
Abstract
Gene transfer efficacy is limited by unwanted immunization against transgene products. In some models, immunization may be avoided by regulating transgene expression with mir142.3p target sequences. Yet, it is unclear if such a strategy controls T-cell responses following recombinant adeno-associated viral vector (rAAV)-mediated gene transfer, particularly in muscle. In mice, intramuscular rAAV1 gene delivery of a tagged human sarcoglycan muscle protein is robustly immunogenic and leads to muscle destruction. In this model, the simple insertion of mir142.3p-target sequences in the transgene expression cassette modifies the outcome of gene transfer, providing high and persistent levels of muscle transduction in C57Bl/6 mice. Such regulated vector fails to prime specific CD4 and CD8 T cells; although, transgene tolerance seems to result from ignorance and could be broken by a robust antigenic challenge. While effective in normal mice, the mir142.3p-regulated transgene remains immunogenic in sarcoglycan-deficient dystrophic mice. In these mice, transgene expression is only prolonged but does not persist as effector CD4 and CD8 T-cell responses develop. Thus, using a mir142.3p-regulated transgene can improve rAAV muscle gene transfer results, but the level of efficacy depends on the context of application. In normal muscle, this strategy is sufficient to prevent immunization and functions even more effectively than tissue-specific promoters. In dystrophic models, additional strategies are required to fully control T-cell responses.
Introduction
Restraining transgene expression to the target tissue can ameliorate gene transfer by reducing antibody or humoral immune responses against the transgene in many systems (De Geest et al., 2003; Follenzi et al., 2004; Franco et al., 2005; Di Domenico et al., 2006). Indeed we also observed that promoting muscle tissue expression of a therapeutic transgene improved rAAV1 gene transfer in sarcoglycan-deficient (sgca−/−) mice, a preclinical murine model of limb-girdle muscular dystrophy (LGMD) type 2D (Fougerousse et al., 2007). In this study, the ubiquitous expression of the human alpha-sarcoglycan gene (SGCA) from a cytomegalovirus (CMV) promoter induced a rapid loss of transgene with the production of anti-sarcoglycan antibodies and CD8+ cell infiltration of muscle parenchyma, whereas muscle tissue-driven expression of SGCA with the C5-12 muscle-driven promoter markedly prolonged muscle transduction (Fougerousse et al., 2007). However, tissue-specific promoters do not always improve gene transfer. The use of the C5-12 promoter for Factor IX (FIX) gene transfer in muscle with rAAV1 did not prevent the formation of anti-FIX antibodies in another model (Liu et al., 2004). It is generally not well understood if regulating transgene expression is sufficient to completely prevent the initiation of gene-specific T-cell responses, particularly following rAAV gene delivery in muscle.
Ameliorations in vector design have provided efficient gene expression control systems exploiting the patterns of expression of micro RNAs. Taking advantage of the restricted expression of mir142.3p in the hematopoietic system, the insertion of mir142.3p-target sequences in the transgene construct was used to prevent expression of the transgene in cells of the immune system, resulting in amelioration of transgene expression in vivo (Brown et al., 2006, 2007a, 2007b). Like tissue-specific expression systems, the mir142.3p-target sequences are thought to act on by preventing expression of the transgene in professional antigen-presenting cells (APCs) that are capable of priming T-cell responses while permitting transgene expression in solid tissues. Using such mir142.3p-regulation strategy can efficiently reduce antibodies and T-cell response against the transgene and improves liver-directed lentiviral gene transfer in rodent models of haemophilia B or Crigler-Najjar type 1 (Brown et al., 2007a; Schmitt et al., 2010). In the particular context of liver-gene transfer, the use of mir142.3p-regulated lentiviral vectors also seems to cause an active immune tolerance against the transgene by inducing natural regulatory T cells (Annoni et al., 2009). However, this strategy has not yet been successfully translated in other gene transfer systems.
The SGCA gene transfer model provides an opportunity to investigate if mir142.3p-regulated vectors can also control T-cell responses in the context of skeletal muscle-targeted rAAV gene transfer. Using a new chimeric transgene SGCA-HY and rAAV1 IM gene delivery, we show that the priming of CD4 and CD8 T-cell responses was simply and effectively controlled with a mir-142.3p-regulated vector. This strategy did not induce active tolerance but enabled the long-term persistence of gene-modified cells in normal muscle. While efficient in normal mice, the strategy was less efficient in sgca−/− mice, highlighting that distinct mechanisms of immunization may occur in pathological models and that the efficacy of mit142.3p-regulated vectors is context dependent.
Materials and Methods
Animals
Six- to eight-week-old C57Bl/6 (CD45.2) and congenic CD45.1 (PtprcaPep3b/BoyJ [CD45.1]) mice were purchased from Charles River. Marilyn and Mata-Hari mice carrying transgenic T-cell receptors that recognize products of the male HY gene, respectively the Dby peptide NAGFNSNRANSSRSS presented by I-Ab (CD45.1, Rag2−/−) (referred to as Dby peptide) and Uty peptide WMHHNMDLI presented by H2-Db (CD45.2, Rag2−/−) (referred to as Uty peptide), were kind gifts from O. Lantz (Thery et al., 2002; Valujskikh et al., 2002). Sgca-null mice were obtained from K. Campbell (Howard Hughes Medical Institute, Iowa City, IA) and were bred in our animal facility (Duclos et al., 1998; Turk et al., 2006). All animal experiments were performed according to institutional and international guidelines for animal care and use.
Construction of the transgene plasmids
Plasmids encoding the human SGCA gene fused or not to HY and expressed from either the CMV or desmin promoter (respectively pAAV CMV haSarco_HY, pAAV Des haSarco_HY, and pAAV Des haSarco) were constructed from the pGG1 CMV haSarco and pGG1 Des haSarco plasmids using standard techniques (Fougerousse et al., 2007). The generation of SGCA-HY was made by amplifying the coding sequence of SGCA (without the stop codon) with oligonucleotides: haSarco F: 5′GAACTTATGGCTGAGACACTCTTCTGGAC-3′haSarco-DBY R: 5′-CATTTGCAGAACCACTGCTGTGCTGGTCCAG-3′ and fusing the HY coding sequence amplified by polymerase chain reaction (PCR) with oligonucleotides: HY F (5′-CTGACCAGCACAGCAGTGGTTCTGCAAATG-3′) and HY XhoI R (5′-CCGCTCGAGCTAGGTATTGTCTCCAATTAGGATCCA-3′) and inserting the products into pAAV_MCS digested with HindIII and XhoI (Biolabs). To generate the mir142.3p-regulated constructs (sometimes abbreviated mir142.3pT in figures), we annealed four repeats of the miR-142-3p target sequences described in (Brown et al., 2006) with the following oligonucleotides: 5′CTAGAGTCGACTCCATAAAGTAGGAAACACTACACGATTCCATAAAGTAGGAAACACTACAACCGGT-3′ 5′
rAAV vector production
Adenovirus-free rAAV2/1 vector preparations were generated by transfection of HEK293 cells with three plasmids, purified by centrifugation over a CsCl gradient, and titered by real-time PCR as previously described (Riviere et al., 2006).
Intramuscular injection, immunofluorescence, and histology
For intramuscular injection, animals were anesthetized by intraperitoneal injection of xylazine (10 mg/kg) and ketamine (100 mg/kg), and the left tibialis anterior (TA) was injected with 25 μl of viral preparation diluted into phosphate-buffered saline (PBS) at the indicated concentration. Mice were sacrificed at indicated time points, and the injected muscles were harvested and snap frozen in liquid nitrogen.
