
Abstract
Immune cells are involved in the pathogenesis of osteoarthritis (OA). CD4+ T cells were activated during the onset of OA and induced macrophage inflammatory protein (MIP)-1γ expression and subsequent osteoclast formation. We evaluated the effects of local knockdown of MIP-1γ in a mouse OA model induced by anterior cruciate ligament transection. The mouse macrophage cell lines and osteoclast-like cells generated from immature hematopoietic monocyte/macrophage progenitors of murine bone marrow were cocultured with either receptor activator of NFκB ligand (RANKL) or CD4+ T cells. The levels of MIP-1γ and RANKL in cells and mice were examined by enzyme-linked immunosorbent assay (ELISA). The osteoclastogenesis was evaluated using tartrate-resistant acid phosphatase and cathepsin K staining. OA was induced in one hind-leg knee joint of B6 mice. Lentiviral vector encoding MIP-1γ small hairpin RNA (shRNA) and control vector were individually injected intra-articularly into the knee joints, which were histologically assessed for manifestations of OA. The expression of MIP-1γ and matrix metalloproteinase (MMP)-13 and the infiltration of CD4+ T cells, macrophages, and osteoclastogenesis in tissues were examined using immunohistochemistry. CD4+ T cells were involved in OA by inducing MIP-1γ expression in osteoclast progenitors and the subsequent osteoclast formation. Neutralizing MIP-1γ with a specific antibody abolishes RANKL-stimulated and CD4+ T-cell-stimulated osteoclast formation. MIP-1γ levels were significantly higher in synovium and the chondro-osseous junction of joints 90 days postsurgery. The number of infiltrated CD4+ T cells and macrophages and IL-1β expression were reduced in the synovial tissues of mice treated with MIP-1γ shRNA. Histopathological examinations revealed that mice treated with MIP-1γ shRNA had less severe OA than control mice had, as well as decreased osteoclast formation and MMP-13 expression. Locally inhibiting MIP-1γ expression may ameliorate disease progression and provide a new OA therapy.
Introduction
Inflammation is pivotal in the pathogenesis of OA. In the synovium of joints with OA, the inflammatory changes include synovial hypertrophy and hyperplasia with an increased number of lining cells, and the infiltration of a mixed population of inflammatory cells within the sublining tissue. Lymphocyte populations included T lymphocytes expressing CD4 and CD8 surface markers (Muir et al., 2011). In patients with severe OA, the extent of inflammation reached that observed in patients with clinically diagnosed rheumatoid arthritis (Farahat et al., 1993). Activated CD4+ T cells contribute to the development of OA through direct cell–cell contact or through soluble mediators (Wagner et al., 1994; Aarvak et al., 1999). We previously showed that CD4+ T cells promoted OA progression by inducing macrophage inflammatory protein-1γ (MIP-1γ) (Shen et al., 2011b), a member of a C-C chemokine family containing MIP-1α, MIP-lβ, MIP-1γ, RANTES, and so on. MIP-1γ is produced by monocytes and other types of leukocytes. By binding to chemokine receptor 1 (CCR1), a specific receptor on neutrophils, MIP-1γ acts as a chemoattractant that induces the chemotaxis of CD4+ T cells, CD8+ T cells, and monocytes (Poltorak et al., 1995; Mohamadzadeh et al., 1996). MIP-1γ promoted osteoclast formation and survival through the receptor activator of the nuclear factor (NF)-κB ligand (RANKL) pathway (Kong et al., 1999; Okamatsu et al., 2004). Osteoclast activation leads to the resorption and subsequent destruction of bone. We previously showed that in synovium infiltrated by CD4+ T cells, MIP-1γ expression was higher, which activated osteoclast formation and exacerbated OA (Shen et al., 2011b). On the basis of these findings, MIP-1γ knockdown may be a promising therapeutic strategy for OA.
Lentivirus-mediated gene transfer causes the efficient expression of transgenes in chondrocytes and synovial tissue (Coughlan et al., 2010; Chen et al., 2012). In this study, we examined (i) how CD4+ T cells induce osteoclast formation and (ii) the effects of MIP-1γ gene silencing mediated by lentiviral gene transfer in mice with OA.
Materials and Methods
Quantitative reverse transcription–polymerase chain reaction assay
Total RNA was extracted from RAW264.7 cells using TRIzol (Invitrogen, Carlsbad, CA) and complementary DNA (cDNA) was synthesized using a cDNA Reverse Transcription Kit (Applied Biosystems, Foster city, CA) according to the manufacturer's directions. The quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was performed on an StepOnePlus real-time PCR system (Applied Biosystems) using SYBR Premix Ex Taq (Tli RNase H Plus) (Takara Bio Inc., Shiga, Japan). The 5′ and 3′ mouse CCR1 gene-specific primers were designed as previously described (Okamatsu et al., 2004). The 5′ and 3′ mouse GAPDH gene-specific primers were 5′-GTT GTC TCC TGC GAC TTC AAC A-3′ (sense) and 5′-TTG CTG TAG CCG TAT TCA TTG TC-3′ (antisense). The qRT-PCR data were analyzed by the ΔΔCt method (Schmittgen and Livak, 2008). The fluorescence signal intensities were quantified as a Ct (threshold cycle) value with the StepOne Software v2.2 (Applied Biosystems). Normalization was performed using mouse GAPDH as an internal control, and relative gene expression was calculated using the comparative 2−ΔΔCt method. The 2−ΔΔCt values of the treated groups were compared with the control groups and presented as fold changes as compared with the control condition.
