
Abstract
Adeno-associated virus (AAV)-U7–mediated skipping of dystrophin-exon-23 restores dystrophin expression and muscle function in the mdx mouse model of Duchenne muscular dystrophy. Soluble activin receptor IIB (sActRIIB-Fc) inhibits signaling of myostatin and homologous molecules and increases muscle mass and function of wild-type and mdx mice. We hypothesized that combined treatment with AAV-U7 and sActRIIB-Fc may synergistically improve mdx muscle function. Bioactivity of sActRIIB-Fc on skeletal muscle was first demonstrated in wild-type mice. In mdx mice we show that AAV-U7–mediated dystrophin restoration improved specific muscle force and resistance to eccentric contractions when applied alone. Treatment of mdx mice with sActRIIB-Fc increased body weight, muscle mass and myofiber size, but had little effect on muscle function. Combined treatment stimulated muscle growth comparable to the effect of sActRIIB-Fc alone and dystrophin rescue was similar to AAV-U7 alone. Moreover, combined treatment improved maximal tetanic force and the resistance to eccentric contraction to similar extent as AAV-U7 alone. In conclusion, combination of dystrophin exon skipping with sActRIIB-Fc brings together benefits of each treatment; however, we failed to evidence a clear synergistic effect on mdx muscle function.
Introduction
Exon-skipping mediated restoration of dystrophin expression may not be a sufficient therapy if muscle wasting has already progressed. Therefore, combination of dystrophin-exon-skipping with strategies that stimulate muscle growth and regeneration may increase therapeutic efficiency especially at advanced disease stages. The transforming growth factor (TGF)-β family member myostatin (Mstn) has been identified as a negative regulator of muscle growth, and loss-of-function mutations of Mstn result in excessive muscle development in different species including one human case (Clop et al., 2006; Grobet et al., 1997; McPherron et al., 1997; Mosher et al., 2007; Schuelke et al., 2004). Mstn signals via activin receptor IIB (Acvr2B or ActRIIB) and inhibition of ActRIIB signaling also enhanced muscle growth (Lee et al., 2005; Lee and McPherron, 2001). Likewise, treatment with the soluble ligand binding domain of ActRIIB fused to the Fc domain of IgG (sActRIIB-Fc) increased muscle mass (Lee et al., 2005). sActRIIB-Fc sequesters Mstn and other TGF-β super family members, such as activin and growth and differentiation factor (GDF)-11, thereby inhibiting signaling by competing with the intrinsic transmembrane ActRIIB receptor (Sako et al., 2010; Souza et al., 2008). sActRIIB-Fc rapidly increased muscle mass and function in mouse and alleviated the disease phenotype in a number of mouse models for neuromuscular disorders and tumor cachexia (George et al., 2011; Lawlor et al., 2011; Morrison et al., 2009; Pistilli et al., 2011; Zhou et al., 2010). Importantly, multiple studies using diverse methods including sActRIIB-Fc showed that Mstn blockade in mdx mice stimulated muscle growth and strength, and some reports indicated an alleviated dystrophic phenotype (Bogdanovich et al., 2002, 2005; Haidet et al., 2008; Morine et al., 2010; Nakatani et al., 2008; Pistilli et al., 2011). These promising results led to first clinical trials using Mstn antibodies (Wagner et al., 2008) and sActRIIB-Fc (ClinicalTrials.gov Identifiers: NCT00952887, NCT00755638, NCT01099761, and NCT01239758).
So far it has not been investigated whether combination of sActRIIB-Fc with dystrophin-exon-skipping results in synergistic therapeutic effects. However, a recent study showed that a combination of adeno-associated virus (AAV)-mediated repression of endogenous ActRIIB with AAV-U7–induced dystrophin-exon-skipping synergistically improved force of mdx muscle, although no effect on muscle growth was observed (Dumonceaux et al., 2010).
In this study we combined AAV-U7 with sActRIIB-Fc and determined individual and combined effects of these treatments on muscle growth, muscle function, and histopathology in mdx mice.
Material and Methods
sActRIIB-Fc production
The human ActRIIB ectodomain was amplified via polymerase chain reaction (PCR) with the following primers: GGACTAGTAACATGACGGCGCCCTGG and CCAGATCTGCGGTGGGGGCTGTCGG from a plasmid containing the human ActRIIB sequence (in pCR-Blunt II-TOPO AM2-G17 ActRIIB, IMAGE clone #40005760, The IMAGE Consortium). A version of the human IgG1 Fc domain incorporating a C-terminal His6 tag was produced by PCR amplifying the Fc domain (GCAGATCTAATCGAAGGTCGTGGTGATCCCAAATCTTGTGAC and TCCCTGTCTCCGGGTAAACACCATCACCATCACCATTGAGCGGCCGCTT) from the pIgPlus expression plasmid. The PCR products were subcloned into the pGEM-T easy (Promega) vectors, sequenced, and fused before cloning into the expression vector pEFIRES-p (Hobbs et al., 1998). Chinese hamster ovary (CHO) cells were transfected with the ActRIIBecd–FcHis6 expression vector via lipofection (Fugene 6, Roche) and selected with puromycin (Sigma–Aldrich) as described by Kaivo-Oja et al. (2003). During expansion, cells were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 2 mmol/l L-glutamine, 100 μl/ml streptomycin, 100 IU/ml penicillin, and 10% fetal calf serum. For production DMEM–Hams F-12 (1:1) was used supplemented with 2 mmol/l L-glutamine, 100 μl/ml streptomycin, 100 IU/ml penicillin, 0.01% bovine serum albumin (BSA), and 6.6 ml/liter Heparin (the heparin preparation was Fragmin 2500 IE/ml [Pfizer]). Cell culture supernatants were clarified by filtration through 0.22-μm membrane (Steritop, Millipore). NaCl (1 M) and imidazole (5 mM) were added into the clarified supernatants, and the solution was pumped through a Ni2+-loaded HiTrap column (GE Healthcare Life Sciences, Uppsala, Sweden) at 4°C. Protein was eluted with raising imidazole concentrations, dialyzed against phosphate-buffered saline (PBS) and finally concentrated with Amicon Ultra concentrator (30,000 MWCO, Millipore).
