
Abstract
Clinical observations and epidemiological surveys indicated that the prevalence of hypertension and heart diseases is increased in cold regions or during winter. Cold exposure increased NADPH oxidase gp91 phox protein expression in heart, kidneys, and aorta in rats. The aim of this study was to investigate if RNA interference (RNAi) silencing of gp91 phox would attenuate cold-induced hypertension and cardiovascular and renal damage. The recombinant adeno-associated virus serotype 2 (AAV-2) vector carrying gp91 phox -shRNA (gp91-shRNA) was constructed for inhibiting gp91 phox protein expression in cold-exposed rats. Blood pressure (BP) was monitored using a telemetry system. BP was increased in the Control-shRNA and PBS groups within 1 week of exposure to moderate cold (5°C) and reached a plateau after 7 weeks. The cold-induced increase in BP was attenuated significantly by intravenous delivery of gp91-shRNA (1.25×1010 particles/rat, 0.5 mL). One single dose of gp91-shRNA controlled hypertension for up to 10 weeks. In addition, gp91-shRNA reversed cold-induced vascular dysfunction. gp91-shRNA abolished the cold-induced up-regulation of gp91 phox protein expression in heart, kidneys, and aorta, confirming effective silencing of gp91 phox . The cold-induced increases in NADPH oxidase activity and superoxide production were eliminated by silencing of gp91 phox , suggesting that the cold-induced up-regulation of NADPH oxidase activity may be attributed to the increased gp91 phox protein expression. RNAi silencing of gp91 phox abolished cold-induced cardiac and renal hypertrophy and attenuated aortic, coronary, and renal remodeling. The up-regulation of gp91 phox may play a critical role in cold-induced cardiovascular dysfunction and organ damage. AAV delivery of gp91-shRNA may be a new and effective therapeutic approach for cold-related cardiovascular disorders.
Introduction
Our previous studies showed that the sympathetic nervous system activates the rennin–angiotensin system (RAS), which plays a role in cold-induced elevation of BP (Sun et al., 1995, 1997, 2003, 2005; Sun, 2010). Inhibition of the sympathetic nervous system or the RAS by pharmacological blockades or by knockout of angiotensinogen or AT1 receptor genes reduced cold-induced elevation of BP, but failed to attenuate cold-induced cardiac hypertrophy (Sun et al., 1995, 1997, 2003, 2005; Sun, 2010). Therefore, cold-induced CV dysfunction is not fully understood. Our preliminary studies showed that gp91 phox (Nox2) protein expression is up-regulated in heart, kidneys, and arteries. It is not clear if gp91 phox is involved in cold-induced CV dysfunction.
Recent studies suggest that the hypertrophic response is mediated by increased oxidative stress (Takemoto et al., 2001; Sawyer et al., 2002). NADPH oxidase–derived reactive oxygen species is involved in endothelial dysfunction, organ hypertrophy, and fibrosis, which are seen in hypertension and diabetes (Levy et al., 1990; Paravicini and Touyz, 2008). The NADPH oxidase is the primary source of superoxide in blood vessels and kidneys (Görlach et al., 2000; Griendling et al., 2000; Touyz and Schiffrin, 2004; Tain and Baylis, 2006). Although Nox1 and Nox4 are found in vasculature, the gp91
phox
containing NADPH oxidase (Nox2) selectively expressed in endothelial cells is a major source of oxygen radical generation in the arterial wall (Görlach et al., 2000). Our preliminary study indicated that cold exposure up-regulated gp91
phox
protein expression, but did not alter Nox1 and Nox4 expression in arteries (Supplementary Fig. S1; Supplementary Data are available online at
The most commonly used Nox inhibitor is the iodonium derivative diphenylene iodonium (DPI). The other Nox inhibitor is apocynin. However, both DPI and apocynin are nonselective Nox inhibitors. There currently is no gp91 phox -specific inhibitor. RNA interference (RNAi) is a powerful tool for inhibiting a target protein expression. Therefore, we designed short hairpin RNA (shRNA) carried by adeno-associated virus serotype 2 (AAV-2) for in vivo specific inhibition of gp91 phox expression. Our recent studies showed that shRNA carried by the AAV-2 vector is an effective approach for long-term inhibition of protein expression in animal models (Wang et al., 2006; Crosswhite and Sun, 2010).
The purpose of this study was to investigate if the up-regulation of gp91 phox plays a role in cold-induced CV dysfunction by RNAi silencing of gp91 phox expression.
Materials and Methods
Construction of recombinant AAV with gp91 phox -shRNA
Three shRNA sequences specifically targeting on gp91 phox (accession no. AF298656) were designed using Dharmacon software and synthesized by Integrated DNA Technologies (Coralville, IA). The inhibition efficiency of the three gp91 phox -shRNA sequences was tested in H9C2 cells.
The gp91 phox -shRNA sequence that achieved the greatest inhibition of gp91 phox expression was chosen for the in vivo study. This sequence was designed to target on rat gp91 phox at 5’-CCA TCG AGC TTC AGA TGA A-3’ (nucleotides 842–860). This gp91 phox -shRNA was then constructed into AAV-2 vector (Stratagene, La Jolla, CA) under the control of RNA polymerase III promoter U6 (AAV.U6-gp91-shRNA), as described in our recent studies (Wang et al., 2006; Crosswhite and Sun, 2010). AAV.U6-gp91-shRNA plasmid DNA was then packaged with pHelper and pAAV-RC to produce recombinant AAV-gp91-shRNA. The titer was determined by real-time PCR as described by Rohr et al. (2002). AAV-2 with a scrambled shRNA sequence was also constructed and served as a control (AAV.U6-Control-shRNA). The scrambled shRNA has been tested by BD Bioscience (Palo Alto, CA) and found not to match with any known gene sequences.
Animal study protocols
This study was carried out according to the guidelines of the National Institutes of Health on the care and use of laboratory animals. The project was approved by the Institutional Animal Care and Use Committee. Four groups of Sprague–Dawley rats (220–250 g, 6 rats/group) were used in this experiment. All rats were housed individually and provided with Purina Laboratory Chow (no. 5001) and tap water.
All animals were implanted with BP devices (aortic cannulation) under anesthesia [sodium pentobarbital 65 mg/kg, intraperitoneally (IP)]. Following a 1-week recovery, three groups of animals were exposed to a moderate cold environment (5±0.2°C) continuously, as we described recently (Crosswhite and Sun, 2010), whereas the remaining group was kept at room temperature (25±2°C) and served as a control. The three cold-exposed groups of rats received intravenous (IV) delivery of AAV-gp91-shRNA (1.25×1010 particles/rat, 0.5 mL), AAV-Control-shRNA (1.25×1010 particles/rat, 0.5 mL), and phosphate-buffered solution (PBS; 0.5 mL/rat), respectively, 3 days before exposure to cold. The group kept at room temperature (warm) received PBS (0.5 mL/rat) and served as a control (PBS-Warm).