The tartrate-resistant acid phosphatase assay
Mouse macrophage RAW264.7 cells were cultured in complete medium consisting of Dulbecco's modified Eagle's medium, 10% cosmic calf serum (Hyclone, Logan, UT), 2 mM L-glutamine, and 50 μg/ml of gentamicin at 37°C in 5% CO2. We treated cells with RANKL (50 ng/ml; Peprotech, Rocky Hill, NJ) to stimulate osteoclast formation. Osteoclasts in culture were identified by histochemically staining tartrate-resistant acid phosphatase (TRAP; Sigma-Aldrich, St. Louis, MO) with or without MIP-1γ neutralization antibody (R&D Systems, Minneapolis, MN). To quantify TRAP signals, areas containing the largest number of osteoclasts were identified by scanning the sections at ×100 magnification. After the fields with the largest number of osteoclasts were determined, individual osteoclasts were counted at ×400 magnification. The number of osteoclasts was determined by averaging the number of osteoclasts in five areas at ×400 magnification.
In vitro coculture system
For osteoclast-like generation, mouse bone marrow cells were isolated and incubated in α-minimum essential medium with 50 ng/ml of mouse M-CSF (R&D Systems) and 100 ng/ml of RANKL to stimulate osteoclast formation. The medium was refreshed on day 3. To generate CD4+ T cells, total splenocytes were prepared from mice after the red blood cells had been lysed. Splenocytes were incubated with mouse CD4 (L3T4) microbeads (Macs; Miltenyi Biotec, Auburn, CA), after which CD4+ T cells were purified by passing them through a mass spectrometry column to enrich CD4+ T cells (Macs; Miltenyi Biotec). The yielded CD4+ T cells were at least 95–97% pure (confirmed using flow cytometry). The bone marrow cells (2.5×105/well) were then incubated with either purified CD4+ T cells (5×105/well) or with CD4+ T cell-conditioned medium (CM; 30%/well).
Enzyme-linked immunosorbent assay
To quantify the MIP-1γ, RANKL, and interferon-γ (IFN-γ), the levels of these cytokines and chemokines in the cell lysate and tissue homogenates were determined using an ELISA kit (R&D Systems).
Screening MIP-1γ small hairpin RNA and producing lentiviral particles
Three heteroduplexes (small hairpin RNA, shRNA) containing different 19 bp RNA segments to target different locations of the MIP-1γ gene were selected (National RNAi Core Facility, Taipei, Taiwan). The number following “sh” indicates the position of the oligonucleotide at the 5′-end of MIP-1γ cDNA (GenBank accession no. NM 011338). The heteroduplexes, supplied as 60-nucleotide oligomers, were annealed and inserted downstream of the U6 promoter into pLKO.1 plasmid, resulting in pLKO.1/sh240, pLKO.1/sh346, and pLKO.1/sh935. The three recombinant lentiviruses, accompanied by two control lentivirus, were produced by transiently transfecting pLKO.1/sh240, pLKO.1/sh346, pLKO.1/sh935, pLKO.1/shLuc, or pWPT, respectively, into 293T cells along with the packaging plasmid psPAX2 and envelope plasmid pMD2.G, using the calcium phosphate precipitation method as previously described (Salmon and Trono, 2007; Wang et al., 2010). After 48 hr, lentiviral particles (VPs) were collected and concentrated from the supernatant using ultracentrifugation. Physical titers of lentiviruses expressed as VPs were determined by analyzing the virus-associated p24 core protein (Lenti-X p24 Rapid titer Kit; Clontech, Mountain View, CA). Effective lentiviral shRNA was screened by infecting these viruses with mouse fibroblast NIH3T3 cells and RAW264.7 cells and, using ELISA, analyzing their inhibitory effect on MIP-1γ expression. The lentivirus containing the most effective shMIP-1γ RNA target sequences (sh935), lentivirus (Lt.) shMIP-1γ, as well as two control viruses, Lt.shLuc and Lt.GFP, was used for the animal study.
Animal models
Seven-week-old male B6 mice were purchased from the Laboratory Animal Center of National Cheng Kung University. The experimental protocol adhered to the rules of the Animal Protection Act of Taiwan and was approved by the university's Laboratory Animal Care and Use Committee. To induce experimental OA, each mouse was anesthetized with Zoletil 50 (10 mg/kg; Virbac, Carros, France) and then subjected to a previously described (Kamekura et al., 2005; Shen et al., 2011b) modified anterior cruciate ligament transection (ACLT) procedure. Immediately after surgery (day 0), 28 mice were randomly divided into 4 groups (n=7 each). They were intra-articularly injected in the ACLT knees with either Lt.shMIP-1γ or Lt.shLuc (5.79×108 VPs) on day 60 and day 75 postsurgery. Mice in the normal saline (NS) group were injected with 15 μl of NS using the same schedule as described above. The sham-operated group received no treatment. Ninety days after surgery, the mice were euthanized for a histological examination.
Histological assessment and immunohistochemistry
To test virus targeting in vivo, Lt.GFP, a lentiviral vector expressing green fluorescent protein (GFP) was intra-articularly injected (2×108 VPs) and the mice were killed 72 hr later (n=5 per group). To evaluate the histopathologic changes in the joints of the mice, ACLT was used to induce OA (n=7 per group). These mice were killed 90 days later. Their cartilage and synovial membranes were removed, fixed, and embedded in paraffin. Serial sections (5 μm thick) were cut and stained with Safranin-O/fast green and hematoxylin and eosin. The histologic change in cartilage was scored using Mankin's histologic grading method (Mankin et al., 1971). The histologic changes in synovial surface tissue and subsynovial tissue were evaluated and scored (Yoshimi et al., 1994). Briefly, the grading system assigns separate scores based on two categories: (a) three subcategories of the synovial lining layer—(i) hyperplasia of the synovial lining cells (0–3 points), (ii) hypertrophy of the synovial lining layer (0–3 points), and (iii) the infiltration of inflammatory cells (0–3 points)—and (b) three subcategories of the subsynovial tissue—(i) the proliferation of granulation tissue (0–3 points), (ii) vascularization (0–3 points), and (iii) the infiltration of inflammatory cells (0–3 points). Total scores in each category were calculated; the maximum score was 18 points. To test specific protein expression, the samples from joints were prepared from mice with different treatment. Serial sections of cartilage (and synovium) were stained with rat antimouse CD4 (L3T4; 1:100; H129.19; BD Biosciences Pharmingen, San Diego, CA), Mac-3 (a macrophage marker; 1:100; M3/84; BD Biosciences Pharmingen), GFP (B-2; 1:100; Santa Cruz Biotechnology, Santa Cruz, CA), IL-1β (H-153; 1:100; Santa Cruz Biotechnology), MIP-1γ antibody (1:100; Peprotech), matrix metalloproteinase (MMP-13; 1:50; Santa Cruz Biotechnology), and cathepsin K (an osteoclast marker; Proteintech, Chicago, IL) at 4°C overnight. After they had been sequentially incubated with the appropriate secondary antibody (1:400; Jackson ImmunoResearch Laboratories, West Grove, PA) for 2 hr at room temperature with aminoethyl carbazole as the substrate chromogen (Invitrogen Zymed Laboratories, Camarillo, CA), the slides were counterstained with hematoxylin.