AAV production
Dystrophin was rescued through U7 snRNA-mediated exon-skipping following AAV2/1 pseudotyped vector injections as described previously (Goyenvalle et al., 2004). Vectors were prepared according to previously published protocols (Riviere et al., 2006). Titer for AAV-U7 was 5×1012 vector genomes (vg)/ml. For control experiments, AAV-U7 was diluted (1/10), which was below the threshold to restore dystrophin expression and therefore termed AAV-U7-diluted.
Mice
For the experiment in wild-type mice, 6-week-old C57Bl/10SnJ mice were injected intraperitoneally twice a week with 10 mg/kg sActRIIB-Fc or PBS. To determine the acute effect of sActRIIB-Fc, six C57Bl/10SnJ mice received one injection of 10 mg/kg sActRIIB-Fc or PBS, after which the muscles were isolated 24 or 48 hr after injection. For the experiment with the combined treatments, 6-week-old mdx mice on a C57Bl/10SnJ background were injected once with either 50 μl AAV-U7 (concentration of 2.5×1011 vg/injection) into the left tibialis anterior (TA) muscles or with AAV-U7-diluted into the right TA muscles. AAV injections were performed under general anesthesia using isoflurane. Two weeks after intramuscular AAV injection, systemic treatment of mice with sActRIIB-Fc was initiated using twice weekly intraperitoneal injections of 10 mg/kg (n=6, three males and three females) or same quantity of PBS for control animals (n=6, three males and three females) for a total of 5 weeks. Experiments were performed according to institutional guidelines after approval of the Animal Experimental Commission (DEC) of the LUMC and the University of Jyväskylä (license number BD.01.284[E21] [LUMC] and ESLH-2009-08528/Ym-23 [University of Jyväskylä]). Total body weight was recorded twice weekly.
Force measurements
The function of TA muscles was evaluated by measuring in situ muscle contraction in response to nerve stimulation, as previously described (Foster et al., 2008; Mouisel et al., 2010; Sharp et al., 2011). Animals were anesthetized using intraperitoneal injection of pentobarbital sodium at 50 mg/kg. The knee and foot were fixed, and the distal tendon of the muscle was attached to a lever arm of a servomotor system (305B, Dual-Mode Lever, Aurora Scientific) using a silk ligature. Data were analyzed using the PowerLab system (4SP, ADInstruments) and software (Chart 4, ADInstruments). The sciatic nerves were stimulated using supramaximal square wave pulses of 0.1 msec duration. Force generation capacity was evaluated by measuring the absolute maximal force that was generated during isometric contractions in response to electrical stimulation (frequency of 75–150 Hz, train of stimulation of 500 msec). Maximal isometric force was determined at L0 (length at which maximal tension was obtained during the tetanus). Force was normalized to the muscle mass as an estimate of specific force. To determine resistance to eccentric contraction–induced damage, muscle fragility was estimated from the force drop resulting from lengthening contraction–induced injury. The sciatic nerve was stimulated for 700 msec (frequency of 150 Hz). A maximal isometric contraction of the TA muscle was initiated during the first 500 msec. Then, muscle lengthening (10% L0) at a velocity of 5.5 mm/sec was imposed during the last 200 msec. All contractions were made at an initial length L0. Nine lengthening contractions of the TA muscles were performed, each separated by a 60-sec rest period. Maximal (isometric) force was measured 1 min after each lengthening contraction and expressed as a percentage of the initial maximal (isometric) force. Following force measurements, animals were sacrificed by cervical dislocation and muscles were dissected, weighed, and frozen in liquid nitrogen or in isopentane precooled in liquid nitrogen. Samples were stored at −80°C for histological analyses.
Reverse-transcription PCR
Snap frozen muscle tissue was lysed with MagNa Lyser beads and instrument (Roche). Total RNA was subsequently isolated using the Nucleospin RNAII kit (Machery-Nagel) according to the manufacturer's protocol. RNA concentration and quality/integrity was checked with Nanodrop (Thermo-Scientific) and Lab-on-Chip (RNA 6000 Nano LabChip kit, Agilent) respectively. Next, cDNA was generated from 100 ng total RNA using Bioscript reverse transcriptase enzyme (Bioline) and random hexamer primers. Nested PCR was performed with primers detecting both full-length dystrophin transcript and the skipped product, as previously described (Spitali et al., 2010).
Histological and immunofluorescence analysis
TA muscles were sectioned at 10 μm on a cryostat (Leica CM3050S). Frozen sections were cut from the midbelly region of each muscle and stained with hematoxylin and eosin. For immunostaining, frozen sections were rinsed once with PBS, and then blocked during 2 hr with a buffer containing 2% fetal bovine serum and 2% BSA (Sigma-Aldrich). Primary antibodies were applied overnight at 4°C. For morphometric studies, co-immunostaining was performed using rabbit anti-laminin (Dako) and rat anti-perlecan (Millipore) together with an antibody against pan-myosin heavy chain (A4-1025, DSHB) to visualize sarcolemma and muscle fibers. Regenerating fibers were visualized using antibodies against embryonic myosin heavy chain (F1.652, DSHB). Restoration of dystrophin expression was studied using rabbit anti-dystrophin antibodies (Thermo Fisher Scientific). After three washes in PBS, muscle sections were then incubated for 1 hr at room temperature with secondary antibodies: goat anti-rabbit Alexa 488 and/or goat anti-mouse Alexa 555 (Molecular Probes, Invitrogen), before being thoroughly washed with PBS and mounted with Fluoromount (Southern Biotech). Images were captured using a digital camera (Hamamatsu ORCA-AG) attached to a motorized fluorescence microscope (Zeiss AxioImager.Z1), and morphometric analyses were made on whole sections using the software MetaMorph 7.5 (Molecular Devices). The fiber diameter was calculated as caliper width, perpendicular to the longest chord of each myofiber, which was called minimal fiber size for simplicity.