BP measurement
BP was monitored continuously using a telemetry system (DSI, St. Paul, MN) as described in our recent study (Wang and Sun, 2010). Body weight was measured weekly.
Hypotensive response to acetylcholine (Ach) and sodium nitroprusside (SNP)
Hypotensive responses to Ach and SNP (Sigma-Aldrich, St. Louis, MO) were tested during week 10. The dose–response curve was generated for Ach (0, 1, 2.5, 10, 20, 150, 250, and 280 μg/kg, 0.25 mL, IV). The dose–response curve was also generated for SNP (0, 10, 20, 50, and 100 μg/kg, 0.25 mL, IV). A sufficient time (10 min) was allowed between doses to ensure a full recovery of BP.
Tissue preparations
At the end of week 10, all animals were euthanized (sodium pentobarbital, 100 mg/kg, IP) and perfused transcardially with heparinized saline. Following perfusion, heart, kidneys, and aorta were removed. Heart and kidney were weighed. One part of heart, kidney, and aorta was saved for quantification of gp91 phox protein expression by western blot analysis. Small parts of these organs were excised rapidly and embedded with O.C.T for assessing in situ superoxide production. Some animals were perfused with 4% paraformaldehyde, and tissues were fixed in 4% paraformaldehyde overnight at 4°C for histological examination and trichrome staining (collagen assay).
Western blot analysis
The procedure for western blotting was described previously (Wang et al., 2006; Wang and Sun, 2009). In brief, heart, kidney, and aorta were homogenized and lysed with cell lysis buffer (10 mM Tris-Cl, 150 mM NaCl, 1 mM EDTA, 0.1% NP40, and 1% proteinase inhibitor cocktail) on ice for 30 min. An equal amount of protein was loaded in 10% Tris-HCl gel (Bio-Rad, Hercules, CA) followed by electronic transfer. After blocking with 5% milk (in Tris-buffered saline with Tween-20), the membranes were incubated with antibody against Nox1, gp91 phox (Nox2), and Nox4 (1:1,000; BD Transduction Laboratories Inc., Franklin Lakes, NJ) at 4°C overnight. The membrane was then incubated with horseradish peroxidase–conjugated secondary anti-mouse antibody (1:2,000) for 1 hr at room temperature. Proteins were visualized by ECL (Amersham, Little Chalfont, Buckinghamshire, UK), exposed to an X-ray film, and developed with an X-ray processor (Canon SRA-101A).
Evaluation of in situ superoxide level
Dihydroethidium (DHE) (Sigma-Aldrich) fluorescence dye can enter the cells and be oxidized by superoxide to yield ethidium, which binds to DNA to produce bright red fluorescence. Unfixed heart, kidney, and aorta were embedded in O.C.T., frozen at −80°C, and sectioned at 10 μm using a cryostat (Microm International Model HM505 E; Microm International GmbH, Walldorf, Germany). Sections were incubated in a humidified chamber (37°C) for 30 min with HEPES-buffered solution (130 mM NaCl, 5 mM KCl, 1.2 mM MgCl2, 10 mM glucose, and 10 mM HEPES, pH 7.3), and then incubated for an additional 30 min in HEPES-buffered solution containing DHE (10–5 M). The sections were counterstained with the nuclear stain 4,6-diamidin-2-phenylindol dichlorohydrate (DAPI; 3×10–7 M) for 5 min at 37°C. Five sections of each group were examined, and the signal was captured with TRITC and UV filters using Leica TCS NT Confocal microscopy at 400× magnification.
Analysis of the collagen level
The paraffin-embedded sections (5 μm) of heart, kidneys, and aorta were used for quantifying connective tissue fibrosis and collagen deposition using trichrome stains (Sigma-Aldrich) as described in our recent study (Wang and Sun, 2009). The collagen level was semiquantified using NIH Imaging J software.
Measurement of NADPH oxidase activity by lucigenin-enhanced luminometer
NADPH oxidase activity was measured as we described recently (Wang and Sun, 2009). In brief, the tissue homogenate was centrifuged at 11,000 g, and the supernatent was mixed (1:1, vol/vol) with PBS containing 20 μM lucigenin (Sigma), before addition of NADPH (0.1 mM) (Sigma) as the substrate. The emitted luminescence was detected by luminometer (Gen5, BioTek, Winooski, VT). The data were expressed as fold changes versus the baseline level of NADPH oxidase activity (prior to the addition of NADPH).
Statistical analysis
Data were analyzed using a two-way ANOVA followed by a one-way ANOVA or an unpaired Student's t test. Significance was set at a 95% confidence limit.
Results
RNAi silencing of gp91 phox attenuated cold-induced elevation of BP and vascular dysfunction
Preliminary studies showed that cold exposure up-regulated vascular gp91 phox protein expression, but did not affect Nox1 or Nox4 protein expression (Supplementary Fig. S1). Therefore, we investigated if the up-regulation of gp91 phox is involved in cold-induced hypertension (CIH) CV and renal damage by RNAi silencing of gp91 phox expression.
Mean aortic BP was elevated significantly within 1 week of exposure to cold in the Control-shRNA and PBS groups (Fig. 1A). BP continued to increase at 3 weeks and reached a plateau after 5 weeks. In contrast, BP of the gp91-shRNA–treated group did not increase until week 2 after exposure to cold (Fig. 1A). The cold-induced increase in BP is significantly decreased in the gp-shRNA–treated group compared with the Control-shRNA and PBS groups (Fig. 1A). Thus, RNAi silencing of gp91 phox delayed and attenuated CIH, although it did not decrease BP to the level of the PBS-Warm group (Fig. 1A).
