
Abstract
On the basis of previous studies suggesting that vascular endothelial growth factor (VEGF) could protect motor neurons from degeneration, adeno-associated virus vectors (serotypes 1 and 9) encoding VEGF (AAV.vegf) were administered in a limb-expression 1 (LIX1)-deficient cat—a large animal model of lower motor neuron disease—using three different delivery routes to the central nervous system. AAV.vegf vectors were injected into the motor cortex via intracerebral administration, into the cisterna magna, or intravenously in young adult cats. Intracerebral injections resulted in detectable transgene DNA and transcripts throughout the spinal cord, confirming anterograde transport of AAV via the corticospinal pathway. However, such strategy led to low levels of VEGF expression in the spinal cord. Similar AAV doses injected intravenously resulted also in poor spinal cord transduction. In contrast, intracisternal delivery of AAV exhibited long-term transduction and high levels of VEGF expression in the entire spinal cord, yet with no detectable therapeutic clinical benefit in LIX1-deficient animals. Altogether, we demonstrate (i) that intracisternal delivery is an effective AAV delivery route resulting in high transduction of the entire spinal cord, associated with little to no off-target gene expression, and (ii) that in a LIX1-deficient cat model, however, VEGF expressed at high levels in the spinal cord has no beneficial impact on the disease course.
Introduction
In this study, we tested the therapeutic effect of the vascular endothelial growth factor (VEGF), a neuroprotective factor, in LIX1-deficient cats using three different gene delivery routes. Initially described as one of the major regulators of blood vessel formation (Ferrara et al., 2003), the VEGF was also shown to induce antiapoptotic and neuroprotective activities both in vitro and in vivo (Greenberg and Jin, 2005; Storkebaum et al., 2005; Sathasivam, 2008). Moreover, intramuscular injections of a VEGF-encoding lentiviral vector increased the life expectancy of a mouse model of ALS (Azzouz et al., 2004), and reduced levels of VEGF seem also to predispose mice and human to MN degeneration (Oosthuyse et al., 2001; Lambrechts et al., 2003; Devos et al., 2004; Brockington et al., 2006). Adequate spinal cord delivery of therapeutic factors represents another critical challenge to develop treatment for MN diseases, especially because the blood–brain barrier prevents penetration of a majority of therapeutics into the central nervous system (CNS) (Miller et al., 1996; Borasio et al., 1998; Ochs et al., 2000; Lauria et al., 2009). Recombinant adeno-associated virus (AAV) was shown to successfully deliver transgenes to the MNs in mice and large animal models after intramuscular (Kaspar et al., 2003; Hollis et al., 2008; Towne et al., 2010) and, more recently, intravenous (IV) injections (Duque et al., 2009; Foust et al., 2010; Bevan et al., 2011; Gray et al., 2011b), resulting in prolonged survival of mouse models of SMA (Foust et al., 2010; Valori et al., 2010; Dominguez et al., 2011). Direct intracranial injections of AAV vectors in structures with spinal cord connections, such as the motor cortex, although invasive can also deliver a therapeutic factor to the spinal cord by an anterograde transport of AAV vectors (Dodge et al., 2008; Foust et al., 2008; Ciron et al., 2009). Finally, AAV intrathecal injections have been shown to mediate transgene expression in the spinal cord of mice (Storek et al., 2008; Vulchanova et al., 2010; Gray et al., 2011a) and pigs (Bevan et al., 2011; Federici et al., 2011) and potentially constitutes another efficient and less invasive gene therapy delivery route.
The purpose of this study was to test the therapeutic action of VEGF overexpression in the spinal cord of LIX1 cats using three different modes of AAV delivery with two AAV serotypes (AAV1 and AAV9) known for their neurotropism in cats (Vite et al., 2003; Duque et al., 2009). These two AAV serotypes were injected into the motor cortex, the cisterna magna, or the jugular vein of young adult cats using an identical dose (1×1012 viral genome [vg]/kg). The biodistribution of AAV vectors and VEGF expression in the spinal cord was analyzed in heterozygous lix1 +/− cats at 3 months postinjection (pi), while the therapeutic efficacy of each gene transfer strategy was evaluated in homozygous affected lix1 −/− cats until 6 months pi. We found that intracisternal (ICis) injections of scAAV9 vector achieved sustained expression of the secreted neurotrophic factor both in the spinal cord and in the nerves while avoiding off-target expression in the liver. However, high and long-term expression of VEGF in the spinal cord did not prevent loss of motor axons and muscle denervation in lix1-deficient cats.
Materials and Methods
Animals
Breeding LIX1 cats (heterozygous lix1 +/− and homozygous lix1 −/− animals) were obtained from John Fyfe (Laboratory of Comparative Medical Genetics, East Lansing, MI) and housed in the Boisbonne Center at Nantes Veterinary School (Oniris, Nantes, France). Kittens were genotyped as previously described (Fyfe et al., 2006). Experiments were approved by the regional ethics committee and were carried out according to European guidelines for the care and use of experimental animals.
Vectors
Pseudotyped AAV1 and AAV9 vectors were generated by packaging AAV2-based recombinant single-strand (ss) or self-complementary (sc) genomes into AAV1 and AAV9 capsids, respectively, as previously described (Rabinowitz et al., 2002) at the University Hospital of Nantes. The AAV2 plasmids contain (i) the gene-encoding green fluorescent protein (GFP) under control of the cytomegalovirus promoter in an sc genome or (ii) the gene encoding murine VEGF under control of the phosphoglycerate kinase promoter in an ss or sc genome. Physical particles were quantified by dot blot hybridization, and vector titers are expressed as vector genomes per milliliter.
In vivo AAV injections
For intracerebral (ICra) injections (Table 1), solutions of ssAAV1 encoding VEGF (ssAAV1.vegf) were bilaterally injected into both the internal capsule and the motor cortex of 7-week-old cats, under general anesthesia obtained by isoflurane inhalation (3% v/v) and morphine injection (0.2 mg/kg). All treated cats received 3 deposits of ssAAV1.vegf (2×15 μl in motor cortex and 20 μl in internal capsule) through 3 stereotactic tracks in each hemisphere (motor cortex: 5 mm caudal to the supraorbital arch and 29 mm rostral to the auditory canal, 3 and 5 mm lateral to the sagittal suture, depth 3 mm; internal capsule: 21 mm caudal to the supraorbital arch and 13 mm rostral to the auditory canal, 8 mm lateral to the sagittal suture, depth 8.5 mm, flow 2 μl/min). No side effect possibly related to surgery or vector deposits was noticed. For ICis injections (Table 1), solutions of both ssAAV1 and scAAV9 encoding VEGF or GFP were injected into the cisterna magna of 7-week-old cats (1×1012 vg/kg in 1 ml/kg per cat) under general anesthesia obtained by isoflurane inhalation (3% v/v). For IV injections (Table 1), scAAV9.vegf was injected into the jugular vein of 7-week-old or 2-day-old kittens (respectively, 1×1012 and 1×1013 vg/kg in a volume of 1–2 ml). Seven-week-old cats receiving ICra, ICis, or IV AAV deliveries were immunosuppressed by mycophenolate mofetil (20 mg/kg/day) combined with cyclosporine, whose dosage was adjusted once a week in order to maintain cyclosporinemia above 100 ng/ml. Before injections of the AAV vectors, the absence of AAV1 or 9-neutralizing antibodies was confirmed in serum of 7-week-old cats and in mothers when neonatal kittens were injected. A unique control group made of three uninjected affected lix1−/− cats (C22–C24) and one healthy lix1+/− cat (C21) was constituted, common to all injected cats (Table 1). This was made because of the restricted availability of lix1−/− cats.
