoded Der f 1 antigen complexed with the 704 polymer. Therefore, the hypothesis of mechanism is that the immune response induced by the vaccination of asthmatic mice with Der f 1/704 led to the systemic production of Der f 1-specific IFN-γ-secreting CD8+ T cells and anti-Der f 1 IgG2a antibodies. All together, these results show the induction of a systemic Tc1 and Th1 profile in the vaccinated asthmatic mice. Then, during the induction of asthma by giving nasally 250 μg of HDM protein extract containing Der f 1, Der f 1-specific IFN-γ-secreting CD8+ T cells migrate to the lungs, where they locally secrete IFN-γ in response to their stimulation by the presence of Der f 1 peptides presented by immune cells. This hypothesis is also supported by Fonseca and colleagues (2012), who have shown that cells transferred from naked DNA-immunized GFP transgenic mice to ovalbumin-induced allergic mice migrated to the allergic sites because they found GFP-positive allergen-specific CD8+ T cells in the lungs. It has also been described in the literature (Iwamoto et al., 1993; Tong et al., 2006) that IFN-γ has a well-known effect on the suppression of airway eosinophilia and on the downmodulation of the allergic response by directly inhibiting Th2 cells expressing IL-5. In the present study, we indeed observed this secretion of IFN-γ in the lungs and the concomitant reduction of (1) interleukin levels, (2) the presence inflammatory cells, and (3) airway hyperresponsiveness.
Therefore, the Tc1 and Th1 cells specific to Der f 1, induced by vaccination via the muscle with a low dose of DNA encoding Der f 1 in the presence of the synthetic vector 704, migrate to the allergic lungs exposed to the allergen and mediate by secreting IFN-γ the local immunomodulatory effect overcoming the deleterious effect of the Th2 response in the allergic condition.
Although the treatment significantly reduced the presence of inflammatory cytokines in the BAL of asthmatic mice, airway hyperresponsiveness was reduced only after the early vaccination protocol, and only a slight effect on airway hyperreactivity was observed in mice immunized concomitantly with the induction of the asthmatic phenotype. In this latter protocol, we analyzed respiratory parameters on day 52 after the first immunization with the synthetic vector, pVAX-Derf1/704. At this time point, the Der f 1-specific IgG antibody titer decreased, as shown in Fig. 4. Indeed, the Der f 1-specific IgG antibody titer was maximal on day 42. Thus, on day 52, IgG levels may not be sufficient to observe a significant reduction in airway hyperreactivity. The immunization protocol could be modified to generate higher levels of Der f 1-specific IgG antibody on day 52, by performing another immunization on day 42 when the IgG antibody level is maximal. We also wonder whether immunization against Der f 1, concomitantly with the sensitization protocol, may impair the efficiency of the immunization protocol. Indeed, as sensitization begins simultaneously with the Der f 1 immunization protocol, the immune system is not yet “educated” to recognize Der f 1 as an antigen, rather than an allergen. Consequently, the immune system produces both Der f 1-specific IgE and Der f 1-specific IgG antibodies, which impairs the efficiency of the Der f 1-specific immunization protocol. Results obtained with the early vaccination protocol are in agreement with this hypothesis, showing that early vaccination led to a better reduction of asthmatic symptoms.
Although a strong Th1/Tc1 bias was demonstrated that is likely to be responsible for the preventive effect of the vaccine on asthmatic inflammation, other mechanisms could be involved. During classical allergen-specific immunotherapy, it is well established that not only a Th2-to-Th1 recommitment but also regulatory T (Treg) cell activation occurs (Pipet et al., 2009). The involvement of Treg cells in the vaccination efficiency in the present mouse model of allergic asthma requires further investigation.
The clinical relevance of the preventive strategy used in the present study may be questionable because it seems difficult to envisage allergen-specific vaccination before the occurrence of sensitization. However, it is not aberrant to consider that in high-risk children with sensitization to allergens other than HDM, the prevention of Der f 1 allergy is crucial to prevent severe rhinitis and asthma.
In conclusion, our present study demonstrates that, in a clinically relevant model of allergic asthma, Der f 1 is a valid target for immunotherapy. DNA vaccination has produced encouraging results in mice, but the production of large amounts of DNA will be difficult to achieve before its transfer to the clinical setting. In contrast to previous studies, we performed a low-dose DNA immunization protocol using the 704 polymer, which offers a number of practical advantages, such as ease of production and purification in large quantities, and the absence of special handling or storage conditions. We have validated this immunotherapy strategy for the prevention of allergic asthma and we suggest that immunotherapy to prevent allergic disease represents an attractive alternative to classical, specific immunotherapy.
Footnotes
Acknowledgments
The authors are grateful to Emilie Goudeau for excellent technical expertise in ELISA and to Marie Aude Muller for excellent technical expertise in animal experimentation. They are also indebted to Benoit Barteau for technical expertise in synthetic DNA vaccination and for stimulating discussions. The authors are grateful to Stallergenes (France) for providing the Dermatophagoides farinae extracts. This work was supported by special grants from the Association Française contre les Myopathies (Evry, France), Vaincre la Mucoviscidose (Paris, France), Fondation Recherche Médicale, Société Française d'Allergologie, and Région Pays de Loire. F. Beilvert is the recipient of a Ph.D. fellowship from Vaincre la Mucoviscidose. M. Langelot is the recipient of a CIFRE contract with Agence Nationale pour la Recherche et la Technologie (ANRT) and Stallergenes.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.