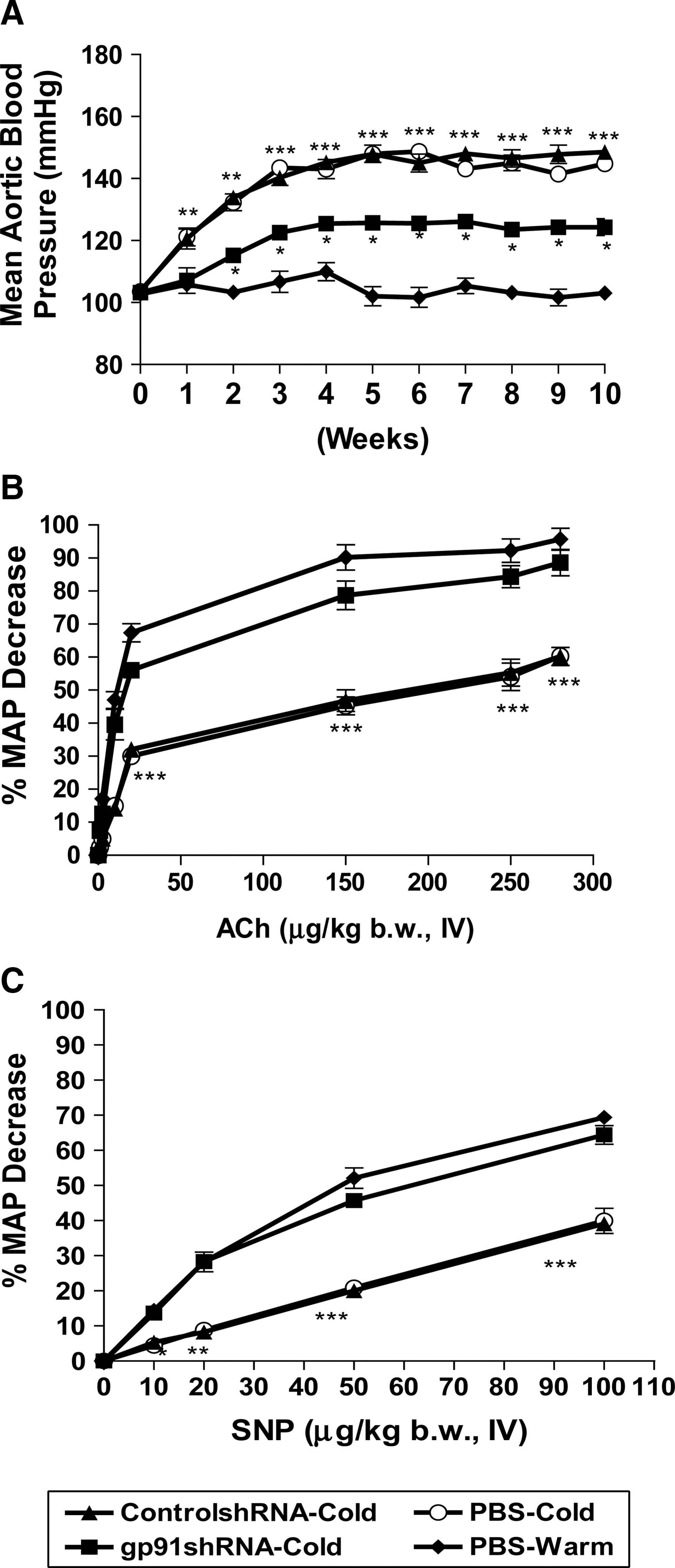
RNAi silencing of gp91
phox
attenuated cold-induced elevation of BP and vascular dysfunction. BP was monitored using a telemetry system.
Cold exposure significantly decreased the hypotensive responses to Ach and SNP (Fig. 1B and C), indicating vascular endothelial dysfunction and vascular smooth muscle dysfunction, respectively. RNAi silencing of gp91 phox significantly increased the hypotensive responses to Ach and SNP (Fig. 1B and C), suggesting improvements in cold-induced vascular endothelial dysfunction and smooth muscle dysfunction.
RNAi silencing of gp91 phox attenuated cold-induced cardiac and renal hypertrophy
Heart and kidney weights were increased significantly in cold-exposed rats, indicating that cold exposure caused cardiac and renal hypertrophy (Fig. 2). It is interesting that gp91-shRNA abolished cold-induced cardiac and renal hypertrophy (Fig. 2). Viral delivery did not affect the body weight gain (data not shown).
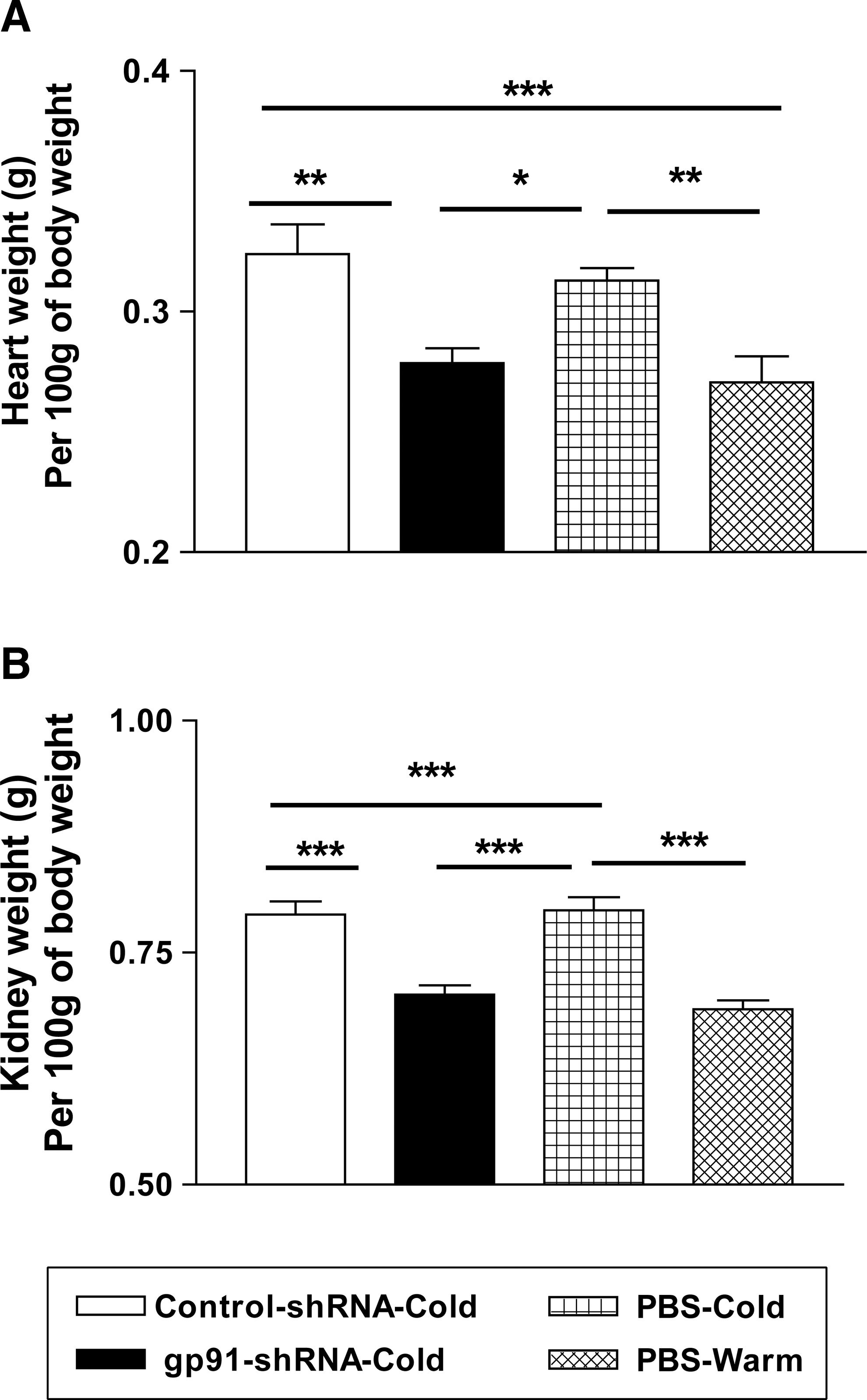
RNAi silencing of gp91
phox
attenuated cold-induced cardiac and renal hypertrophy.