Transverse cryosections (8 μm thick) of frozen muscle were either stained with hematoxylin and eosin or used for immunodetection of muscle proteins. In such cases, slices were rehydrated with PBS for 5 min, incubated for 20 min at room temperature (RT) with H2O2 to inhibit endogenous peroxidase activity, washed with PBS, and incubated with blocking medium consisting of PBS with 10% goat serum for 30 min before incubation for 1 to 2 hr at RT with either 1:1000 dilution of a rabbit polyclonal primary antibody directed against amino acids 366–379 of the human a-sarcoglycan protein (AC-ahSarco57) (Fougerousse et al., 2007), or antibodies against dystrophin (1/200 dilution, Thermo Scientific), γ-sarcoglycan (1/200 dilution, Santa-Cruz), or β-sarcoglycan (1/40 dilution, Novocastra). After washing with PBS, sections were incubated for 1 hr at RT with secondary antibody conjugated with horseradish peroxidase (HRP) diluted 1/200 (Dako), washed with PBS before revealing the stain with diluted diaminobenzidine (Dako) for 2 to 5 min, fixed with ethanol (5 min) and xylene (twice for 5 min), and mounted with Eukkit before being examined with a Nikon microscope. Digital images were captured using CCD camera (Sony) and processed using Cartograph 5.5 (Microvision software). For immunofluorescence stainings, sections were incubated with the AC-ahSarco57 antibody revealed by a goat anti-rabbit Alexa 488 (Invitrogen) and also with anti-mouse CD8a antibody (Caltag), followed by a biotinylated rabbit anti-Ig antibody (Dako) revealed by streptavidin Alexa 594 (Molecular Probes – Invitrogen). Slides were mounted with a Fluoromount and Dapi mix (Southern Biotech). The slides were examined with a Leica DMRB microscope (25x objective), and digital images were captured using Retiga 2000R camera and processed using Cartograph 5.5 (Microvision software).
Measure of transgene expression by real-time reverse transcriptase–PCR
Total RNA was extracted from muscles using Trizol (Invitrogen). Residual DNA was removed from the samples using the Free DNA kit (Ambion). cDNA was synthesized from 1 μg of RNA using random hexamers according to the protocol Superscript II first-strand synthesis system for reverse transcriptase (RT)-PCR (Invitrogen). Real-time PCR was performed using ABI PRISM 7700 system (PE biosystems) with 0.2 mM of each primer and 0.1 mM of the probe, according to the protocol Absolute QPCR Rox Mix (ABgene). The primer pairs and Taqman probes used for ha-sarcoglycan amplification were: 920hasarco. F: 5′-TGCTGGCCTATGTCATGTGC-3′, 991hasarco.R:5′-TCTGGATGTCGGAGGTAGCC-3′, and 946hasarco.P: 5′-CGGGAGGGAAGGCTGAAGAGAGACC-3′.
The ubiquitous acidic ribosomal phosphoprotein gene (PO) was used to normalize the data across samples. The primer pairs and Taqman probe used for PO mRNA amplification were: m181PO.F: 5′-CTCCAAGCAGATGCAGCAGA-3′, m267PO.R:5′-ACCATGATGCGCAAGGCCAT-3′, and m225PO.P: 5’-CCGTGGTGCTGATGGGCAAGAA-3’.
Adoptive transfer of T-cell receptor (TCR)-transgenic T cells
C57Bl/6 mice were injected with 106 splenocytes from congenic MataHari mice or Marilyn mice, and the following day, the injected C57Bl/6 mice received the indicated dose of rAAV2/1 vector. The levels of adoptively transferred cells in peripheral blood lymphocytes were monitored by fluorescent-activated cell sorter (FACS) over time. To analyze antigen-specific CD4+ T-cell proliferation, the cells from Marilyn mice were stained prior to the adoptive transfer with 2 μM carboxyfluorescein succinimidyl ester (CFSE) (Molecular Probes) in RPMI-1640 (10.106 cells/ml) at RT for 8 min and at 37°C for 2 min. The reaction was stopped by adding an equal volume of RPMI 1640 10% fetal calf serum (FCS) followed by a 5 min incubation at 4°C. Cells were extensively washed, resuspended in PBS, and injected in the tail vein (0.2 ml per injection).
Peptides and adjuvants
The Dby and Uty peptides were synthesized by Genepep. To analyze immune tolerance, 21 days after injection of the rAAV, mice were injected subcutaneously with 100 μM Dby peptide or PBS emulsified in complete Freund's adjuvant (CFA) (v/v) (Sigma Aldrich). At day 35, the frequency of Dby-activated T cells was measured by IFN-γ enzyme-linked immunospot assay (ELISPOT).
Cytokine determination
IFN-γ and IL-2 ELISPOT were performed as previously described (Boisgerault et al., 2001; Gross et al., 2003), culturing 0.5 and 1×106 spleen cells per well with or without 1 μM of peptide. As a positive control, cells were stimulated with Concanavalin A (5μg/ml). After 24 hr, spots were revealed and counted using a Bioreader 2000 (BIO-SYS). Spot-forming units (SFU) are represented after subtraction of background values obtained with nonpulsed splenocytes (background values are below 30 SFU/well), unless indicated otherwise. The secretion of IL-2, IL-10, and IFNγ after restimulation of cells for 24 hr or 96 hr was measured by capture ELISAs using eBioscience reagents according to the manufacturer's instructions. Results were expressed as cytokine concentration in pg/ml. The detection limit was 2 pg/ml for IL-2, 30 pg/ml for IL-10, and 15 pg/ml for IFNγ.
In vivo killing assay
Female and male C57Bl/6 CD45.1 spleen cells were incubated in obscurity with 2 μM and 0.2 μM CFSE respectively. After washing, 2×107 cells were injected in the tail vein in 0.2 ml PBS. Two days post-cell injection, spleen cells were collected from individual mice and analyzed by FACS. The percentage of specific lysis of male over female cells (m/f) was calculated on CD45.1+ cells as follows: % specific lysis=[(m/f)t0 − (m/f)tx] / [(m/f)t0] × 100. The quantities m and f are measured on standard gates placed on the male CFSE low and female CFSE high histograms peaks.
Flow cytometry
All reagents used for flow cytometry were purchased from BD-Biosciences. For spleen and blood staining, erythrocytes were eliminated by hypotonic shock with PharMLysis buffer. Cell suspensions were first incubated with anti-FcγRIII/II (2.4 G2) monoclonal antibodies (mAb) for 15 min at 4°C and then stained for 30 min at 4°C in PBS with 0.1% bovine serum albumin (BSA) using saturating amounts of the following mAbs: fluorescein-conjugated (FITC) anti-CD8, FITC-conjugated anti-CD4, pacific blue–conjugated anti-CD4, biotinylated anti-CD45.1 and anti-CD45.2, and APC-conjugated streptavidin. Dead cells were excluded using 7-actinomycine D (Sigma Chemical Co) staining. MHC class I pentamer staining was performed on a minimum of 1×106 splenic cells, incubated for 10 min in obscurity at RT with the R-PE labeled pro5 MHC Pentamer (H-2Db WMHHNMDLI) (Proimmune). Cells were further incubated for 15 min with 2.4 G2 antibody against FcII/III receptor, then stained for 30 min in PBS with 0.1% BSA and FITC-conjugated anti-CD8 mAb, washed, and immediately analyzed. Analyses were performed on a FACSCalibur or LSRII using CELLQuest or Diva software (BD-Biosciences).
Statistical analysis
Two-group comparisons were performed using a Mann and Whitney two-tailed analysis. Values were considered to be statistically significant if p<0.05.
Results
A model to monitor T-cell responses to a skeletal muscle protein
Our prior studies using human SGCA gene transfer in mice (Fougerousse et al., 2007) showed that regulating transgene expression had a positive impact on sustained correction of muscle. To develop this model and to facilitate the study of transgene-specific T-cell responses, we generated a reporter chimeric/fusion transgene SGCA-HY enabling studies of T-cell responses in scga−/− and C57Bl/6 mice. Male HY gene sequences were inserted in-frame 3’ of SCGA (Fig. 1a, top), generating a chimeric human alpha-sarcoglycan. In this fusion protein, the Dby and Uty peptides respectively presented by major histocompatibility complex (MHC) I-Ab and H2-Db to CD4 and CD8 T cells (Thery et al., 2002; Valujskikh et al., 2002) are inserted in a structurally permissive portion of the C-terminus of the alpha-sarcoglycan transmembrane protein (Draviam et al., 2006). The chimeric protein is expressed at the membrane of skeletal myofibers (Fig. 1b) like the nonmodified alpha-sarcoglycan and is biologically functional. The dystrophin-sarcoglycan complex is destabilized in muscle fibers of sgca−/− mice (Draviam et al., 2006) and can be restored by SGCA gene transfer (Fougerousse et al., 2007). The dystrophin-sarcoglycan complex is also restored by SGCA-HY gene transfer (Fig. 1b) as shown by expression of alpha-sarcoglycan in the same myofibers expressing beta-sarcoglycan and gamma-sarcoglycan proteins in proximity to dystrophin. Altogether, SGCA-HY is a pertinent tool to study the consequences of gene transfer on a skeletal muscle component.