AAV, adeno-associated virus; C, cat; CyA, cyclosporine A; MMF, mycophenolate mofetyl; sc, self-complementary; ss, simple strand; vegf, vascular endothelial growth factor; vg, vector genome.
Tissue preparation
Respectively, 100 and 200 days after ICra, ICis, or IV AAV.vegf injections, heterozygous lix1 +/− and homozygous lix1 −/− animals were anesthetized by isoflurane inhalation (3% v/v) and euthanized by an overdose of pentobarbital administered in the cephalic vein. Brain and spinal cord tissues were removed, embedded in O.C.T. Tissue Tek (Sakura Finetek, Torrance, CA), and frozen in cold isopentane (−50°C), and serial sections were cut on a cryostat (3×100 μm followed by 5×10 μm) and stored at −80°C for further analysis.
Cats receiving ICis injections of scAAV9.gfp were anesthetized 30 days after injection (150 μg/kg medetomidine, 10 mg/kg ketamine) and perfused transcardially with 10 ml phosphate buffered saline (PBS) followed by 100 ml 4% paraformaldehyde. Brains and spinal cords were removed and cut into coronal 5 mm blocks, postfixed by incubation in 4% paraformaldehyde, and cryoprotected by overnight incubation in 30% sucrose. Samples were embedded in optimal cutting temperature compound and frozen on dry ice. Spinal cord slices were cut on a cryostat in intervals of 100 μm followed by 5×10 μm.
Quantitative polymerase chain reaction analysis
Genomic DNA was extracted from frozen tissues using the following buffer: 50 mM Tris pH 8, 100 mM EDTA, 1% sodium dodecyl sulfate, and 100 mM NaCl containing 500 μg/ml proteinase K (Eurobio, Les Ulis, France). Samples were digested overnight at 55°C and then subjected to phenol-chlorophorme extraction. Real-time polymerase chain reaction (PCR) was conducted in duplicate with a LightCycler 2.0 instrument (Roche Diagnostics, Rotkreuz, Switzerland) using 50 ng of DNA in a 20 μl volume of the following solution: 10 μl Premix ExTaq (Takara Bio Inc., Shiga, Japan), 0.4 μl of 10 μM Primer, 0.4 μl of 10 μM FAM/TAMRA TaqMan probe, 5 μl template, and 3.8 μl H2O. The feline β-glucuronidase (βGLU) gene was employed as a reference, and for each sample, Ct values were compared to those obtained with plasmid standard dilutions containing mVEGF and βGLU cDNA sequences. The ratio between transgene (VEGF sequence) and genomic DNA (βGLU sequence) copy number provided the amount of transgene copy per cell. Primers and probe for mVEGF were as follows: forward primer 5′-CTTTCTCTCCACAGGTGTCCA-3′, reverse primer 5′-CAGTAAAGCCAGGGTCCA-3′, probe 6FAM 5′-CCTCCGAAACCATGAACTTTCTG-3′-TAMRA, and for feline βGLU: forward primer 5′-ACGCTGATTGCTCACACCAA-3′, reverse primer 5′-CCCCAGGTCTGCTTCATAGTT-3′, probe 6FAM 5′-CCGGCCCGTGACCTTTGTGA-3′-TAMRA. Samples were heated at 95°C for 20 sec followed by 45 cycles of 95°C for 10 sec and 60°C (βGLU) or 66°C (mVEGF) for 40 sec. Control tissues extracted separately from vector-treated tissues were checked for VEGF amplification. The sensitivity of the quantitative PCR is 10−4 copies/cell or 10 copies/50 ng of genomic DNA.
Reverse transcriptase PCR analysis
The RNA was isolated from frozen brain and spinal cord tissues with Trizol Reagent (Invitrogen, Carlsbad, CA) and treated with DNAse (Turbo DNA-free; Ambion, Austin, TX). About 1 μg of RNA from each sample was reverse transcribed using MMLV reverse transcriptase (Invitrogen), and PCR proceeded using the cDNA as a template with AmpliTaq Gold DNA Polymerase (Applied Biosystems, Foster City, CA). The amplification was done for 40 cycles (94°C for 30 sec, 62°C for 1 min) with specific primer pairs for VEGF (forward primer 5′-GCCCTCCGAAACCATGA-3′, reverse primer 5′-AGCAGCCTGCACAGCG-3′) or β-actin primers (forward primer 5′-TGACGGGGTCACCCACACTGTGCCCATCTA-3′, reverse primer 5′-CTAGAAGCATTTGCGGTGGACGATGGAGGG-3′). Appropriate no-reverse transcripted and no-template controls were included in each PCR to confirm the specificity of the reaction. The sensitivity of the reverse transcriptase (RT)-PCR is 10 copies of vegf cDNA/1 μg RNA.
VEGF enzyme-linked immunosorbent assay analysis
Quantitative analysis of VEGF levels was performed using a mouse VEGF immunoassay kit (Quantikine; R&D Systems, Lille, France). About 300 μm brain slices and 5-mm-long spinal cord blocks, frozen at −80°C, were homogenized in a solution of tris-buffered saline containing 1% Triton X-100 (Sigma, St. Louis, MO) containing a cocktail of protease inhibitor (Complete Mini; Roche, Meylan, France). The lysates were then centrifuged for 5 min at 4°C, supernatants were collected, and enzyme-linked immunosorbent assay was performed following the manufacturer's protocol. Protein concentration was assessed using the Protein Assay Kit (Biorad, Hercules, CA). Data are expressed as pg of VEGF per mg of total proteins. The sensitivity of the enzyme-linked immunosorbent assay (ELISA) test was estimated to be 1 pg/mg of total proteins.