Statistical analysis
All data are means±standard deviation. The quantification of the number of osteoclasts and MIP-1γ, RANKL, and IFN-γ expression was determined using the Student's t-test. Significance was set at p<0.05. JMP 8.0 (SAS Institute, Cary, NC) was used to analyze the histological data. Significance between groups was estimated using one-way analysis of variance. To evaluate the differences between groups, we used Tukey's Honestly Significant Difference test set at p<0.05.
Results
MIP-1γ antibody inhibited in vitro osteoclast formation
Our results showed that there was endogenous expression of MIP-1γ in RAW264.7 cells (782.48±59.96 pg/ml) (Fig. 1A). However, the levels of MIP-1γ were significantly increased in RANKL-treated RAW264.7 cells (4,168.43±1,343.50 pg/ml; p=0.012). The expression of CCR1 in cells induced by RANKL is more than five times as that in controls (5.76±2.76 vs. 1.07±0.51; p=0.0445; Fig. 1B). In vitro osteoclast differentiation was significantly and dose dependently (52.20±13.33, 0.5 μg/ml, p=0.0033; 18.21±6.69, 2.5 μg/ml, p=0.000024] inhibited in RAW264.7 cells treated with MIP-1γ antibody plus RANKL (50 ng/ml), but not in those treated with RANKL only (94.22±18.39) (Fig. 1C and D). The levels of MIP-1γ and soluble RANKL secreted by concanavalin A (ConA)-activated CD4+ T cells were significantly increased when compared with the controls (MIP-1γ: 338.52±115.71 pg/ml vs. 86.67±63.29 pg/ml, p=0.03; RANKL: 488.10±35.82 pg/ml vs. 261.45±102.35 pg/ml, p=0.0224, respectively) (Fig. 2A). Nevertheless, there was no significant difference in the levels of IFN-γ 7 days after ConA induction (337.23±7.73 pg/ml vs. 327.02±4.52 pg/ml; p=0.119). Furthermore, we isolated osteoclast-like cells from mice and cocultured them with activated CD4+ T cells. The levels of MIP-1γ in CD4+ T cell-activated cells were increased compared with controls (107.22±7.01 ng/ml vs. 0.76±0.04 ng/ml; p=0.000012) (Fig. 2B). Since the level of MIP-1γ secreted by activated CD4+ T cells was 338.52±115.71 pg/ml (Fig. 2A), most increased MIP-1γ should come from activated osteoclast-like cells. Although at lower levels in CD4+ T cell-CM–activated osteoclast-like cells, significant increase of MIP-1γ expression was seen (0.96±0.11 ng/ml; p=0.0478). CD4+ T cells activated osteoclast differentiation. This was true for cocultures of osteoclast progenitors with CD4+ T cells and for those progenitors treated with CD4+ T cell-CM (Fig. 2C). However, osteoclast differentiation was inhibited after the MIP-1γ antibody was added. The number of osteoclasts fell in CD4+ T-cell-treated TRAP-positive multinucleated cells (from 19.20±5.45 to 7.20±1.09; p=0.0013) and CM-treated TRAP-positive multinucleated cells (from 11.20±2.59 to 3.40±1.14; p=0.00026) (Fig. 2D).

The effect of MIP-1γ antibody on murine osteoclast differentiation stimulated by receptor activator of nuclear factor (NF)-κB ligand (RANKL).
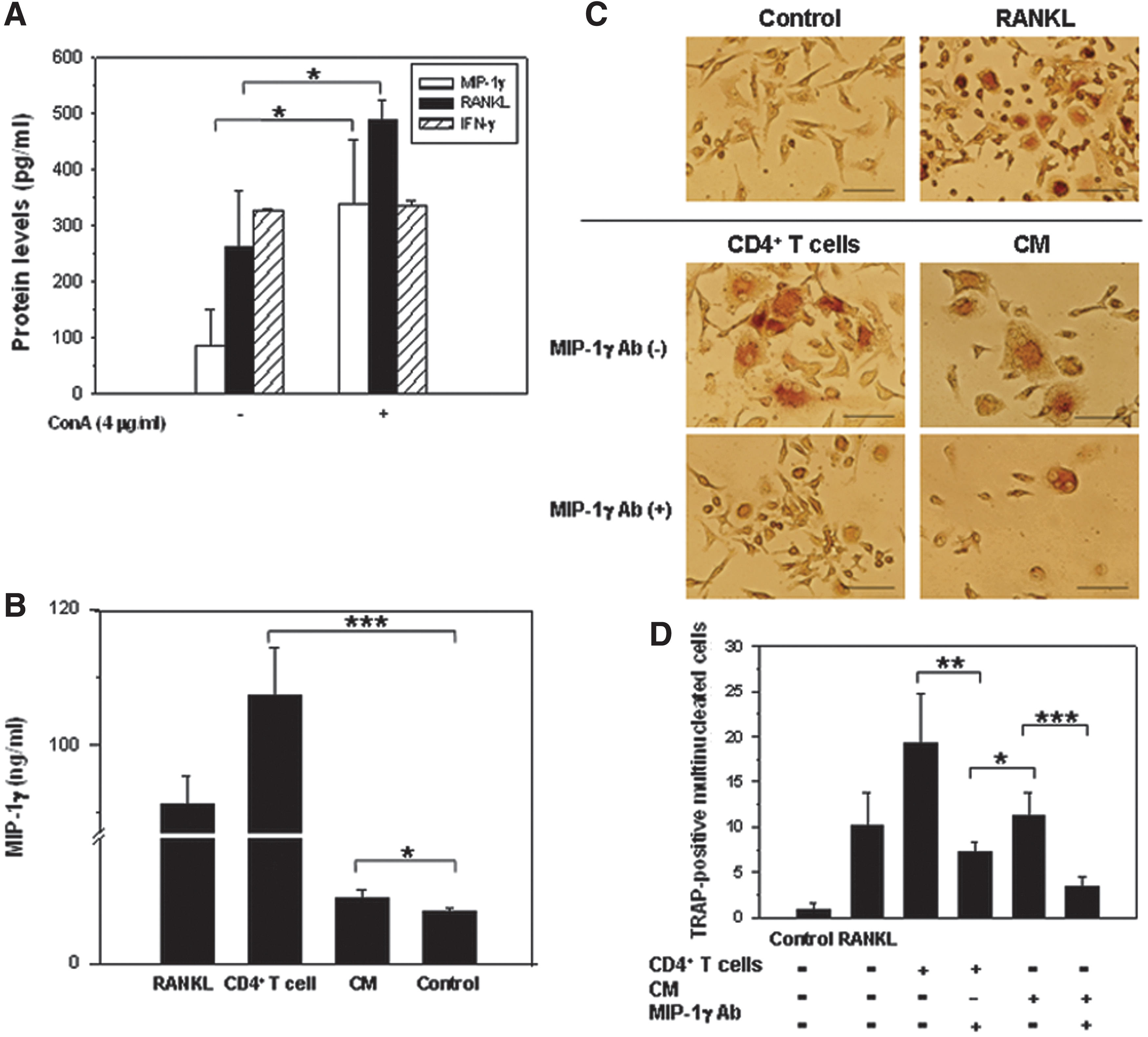
The effect of MIP-1γ antibody on murine osteoclast differentiation after stimulation by CD4+ T cells.
MIP-1γ expression during the progression of OA
MIP-1γ levels in the knee joints of mice with OA in the ACLT and Sham groups were not significantly different 30 days postsurgery (422.24±163.01 pg/ml vs. 441.61±103.06 pg/ml); however, they were significantly higher in the ACLT group 90 days postsurgery (Sham group: 464.75±70.24 pg/ml vs. ACLT group: 729.62±68.26 pg/ml; p=0.0007) (Fig. 3A). MIP-1γ expression was detected only in the lower metaphysis in the Sham group (Fig. 3B), whereas it was detected in the chondro-osseous junction of the ACLT group, and in the synovium of mice 90 days postsurgery.
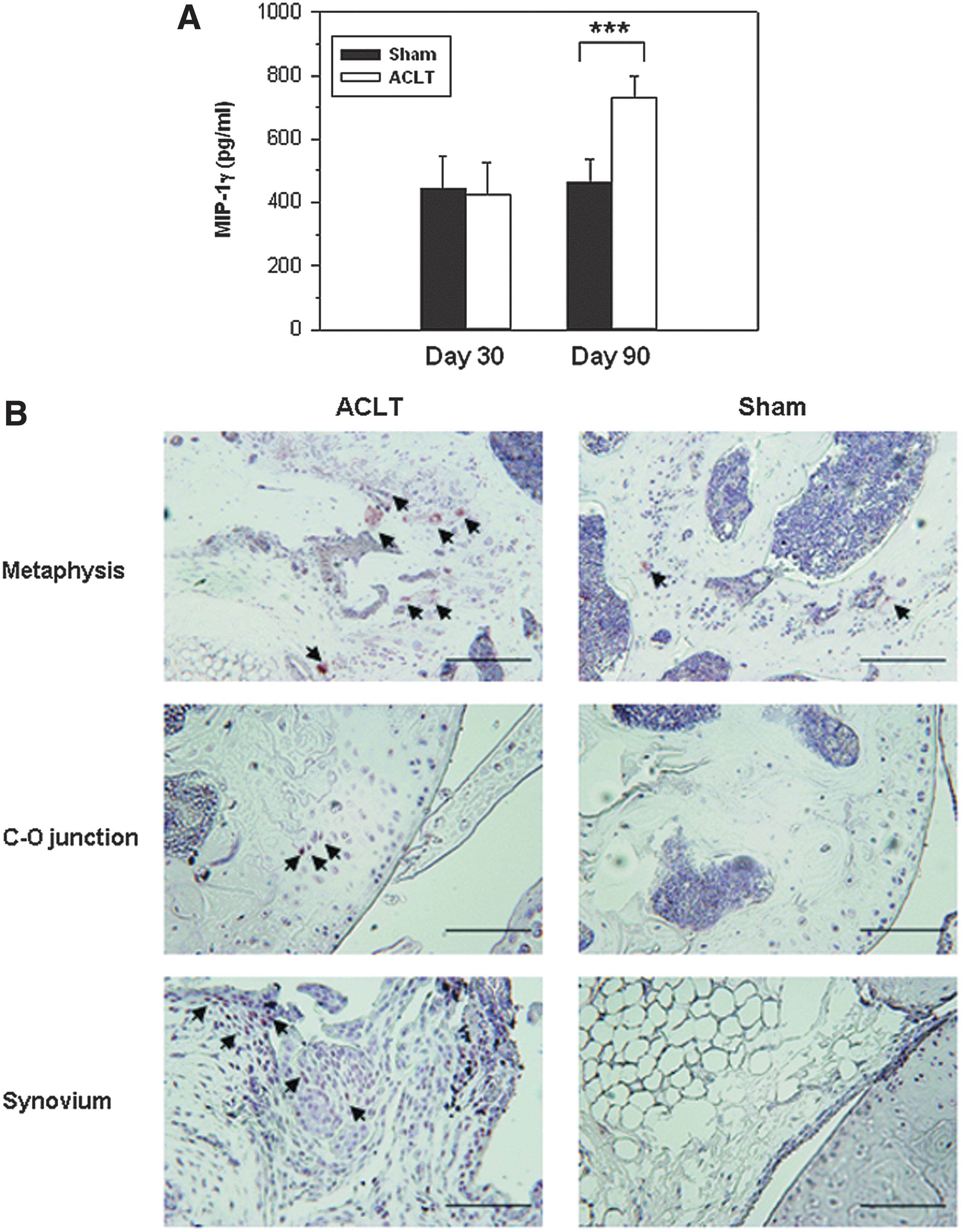
The expression of MIP-1γ in the knee joints of mice with OA.