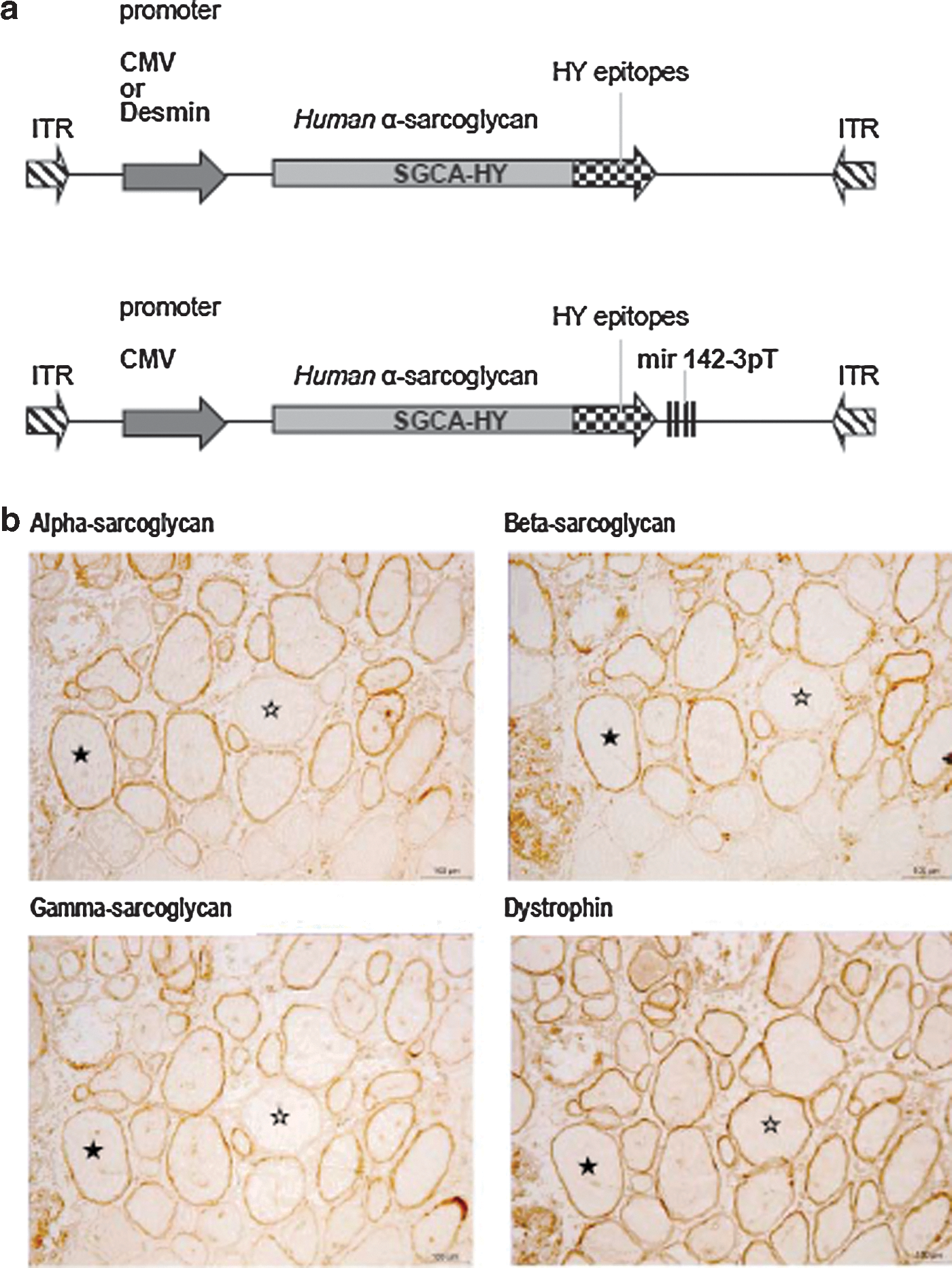
Validation of the SGCA-HY transgene model.
At the immunological level, and as expected from the presence of HY peptides, the human alpha sarcoglycan-HY protein was immunogenic in female but not male mice as shown by specific IFN-γ production in the spleen following IM gene transfer of SGCA-HY (Supplementary Fig. S1a; Supplementary Material available online at
Loss of transgene over time is associated with the induction of specific CD4 and CD8 T-cell responses
In female C57Bl/6 mice, the IM administration of rAAV1 encoding the CMV-driven SGCA-HY transgene (CMV_SGCA-HY) led initially to high levels of gene expression as measured by q-RT-PCR (Fig. 2a, day 8), but mRNA expression rapidly declined, reaching background levels within 3 weeks. This rapid loss of expression coincided with muscular inflammation, which was extensive 2 weeks post vector injection as shown by histology (Fig. 2b). Myofibers were destroyed and skeletal muscle tissue remodeling was complete within 4 weeks of vector administration. At this time point, a mild infiltrate of the tissue remained visible, but a generalized centro-nucleation of myofibers indicated that extensive regeneration of the tissue had occurred (Fig. 2b, day 30). Concomitantly, potent transgene-specific CD4 and CD8 T-cell responses were measured in the spleen. Two weeks after vector injection, transgene-induced CD4 T-cell responses were characterized by high frequencies of Dby-activated IFN-γ-producing cells (Fig. 3a). The induction of transgene-specific Th1 cells was confirmed by the secretion of IFN-γ and IL-2 with low levels of IL-10 (Supplementary Fig. S2). Transgene-induced CD8 T-cell responses were characterized by high levels of Uty-pentamer+ CD8+ T cells, representing 5 to 10% of CD8+ T cells in the spleen and blood of the mice (Fig. 3b). Such splenic CD8+ T-cell response was functional as shown by IFN-γ production (Fig. 3c) and by male-specific cytolytic activity (Fig. 3d). In summary, rAAV1 IM administration of CMV-driven SGCA-HY induces the rapid development of robust transgene-specific CD4 Th1 and CD8 cytolytic T-cell responses coinciding with the destruction of gene-modified tissue and loss of transgene expression.