GFP expression in the spinal cord and in the brain
The brain and spinal cord sections were examined for GFP expression using a laser scanning confocal microscope (Nikon, C1, Champigny sur Marne, France) equipped with a blue argon ion laser at 488 m. For choline acetyltransferase (ChAT) and NeuN immunofluorescence, the brain and spinal cord sections were blocked, respectively, by incubation with 1% rabbit or 5% goat serum (Dako, Glostrup, Denmark) in 0.4% Triton X-100 in PBS and incubated overnight at room temperature with a goat polyclonal anti-ChAT antibody (1:100, AB144P; Chemicon International) or a monoclonal anti-NeuN antibody (1:8000, clone A60; Millipore); the sections were then incubated with a biotinylated rabbit anti-goat antibody (Dako) followed by incubation with Alexa fluor 555–conjugated streptavidin (Invitrogen) or an Alexa fluor 555 goat antimouse antibody (Molecular Probes).
Sections were then mounted in Mowiol (Calbiochem, San Diego, CA). The immunolabeled sections were scanned serially using the argon ion laser (488 nm) to observe GFP signals, and with a helium neon laser (543 nm) to observe Alexa fluor 555 signals. Each image was recorded in a separated channel (channel green for GFP and channel red for Alexa fluor 555) and overlayed to allow detection of colocalized fluorescent signals.
Neurological and electromyographic evaluation of the feline SMA disease
To evaluate the severity and the course of the feline SMA disease, neurological examinations were weekly performed in injected animals compared with three homozygous lix1 −/− affected kittens and three heterozygous lix1 +/− nonaffected kittens as recently described (Wakeling et al., 2011). Briefly, locomotion, limb position, muscular atrophy, postural reactions, and reflexes were scored according to severity as follows: 0, normal; 1, moderately affected; 2, clearly affected; 3, severely affected. The scores obtained at different ages for each animal allowed us to follow the disease evolution in the affected kittens.
Additionally, EMG examinations were achieved every 2 weeks in affected and normal carrier animals with a Neuropack 2 (Nihon Kohden, Tokyo, Japan) as previously described (Wakeling et al., 2011). The biceps femoris, sartorius, tibialis anterior, gastrocnemius, and plantaris muscles were tested. Briefly, for each muscle, a global score accounting for the presence and the frequency of fibrillations (0, absence; 1, presence in some regions; 2, presence in multiple regions; 3, presence in all regions tested), abnormal insertion potentials (0, absence; 1, presence), and high-frequency discharges (0, absence; 1, presence) was assigned.
Clinical examinations were completed by motor axon count and motor axon morphometric analysis in the ventral root of the spinal cord at the C6 level in injected C13 lix1 −/− cat, as previously described (Wakeling et al., 2011). The results were compared with MN axon counts and morphometric analyses previously obtained in cohorts of affected and healthy cats of the same age.
VEGF Western blot and Miles test
After transfection of HEK293 cells with the plasmid vector containing the mVEGF164 cDNA fused in the C-terminus with a cmyc epitope or the GFP cDNA as control, 50 μg of total proteins for the GFP extracts and 2.5, 5, and 10 ng of VEGF protein as measured by ELISA for the VEGF supernatant were transferred to a nitrocellulose membrane (Hybond; Amersham, Les Ulis, France) and blocked with a solution of PBS supplemented with 5% nonfat dry milk, 1% NP-40, and 0.1% Tween-20. Membranes were then immunoblotted for 2 hr with rabbit polyclonal anti-cmyc antibody (Abcam ab9106) diluted at 1:15,000 and next incubated with peroxidase-labeled antirabbit IgG polyclonal goat antibody (Dako PO448; 1:15,000) for 45 min. All membranes were visualized using ECL and exposure to ECL Hyperfilm.