RNAi silencing of gp91 phox attenuated cold-induced superoxide production
The in situ superoxide production was assessed using DHE staining (red fluorescence). The superoxide level was increased significantly in the heart (Fig. 3A and D), kidneys (Fig. 3B and E), and aorta (Fig. 3C and F) in cold-exposed groups versus that of the warm PBS group, indicating that cold exposure stimulated superoxide generation. gp91-shRNA prevented the cold-induced increase in superoxide levels (Fig. 3A–F).
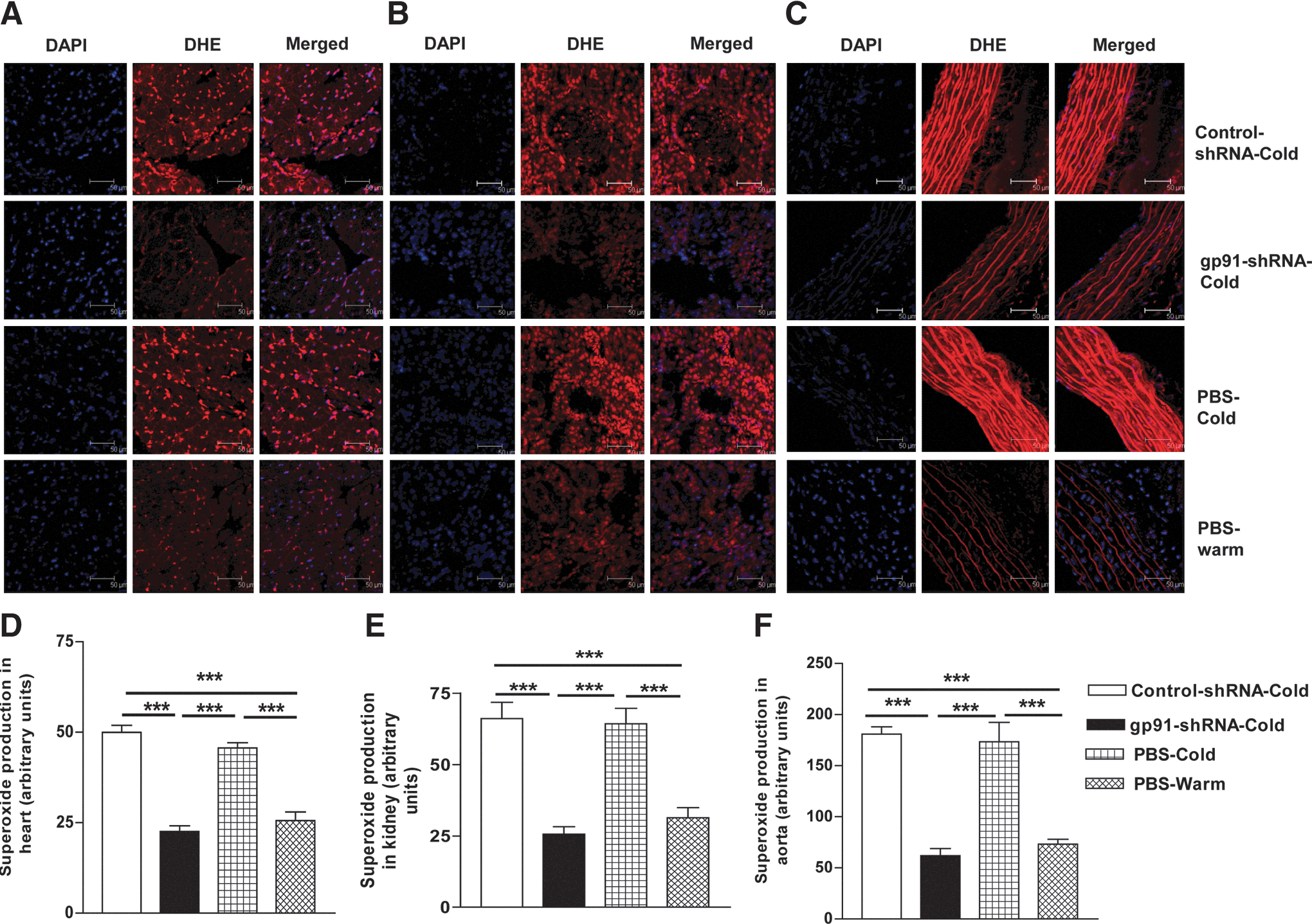
RNAi silencing of gp91
phox
attenuated cold-induced superoxide production. The frozen sections (10 μm) of heart
RNAi silencing of gp91 phox attenuated the cold-induced up-regulation of NADPH oxidase activity
NADPH activity of the heart (Fig. 4A), kidneys (Fig. 4B), and aorta (Fig. 4C), as assessed by lucigenin-enhanced chemiluminescence, was markedly increased significantly in cold-exposed rats. gp91-shRNA abolished the cold-induced up-regulation of NADPH oxidase activity.
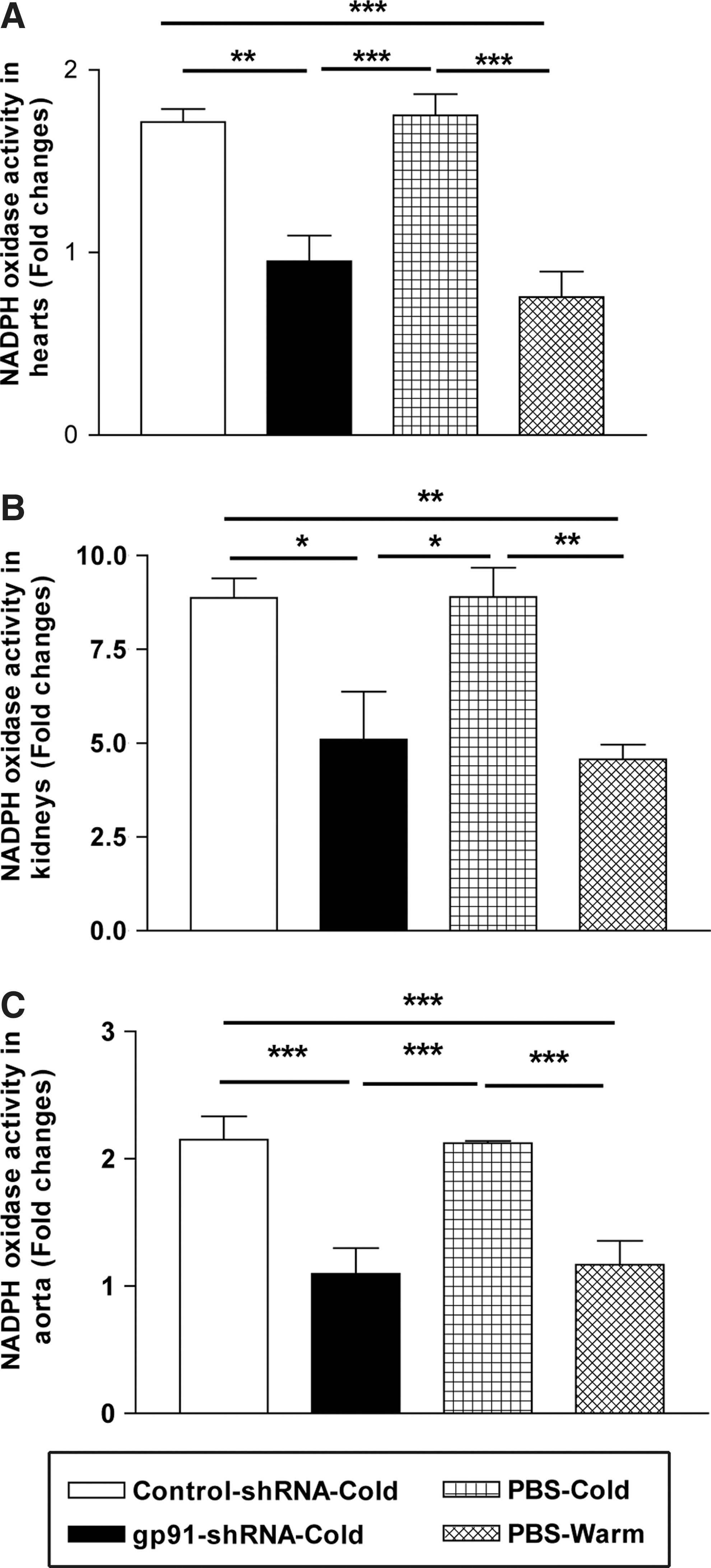
RNAi silencing of gp91
phox
attenuated the cold-induced up-regulation of NADPH oxidase activity. NADPH activity in heart
gp91-shRNA effectively silenced gp91 phox protein expression