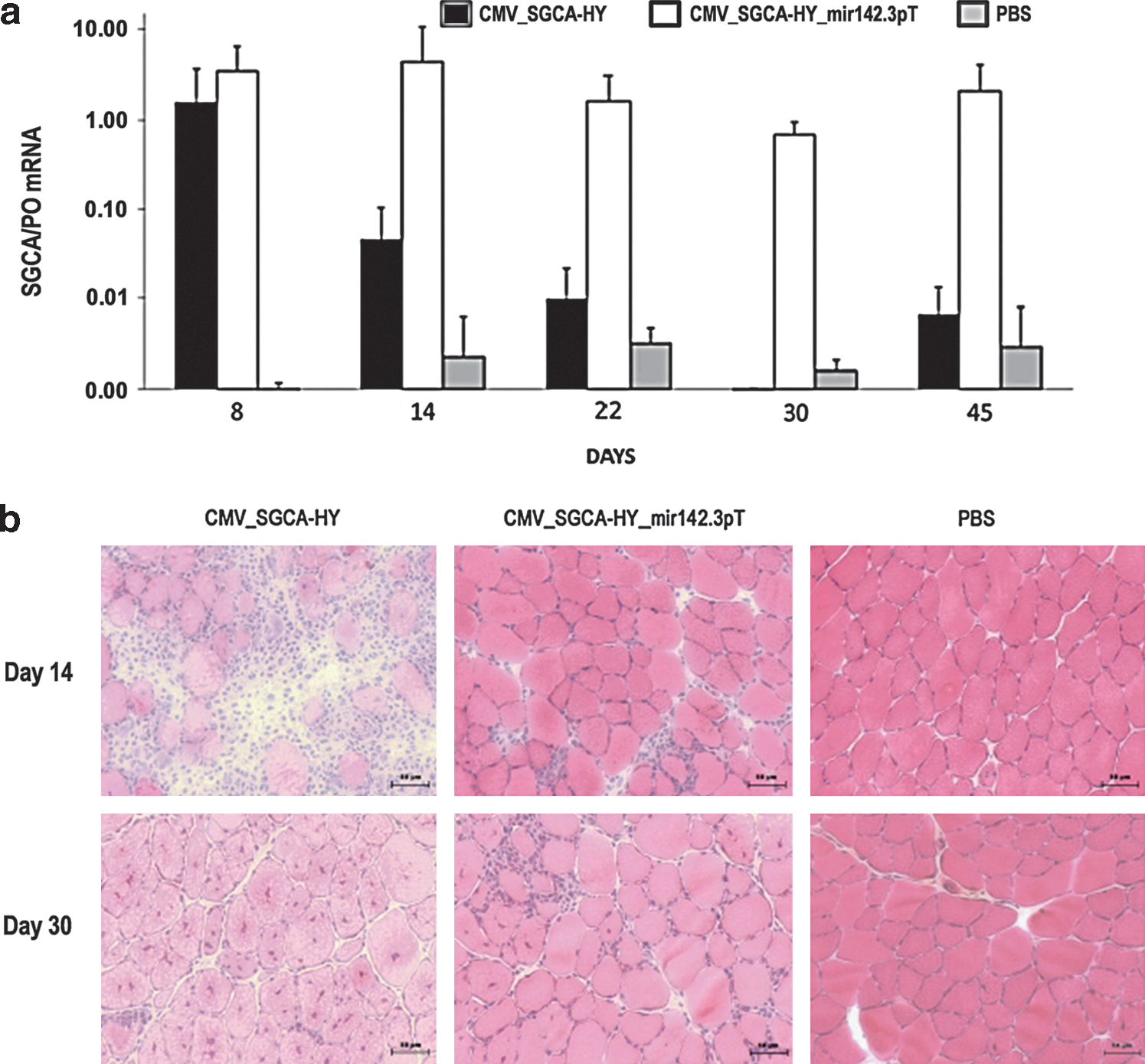
Effect of SGCA-HY gene transfer on muscle structure and transgene expression in C57Bl/6 mice. Female C57BL/6 mice were injected with PBS or with vector (5×109 vg of rAAV1_CMV_SGCA-HY or of rAAV1_CMV_SGCA-HY_mir142.3pT) in the left TA and muscle transduction was analyzed over time.
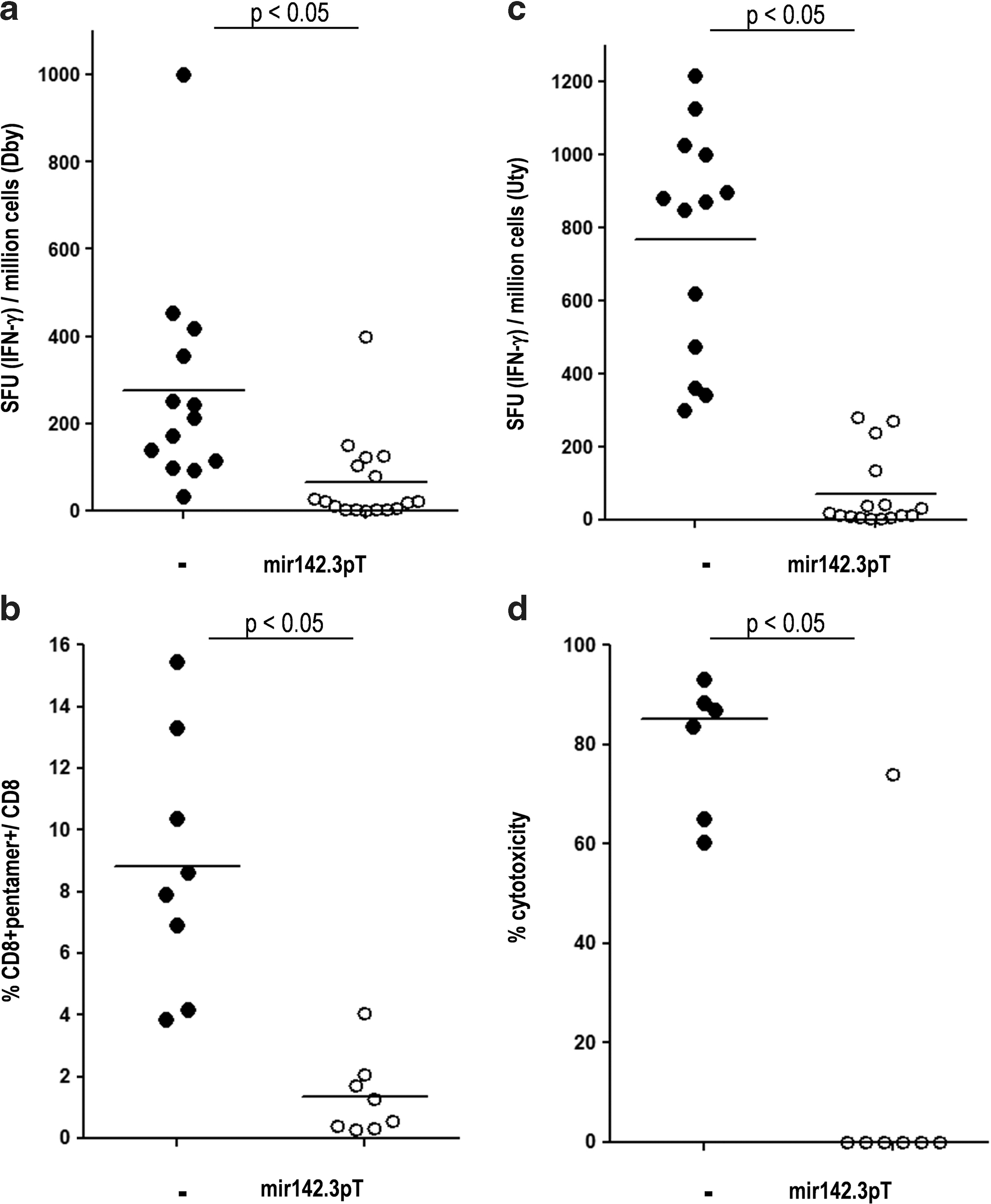
Analysis of transgene-specific T-cell responses. C57Bl/6 mice were injected IM into the TA with 5×109 vg of rAAV1_CMV_SGCA-HY vector carrying (open dots) or not carrying (solid dots) mir142.3pT sequences. Fourteen days later, spleen cells were harvested to measure
Mir142.3p-regulated SGCA-HY expression prolongs transgene expression, prevents muscle destruction, and abrogates CD8 and CD4 T-cell responses
The regulation of transgene expression with mir142.3p target sequences prevents transgene expression in the hematopoietic system and reduces immune responses following lentiviral-mediated gene transfer (Brown et al., 2006). To determine if this particular strategy is able to control transgene-specific T-cell responses in our system, we inserted four repeats of the mir142.3p target sequence in the 3’ untranslated region of CMV-driven SGCA-HY expression cassette (Fig. 1a, bottom). Following rAAV1 IM delivery, the mir142.3p-regulated SGCA-HY transgene permitted expression of high levels of muscle transduction, as high as initially reached with the nonregulated CMV-driven construct (Fig. 2a).
Contrary to the nonregulated construct that rapidly failed, high levels of SGCA mRNA persisted in muscle for up to 45 days (Fig. 2a) at values corresponding to therapeutically efficient levels in the LGMD2D mouse model (Fougerousse et al., 2007). The mir142.3p-regulated rAAV1 vector caused minimal infiltration of the muscle parenchyma (Fig. 2b). The tissue remained intact without evidence of ongoing regeneration as few centro-nucleated myofibers were observed 30 days post vector injection. The long-term maintenance of muscle transduction without tissue damage correlated with the absence of transgene-specific T-cell immune responses (Fig. 3a–d). All assays including the cytokine production assays, the detection of Uty-pentamer+ CD8 T cells, and the cytotoxicity assays concurred to show very low to undetectable levels of transgene-specific CD4 and CD8 T-cell responses following IM administration of rAAV1 CMV_SGCA-HY_mir142.3pT. Taken together, the data show that mir142.3p-regulated expression of the transgene efficiently prevents the development of CD4 and CD8 T-cell responses against the transgene product, thereby allowing sustained transgene expression in muscle.