Western Blot
Protein extracts were obtained from pooled tissue sections treated with 125 mM sucrose, 5 mM Tris-HCl pH 6.4, 6% of XT Tricine Running Buffer (Bio-Rad), 10% sodium dodecyl sulfate (SDS), 10% glycerol, 5% β-mercaptoethanol. The samples were purified with the Compat-Able™ Protein Assay preparation Reagent Set (Pierce) and the total protein concentration was determined with the BCA Protein Assay Kit (Pierce). Samples were denatured at 95°C for 5 min, and 50 μg of protein were loaded onto Criterion XT tris-acetate precast gel 3%–8% (Bio-Rad). Dystrophin protein was detected by probing the membrane with 1:50 dilutions of NCL-DYS2 primary monoclonal antibody (Novocastra), followed by incubation with a sheep anti-mouse secondary antibody (horseradish peroxidase conjugated; 1:15,000) and ECL Analysis System (Amersham). Actin (Sigma) was used as internal control. Western blot analysis for the total and phosphorylated ribosomal protein S6 was performed as previously published (Hulmi et al., 2012).
Statistics
For statistical analysis of experiments in TA muscles of the treated mdx mice, a two-way repeated measures analysis of variance (ANOVA) was applied to account for the effect of AAV-U7 (high vs. diluted) and the effect of the sActRIIB-Fc (vs. PBS) measured in the same animals. Addition of sex as a factor to the ANOVA analysis did not reveal statistical differences between the different sexes, and sex was therefore not included in the final analysis (data not shown). For statistical analysis of fiber diameter, we compared the mean minimal fiber diameter of individual muscles. To compare the effect of sActRIIB on body weight development, we calculated weight increase of individual mice by subtracting initial body weight (measured before the first injection) from body weight at each time point. Two-way repeated measures ANOVA to compare the effect of sActRIIB-Fc on body weight were followed by Bonferroni's post hoc test to determine significant differences between groups at individual time points. Weights of muscles other than TA (not treated with AAV-U7) were compared using Student's t-test. Results were expressed as the mean together with the mean standard error (±SEM) or standard deviation (±SD) as stated in the figure legends.
Results
Bioactivity of sActRIIB-Fc in wild-type mice
We first tested the bioactivity of sActRIIB-Fc on skeletal muscle in vivo. We injected adult wild-type mice intraperitoneally with either sActRIIB-Fc (10 mg/kg) or PBS (control) at days 0, 4, 7, and 10 and monitored the effect on total animal weight and determined muscle wet weights 14 days after the first injection. sActRIIB-Fc induced a larger increase in body weight, which was evident already 2 days following the first injection when compared to control injected mice, and the maximal body weight increase was reached after 12 days (Fig. 1A). Two-way repeated measures ANOVA revealed a significant effect of sActRIIB-Fc as well as a significant effect of time on body weight increase (both p<0.001), however, Bonferroni's post hoc test failed to demonstrate differences between the two groups at individual time points. In addition, treatment with sActRIIB-Fc significantly increased muscle weights of gastrocnemius, quadriceps, and TA (Fig. 1B) compared to respective muscles from control mice. The mammalian target of rapamycin (mTOR)–S6 kinase pathway is an important regulator of muscle protein synthesis and myofiber size (Bodine et al., 2001; Zoncu et al., 2011) and is up-regulated following postdevelopmental myostatin depletion (Welle et al., 2011). To elucidate whether sActRIIB-Fc signals via the mTOR-S6 kinase pathway, we monitored the level of phospho-rpS6, a marker of S6 kinase activity, 1 and 2 days after a single injection with sActRIIB-Fc (10 mg/kg) or PBS. Whereas there was little effect on total rp-S6, we found a robust increase in phospho-rpS6 in gastrocnemius muscles from day 1 to day 2 following a single injection of sActRIIB-Fc compared to PBS-treated muscles (Fig. 1C). These experiments prove that our sActRIIB-Fc is bioactive and that it acts on skeletal muscle via the presumed molecular mechanism.