Cold exposure increased gp91 phox protein expression in the left ventricle (Fig. 5A and B), kidneys (Fig. 5C and D), and abdominal aorta (Fig. 5E and F). gp91-shRNA abolished the cold-induced up-regulation in gp91 phox protein expression in these organs, confirming effective silencing of gp91 phox gene. It is noted that gp91-shRNA kept gp91 phox protein at, but not below, the control level (PBS-Warm) (Fig. 5A–F). Nox1 or Nox4 was not affected by cold exposure or gp91-shRNA (Supplementary Fig. S1). Nox5 was not measured, because it is not expressed in rodents (Schulz and Munzel, 2008). Manganese superoxide dismutase (Mn-SOD) protein expression was not affected by either cold exposure or gp91-shRNA (Supplementary Fig. S2). The AAV transfection rate was about 60–76% (Supplementary Fig. S3). The reporter genes [green fluorescent protein (GFP) and LacZ] were still expressed at the time of animal sacrifice, indicating that AAV is a long-term vector (Supplementary Figs. S3 and S4).
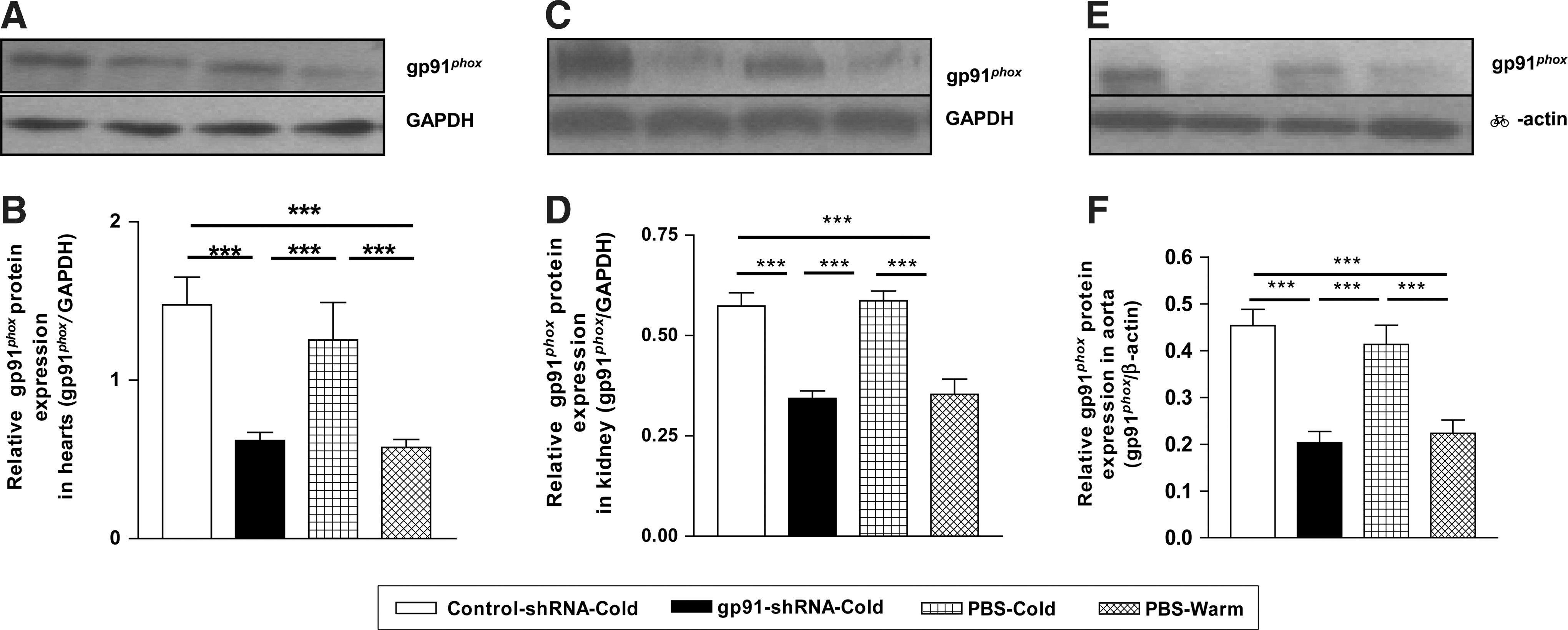
gp91-shRNA effectively silenced gp91
phox
protein expression.
RNAi silencing of gp91 phox attenuated cold-induced coronary artery hypertrophy and collagen deposition
Trichrome staining revealed collagen deposition around the intramyocardial coronary arteries. Cold exposure increased the medial wall thickness of coronary arteries and the collagen deposition (fibrosis formation) (Fig. 6), indicating coronary remodeling. RNAi silencing of gp91 phox prevented cold-induced coronary remodeling (Fig. 6).