Selecting shRNA sequences to target MIP-1γ
Three shRNA sequences, sh935, sh346, and sh240, were designed to specifically target mouse MIP-1γ. NIH-3T3 and RAW264.7 cells were infected with viruses that coded the three different shRNAs; then, the expression of MIP-1γ proteins was analyzed. The inhibitory efficacy of sh935 (73.32%; p=0.0139) and sh346 (72.47%; p=0.016) on MIP-1γ expression was similar in NIH3T3 cells (Fig. 4A) compared with the shLuc, a control virus encoding luciferase shRNA. In RAW264.7 cells, the inhibitory efficacy of sh935 was 78.25% (p=0.005) and of sh346 was 54.9% (p=0.011) compared with shLuc (Fig. 4A). We then named the recombinant virus carrying sh935-targeted sequence as Lt.shMIP-1γ, and used it for the subsequent studies. When induced with RANKL, the levels of MIP-1γ in Lt. shLuc-treated cells increased (2,037.76±491.41 pg/ml), and its expression could be significantly inhibited (Fig. 4B) by treating cells with Lt.shMIP-1γ (1,136.44±74.84 pg/ml, p=0.0348).

Targeting MIP-1γ using shRNA. Screening of effective MIP-1γ shRNA by infecting
In vivo histopathologic evaluation in mice with OA after recombinant lentivirus shMIP-1γ treatment
To test the virus targeting specificity in vivo, Lt.GFP, a lentiviral vector expressing GFP, was intra-articularly injected (2.0×108 VPs) into mice. The GFP could be detected in most of the superficial and deep zones of cartilage as well as synovia 72 hr after virus injection (Fig. 5A). After surgery, we intra-articularly injected in the ACLT knees with either Lt.shMIP-1γ or Lt.shLuc (5.79×108 VPs) on day 60 and day 75 postsurgery. Ninety days after surgery, more CD4+ T cells were infiltrated in the synovial membranes of NS and Lt.shLuc group mice than in those of Lt.shMIP-1γ group mice (Fig. 5B). Many macrophages were localized in the synovial membranes of NS and Lt.shLuc group mice, but only occasionally found in those of Lt.shMIP-1γ group mice. Abundant expression of IL-1β was also observed in the synovia of NS and Lt.shLuc group mice compared with that of Lt.shMIP-1γ group ones. The levels of RANKL in mice treated with Lt.shMIP-1γ were significantly reduced compared with those treated with Lt.shLuc and NS (Lt.shMIP-1γ group: 876.35±35.13 pg/ml vs. Lt.shLuc group: 998.00±79.94 pg/ml, p=0.0143; Lt.shMIP-1γ group: 876.35±35.13 pg/ml vs. NS group: 1,024.67±120.97 pg/ml, p=0.0331; Fig. 5C). Nevertheless, there was no significant difference in levels of IFN-γ among the four