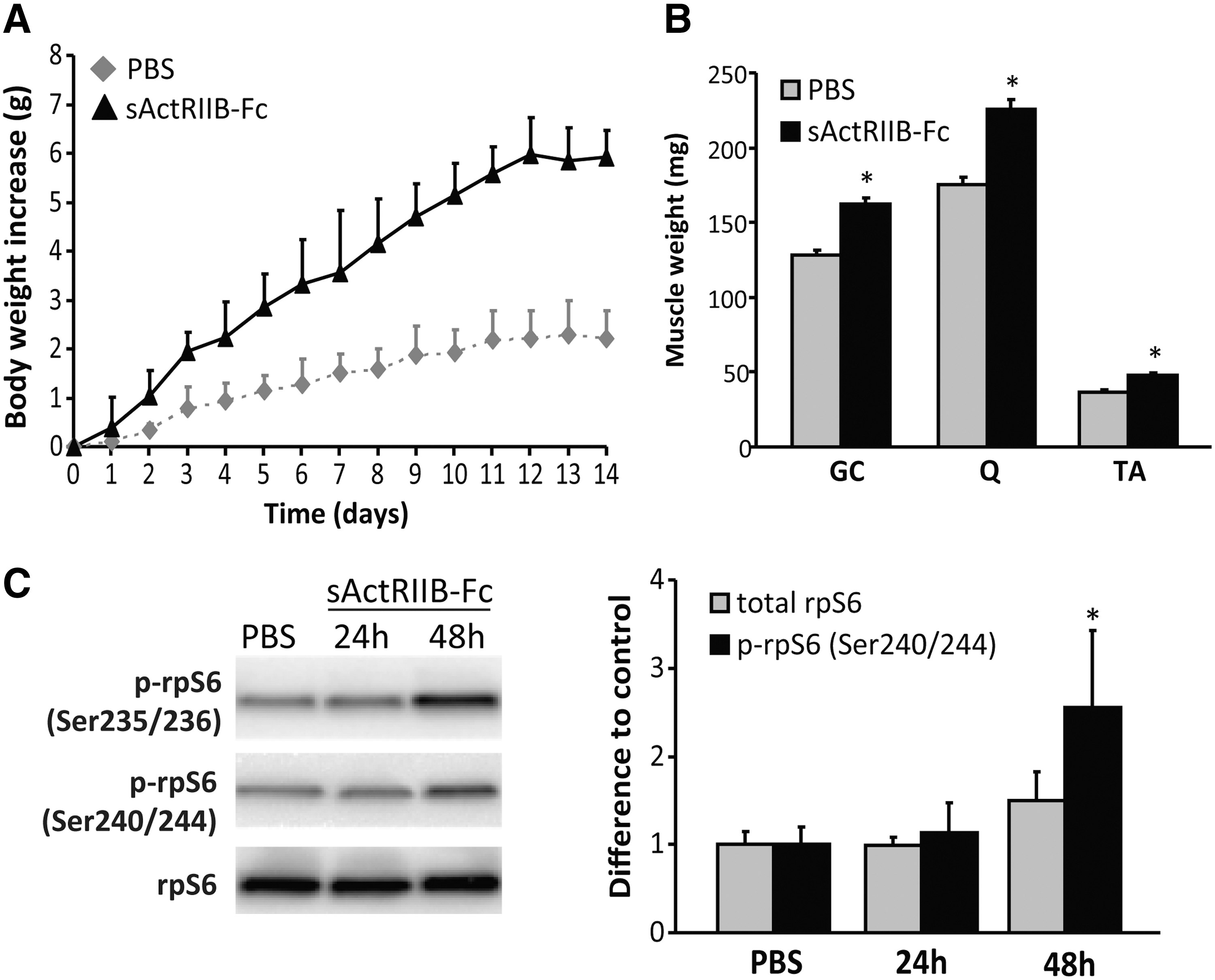
Effect of sActRIIB-Fc on muscle mass in wild-type mice.
Combined treatment in mdx mice
We next tested the effect of sActRIIB-Fc–mediated inhibition of ActRIIB signaling on dystrophic mdx muscle and explored the combined effect when applying together with AAV-U7–induced dystrophin exon skipping. Single strand AAV-vectors require about 2 weeks for initiating transgene expression, whereas treatment with recombinant proteins such as sActRIIB-Fc have no delayed effects. Since we aimed to simultaneously treat mdx mice with AAV-U7 and sActRIIB-Fc, we injected AAV-U7 2 weeks prior initiation of sActRIIB-Fc treatment (Fig. 2A). A total of 12 young adult mdx mice (aged of 6 weeks at the beginning of treatment) were included in this study. All mice were intramuscularly injected with high titer AAV-U7 (2.5×1011 vg/injection; labeled as AAV-U7-high) into the left TA muscles and with a similar volume of 10×diluted AAV-U7 into the right TA muscles (labeled as AAV-U7-diluted), which served as a control since dystrophin protein expression was below detection limit (see below, Fig. 2C). Two weeks after AAV injection, mice were assigned randomly to two different treatment groups; one group was intraperitoneally injected with 20 mg/kg/week sActRIIB-Fc (n=6 consisting of three males and three females) and the second group was treated with an equal volume of PBS (n=6 consisting of three males and three females) for a total of 5 weeks. This established four different treatment schedules for the mdx TA muscles: AAV-U7-highPBS, AAV-U7-dilutedPBS, AAV-U7-highsActRIIB-Fc, and AAV-U7-dilutedsActRIIB-Fc.