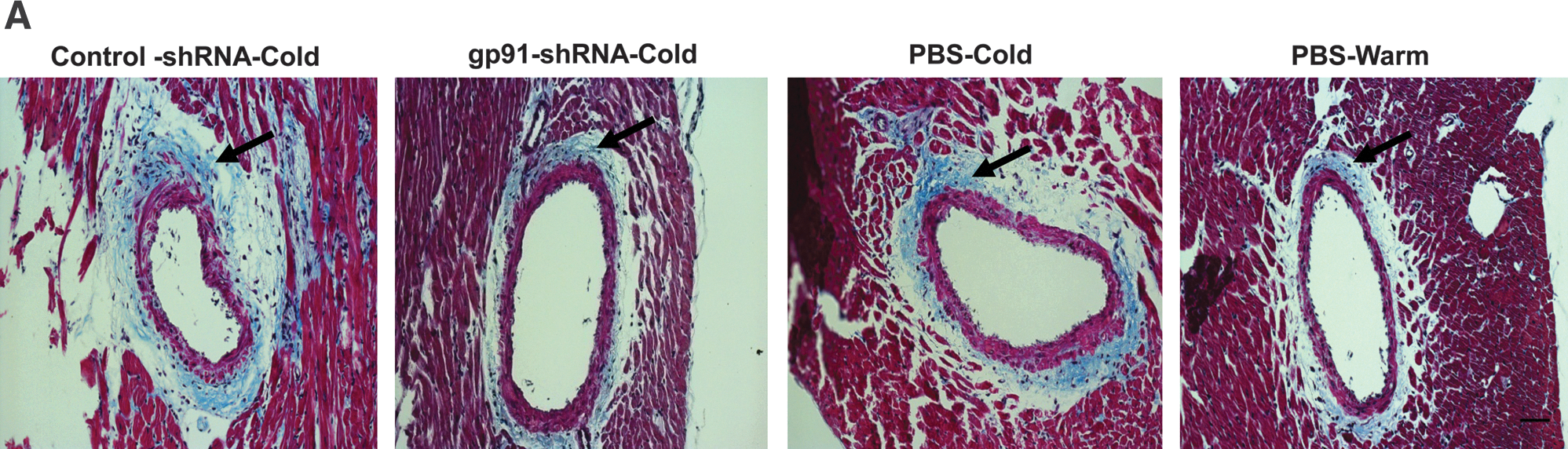
RNAi silencing of gp91
phox
attenuated the cold-induced coronary artery hypertrophy and collagen deposition.

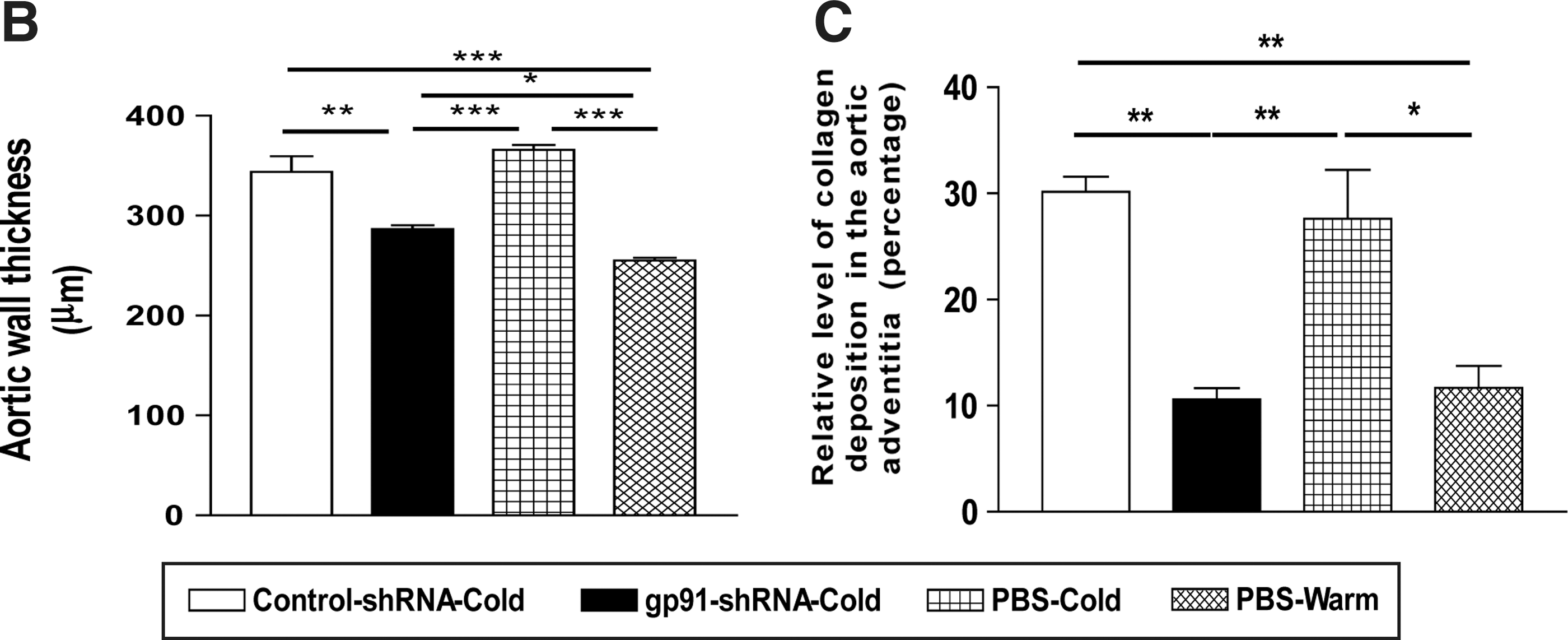
RNAi silencing of gp91 phox attenuated cold-induced aortic hypertrophy and collagen deposition
Cold exposure increased medial wall thickness of aorta and perivascular collagen deposition (Fig. 7). RNAi inhibition of gp91 phox abolished cold-induced aortic remodeling.
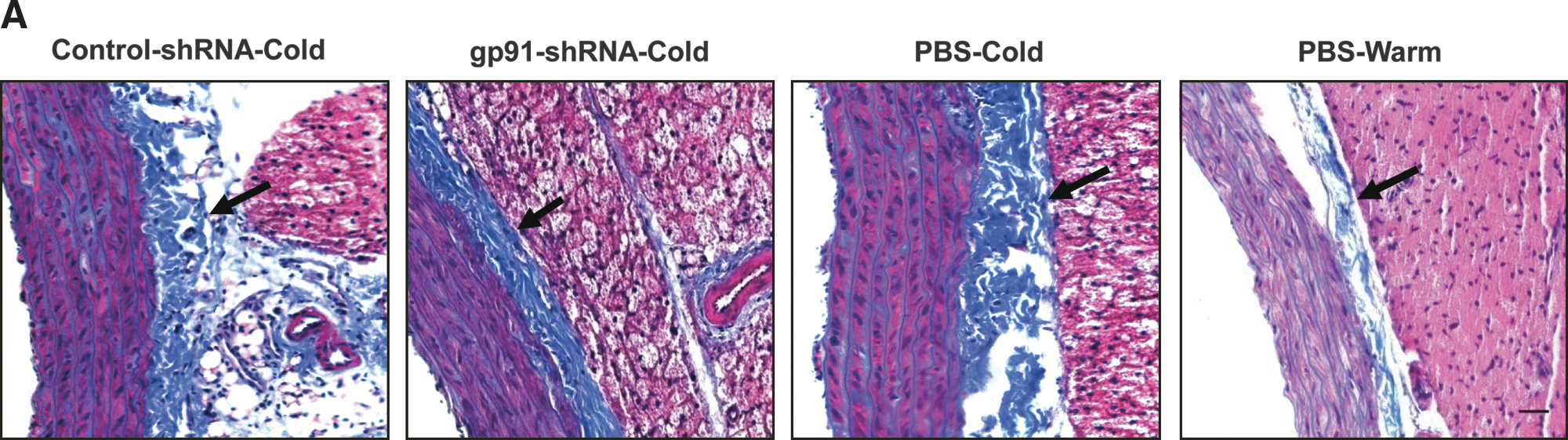
RNAi silencing of gp91
phox
attenuated the cold-induced aortic hypertrophy and collagen deposition.