In parallel to the detection of the VEGF protein by Western blot, we verified that the recombinant murine VEGF (Peprotech, Levallois Perret, France) and the murine VEGF protein produced by our AAV-vector had a functional effect in vivo in cats. For this purpose, we transduced HEK293 cells (multiplicity of infection: 105) with the AAV9-VEGF vector, verified the expression of the murine VEGF protein in the culture supernatant by a VEGF-specific ELISA test, and subsequently performed Miles assay in a cat. Miles test consisted of intravenously injecting 3% Evans blue dye (1 ml/kg) and 10 min later in the dermis 0.1 ml of PBS, of recombinant murine VEGF at 10, 50, and 150 ng, of histamine at 1 μM, or the culture supernatant obtained after transduction of HEK293 cells with scAAV9-VEGF, scAAV9-VEGF-cmyc, and scAAV9-GFP vector as control.
Statistical analyses
Data were expressed as means±SEM. Differences between the vector groups or between the treated and nontreated groups were analyzed using two-way ANOVA. p-Values<0.05 were considered significant.
Results
ICrs injection of ssAAV1.vegf results in widespread detection of the vector genome in the spinal cord and in the peripheral nerves
Since AAV vector is capable of anterograde transport (Dodge et al., 2008; Foust et al., 2008; Ciron et al., 2009) and ss AAV1 was shown to mediate high transgene expression in the cat brain after intraparenchymal injection (Vite et al., 2003; Duque et al., 2009), ICra delivery of ssAAV1 encoding the mouse VEGF was tested in cats from the LIX1 strain. Using the Miles assay, that is, Evans blue leakage in the skin (Murohara et al., 1998; Robinson et al., 2004; Gozes et al., 2006), we demonstrated that the murine VEGF was effective in cats (Supplementary Fig. S1; Supplementary Data are available online at
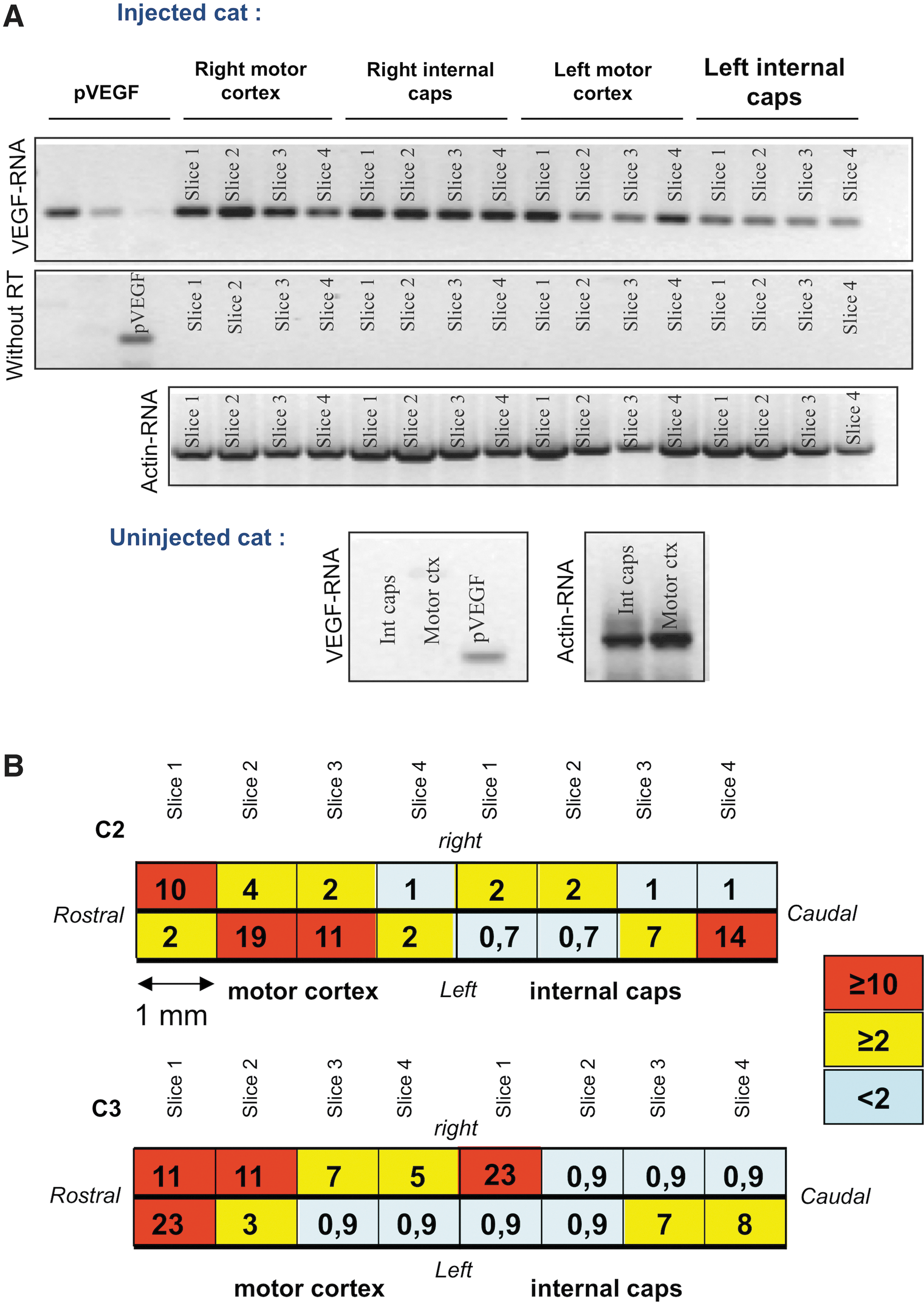
Intracerebral injections of ssAAV1.vegf result in VEGF expression in the motor cortex and in the internal capsule. Results of VEGF transcript and protein detection in the cortex. Each injection site was cut into 100 μm slices for PCR, RT-PCR, and ELISA analyses; each analysis was performed every millimeter.
We further tested the distribution of the vector genome in the spinal cord and in the peripheral nerves both by vegf-specific quantitative PCR and by vegf-specific PCR followed by Southern blot, whose sensitivity is higher than that of classic quantitative methods (sensitivity: 10 copies of vegf cDNA/1 μg of genomic DNA for the PCR–Southern blot and 200 copies of vegf cDNA/1 μg of genomic DNA for the quantitative PCR). In all ICra injected animals, the viral genome was detected by PCR and Southern blot (Fig. 2A) all along the spinal cord from the cervical to the lumbar part, in the ventral, as well as in the dorsal sections and by quantitative PCR (mean of 0.02±7×10−3 copies/cell for C1 and C2, <10−4 copies/cell for the uninjected cats; Fig. 3A). Vector genomes were also detected in the peripheral nerves but only by PCR and Southern blot (Fig. 2C). Consistent with these data, vegf-specific transcripts were found in the different regions of the spinal cord (Fig. 2B). However, despite the presence of the vector genome in the spinal cord, the VEGF protein was not detected; that is, values obtained were similar to those of the uninjected animals. Accordingly, no beneficial therapeutic effect of ssAAV1.vegf ICra injections was seen by neurological and EMG tests performed as previously described (Wakeling et al., 2011; data not shown) in the three lix1 −/− animals (C3–C5) compared with uninjected lix1 −/− animals (C22–C24; Table 1).

Intracerebral injections of ssAAV1.vegf result in a diffusion of vector DNA and RNA transcripts in the spinal cord and in the nerves.
Intracisternal is superior to ssAAV1 intracerebral injection, and intracisternal scAAV9 is superior to ssAAV1 for VEGF delivery to the spinal cord.