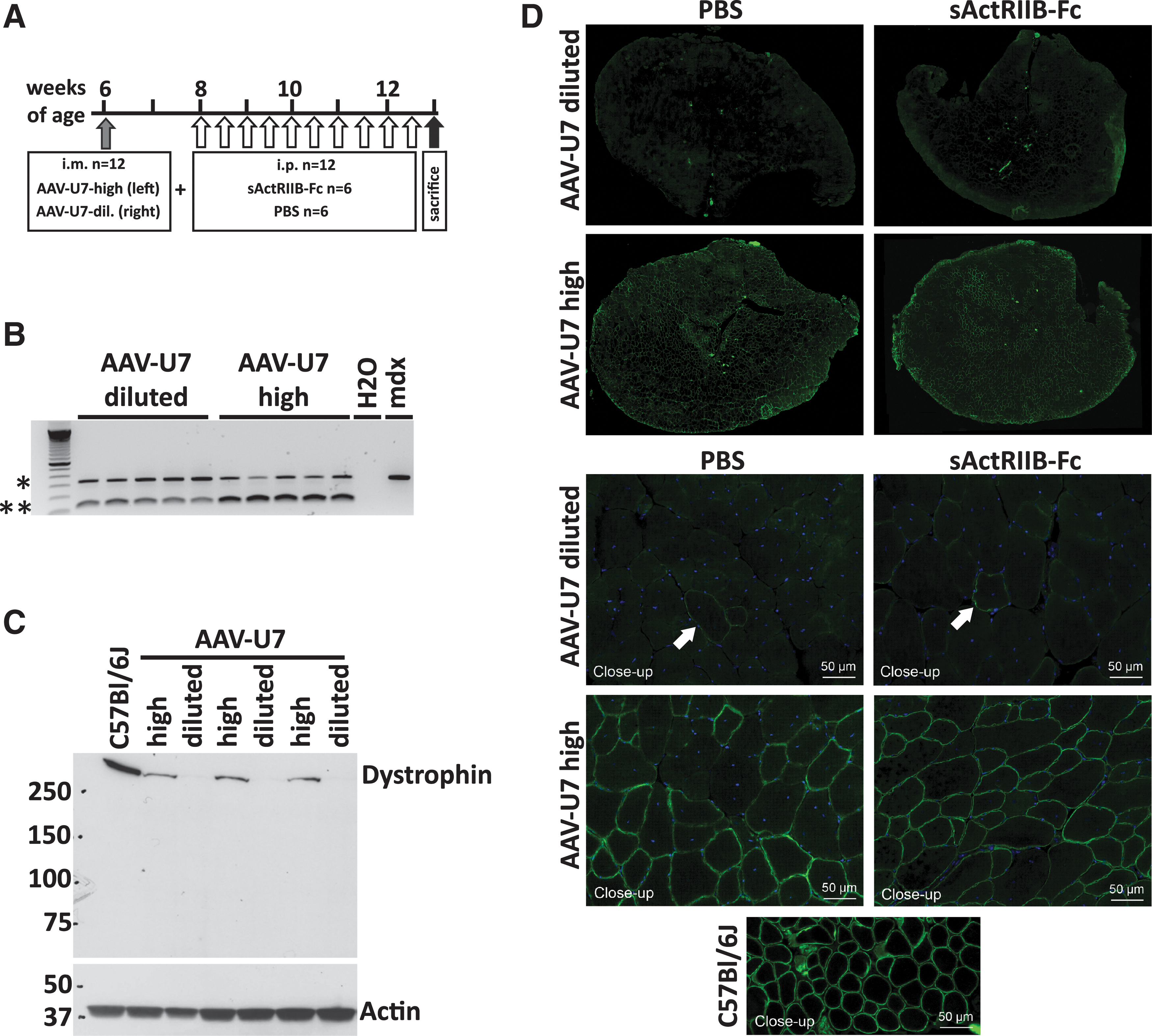
Effect of AAV-U7-high and AAV-U7-diluted (control) on exon skipping and restoration of dystrophin expression.
Restoration of dystrophin expression
We first qualitatively analyzed the ability of high titer AAV-U7 and diluted AAV-U7 (control) to transduce muscle and skip exon-23 of the dystrophin gene by using nested reverse-transcription (RT)-PCR. Intramuscular injection of mdx TA muscles with AAV-U7-high and also with AAV-U7-diluted resulted in partial skipping of exon-23 of the DMD gene of the injected muscles as evidenced by the presence of two bands representing the normal mRNA species and a shorter exon-23–skipped mRNA fragment (Fig. 2B). This demonstrated the bioactivity of AAV-U7-high and AAV-diluted vectors; however, nested RT-PCR is an insufficient method to quantify exon-skipping efficiency (Spitali et al., 2010). Importantly, only high titer AAV-U7, but not diluted AAV-U7, restored dystrophin protein expression to detectable levels (Fig. 2C, 2D). Immunohistochemistry revealed a mosaic of dystrophin expressing muscle fibers following transduction with AAV-U7-high, whereas only sporadic dystrophin positive fibers were observed in contralateral AAV-U7-diluted treated muscles (Fig. 2D). The AAV-U7-high–mediated dystrophin rescue was confirmed by Western blot (Fig. 2C) but was considerably lower as compared with the dystrophin expression of wild-type control muscle. AAV-U7-diluted–treated muscles revealed no detectable dystrophin expression in three out of six injected TA muscles and a very faint band of expression in the other three muscles (Fig. 2C). Importantly, the quantity of dystrophin in TA muscles following AAV-U7-high transduction was similar between mouse groups that were systemically treated with sActRIIB-Fc and with PBS (Supplementary Fig. S1; Supplementary Data are available at
Body mass and muscle size development
Treatment with sActRIIB-Fc rapidly increased total body weight as soon as 7 days following the first injection and remained stable thereafter (Fig. 3A). sActRIIB-Fc–treated mdx mice were always heavier than PBS-treated control mice; however, there were some fluctuations in body weight throughout the 5-week treatment protocol. The mean weight increase at the end of the treatment period compared to the start was 18% for sActRIIB-Fc–treated mice and 5% for PBS-treated control mice. Two-way repeated measures ANOVA revealed a significant effect of sActRIIB-Fc as well as a significant effect of time on body weight increase (both p<0.001); however, Bonferroni's post hoc test failed to demonstrate differences between the two groups at individual time points. The body weight increase following treatment with sActRIIB-Fc was accompanied by a gain in skeletal muscle mass to various extents (Fig. 3B, C), however, not for all muscles at significant levels (Fig. 3C). Importantly, treatment with sActRIIB-Fc increased weights of TA muscles to similar extents, irrespective of whether muscles were treated with AAV-U7-high or with AAV-U7-diluted. TA muscles treated with AAV-U7-dilutedsActRIIB-Fc gained 19% in weight compared with AAV-U7-dilutedPBS muscles (Fig. 3D). Similarly, AAV-U7-highsActRIIB-Fc–treated TA muscles gained 20% in weight compared with AAV-U7-highPBS–treated TA muscles (Fig. 3D). Morphometric analysis revealed a significant shift towards larger fiber diameters for both AAV-U7-dilutedsActRIIB-Fc– and AAV-U7-highsActRIIB-Fc–treated TA muscles compared with respective controls, suggesting that muscle mass increased from fiber hypertrophy following treatment with sActRIIB-Fc (Fig. 3E, F). These results demonstrate that the positive effect on muscle mass following treatment with sActRIIB-Fc was maintained in combination with AAV-U7-high–mediated exon skipping. Interestingly, we noted that treatment with AAV-U7-high caused a modest and nonsignificant trend towards lower muscle weights and this modest decrease occurred in presence as well as in the absence of an additional treatment with sActRIIB-Fc.