Mir142.3p-regulated expression prevents the priming of transgene-specific CD4+ and CD8+ T cells more effectively than tissue-specific promoters
To analyze the initial development of transgene-specific T cells, we measured the amplification of adoptively transferred naive T cells following administration of vector. We used transgene-specific naive CD4 and CD8 T cells that were respectively obtained from Marilyn or Mata-Hari female mice bearing TCRs specific to the male Dby or Uty peptides (Thery et al., 2002; Valujskikh et al., 2002). Following injection of the nonregulated CMV_SGCA-HY rAAV1 vector, the circulating blood levels of Dby-specific CD4+ T cells rose sharply, peaking at day 6, and returned to baseline in about 2 weeks (Fig. 4a). Such kinetics were confirmed in the spleen (data not shown). The CD4+ T-cell response was followed by the amplification of Uty-specific CD8+ T cells, which started rising 1 week after vector injection, peaking at 3 weeks, and returning to basal levels by about 10 weeks (Fig. 4b). In sharp contrast, no expansion of CD4 or CD8 T cells occurred following administration of the mir142.3p-regulated vector over the periods of time examined, which spanned 30 days for CD4 cells and more than 70 days for CD8 T cells (Fig. 4b).
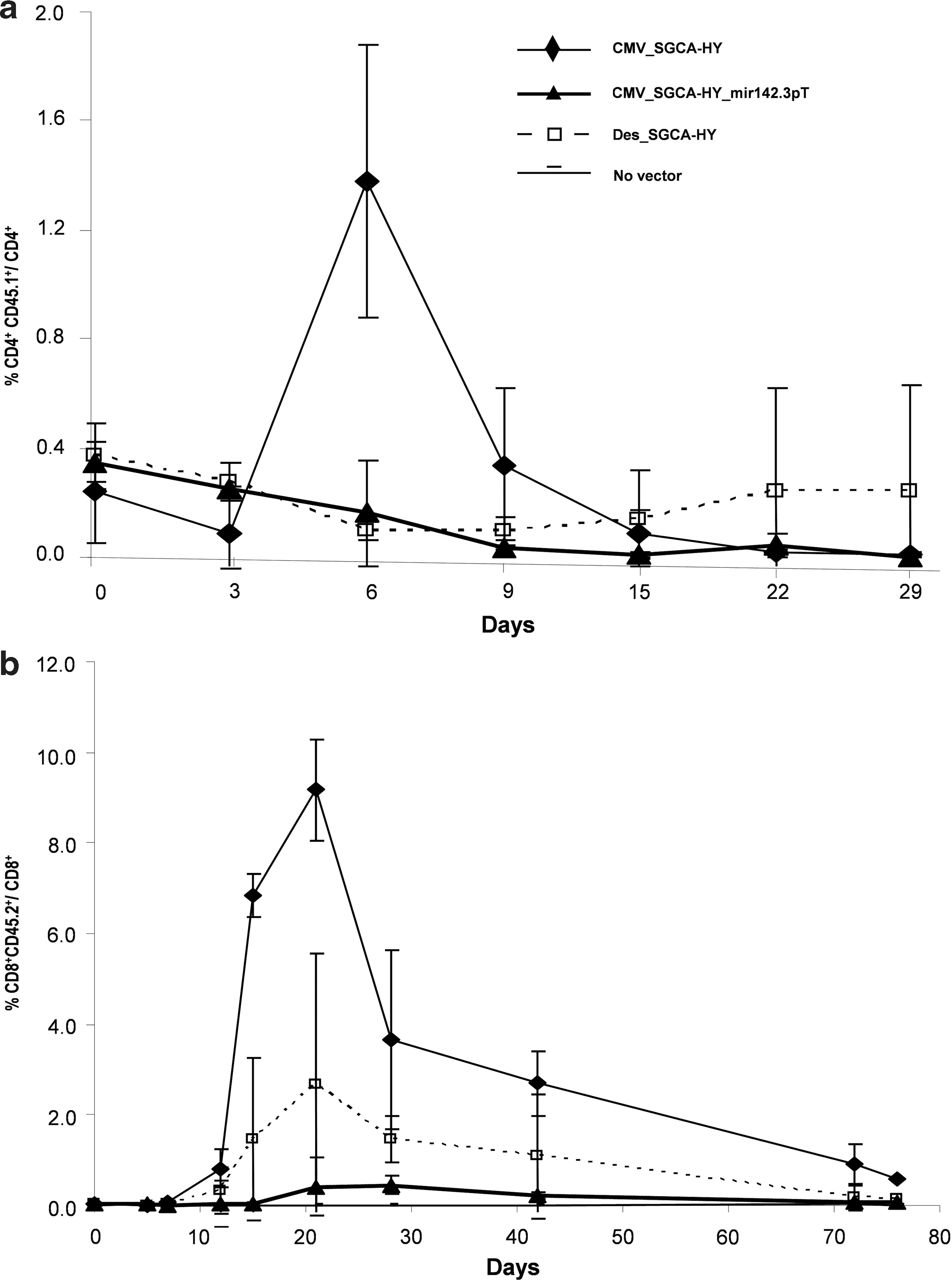
Adoptive transfer of HY-specific TCR transgenic CD4 and CD8 T cells.
Since the use of tissue-specific promoter can reduce immune responses induced by gene transfer (De Geest et al., 2003; Follenzi et al., 2004; Franco et al., 2005; Di Domenico et al., 2006), we tested whether or not the use of a desmin promoter described for muscle tissue–specific expression (Talbot et al., 2009) could impact T-cell responses in our system. The desmin_SGCA-HY rAAV1 vector induced remarkably less transgene-specific CD4 or CD8 T-cell expansion than the nonregulated CMV_SGCA-HY rAAV1 vector (Fig. 4b), showing that tissue-driven promoters can reduce the intensity of the T-cell immunization process. However, the desmin promoter did not completely control the induction of CD8 and CD4 T-cell responses since a small peak of amplification was detectable by day 21. This weak signal contrasted with the complete lack of response under mir142.3pT regulation (Fig. 4b).
The kinetics of CD4+ T -cell priming induced by IM gene transfer was assessed by measuring the division of adoptively transferred naive T cells in the spleen. The nonregulated vector induced an extensive division of the entire population of adoptively transferred Dby-specific CD4+ T cells within 6 days (Fig. 5), matching the blood CD4+ T-cell amplification curves (Fig. 4a). In contrast, the mir142.3p-regulated vector did not prime CD4 T cells as the great majority of adoptively transferred cells remained undivided in the spleen for at least 9 days (Fig. 5). The desmin-driven vector induced an intermediate and protracted response between the nonregulated and mir142.3p-regulated vectors. The desmin promoter reduced but did not completely prevent the induction of transgene-specific CD4+T cell responses.