RNAi silencing of gp91 phox attenuated cold-induced kidney damage
Representative histological images of kidneys are presented in Fig. 8A and B. Glomerular sclerosis (blue fibrotic tissue) and collapsed capillary structure were found in kidneys of cold-exposed rats, indicating severe glomerular injury (Fig. 8A and C). Renal tubular damage, including blocked tubules in the outer medulla (red protein deposition casts), was also found in cold-exposed rats (Fig. 8B and D), indicating impairment of tubular function. RNAi silencing of gp91 phox abolished cold-induced kidney damage and remodeling (Fig. 8A–D). Cold exposure significantly increased plasma creatinine, indicating functional damage of the kidneys (Fig. 8E). RNAi silencing of gp91 phox improved kidney function (Fig. 8E).
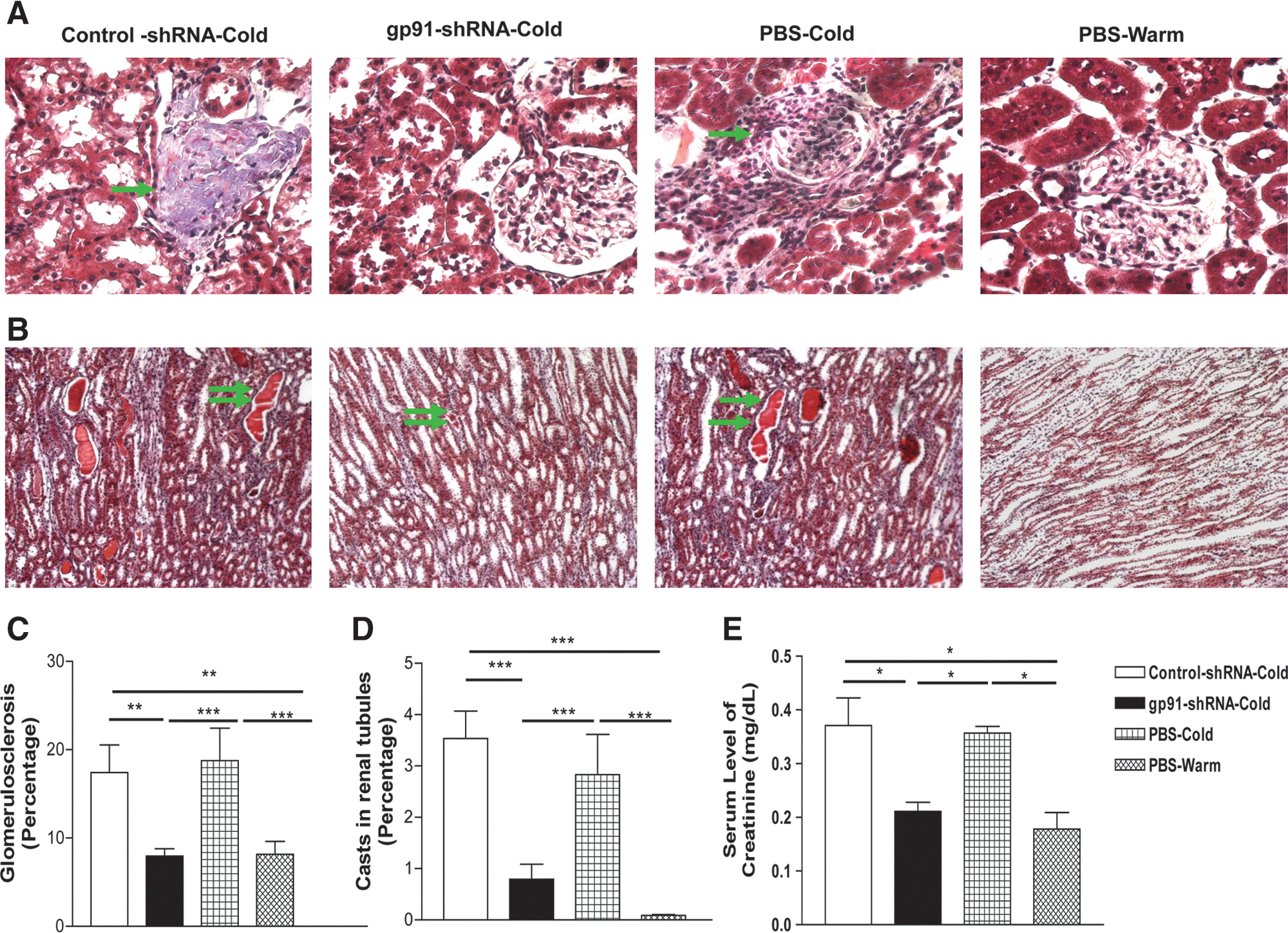
RNAi silencing of gp91
phox
attenuated the cold-induced kidney damage. Light microscopy images of renal cortex (20× original magnification;
Discussion
It is well established that the prevalence of hypertension and heart diseases is increased in cold regions or during winter (Baker-Blocker, 1982; Thorvaldsen et al., 1995; Gorjanc et al., 1999; Fu et al., 2002; Velázquez Monroy et al., 2002). The mortality and morbidity of CV disease is peaked in the winter (Baker-Blocker, 1982; Seretakis et al., 1997; Gorjanc et al., 1999; Crawford et al., 2003). Cold weather exacerbates hypertension (Hata et al., 1982; Verdon et al., 1993; Minami et al., 1996; Donaldson et al., 1997) and triggers myocardial infarction and stroke in hypertensive patients. Thus, cold temperatures have adverse effects on the human CV system. However, the mechanism of cold-induced CV dysfunction is not fully understood. We noticed that cold exposure up-regulated NADPH oxidase gp91 phox protein expression in aorta, heart, and kidneys (Fig. 5). RNAi silencing of gp91 phox gene effectively prevented the cold-induced up-regulation of gp91 phox protein expression, delayed and attenuated cold-induced elevation of BP, and abolished cold-induced vascular dysfunction and cardiac, vascular, and renal hypertrophy and remodeling (Figs. 1 –8). These findings demonstrate, for the first time, that up-regulation of NADPH oxidase gp91 phox plays a critical role in the pathogenesis of CIH, vascular dysfunction, and organ damage. Renal function was impaired in cold-exposed rats (Fig. 8E). RNAi silencing of gp91 phox significantly improved the renal function in cold-exposed animals (Fig. 8E). Because RNAi silencing of gp91 phox attenuated cold-induced cardiac remodeling, one would expect improved cardiac function in cold-exposed rats treated with gp91-shRNA. The limitation of this study is that the in vivo cardiac function was not assessed due to technical challenges.