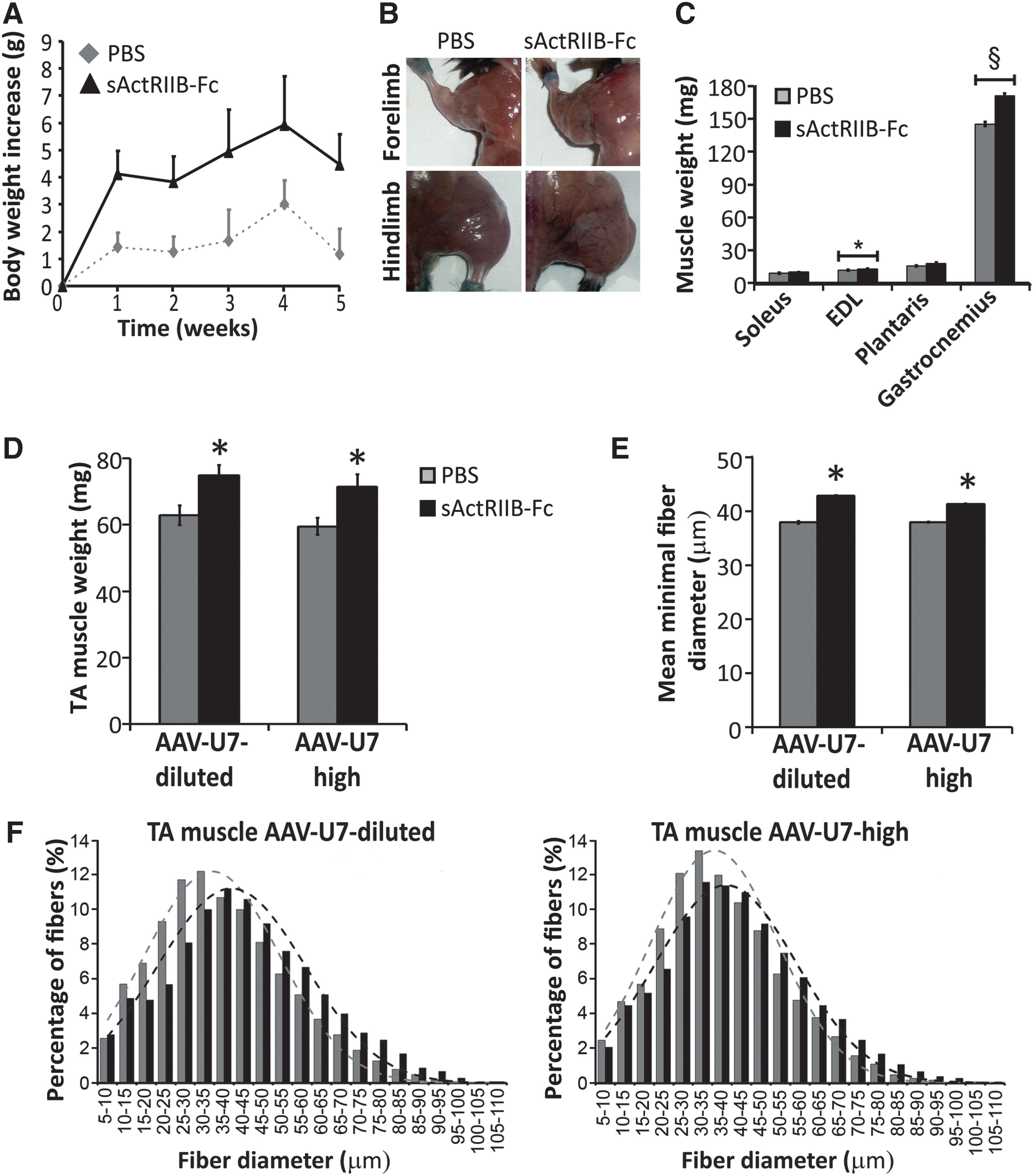
Body mass and muscle size development following systemic treatment with sActRIIB-Fc.
Effect on muscle histopathology
We next determined the effect of AAV-U7 and sActRIIB-Fc on the histopathology of dystrophic mdx muscle. Hematoxylin and eosin staining revealed histological features consistent with previous descriptions of the mdx phenotype such as strong variation in myofiber sizes, presence of many fibers with centrally located nuclei, hypercontractive fibers, degenerating fibers, small regenerating fibers, and increased endomysial fibrosis. There were no obvious differences in these histopathological features between the different treatment groups and some typical images are presented (Fig. 4A). Generally, degenerating muscles fibers were not very frequent and were mostly isolated, and clusters of degenerating fibers were extremely rare. Quantitative analysis revealed a considerable interindividual variation in the number of degenerating fibers as well as a high variation between left and right legs, which ranged between 2 and 42 degenerating fibers per TA muscle cross section. Because of these high variations, no differences were observed between the different treatment groups (data not shown). Mdx mice undergo substantial degeneration and regeneration between 3 and 5 weeks of age, which is reflected by a high number of fibers with centrally located myonuclei at young adult age. As we here treated young adult mdx mice, we considered central nucleation as an unsuitable marker for changes in regeneration activity. We therefore quantified the number of fibers expressing embryonic myosin heavy chain (MHCemb), because regenerating fibers only transiently express this MHC isoform, thus their appearance fell into the treatment period. Again, we found a high variation in the number of MHCemb positive regenerating fibers between different individuals and between left and right muscles, and no differences could be revealed between the different treatment groups (see example image in Fig. 4B).