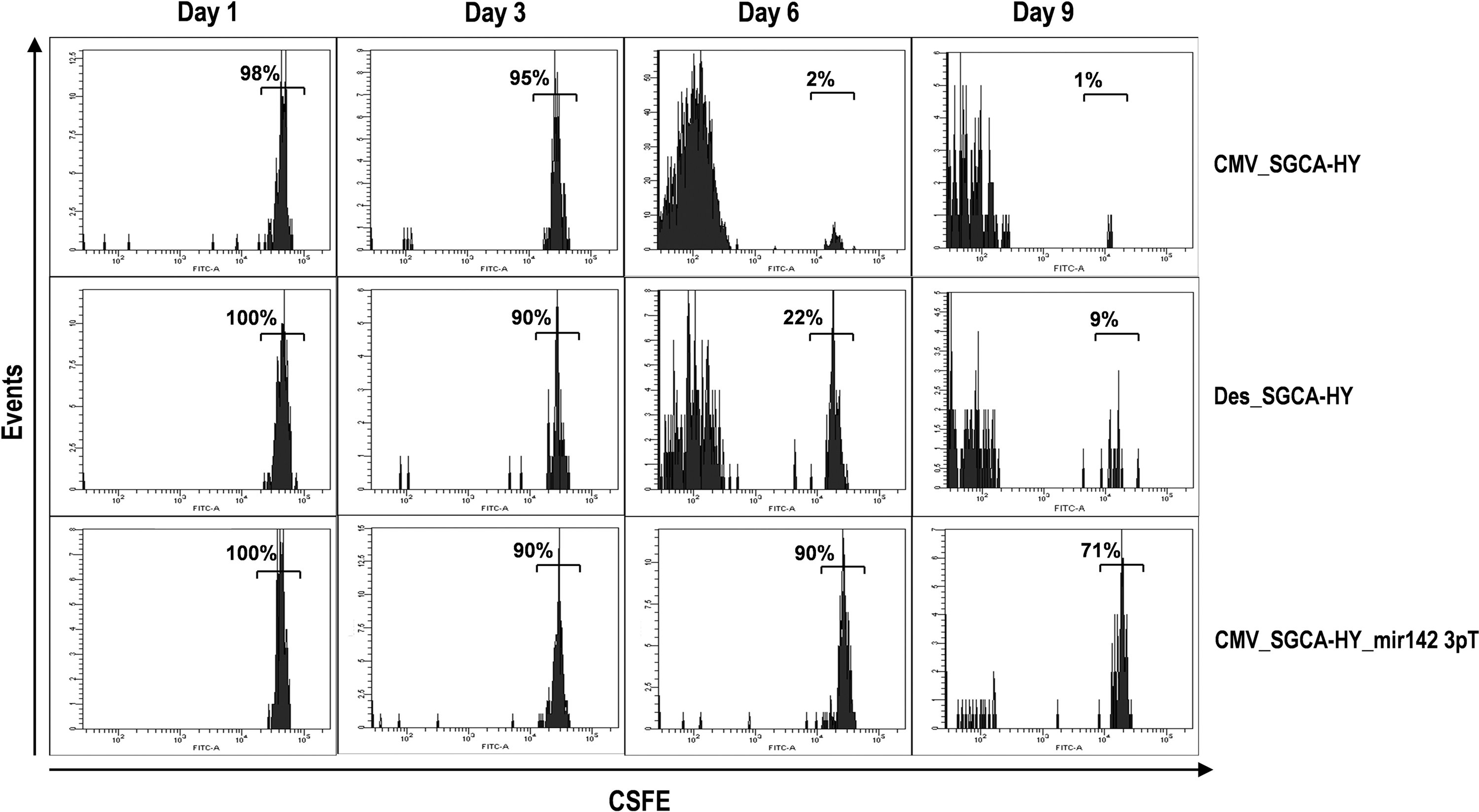
Division of transgene-specific CD4 T cells following vector administration. CFSE-labeled Marilyn CD4+ T cells were injected into C57Bl/6 mice, and the following day, the indicated vectors were injected IM into the TA. At indicated time points following vector injection, mice were sacrificed and spleen cells analyzed by FACS to determine the levels of CFSE staining on adoptively transferred cells.
Thus, various strategies preventing transgene expression in hematopoietic cells can reduce the priming of naive transgene-specific T cells. The use of mir142.3p-target sequences appears to be more effective than some muscle tissue-specific promoters for this purpose.
Induction of transgene persistence but without active immune tolerance
As shown in Figure 2, the mir142.3p-regulated vector provides long-term muscle transduction in C57Bl/6 mice. To test if such long-term persistence was due to the establishment of an active immune tolerance against epitopes of the transgenic protein, mice treated with the rAAV1 CMV_SGCA-HY_mir1423.pT vector were subsequently challenged 3 weeks later with the Dby peptide administered in complete Freund's adjuvant (CFA). Similar peptide challenge protocols have been used to demonstrate peripheral tolerance in mice (Liu et al., 2002). Upon such challenge, mice developed a robust Dby-specific IFN-γ+ CD4+T-cell response in the spleen, which was identical whether or not they had received initially the mir1423p-regulated vector, the nonregulated vector, or PBS (Fig. 6). Thus, the IM administration of the miR142.3p-regulated vector does not lead to a state of robust active immune tolerance.

The mir142.3p-regulated rAAV1 vector does not induce an active immune tolerance against the transgene. C57Bl/6 mice were injected IM with PBS or with 5×109 vg of rAAV1_CMV_SGCA-HY vector with or without mir142.3pT sequences. Twenty one days later, mice were challenged with PBS or Dby peptide emulsified in complete Freund's adjuvant
The mir142.3p-regulated vector fails to control T-cell responses in scga−/− mice and only temporarily prolongs muscle transduction
In sgca−/− mice, IM administration of the CMV_SGCA-HY rAAV1 vector led to a rapid clearance of gene-modified fibers with similar kinetics as in C57Bl/6 mice. By 2 weeks, the muscle of sgca−/− mice was infiltrated and transgene expression was reduced at the protein (Fig. 7a) and mRNA levels (Fig. 7b). By 4 weeks, the muscle was entirely remodeled as seen by extensive areas of centro-nucleated fibers, and SGCA mRNA was no longer detectable. In parallel, robust effector HY-specific CD4+ and CD8+ T-cell responses were measured at comparable levels than in C57Bl/6 mice (Fig. 7c and d). The administration of the mir142.3p-regulated vector effectively prolonged transgene expression in the muscle of sgca−/− mice compared to the nonregulated vector. After 2 weeks, the alpha-sarcoglycan protein immunostaining was clearly detected at the rim of myofibers accompanied by high levels of mRNA expression in muscle, thus contrasting sharply with the results obtained with the nonregulated vector at this time point. However, in contrast to C57Bl/6 mice, muscle transduction achieved with the mir142.3p-regulated vector in sgca−/− mice was only transient. The levels of alpha-sarcoglycan protein declined after 2 weeks. At 4 weeks, immunostainings showed persistent protein, but the pattern was abnormally diffuse in contrast to the well-structured membrane marking observed in normal muscle (Fig. 7a). The demise of muscle transduction was confirmed at 4 weeks by strongly reduced levels of SGCA mRNA, although some expression persisted at low levels over time (Fig. 7b). The induction of HY-specific CD4 and CD8 T-cell responses was evident following administration of the mir142.3p-regulated vector in sgca−/− mice, and levels were comparable to those induced by the nonregulated vector (Fig. 7c and d). Thus, in sgca−/− mice, the failure to maintain transgene expression over time with a mir142.3p-regulated CMV-driven construct, is clearly correlated to an uncontrolled T cell responses.