groups. No change has been observed either in the levels of MIP-1α (data not shown). These results indicated that MIP-1γ was the predominant C-C chemokine produced by CD4+ T cell-stimulated cells at the late stage of OA. The blocking of MIP-1γ may inhibit the recruitment of CD4+ T cells and macrophages into the synovial membranes and the subsequent inflammatory responses.
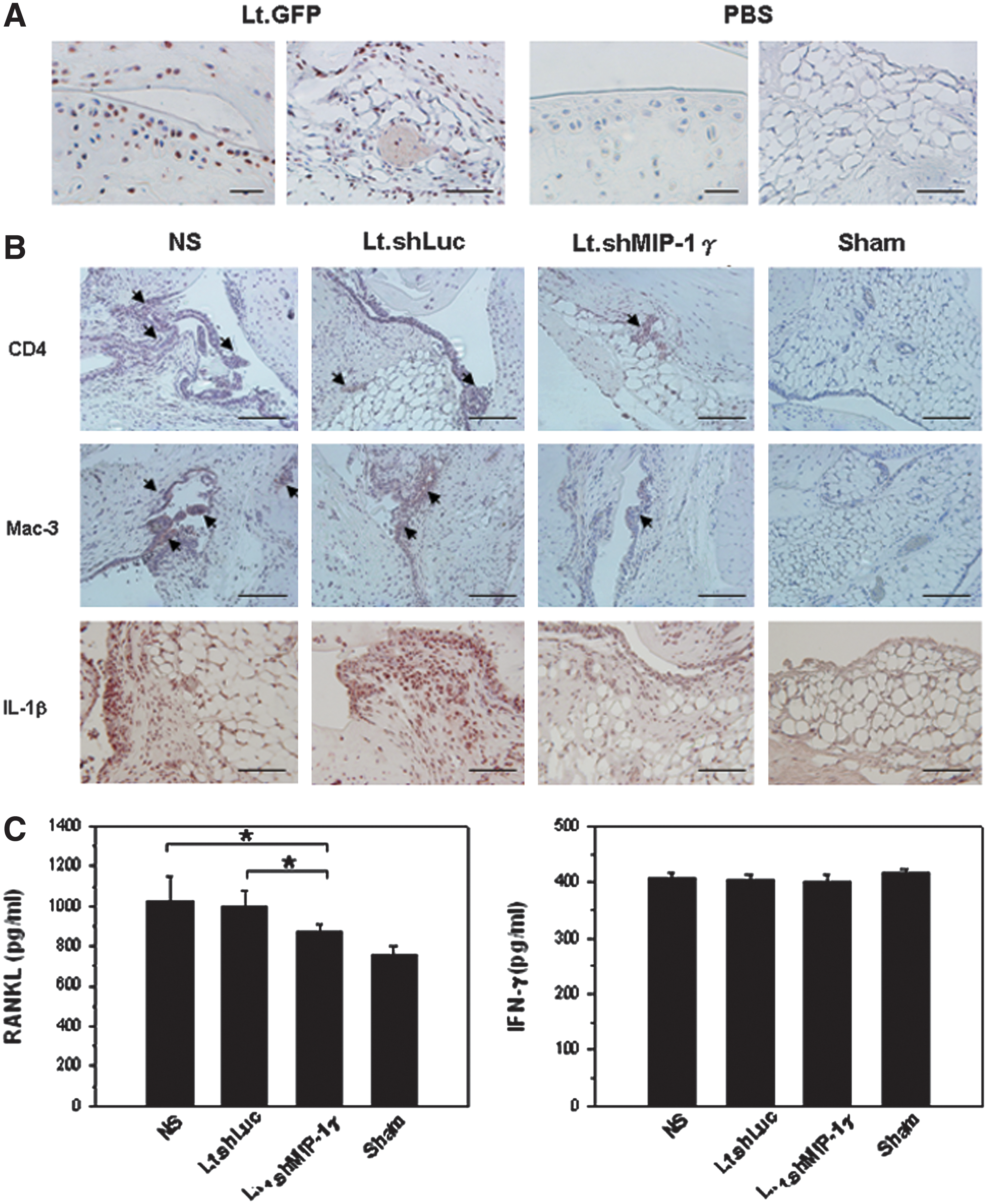
Inflammatory responses in the knee joints in Lt.shMIP-1γ-treated mice.
Histopathologic analysis showed a severe reduction of cartilage thickness and loss of chondrocytes both in the NS group and in the Lt.shLuc group (Fig. 6A). However, in the Lt.shMIP-1γ group, the severity of the lesions was reduced, and some fibrillation and irregularities were seen on the surface of the cartilage. In the sham-operated knee joints, most of the articular cartilage had a smooth surface. Synovium in the NS and Lt.shLuc groups showed hyperplasia and hypertrophy of the lining cells and greater cellular infiltration (Fig. 6B). In the Lt.shMIP-1γ group, only mild hyperplasia and mononuclear infiltration were seen compared with the Sham group. The osteoarthritic score in the Lt.shMIP-1γ group was significantly lower than that in the NS and Lt.shLuc groups (p<0.001; Table 1). The synovitis score in the Lt.shMIP-1γ group was also significantly lower than those in the NS and Lt.shLuc groups (p<0.001). MIP-1γ expression was higher in the metaphysis (Fig. 6C) and synovium of the NS and Lt.shLuc groups, but lower in the Lt.MIP-1γ group. Furthermore, osteoclast immunoreactivity was commonly detected in the proximal tibial growth plate. More cathepsin K-positive cells were detected in the NS and Lt.shLuc groups than in the Lt.shMIP-1γ group (Fig. 6D). Chondrocytes positive for the expression of MMP-13, a key regulator of cartilage degradation, were localized to all zones (superficial, deep, and subchondral) of articular cartilage in the NS and Lt.shLuc groups, but only occasionally found in the deeper zones of cartilage in the Lt.shMIP-1γ group (Fig. 6E). Taken together, these results suggested that inhibiting the MIP-1γ gene had modulated the formation of osteoclasts in the knee joints of mice after the induction of OA and that it had attenuated the development of OA.
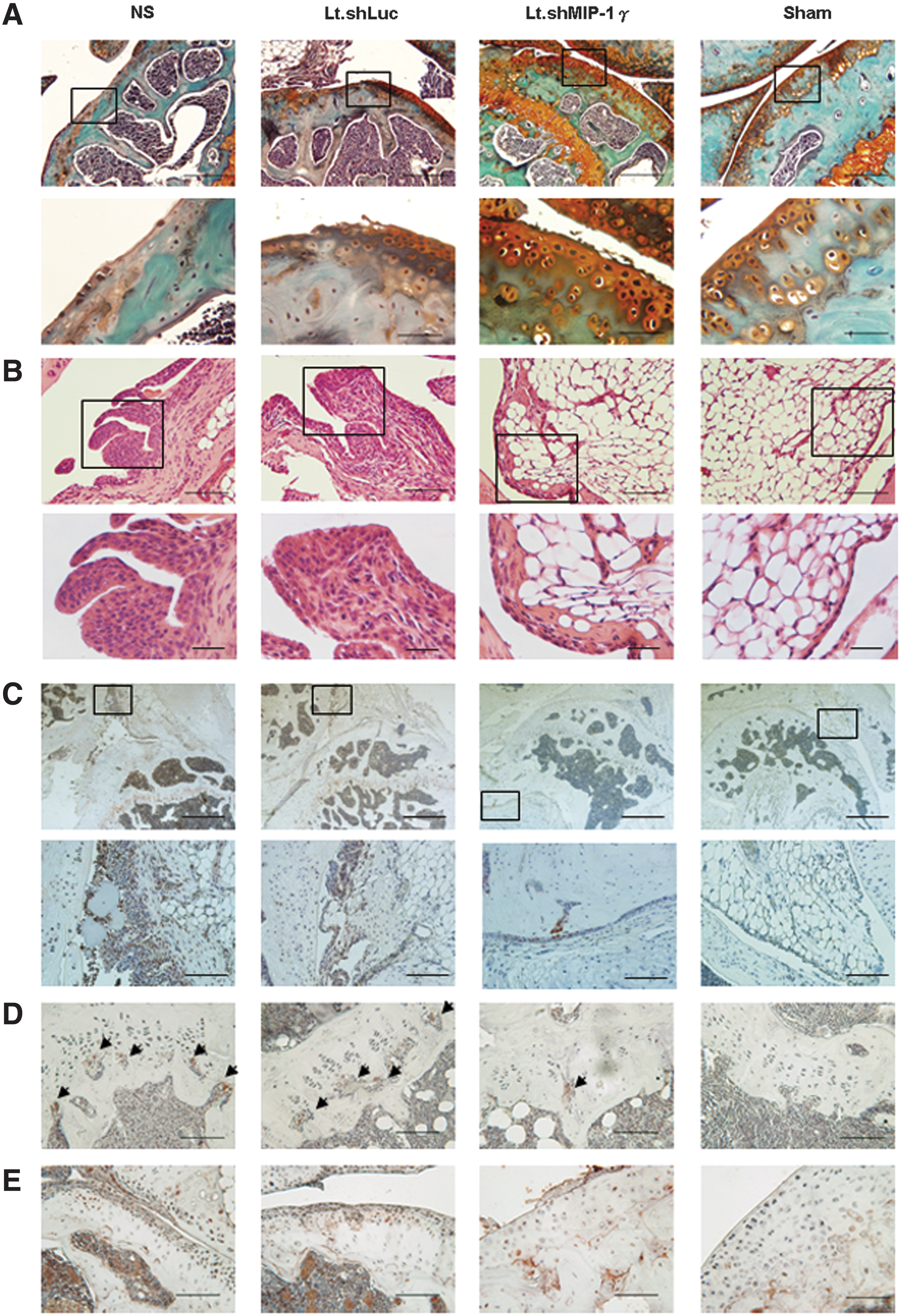
Histological analysis in the knee joints in Lt.shMIP-1γ-treated mice (n=7 per group). Cartilage on the posterior tibia condyle is shown.