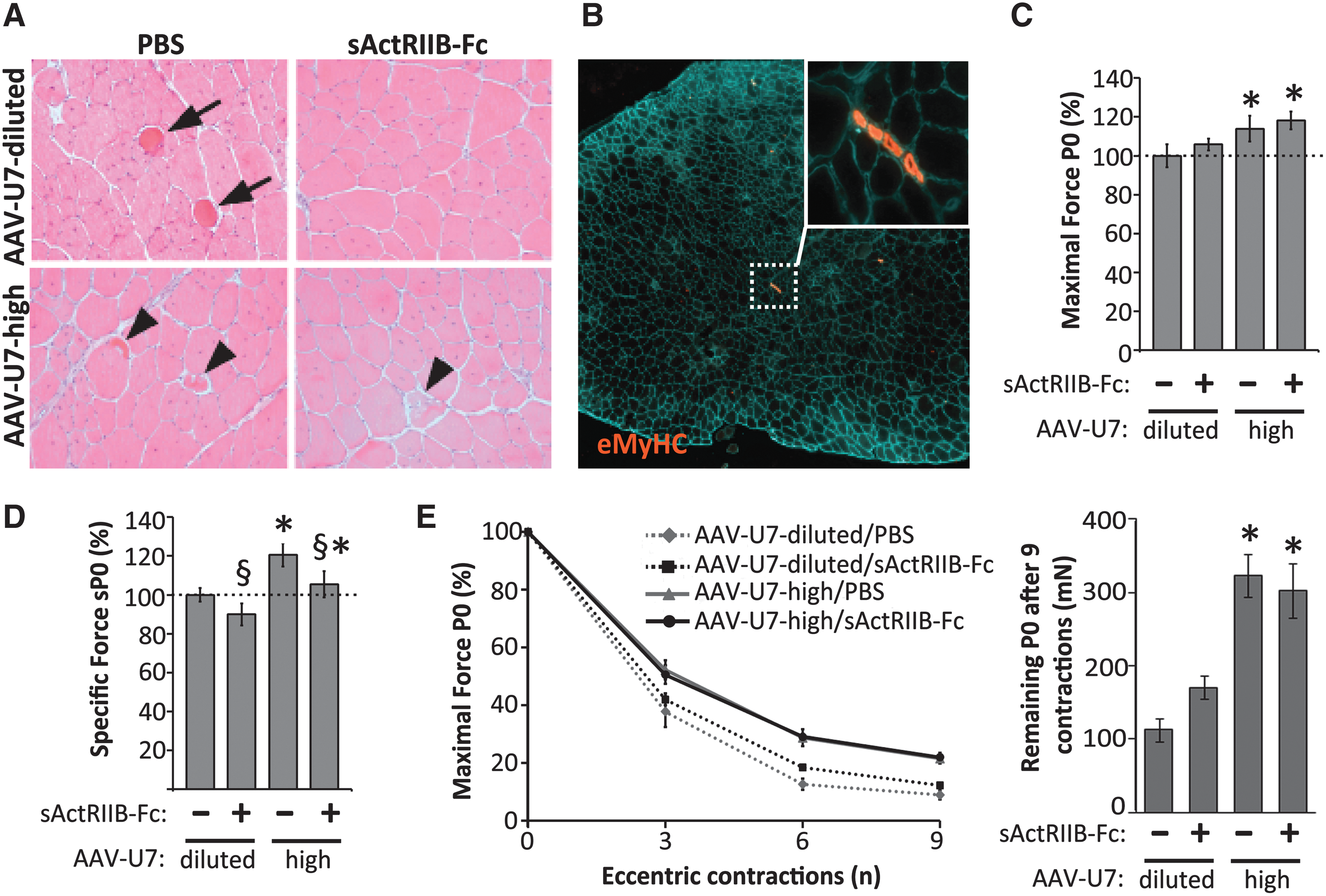
Effect of AAV-U7 and sActRIIB-Fc on muscle histology and function.
Effect on force generation capacity and vulnerability to contraction-induced muscle damage
Next, we determined the effect of the different treatment schedules on the force generation of TA muscles in situ. The data corresponding of the different experimental groups were expressed as percentage of AAV-U7-dilutedPBS. Using a two-way repeated measures ANOVA, we compared the effect of AAV-U7-high versus AAV-U7-diluted and the effect of sActRIIB-Fc versus PBS. Treatment with AAV-U7-highPBS increased absolute maximal force to 114%, which was significant in comparison to AAV-U7-dilutedPBS (p=0.02; Fig. 4C), and robustly increased specific maximal force (absolute maximal force normalized to muscle weight) to 120% (p=0.004; Fig. 4D). Furthermore, AAV-mediated restoration of dystrophin expression improved muscle resistance to contraction-induced damage, which is an indicator of the structural integrity of muscle fibers. This was shown by a reduced force drop following nine eccentric contractions in AAV-U7-highPBS–treated muscles compared to AAV-U7-dilutedPBS (p<0.001; Fig. 4E). We next studied the effect of single treatment with sActRIIB-Fc. Treatment with AAV-U7-dilutedsActRIIB-Fc hardly changed absolute maximal force as compared with AAV-U7-dilutedPBS (p=0.35; Fig. 4C), however specific force decreased to 90% (p=0.036; Fig. 4D). Resistance to eccentric contractions-induced damage remained unchanged following treatment with sActRIIB-Fc (p=0.47; Fig. 4E). We finally determined the combined effect of AAV-U7-high and sActRIIB-Fc on TA muscle function. Combined treatment increased absolute maximal force to 118% (p=0.02; Fig. 4C) and this gain in force was similar as AAV-U7-high on its own (114%; see above, no combined effect; p=0.88). It is important to note that combined treatment prevented the improvement of specific maximal force compared to the treatment with AAV-U7-high alone (p=0.036, Fig. 4D). Combined treatment also improved muscle resistance to eccentric contraction; however, there was no additive effect on top of the effect that AAV-U7-high achieved on its own (p=0.14, Fig. 4E).