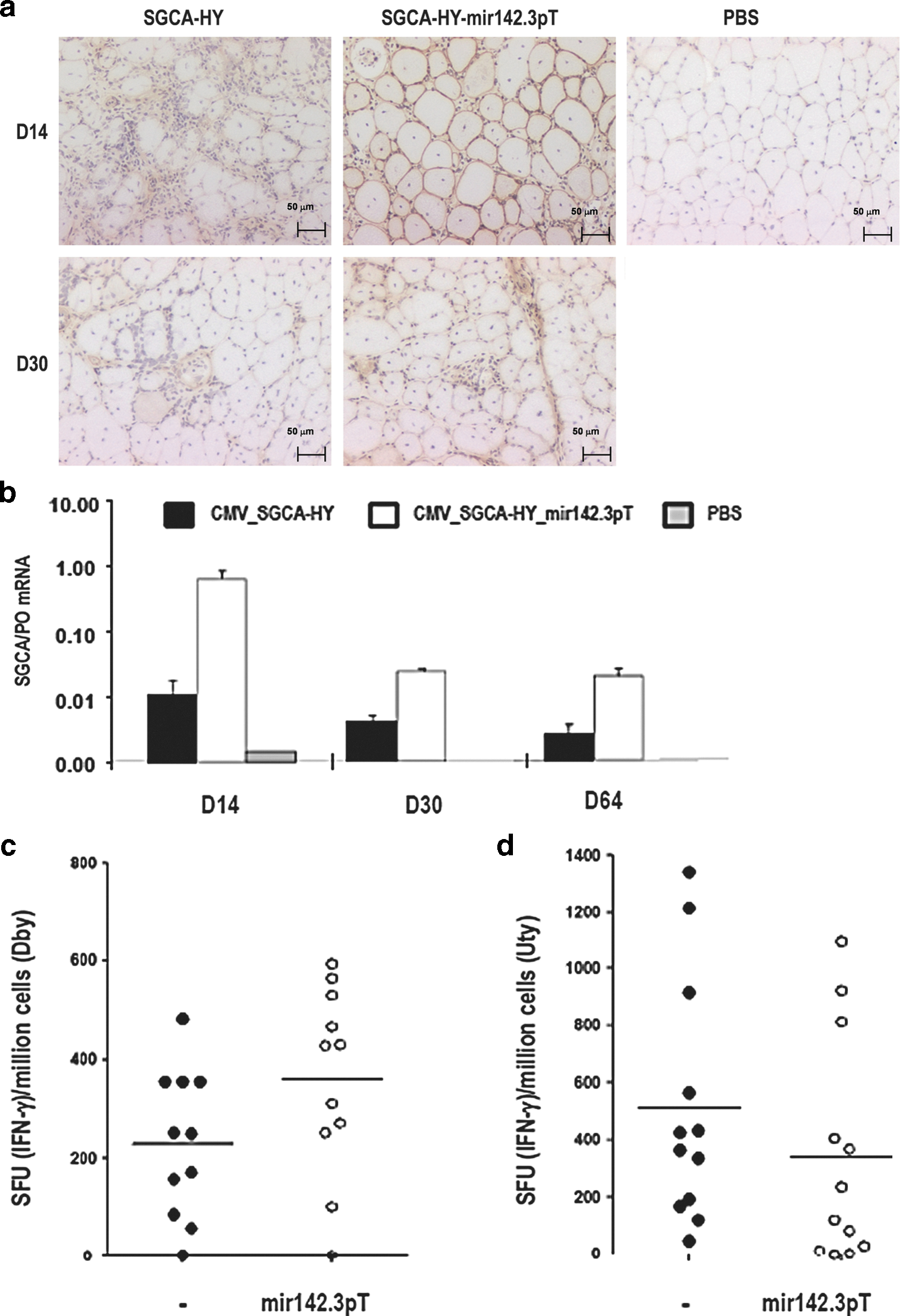
Transgene expression and transgene-specific immune responses in sgca−/− mice were injected IM in the TA with 5×109 vg of rAAV1_CMV_SGCA-HY vector with or without mir142.3pT sequences or with the same volume of PBS.
Since the CMV promoter-driven SGCA-HY transgene is strongly immunogenic, we tested if using a potentially less immunogenic desmin promoter-driven SGCA transgene (lacking the HY neoantigen tag) would provide long-term expression in scga−/− mice. Such desmin_SGCA vector induced initially high levels of transgene expression following IM injection in sgca−/− mice, as shown by the alpha-sarcoglycan immunohistochemical marking at day 14 (Fig. 8a). Yet, these levels did not persist over long periods of time as documented by an almost complete loss of transgene marking at day 56 (Fig. 8a), which was confirmed by qRT-PCR (data not shown). The loss of transgene was accompanied by CD8 T-cell infiltration in the injected muscles, which was already evident at day 14 (Fig. 8b). Inserting mir142.3p-target sequences in the desmin_SCGA vector seemed to improve gene transfer because a few alpha-sarcoglycan-positive fibers remained at day 56 (Fig. 8a). Nevertheless, this strategy was not sufficient to fully prevent the loss of transgene and the induction of CD8 T-cell responses, which were also apparent in the injected muscle from day 14 and later (Fig. 8a and b). These data confirm with a less immunogenic system than SGCA-HY that mir142.3p-regulated rAAV1 gene delivery in muscle is not as effective in sgca−/− mice than in C57Bl/6 mice.
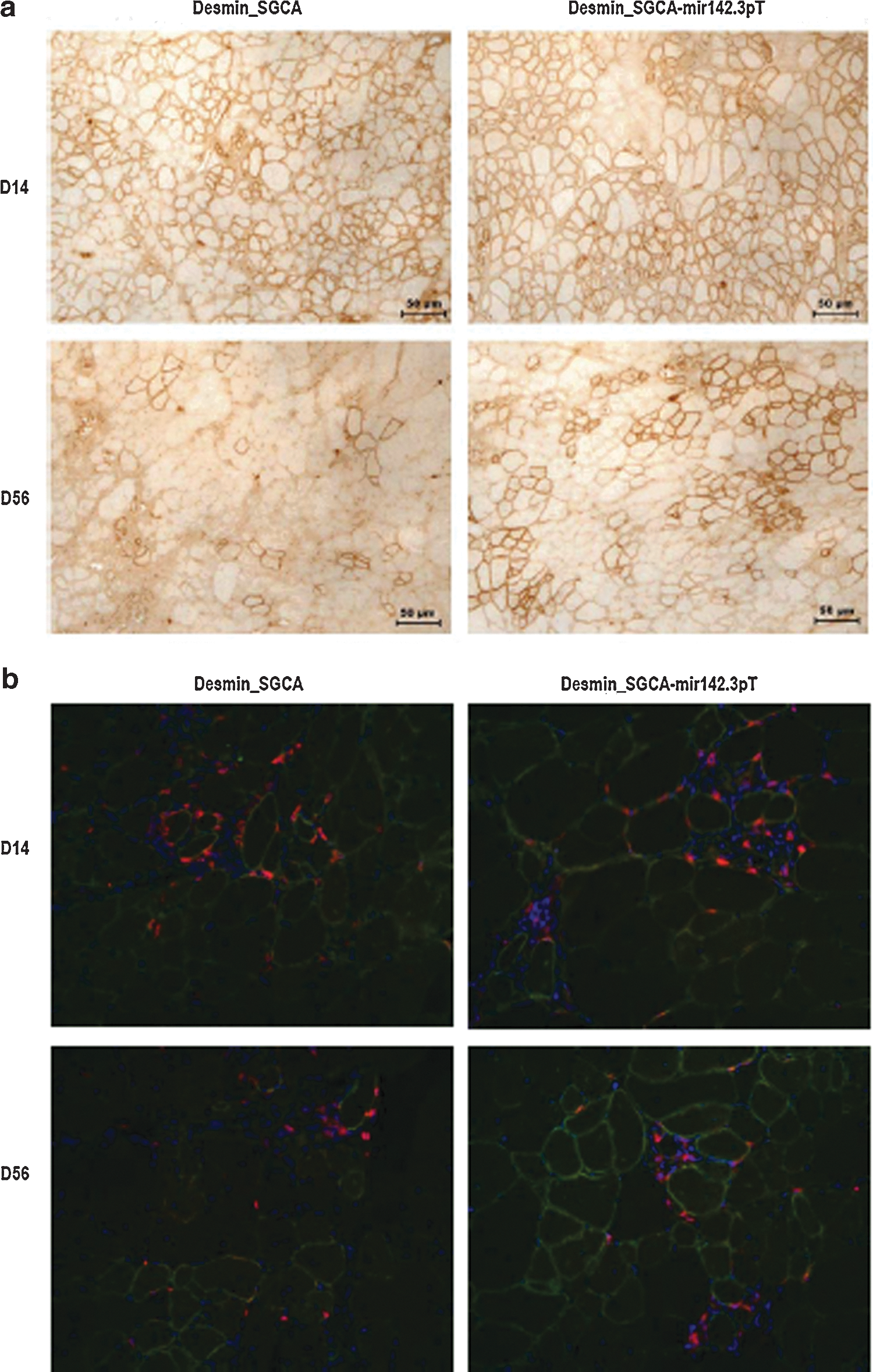
Failure to sustain long-term transgene expression in sgca−/− mice with the desmin_SGCA_mir142.3pT vector. The sgca−/− mice were injected IM in the TA with 5×109 vg of rAAV1_desmin_SGCA vector with or without mir142.3pT sequences.
Discussion
Efficient strategies are needed to prevent gene-specific immunization following rAAV gene delivery in vivo. Our results show that regulating transgene expression with mir142.3p target sequences can prolong muscle transduction and preserve the integrity of the treated muscle by preventing unwanted T-cell immunization, but this approach has some limitations in pathological models. The ability to investigate transgene-specific T-cell responses with a new artificial reporter transgene SGCA-HY has been a key factor to understanding the mechanisms of immunization by gene transfer in normal and pathological conditions. The development of both CD4 and CD8 T-cell responses could be monitored with high sensitivity in mice using HY-specific TCR transgenic T cells, peptides, and tetramers. The chimeric product was robustly immunogenic, in part because it is a transmembrane muscle protein known to more effectively immunize and promote muscle tissue destruction than other types of transgenic proteins expressed in muscle (Sarukhan et al., 2001). Both SGCA and HY are neoantigens in female mice. In addition, the IM delivery route for rAAV1 is also known to be strongly immunogenic (Toromanoff et al., 2010).