ICis delivery of ssAAV1.vegf results in detection of the VEGF protein in the spinal cord
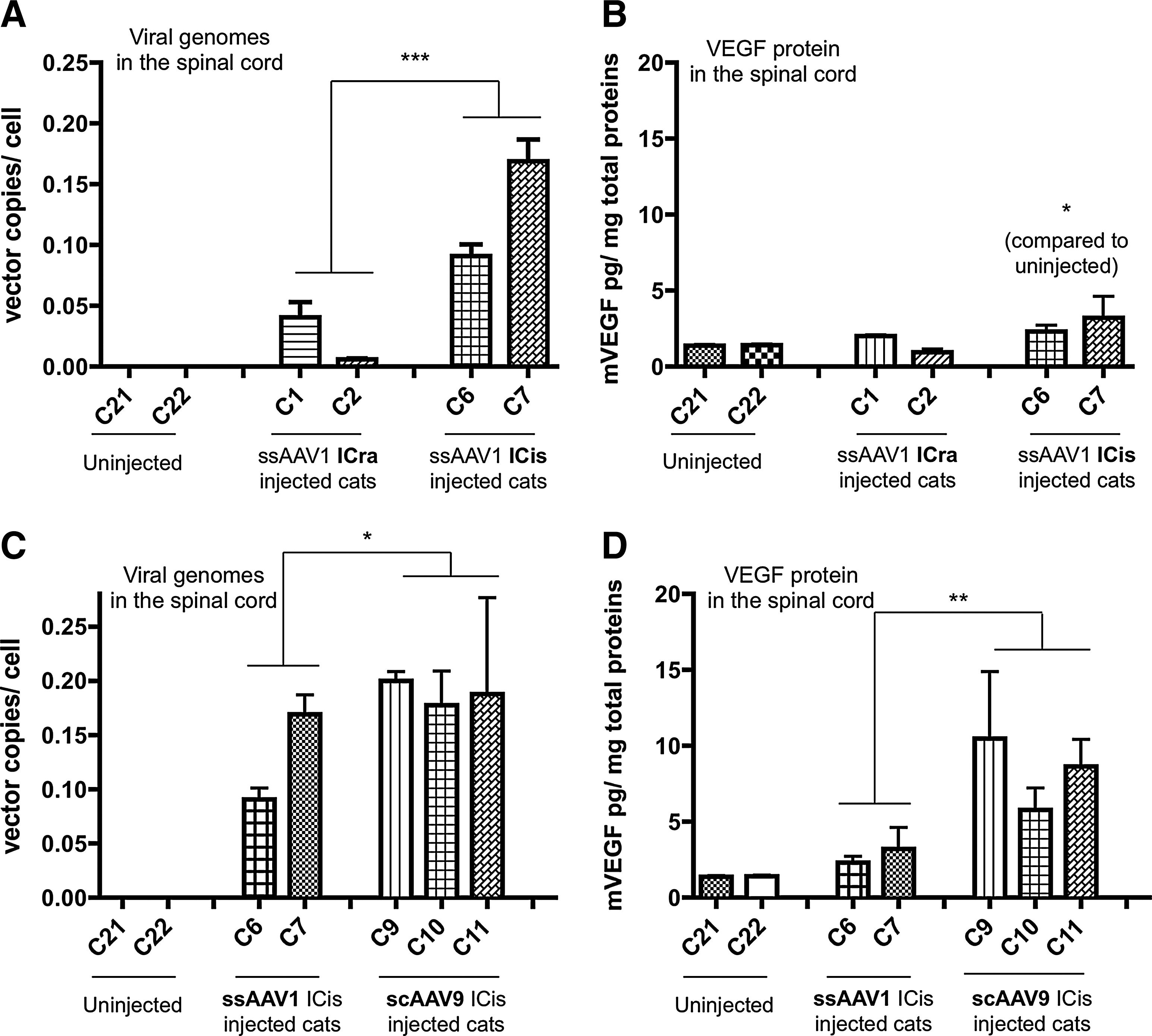
Since intrathecal delivery is less invasive and was previously reported to target MNs in mice and in pigs (Bevan et al., 2011; Federici et al., 2011; Gray et al., 2011a; Snyder et al., 2011), we tested ssAAV1.vegf vector into the cerebrospinal fluid (CSF) via ICis delivery. Two lix1 +/− cats (C6 and C7) were injected in the cisterna magna with ssAAV1.vegf (1×1012 vg/kg) and transgene expression was investigated 100 days later. Compared with ssAAV1 ICra injections (see above), ICis injections of ssAAV1.vegf led to a 5–6-fold increase in the number of viral copies per cell (p<0.001, mean of 0.12±0.01 copies/cell for C6 and C7; Fig. 3A) and to the significant detection (p<0.05) of the VEGF protein in the spinal cord (mean over the entire spinal cord of 2.69±0.64 pg/mg for C6 and C7; Fig. 3B). In contrast, the level of brain transduction especially in the motor cortex and internal capsule regions was lower after ICis delivery compared with direct intracortical injections of the vector (mean of 0.27±0.09 copies/cell in the motor cortex and internal capsule for C6 and C7, and 5±2.3 copies/cell in these regions for C1–C5; data not shown).
ICis delivery of scAAV9 is superior for VEGF expression in the spinal cord
Because sc AAV serotype 9 vector was recently described as efficient in transducing the MNs of various animal species (Duque et al., 2009; Foust et al., 2009; Foust et al., 2010; Valori et al., 2010; Bevan et al., 2011; Dominguez et al., 2011; Gray et al., 2011b; Glascock et al., 2012), we injected 1×1012 vg/kg of scAAV9 intracisternally in 7-week-old lix1 +/− cats (C9–C11). One hundred days later, we found a moderate increase in the number of vector copies per cell in scAAV9-injected cats compared with ssAAV1-injected animals (respectively, C9–C11 cats vs. C6 and C7; p<0.05) (Fig. 3C) associated with an increase of VEGF expression in the spinal cord (p<0.01) (mean of 8.33±1.34 pg/mg of total proteins), which was 6 times higher than the level measured in uninjected cats and ∼3 times superior than the level measured in ssAAV1-injected cats (Fig. 3D). Distinct cell tropism, which could influence vector expression in the spinal cord, may partly explain these different results concerning both vector genomes and transgene expression levels after AAV1 and AAV9 ICis delivery. Importantly, cat C12, an lix1 −/− animal euthanized 200 days post-scAAV9 injection, showed sustained and comparable high-level VEGF expression (Fig. 4B), suggesting that ICis injection of scAAV9 is an efficient strategy to mediate long-term therapeutic gene expression in the feline spinal cord. Lastly, injecting 10 times more scAAV9 vector (1×1013 vg/kg) in one affected lix1 −/− cat (C13) (Fig. 4A and B) resulted in ∼4–5 times more vector genome per cell (0.93±0.21 copies/cell) and ∼5 times more VEGF expression (43.1±18 pg/mg of total proteins) over the 1×1012 vg/kg dose of scAAV9 in the spinal cord after 200 days pi. Yet, despite this high level achieved (∼30-fold more than the uninjected cat level) and long-term expression of VEGF in the spinal cord, no phenotypic benefit was seen in the two affected cats (C12 and C13) (Supplementary Fig. S2). These results were confirmed by MN axon counts in the ventral root at the C6 level of the spinal cord of the injected C13 lix1 −/− cat and axon morphometric analysis that showed no increase of the number of motor axons, especially of large caliber (>10 μm) in VEGF-injected C13 cat compared with uninjected affected cats of the same age (Supplementary Fig. S3).
Intracisternal is superior to intravenous scAAV9 injection for long-term VEGF delivery to the spinal cord.