Discussion
We here investigated for the first time a therapeutic strategy for DMD using sActRIIB-Fc to block Mstn and homologous growth factors in combination with AAV-U7–mediated dystrophin exon-skipping. Our results demonstrate that treatment with sActRIIB-Fc alone stimulates growth of both wild-type and mdx muscle. We also show that treatment with AAV-U7 alone restores dystrophin expression and improves mdx muscle function. These observations are in good agreement with previously published data (Goyenvalle et al., 2004; Morine et al., 2010; Pistilli et al., 2011). The novelty of our work is that combined treatment brings together these positive effects, illustrating the potential value of such multidrug therapies for DMD to restore wasted muscle and muscle function, especially for patients at advanced disease stages.
Our results demonstrate that despite larger muscles following treatment with sActRIIB-Fc alone, specific maximal force decreased, opposing previous observations on the effect of this molecule on mdx muscle (Morine et al., 2010; Pistilli et al., 2011). We can only speculate about the nature for these different physiological responses following treatment with sActRIIB-Fc. One possible reason for the failure of force increase in our study could be the shorter treatment period during which skeletal muscle has not yet fully adapted to its larger size. In contrast to the effect of sActRIIB-Fc, we here showed that high titer AAV-U7 improved absolute maximal force and this effect was not enhanced following combination with sActRIIB-Fc.
An important hallmark of the muscle dysfunction of dystrophin-deficient mdx mice is the strongly decreased specific maximal force. We here showed that AAV-U7-high improved specific maximal force, further evidencing its therapeutic benefit, which is in accordance with previous data on the effect of dystrophin exon skipping (Goyenvalle et al., 2004). However, specific maximal force failed to increase when AAV-U7-high was combined with sActRIIB-Fc.
The massively decreased resistance to eccentric contractions is another hallmark of mdx pathology and its improvement is an important parameter to test the efficiency of therapeutic interventions (Blaauw et al., 2009, 2010; Deconinck et al., 1997; DelloRusso et al., 2001; Tinsley et al., 1996). As expected, a high dose of AAV-U7 improved the resistance to eccentric contractions, which also is in line with previous data on the effect of dystrophin exon skipping (Goyenvalle et al., 2004). Differently to AAV-U7, treatment with sActRIIB-Fc did not improve the force drop following eccentric contractions, similar to what was found previously (Pistilli et al., 2011), and combined treatment had no synergistic effect compared with the effect of AAV-U7 alone.
Dumonceaux et al. (2010) previously investigated a different combined strategy and used shRNA-mediated knockdown of ActRIIB together with U7-mediated exon skipping. Similar to our results, they presented improved maximal muscle forces following AAV-U7–mediated exon skipping. Differently to our study, ActRIIB knockdown strongly boosted this force increase, although it failed to induce muscle growth. It remains to be elucidated why different strategies of ActRIIB blockade results in different results when combined with dystrophin exon skipping.
In this study we used 10-fold diluted AAV-U7 as a control vector, because we favored a control that initiated to a low extent the identical molecular mechanism as AAV-U7-high, however, at a concentration that was ineffective to restore dystrophin expression. The AAV consisted of an AAV-1 capsid and AAV-2–based vector. Previous work of our laboratory using different AAV2/1 vectors including control vectors such as AAV-U7-scramble or AAV-muSeAP did not reveal widespread immune response in muscle apart from some local inflammation and regenerating fibers along the injection trajectory following intramuscular application (Amthor et al., 2009; Goyenvalle et al., 2004; Lorain et al., 2008; Pietri-Rouxel et al., 2010). However, AAV-1 capsid, which was used in our study, causes a humoral immune response against the AAV-1 itself, which makes subsequent injection inefficient, but no cytotoxic T-cell response (Lorain et al., 2008). Lorain et al. (2008) also showed that ascending AAV-1 titers (10×109 to 10×1011 vg) provoked the same magnitude of humoral immune response. This made diluted AAV-U7 a convenient control for our experiments.
It remains difficult to conclude from our study whether a combination of both therapeutic strategies could be beneficial for DMD patients. We here used AAV-U7 to restore dystrophin, which is far more potent to restore dystrophin expression than synthetic antisense oligonucleotides. However, AONs are far more advanced for clinical use, and sActRIIB-Fc could possibly boost the effect of AONs better than in combination with AAV-U7. In mdx mice, dystrophin restoration using exon skipping largely restores dystrophic muscle function and this may obscure positive effects of sActRIIB-Fc. However, this may be different for DMD patients. It is also important to consider that we here tested the effect of combined approach only on a single muscle and we lack knowledge about the effect on whole body function when both strategies are systemically applied. Lastly, it is well established that the dystrophic pathology is marginal in mdx mouse and cannot be compared with the progressive muscle dystrophy in DMD patients; therefore, future efforts should focus on determining the efficacy of combined treatment in other DMD animal models such as the more severely affected mdx/utrophin−/− mouse.
Footnotes
Acknowledgements
The authors would like to thank Annemieke Aartsma-Rus and Gert-Jan Ommen for their contribution and expert advice and Christophe Hourdé, Cécile Peccate, Guillaume Precigout, and Bernardo Oliveira for technical assistance. This work was funded by the Dutch Duchenne Parent Project (to W.M.H.H. and P.A.C.tH.), the Association Française contre les Myopathies, the Association Monégasque contre les Myopathies, the Parents Project France, Aktion Benni & Co, ANR-Blanc Androgluco, the Agence Française de Lutte contre le Dopage, MyoGrad International Graduate School for Myology (DRK 1631/1), and the Academy of Finland (Decision No. 137787).
Author Disclosure Statement
The authors state no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.