In such an immunogenic model, it is therefore remarkable that simply adding mir142.3p target sequences to the CMV-driven expression cassette could prevent the priming and development of transgene-specific CD4 and CD8 T-cell responses. This effect was indeed specific of the mir142.3p target sequences inserted in the vector, since we tested several other sequences that were targets of other microRNAs and found no regulation of immune responses against SGCA-HY (data not shown). The mir142.3p-regulation strategy has been successfully used in the context of liver-directed lentiviral vector gene transfer to prevent FIX immune responses in FIX-deficient mice and UGT1A1 immune responses in the Gunn rat model (Brown et al., 2007a; Schmitt et al., 2010).
We herein show that this strategy can also be applicable successfully with rAAV gene delivery in muscle and is at least as efficient, if not more, than tissue-specific promoters to prevent gene-specific T-cell immune responses. One advantage of mir142.3p-target sequences, which act at the post-transcriptional level, is the ability to regulate transgene expression and to reduce immunization while providing high levels of expression in permissive tissues through the use of strong transcriptional elements such as the CMV promoter. Muscle tissue–specific promoters can also function effectively to express the SGCA transgene. Recently, an rAAV1 with a muscle tissue–specific promoter (rAAV1.tMCK) was used in LGMD2D patients and demonstrated persistent gene expression in two of three patients for as long as 6 months (Mendell et al., 2010). In principle, tissue-specific promoters should achieve the same objective as mir142.3p-regulation, which is to prevent immunization by reducing expression of the transgene in the hematopoietic system. In our study, we tested the desmin promoter to drive SGCA-HY expression. In C57Bl/6 mice the desmin promoter was clearly effective to reduce immunization compared to the CMV promoter as shown by considerably lower levels of T-cell responses and lower muscle tissue destruction (data not shown). However, the desmin promoter was not sufficient to permit long-term transgene expression in sgca−/− mice following IM injection. Low levels of T-cell responses were detected. Possibly, such promoters may enable transgene expression in the immune system. In human cells, the desmin promoter, like the C512 promoter, is not strictly restricted to muscle but functions also in human APCs such as plasmacytoid dendritic cells (Veron et al., 2009), supporting this hypothesis. Even at low levels, the priming of a few T cells may be sufficient to reject the transgene over time, which could explain why muscle tissue–specific expression systems have failed to prevent gene-specific immunization in other contexts (Liu et al., 2004).
While mir142.3p-target sequences can improve rAAV1 gene transfer in normal muscle by reducing immune responses, there are important differences with other models and some limitations to consider. In our model, transgene expression is prolonged because the vector simply fails to prime CD4 and CD8 T-cell responses. We obtained no evidence for immunomodulation or tolerance induction. There was no elevation of IL-10, and we failed to detect the induction of significant levels of CD4+ FoxP3+ regulatory T cells in the spleen in response to the mir142.3p-regulated SGCA-HY delivered with rAAV1 (data not shown). More importantly, mice were not actively tolerized against the transgene.
The lack of gene-specific T-cell responses achieved with mir142.3p-regulated rAAV1 vectors in muscle could be broken by a robust antigenic challenge. In other models using liver gene transfer, it was shown that the use of mir142.3p-regulated LV prolongs transgene expression by reducing immune responses but also by establishing an active immune tolerance via the induction of CD4+CD25+FoxP3+ regulatory T cells (Annoni et al., 2009). The differences between our results and results from other studies is not surprising because different outcomes are expected from the use of different types of vector in liver or muscle due to specific properties of the microenvironment. Hepatic gene transfer with AAV vectors can induce a very robust immune tolerance involving regulatory T cells capable of preventing the development of CTL responses following immunological challenge with an adenovirus expressing the transgene (Dobrzynski et al., 2006). In the context of rAAV1 mir142.3p-regulated gene transfer in skeletal muscle, the lack of T-cell response against gene-modified cells appears to result essentially from immune ignorance and not from the induction of a state of active immune tolerance against the transgene.
One limitation of the mir142.3p-regulation strategy is the lack of efficacy in a dystrophic model. Administering the mir142.3p-regulated rAAV1 in the muscle of sgca−/− mice initially provides high levels of transduction but without persistence. Eventually, a protracted immune response develops against gene-modified cells. It is not clear what is causing the lack of control of immune responses in sgca−/− mice. These female mice are equally naive as C57Bl/6 female mice against the male HY neoantigen and respond with the same amplitude and kinetics to SGCA-HY. The use of SGCA devoid of the HY tag combined with a desmin promoter and mir142.3p target sequences constitutes a less immunogenic system, yet this was not sufficient to prevent T-cell responses and transgene loss in the sgca−/− murine model. It is possible that the mir142.3p could be abnormally expressed or functionally different in scga−/−, which are dystrophic animals.
Various types of muscular dystrophies in humans are associated with abnormal patterns of miRNA expression (Eisenberg et al., 2007). If this is the case, the selection of microRNAs differentially expressed in hematopoietic versus muscular cells in the context of muscular dystrophy could be considered. The lack of efficacy in dystrophic mice may simply reflect that various modalities of immunization take place following gene transfer in different contexts. Preventing transgene expression in hematopoietic cells with mir142.3p-target sequences is designed to reduce direct antigenic presentation of the transgene by directly controlling transgene expression in professional APCs (Brown et al., 2006). This strategy will work only if direct antigenic presentation is the sole modality of immunization, and this seems to be the case following rAAV1 IM delivery in normal muscle. Indeed, such results suggest that rAAV1 is able to transduce professional APCs in vivo, even though rAAV1 may not be the most effective serotype as shown in vitro with the transduction of cultured dendritic cells (Xin et al., 2006; Lu and Song, 2009). However, the mir142.3pT strategy is not expected to prevent indirect antigenic presentation occurring from the uptake of transgenic proteins from the environment. Because dystrophic sgca−/− mice have weakened myofibers and their skeletal muscle is inflamed (Fougerousse et al., 2007), indirect antigenic presentation may occur more extensively than in normal mice and could significantly contribute to T-cell immunization. Further studies are needed to explore these mechanisms. Yet, the limitations identified with our model may explain in part why mir142.3p-regulated constructs have not been universally effective in all systems. This approach was not found to be efficient to prevent the loss of transgene expression in the case of plasmid gene delivery (Wolff et al., 2009) or in the case of rAAV9 or rAAV8 gene delivery to liver (Qiao et al., 2011; Cotugno et al., 2012). Thus, different outcomes are expected from the use of mir142.3p-regulated expression cassettes according to the target tissue, environment, and possibly other factors.
While the efficacy of mir142.3p-regulated constructs to prevent transgene-specific immunization may not be universal, there are some circumstances in which the possibility to avoid immune system activation with mir142.3p-regulated rAAV1 gene transfer could benefit certain gene therapy applications. This approach could be used as an alternative to tissue-specific promoters, or more probably in combination with other strategies, to facilitate long-term muscle transduction. There are also multiple therapeutic applications based on protein delivery platforms in normal muscle that could benefit from the use of mir142.3p-regulated vectors.
Footnotes
Acknowledgments
We are grateful to Genethon support groups for help with animal care, animal experiments, histology, and rAAV vector production. We thank Marc Bartoli (DDC group) and Smahane Chalabi (information technology) for discussions. We are also indebted to O. Lantz and K. Campbell for providing mice and acknowledge the support from AFM (French Muscular Dystrophy Association) and from the EC FP7-HEALTH-2007 project PERSIST (GA 222878). FB and DAG conceived, supervised, performed experiments, and wrote the manuscript; MF, JP, and SD performed experiments; IR provided reagents and supervision; and AG conceived and directed the study and wrote the manuscript.
Author Disclosure Statements
The authors have no conflicts of interest to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.