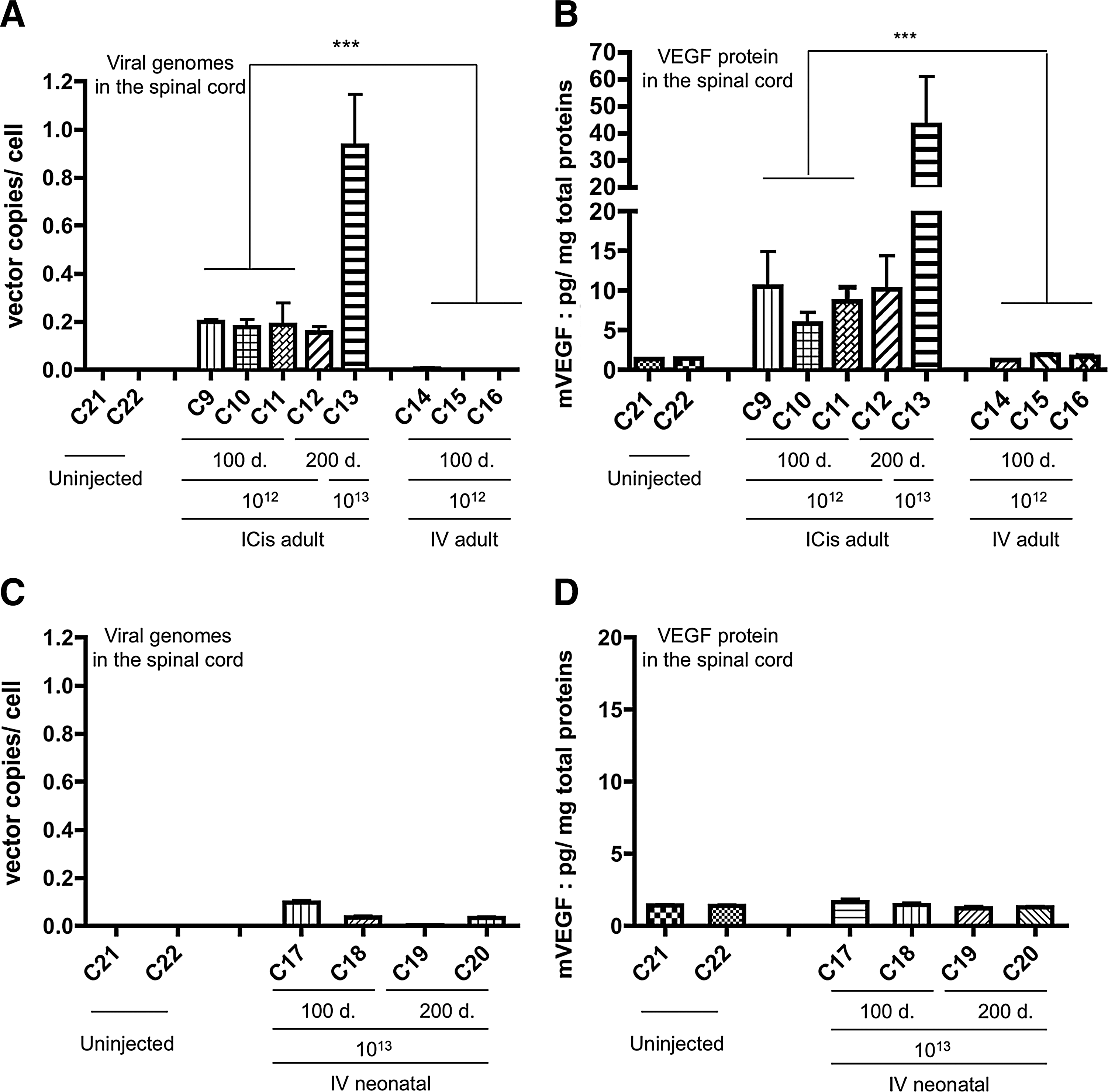
ICis versus IV delivery of scAAV9 results in ∼50 times more vector copies per cell in the spinal cord
Since we previously reported MN transductions after IV administration of 1×1012 vg/kg of scAAV9.gfp in adult cats (Duque et al., 2009), we decided to compare ICis and IV administration of the same dose of scAAV9.vegf. For this, three 7-week-old lix1 +/− cats (C14–C16) received 1×1012 vg/kg of scAAV9.vegf into the jugular vein and results were compared with the ICis delivery group (C9–C11). One hundred days later, the number of vector copies per cell was found ∼50 times higher after ICis delivery with a mean of 0.19±0.03 copies/cell versus 3 10−3±2 10−3 copies/cell after IV delivery of the same vector dose (Fig. 4A). Accordingly, the IV route resulted in a lack of detectable recombinant VEGF protein in the spinal cord (Fig. 4B).
Finally, we looked at whether one log higher dose of scAAV9.vegf vector injected IV at a neonatal permissive age (Foust et al., 2010) could transfer the vegf cDNA in the feline spinal cord. Neonatal IV injections of 1×1013 vg/kg of scAAV9.vegf were performed in two lix1 +/− (C17 and C18) and two lix1 −/− (C19 and C20) 2-day-old kittens (Table 1). We found 0.07±0.03 copies/cell, n=2 (100 days pi), and 0.02±0.004, n=2 (200 days pi), in the spinal cord of heterozygous and homozygous animals, respectively (Fig. 4C). Although more vector genomes per cell were found in the spinal cord of injected newborns compared with the adult group, no recombinant VEGF protein was detected in the spinal cord after neonatal IV injection of 1×1013 vg/kg scAAV9.vegf (Fig. 4D). Additionally, no clinical improvement was observed in the two affected cats (C19 and C20) (Supplementary Fig. S2).
ICis delivery of scAAV9 results in transduction of MNs in the spinal cord as well as neurons in the cerebellum, the cortex, and the thalamus
To confirm that scAAV9 is able to target MNs after ICis delivery in cats and to determine the distribution of the transduced cells in the whole CNS, two adult lix1 +/− cats were injected in the cisterna magna with 1×1012 vg/kg of an scAAV9 vector encoding the GFP. GFP-expressing cells were observed in the cervical and the lumbar portions of spinal cord at 30 days pi (Fig. 5). Double-staining combining GFP detection and an antibody raised against ChAT revealed that MNs were efficiently transduced in the cervical and lumbar segments of the spinal cord (Fig. 5). GFP-positive neurons were also observed in brain structures such as the cerebellum, the thalamus, and the sensory and motor cortex, especially in neurons of the layer 5 (Supplementary Fig. S4). In the cerebellum, Purkinje cells expressed mainly GFP (Supplementary Fig. S4A). In the brainstem, the GFP protein was localized in axonal tracts.