Lt.shLuc, lentivirus encoding luciferase small hairpin RNA; Lt.shMIP-1γ, lentivirus encoding macrophage inflammatory protein-1γ small hairpin RNA; NS, normal saline.
Data are means±SD and analyzed using one-way analysis of variance and Tukey's Honestly Significant Difference tests.
p<0.001 compared with the NS and Lt.shLuc groups.
Discussion
The dynamic interplay of the immune and skeletal systems is thought to influence homeostasis and contribute to degenerative diseases (Nakashima and Takayanagi, 2009). CD4+ T cell regulation of osteoclastogenesis causes subchondral bone remodeling and initiates OA (Kotake et al., 2001; Crema et al., 2010). Patients with progressive OA usually have high subchondral bone turnover (Buckland-Wright et al., 2007). Osteoclast activation causes excessive bone resorption and subsequent bone destruction. We previously (Shen et al., 2011b) showed that CD4+ T cells induced osteoclast formation through the expression of MIP-1γ. In this study, we showed that activated CD4+ T cells could increase the production of MIP-1γ and RANKL (Fig. 2A). Both of these factors were able to induce osteoclast formation. When cocultured with activated CD4+ T cells, osteoclast progenitors were able to express high levels of MIP-1γ protein and differentiate into osteoclasts. Similar results were found in those osteoclast progenitors cocultured with CD4+ T cells CM. T cells themselves may produce MIP-1γ or stimulate other cells, such as multinucleated osteoclast-like cells, to produce MIP-1γ through the RANKL signaling pathway. The effects could be blocked by the addition of MIP-1γ neutralization antibody. These results suggested that through direct or indirect contact with a macrophage cell line or mouse osteoclast-like cells, activated CD4+ T cells may induce osteoclastogenesis.
When setting up OA animal models, we classified the disease stages based on the radiologic, macroscopic, and histopathologic evaluation. Moderate cartilage damage and synovia inflammation were observed 60 days after surgery, and it continued to get more severe till 90 days at sacrifice. We did not detect a significant increase of MIP-1γ in the joints of mice until 90 days after OA induction, which corresponded to severe OA. On the basis of this evidence, we therefore gave the mice two doses of lentiviral vectors (5.79×108 VPs) on days 60 and 75 postsurgery, which corresponded to moderate to severe OA. Our results showed that the severity of disease could be attenuated by intra-articular injections of shMIP-1γ using this regimen. This treatment is more close to a delayed intervention. Intra-articular administration of Lt.shMIP-1γ could both inhibit an abnormal cartilage turnover and inflammation.
Although cartilage lesions are the main feature of OA, subchondral bone destruction can trigger further cartilage degradation and contribute to the progression of disease. Lesions occur in subchondral bone are accompanied with a decrease of bone volume, suggesting that the bone resorption might impact cartilage remodeling and trigger OA. Accumulating evidence suggests that modifying subchondral bone provides alternative therapy for OA (Strassle et al., 2010). Bisphosphonates, commonly used to prevent and treat osteopenia and osteoporosis, were recently used for treating OA (Muehleman et al., 2002; Hayami et al., 2004; Funck-Brentano et al., 2012). Although promising results showed in animals, clinical trials using bisphosphonates to target osteoclasts have mixed and disappointing therapeutic effects (Bingham et al., 2006; Ramnaraine et al., 2012). When using this kind of drug, better results were seen in patients taking protective chronic doses. As a matter of fact, patients came to hospitals often have heterogeneous or advanced cases of OA, or both. To develop a therapeutic agent for symptomatic OA would be more desirable than using a preventive agent. The effects of shMIP-1γ on a later stage of OA have potentials for treatment of this disease.