It is noted that silencing of gp91 phox delayed and attenuated, but did not prevent, cold-induced elevation of BP (Fig. 1), suggesting that a mechanism other than the up-regulation of gp91 phox also contributes to CIH. In contrast, the up-regulation of gp91 phox may be responsible for cold-induced cardiac and renal hypertrophy, because they can be abolished by RNAi silencing of gp91 phox expression (Fig. 2). These data suggest that cold-induced cardiac and renal hypertrophy may be independent of CIH. On the other hand, prevention of the up-regulation of gp91 phox protein expression by gp91-shRNA abolished cold-induced cardiac, vascular, and renal remodeling although BP remains elevated. Therefore, up-regulation of gp91 phox is the key player in cold-induced organ damage and remodeling. It was reported that overproduction of superoxide (O2 −) leads to oxidative damage, fibrosis formation, and tissue remodeling (Levy et al., 1990; Sawyer et al., 2002; Paravicini and Touyz, 2008). Indeed, cold-induced up-regulation of gp91 phox is associated with increases in NADPH oxidase activity and O2 − generation (Figs. 3 and 4). It seems that the up-regulation of gp91 phox protein expression may account for the cold-induced increases in NADPH oxidase activity and O2 −generation, which can be abolished by silencing of gp91 phox . The alteration in O2 − levels may not be due to Mn-SOD, because it was not affected by either cold exposure or silencing of gp91 phox (Supplementary Fig. S2).
Nox2 (gp91 phox ) was first described in neutrophils and macrophages and is often referred to as the phagocyte NADPH oxidase. Recent studies indicated that gp91 phox is also expressed in nonphagocytic cells, including neurons (Serrano et al., 2003), cardiomyocytes (Heymes et al., 2003), skeletal muscle myocytes (Javesghani et al., 2002), hepatocytes (Reinehr et al., 2005), endothelial cells (Jones et al., 1996; Görlach et al., 2000; Li and Shah, 2002), and hematopoietic stem cells (Piccoli et al., 2005). The NADPH oxidase is the major source of superoxide in blood vessels and kidneys (Görlach et al., 2000; Griendling et al., 2000; Touyz and Schiffrin, 2004; Tain and Baylis, 2006). Increased superoxide levels inactivate the vasodilator nitric oxide leading to endothelial dysfunction and vasoconstriction, which are associated with several CV diseases, including hypertension (Touyz, 2004; Paravicini and Touyz, 2008). Cold exposure caused vascular endothelial and smooth muscle cell dysfunction as evidenced by disrupted hypotensive responses to Ach and SNP, respectively, in cold-exposed rats (Fig. 1). It appears that cold-induced vascular dysfunction is mainly mediated by the up-regulation of gp91 phox protein expression, because it can be restored by silencing of gp91 phox . Indeed, the cold-induced increases in vascular NADPH oxidase activity and superoxide generation were abolished by silencing of gp91 phox (Fig. 4).
It is interesting that up-regulation of a single NADPH oxidase gp91 phox plays a critical role in cold-induced CV dysfunction. The gp91 phox gene expression is inducible (Newburger et al., 1988; Nicotera et al., 1993; Krijnen et al., 2003). The gp91 phox expression was increased in phagocytes in response to interferon-γ (Newburger et al., 1988), in myofibroblasts after carotid artery injury (Nicotera et al., 1993), and in cardiomyocytes after acute myocardial infarction (Krijnen et al., 2003). The gp91 phox expression is also increased in response to angiotensin II in adipose tissue, aorta, resistance artery smooth muscle cells, and heart (López-Barneo et al., 1999; Touyz et al., 2002). The present study showed that cold exposure resulted in up-regulation of gp91 phox protein expression (Fig. 5). Our previous studies showed that cold exposure activates the RAS (Sun et al., 2003, 2005; Sun, 2010). However, the RAS may play a role in CIH, but not cold-induced cardiac and renal hypertrophy, because inhibition of the RAS reduces cold-induced elevation of BP, but does not attenuate organ hypertrophy (Sun et al., 1995, 1997, 2003, 2005; Sun, 2010). In contrast, the up-regulation of gp91 phox played a critical role in both CIH and cardiac and renal hypertrophy and remodeling (Figs. 1 –8). Thus, the activation of the RAS cannot fully explain the up-regulation of gp91 phox . Further studies are required to determine the mechanism of cold-induced up-regulation of gp91 phox .
The present study convincingly demonstrated that one single dose of AAV-gp91-shRNA attenuated hypertension and CV and renal damage for at least 10 weeks (length of the study). The reporter genes were still expressed at the endpoint of the study (Supplementary Figs. S3 and S4), indicating that AAV is indeed a long-term vector. AAV also had a decent transfection rate (Supplementary Figs. S3 and S4). AAV-2 is a clinical-grade vector and has been approved by the U.S. Food and Drug Administration for clinical trial in humans (Flotte et al., 2011; Mingozzi and High, 2011). We and others have shown that AAV is a safe vector for animal studies (Wang et al., 2006; Wang and Sun, 2009; Crosswhite and Sun, 2010; Flotte et al., 2011; Mingozzi and High, 2011). shRNAs delivered by AAV-2 have been used for regulating gene expression in cultured cells (Tenenbaum et al., 2004; Michel et al., 2005) and in animal models (Hasuwa et al., 2002; Wang et al., 2006; Sun et al., 2008). It was reported that high doses of shRNAs, particularly the 25-mer shRNAs, caused toxicity and morbidity in animals within 1 month (Grimm et al., 2006). In our study, AAV-2 delivery of 19-mer gp91 phox -shRNA at the dose of 1.25×1010 particles/rat did not cause obvious adverse effects in rats. The viral complexes did not affect the body weight gain.
Clinical Perspective
The present findings are of clinical significance and entail translational potential. The results will arouse clinicians' awareness of the adverse effects of cold temperatures on the CV function. There currently is no specific inhibitor for gp91 phox . To the best of our knowledge, this is the first study showing that gp91-shRNA, a gp91 phox -specific inhibitor, effectively prevented the cold-induced up-regulation of gp91 phox expression and superoxide production. The inhibition efficiency was over 50% (Figs. 4 and 5). It is noted that one single dose of AAV.gp91 phox -shRNA attenuated cold-induced CV dysfunction for at least 10 weeks (length of the study). The present findings suggest that AAV-2 delivery of gp91-shRNA may serve as a novel approach for ameliorating hypertension, vascular dysfunction, and organ damage.
Footnotes
Acknowledgments
This work was supported by NIH grants R01 HL077490, HL105302, and HL102074.
Author Disclosure Statement
The authors confirm that there are no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.