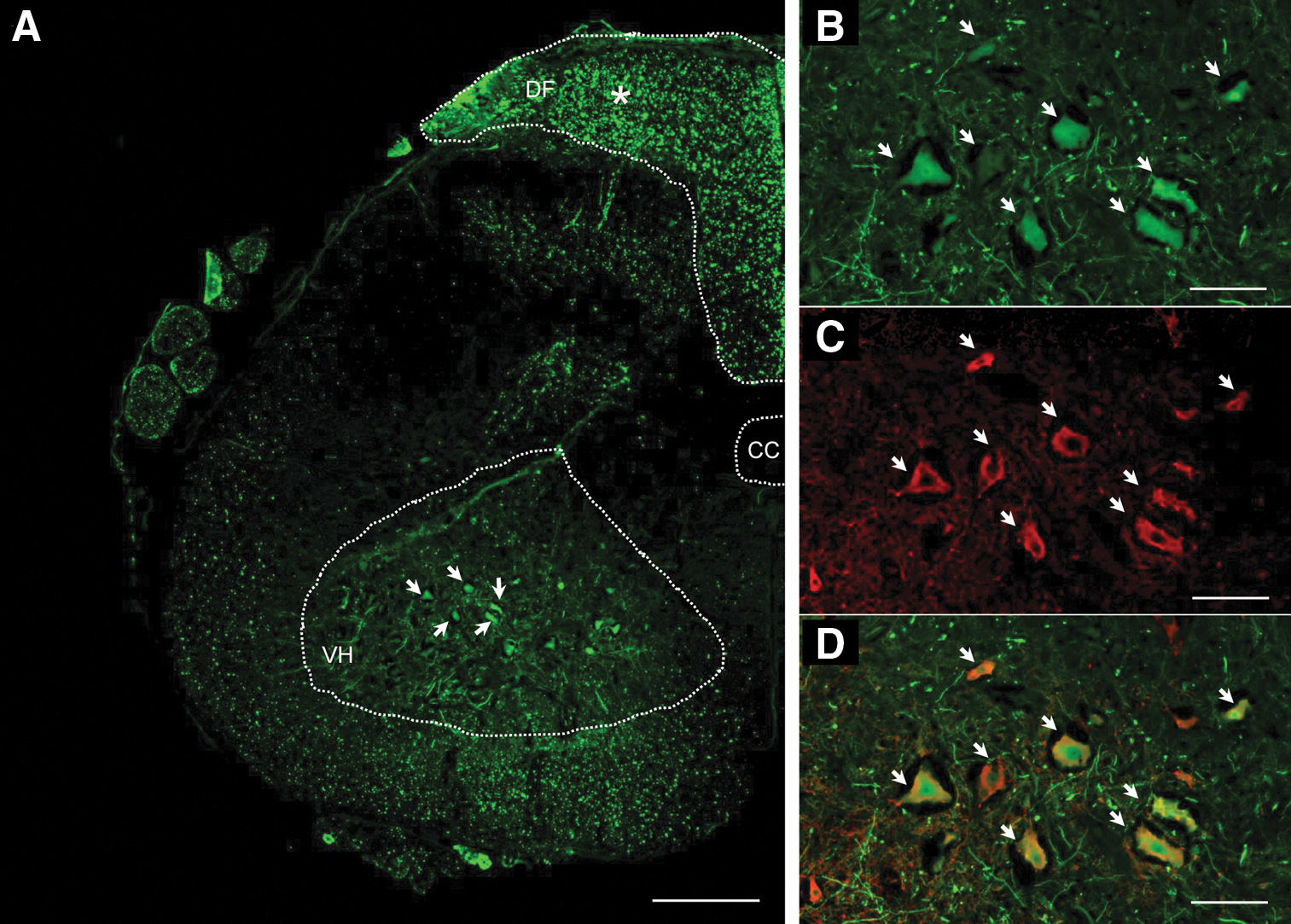
Intracisternal delivery of scAAV9.gfp in 7-week-old cats mediates transgene expression throughout the spinal cord and within MNs.
Off-target VEGF expression after ICis delivery of scAAV9 vector
We finally analyzed if ICis delivery could result in a restricted transduction pattern, that is, limited to the CNS. VEGF concentration was measured in the nerves, the skeletal muscles (triceps brachii and biceps femoris), and the liver of the adult cats intracisternally injected with 1×1012 vg/kg (C9–C12) and 1×1013 vg/kg (C13) of scAAV9.vegf (Fig. 6). The IV injected cats receiving 1×1012 vg/kg of the vector represented the comparative group (C14–C16). The ICis cats exhibited 33.8±9.1 pg (C9–C12) and 75.14±20.54 pg (C13) of murine VEGF per mg of total protein extracts from the peripheral nerves of the forelimb (ulnar and radial nerves) and the hindlimb (sciatic, femoral, and tibial nerves), whereas ∼6 times less murine VEGF expression was detected in the same tissues in the IV group compared with C9–C12 cats (mean of 5±2.5 pg/mg of total protein) (Fig. 6A).
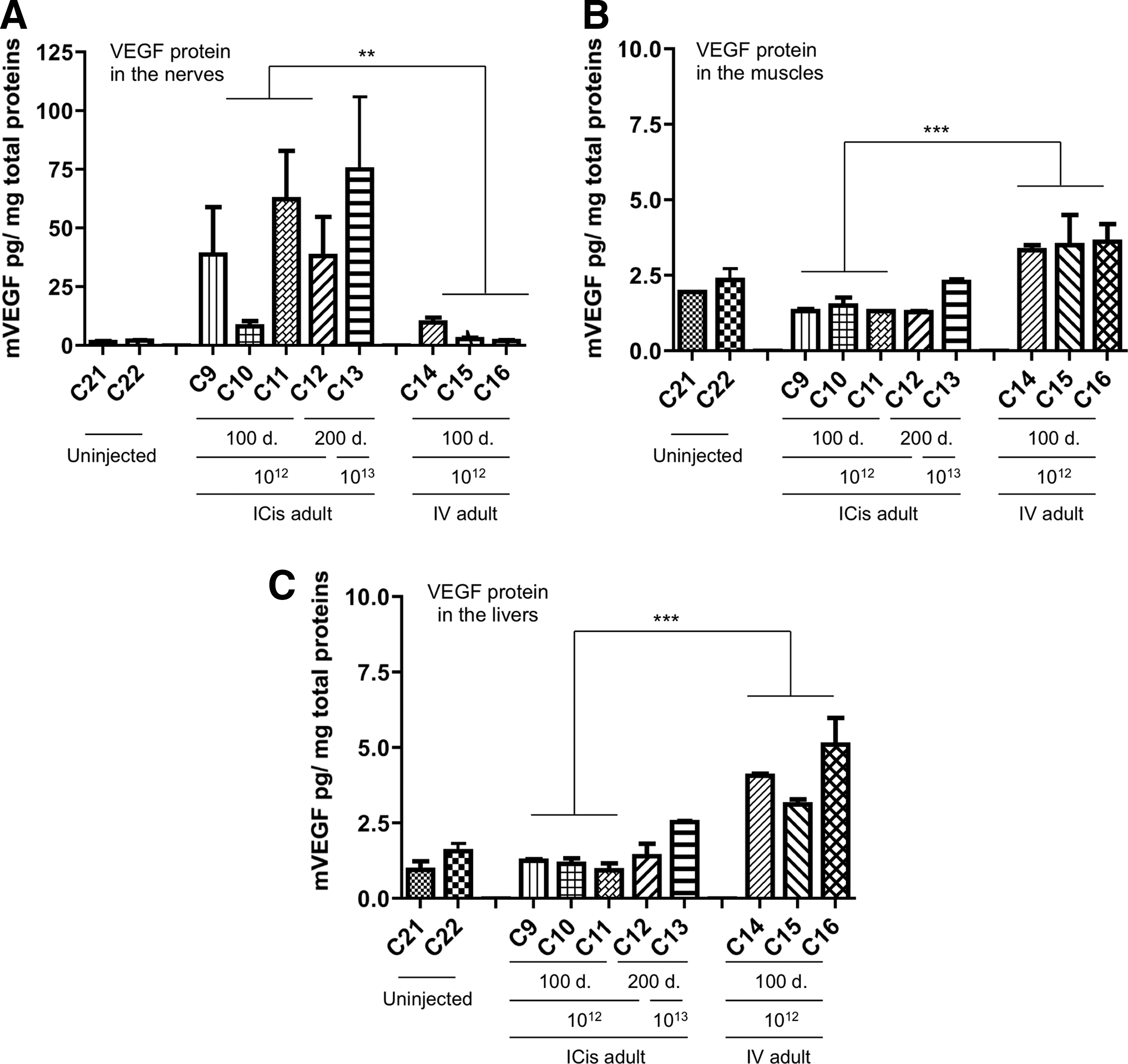
Intracisternal injections of scAAV9.vegf result in a sustained VEGF expression in the nerves but not in the livers or in the muscles.