Instead of using the MIP-1γ-neutralizing antibody, we chose lentiviral vector–encoding MIP-1γ shRNA to inhibit osteoclast formation based on the following reasons: First, according to the results from clinical trials, utilizing neutralizing antibody such as anti-TNF agents required long-term continuing treatment to control chronic diseases such as rheumatoid arthritis (Maini et al., 1999). However, such treatments used for patients are extremely expensive and time-consuming (Thrall, 2005). In contrast, using recombinant viral vectors for gene delivery to patients with chronic disease would benefit from both cost and therapeutic efficiency. Lentiviral vectors are able to infect a wide variety of dividing and nondividing cells, integrate stably into the host chromosomes, and result in long-term expression of the transgene (Singer and Verma, 2008). Upon one or two shots, the therapeutic levels of transgene can be usually achieved. Although there are safety concerns regarding to insertional mutagenesis, their integration profile seems more favorable than that of retroviral vectors. Several clinical trials are ongoing using lentiviruses for the treatment of congenital and acquired diseases and showing promising results (Mátrai et al., 2010). Additionally, we and other laboratories (Wazen et al., 2006; Buckland-Wright et al., 2007; Chen et al., 2011, 2012; Marzaioli et al., 2012; Ramnaraine et al., 2012) showed that lentiviral vector can efficiently transduce the synovial-lining cells, cartilage, and even calcified tissue, including osteoclasts, in the knees of animals without eliciting an inflammatory response. An efficient and stable gene transduction using lentiviral vectors in human osteoclast could be obtained for 4 weeks at a multiplicity of infection of 10 (Chu et al., 2008). Furthermore, the lentiviral transduction did not affect the function of osteoclast. We used Lt.GFP to test the virus targeting specificity. The GFP expression could be detected in the chondrocytes and synoviocytes, demonstrating that the lentiviral vectors were able to transduce genes efficiently into the cartilage and synovia.
Previously, we showed that 30 days after ACLT induction, both total CD4+ T cells and active CD4+ T cells increased in spleens (Shen et al., 2011b). In this study, our results revealed a more significant elevation of MIP-1γ levels in the joints at the late stage than the early stage of OA (Fig. 3). It seemed to have different dynamics between the CD4+T cells infiltration and the MIP-1γ expression in joints. In fact, the CD4+ T cells infiltration, although decreased along with disease progression, occurred in the synovium at all time from OA initiation to 90 days at sacrifice. We were thus able to detect a colocalization of CD4+ T cells and MIP-1γ expression in synovial tissues at the late stage of OA (Shen et al., 2011b). Although the activation of CD4+ T cells may occur during the onset of OA, the effects caused by activated CD4+ T cells on the composition of extracellular matrix can be more persistent. The other possibility is that different factors, such as IFN-γ and RANK secreted by different Th cell subsets, Th1 and Th17, respectively, may act at different time-points during OA progression. At the early stage of OA, when a large number of Th cells are activated, Th1 cells and the cytokines they produce, such as IFN-γ, play essential roles in initiating OA. At a later stage of OA, despite a decrease in the total number of Th cells, Th17 cells may activate now and continuously induce RANK production, resulting in subsequent disease exacerbation. However, this hypothesis required further investigation. IFN-γ produced by CD4+ T cells (Th1 and γδ T) had functions in macrophage activation, inflammation, T cell differentiation, and tissue remodeling (Hu and Ivashkiv, 2009; Pappalardo and Thompson, 2013). During OA progression, it was believed that IFN-γ exerted an antiosteoclastogenic effect on arthritis (Rho et al., 2004). It was recently reported that INF-γ was able to suppress the collagen-induced arthritis through the regulation of Th17 (Lee et al., 2013). Others demonstrated that IFN-γ tended to express in the early inflammatory arthritis or even at the prearthritic state (Adarichev et al., 2005; Raza et al., 2005). Our previous study showed that more substantial expression of IFN-γ was observed in synovium 30 days after OA induction than 90 days at sacrifice (Shen et al., 2011b). It appeared that INF-γ expressed and affected the disease in the initiation of OA. In the current study, our in vitro data showed that 7 days after ConA induction, the RANKL and MIP-1γ were both increased in activated CD4+ T cells except for IFN-1γ. In addition, the in vivo data showed that at a later stage of OA, no significant change of INF-γ expression was detected among the four groups (Fig. 5C). The temporal expression difference of INF-γ and MIP-1γ is probably the reasons why more osteoclasts and severe tissue damage were seen at the late stage of OA.
Several mediators, such as RANKL and IL-17 induced by activated Th cells, can stimulate osteoclast differentiation and formation. Like MIP-1γ, targeting these factors could inhibit the osteoclastogenesis and the following bone sorption (Dimitrova et al., 2012; Park et al., 2012). Using antibodies that specifically targeted to RANKL, clinical trials were conducted to treat osteoporosis and rheumatism arthritis, and showed promising results (Cohen et al., 2008; Cummings et al., 2009; Dore et al., 2010). Okamatsu et al. (2004) reported that among all the mediators screened, MIP-1γ was the most strongly upregulated cytokines in osteoclasts derived from RANKL-stimulated monocytes/macrophages. The prosurvival effect of RANKL was in part depends on its ability to induce MIP-1γ. In consistent with the previous reports, our data showed that MIP-1γ-neutralizing antibody could block the RANKL-stimulated osteoclastogenesis in vitro. Not only CD4+ T cells, but also most infiltrated cells (such as monocytes, neutrophils, and osteoclasts) in joints could produce MIP-1γ. Since MIP-1γ chemoattracts these cells into synovium, it may therefore affect the subsequent cell differentiation, survival, and activation likely via an autocrine way. An amplified therapeutic effect would be achieved when knocking down MIP-1γ. This might be the reason that inhibiting MIP-1γ expression in joint tissues would also reduce the RANKL expression in joints (Fig. 5C). Although there is no known human homolog, using shRNA to target MIP-1γ proves that effectively modulating the osteoclast and subchondral bone can significantly attenuate the OA.
In conclusion, we have provided evidence that CD4+ T cells directly and indirectly induce the formation of osteoclasts. The lentivirus that encoded MIP-1γ shRNA was used to treat OA. Our data showed that locally inhibiting MIP-1γ expression reduced the formation of osteoclasts and cartilage damage and led to the amelioration of OA.
Footnotes
Acknowledgment
This work was supported by Grant NSC 98-2314-B-273-001-MY3 from the National Science Council, Taiwan.
Author Disclosure Statement
No competing financial interests exist.