When we looked for the presence of recombinant VEGF in the triceps brachii, biceps femoris muscles, and the liver, only the C13 cat showed a measurable level of the murine transgene product in the latter (2.55±0.05 pg/mg). Similar levels were found in livers and skeletal muscles from the IV cats group that had received 10 times less vector (4.1±0.6 pg/mg and 3.5±0.08 pg/mg, respectively) (Fig. 6B and C).
Another critical point in evaluating a delivery route is the exposure of the vector to the host immune system since gene therapy efficacy can be limited by immunotoxicity. As shown in Table 2 and despite an immunosuppressive regiment, the ICis route resulted in the detection of AAV9 capsid neutralizing antibodies in the sera as high as after IV administration. Yet, no antibodies against the murine VEGF could be detected by Western blot when incubated with serum from ICis and IV-injected cats (data not shown). Despite the presence of neutralizing antibodies in the serum of the ICis and IV-injected animals, none were detected in the CSF (Table 2).
CSF, cerebrospinal fluid; inj., injection; NA, not available.
Discussion
The main goal of this study was to test the therapeutic efficiency of a neuroprotective factor, the VEGF protein, in an LIX1-deficient cat—a large animal model of lower MN disease—using different modes of AAV delivery into the CNS. We found no clinical benefit in lix1−/− cats after ICra, ICis, or IV injections of AAV-VEGF.
The lack of therapeutic benefit could not likely be attributed to the levels of VEGF expression in the spinal cord since it was found 10–30 times higher in ICis-injected cats than in lix1 −/− uninjected animals at a concentration previously shown to exhibit both neuroprotective and vascular activities in the CNS (Wang et al., 2005). The absence of clinical benefit could also not be related to inflammation-mediated instability of the transgene expression since no signs of inflammation were detected in the spinal cord of the cat lix1 −/− C13 after hematoxylin–eosin–saffron staining and after “ionized calcium-binding adapter molecule 1” immunostaining, a marker upregulated in activated macrophages/microglia (Ito et al., 1998; data not shown). These results suggest that the neuroprotective activity of VEGF has no impact on motor axonal growth defect described in the LIX1 cat (Wakeling et al., 2011). Interestingly, another well-known neuroprotective factor, that is, insulin growth factor 1, was also recently described to be ineffective to treat neuromuscular junction abnormalities and to improve motor function in type III SMA mice (Tsai et al., 2012), suggesting that growth factor overexpression in the spinal cord is not sufficient to treat motor axonal defects and muscle denervation.
On the other hand, our study clearly provides evidences that AAV ICis delivery is an efficient strategy to mediate high transduction of the whole spinal cord including MNs in a large animal model. Although ICra injections of AAV1 resulted in the presence of detectable vector DNA in the spinal cord, recombinant DNA was found predominantly in the cortex, that is, at the injection sites. Our results confirm that AAV1 is efficient in the cat brain (Vite et al., 2003) and could target upper MNs (Hutson et al., 2012), resulting in anterograde transport of AAV particles in axons of the corticospinal tract as previously described (Foust et al., 2008; Ciron et al., 2009; Salegio et al., 2013). However, the efficiency of such strategy is too low to achieve high transduction of the spinal cord probably because a part of the viral genome is trapped in axons without being translated. These results indicate that ICra AAV injection is likely not an effective delivery route to transfer a therapeutic AAV vector to the whole spinal cord.
A promising alternative to target the MNs is the IV administration of AAV vectors, especially with AAV9 serotype (Duque et al., 2009; Foust et al., 2009, 2010; Valori et al., 2010; Bevan et al., 2011; Dominguez et al., 2011; Gray et al., 2011b). If our study confirmed that transduction of the spinal cord occurs in neonatal and adult cats after scAAV9 systemic injection as previously described (Duque et al., 2009), yet, the level of transgene expression remained low in both adult and neonate-injected animals. Indeed, transduction was detected essentially in peripheral tissues, that is, liver. Therefore, if, for instance, severe phenotypes of MN diseases such as type I SMA may require such transduction profile (Vitte et al., 2004; Shababi et al., 2010; Hua et al., 2011), IV delivery may be relevant.
In contrast, the ICis route of injection resulted in strong expression of the transgene in the spinal cord of our cat model, even at the lowest dose of 1012 vg/kg. Interestingly, although AAV1 was not previously shown to be particularly efficient to transduce MNs after intraspinal or intrathecal delivery in rodents (Iwamoto et al., 2009; Snyder et al., 2011), VEGF expression was found in the feline spinal cord after AAV1.vegf ICis delivery in contrast to ICra injections. These results suggested that AAV1 vectors delivered in the CSF can directly transduce neurons of the spinal cord as previously described for the sensory neurons of the DRG with different AAV serotypes (Storek et al., 2008; Iwamoto et al., 2009; Towne et al., 2009; Fu et al., 2010; Vulchanova et al., 2010). Known to have a certain propensity to cross complex biological barriers such as the brain–blood barrier, AAV9 may also have an increase ability to traverse the pia mater and astrocyte layer that separates the spinal cord from the CSF.
Another potential advantage using this local regional delivery route is the limited off-target expression, that is, in the liver and muscles. Lastly, this strategy resulted in high transgene expression in peripheral nerves, suggesting an anterograde transport of the recombinant protein all along the axons, which in turn may have a therapeutic impact on neurodegenerative diseases characterized by defects of axonal growth process.
These results are consistent with two recent studies that compared intravascular and ICis delivery of scAAV9 in mice (Glascock et al., 2012) and in primates (Samaranch et al., 2012, 2013). Both studies indeed showed that delivery of scAAV9 vectors in the CSF mediates higher transgene expression, respectively, in the spinal cord of a murine model of SMA and in the spinal cord, the cortex, and the cerebellum of primates. We found also numerous GFP-positive neurons in the same localization in the brain, that is, the cortex and the cerebellum, as well as in the thalamus, suggesting that this route of delivery could be relevant for the treatment of neurodegenerative disorders affecting simultaneously the spinal cord and the brain.
In conclusion, the present study identified AAV ICis delivery as an efficient method to mediate high transduction of the spinal cord, the brain, and the nerves of a large animal model while reducing peripheral off-target transduction.
Footnotes
Acknowledgments
We would like to thank Lydie Guigand and Mireille Ledevin of the UMR703 Laboratory (Nantes, France) for their assistance with autopsies and congelation of samples. We also thank Oumeya Adjali, who carefully read this article and provided us critical comments, and Johonne Le Duff for the dosage of anti-AAVg antibodies.
This work was supported by the “Association Française contre les Myopathies” and SMA-Europe.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.