
Abstract
Over the past decades, lentiviral vectors have evolved as a benchmark tool for stable gene transfer into cells with a high replicative potential. Their relatively flexible genome and ability to transduce many forms of nondividing cells, combined with the potential for cell-specific pseudotyping, provides a rich resource for numerous applications in experimental platforms and therapeutic settings. Here, we give an overview of important biosafety features of lentiviral vectors, with detailed discussion of (i) the principles of the lentiviral split-genome design used for the construction of packaging cells; (ii) the relevance of modifications introduced into the lentiviral long terminal repeat (deletion of enhancer/promoter sequences and introduction of insulators); (iii) the basic features of mRNA processing, including the Rev/Rev-responsive element (RRE) interaction and the modifications of the 3′ untranslated region of lentiviral vectors with various post-transcriptional regulatory elements affecting transcriptional termination, polyadenylation, and differentiation-specific degradation of mRNA; and (iv) the characteristic integration pattern with the associated risk of transcriptional interference with cellular genes. We conclude with considerations regarding the importance of cell targeting via envelope modifications. Along this course, we address canonical biosafety issues encountered with any type of viral vector: the risks of shedding, mobilization, germline transmission, immunogenicity, and insertional mutagenesis.
Introduction
Split Genome Design to Avoid the Formation of Replication-Competent Species
Initial concepts of retroviral vector design focused on retroviral genera that have a rather simple genome configuration, are quite well understood in their life cycle, and are not known to cause human disease. The murine leukemia viruses (MLV), members of the gammaretroviral family, fulfilled these criteria and thus were used to generate the first generation of retroviral vectors exploited for studies in the context of human gene marking and gene therapy (Williams et al., 1984; Mulligan, 1993; Brenner and Heslop, 2003). Several important principles have emerged from the development of these vectors: (i) the split genome design to avoid the formation of replication-competent species in producer cells (Miller, 1990); (ii) the deletion of enhancer/promoter sequences from the long terminal repeats (LTRs) to generate so-called self-inactivating (SIN) vectors (Yu et al., 1986); and (iii) the decoration of the membrane coat with envelope proteins that modify the host range of the vector particles (pseudotyping) (Emi et al., 1991).
In our current era of clinical trials, insertional mutagenesis is perceived as the major limitation of retrovirus-based (including lentiviral) vectors. It tends to be forgotten that the formation of replication-competent species causes substantially greater safety concerns, because the consequences can be severe (Donahue et al., 1992) and might not be confined to the individual receiving the treatment. With the notable exception of the targeted formation of replication-competent viruses, including MLV, for oncolytic therapy (Tai and Kasahara, 2008), it is a widely accepted standard that intercellular transmission of vector sequences needs to be prevented in somatic gene therapy.
The split-genome design follows a simple principle to avoid the formation of replication-competent species: all sequences that encode retroviral proteins are deleted from the vector, while all sequences that are required for highly efficient packaging into nascent particles are retained. Lentiviruses contain two copies of their full-length, positive-strand RNA genomes, which are specifically selected from a cellular RNA pool and packaged into the lentiviral particles through an interaction of the lentiviral RNA and lentiviral Gag components. Packaging is mediated by interactions between a specific segment of the 5′-untranslated region (UTR) of the lentiviral genome, called the packaging signal ψ, and the nucleocapsid (NC) of the assembling Gag proteins (Berkowitz et al., 1995; Hibbert et al., 2004; Paillart et al., 2004; D'Souza and Summers, 2005). In the case of the human immunodeficiency virus (HIV), this interaction is, at least in part, evolutionarily conserved since HIV-1 can also package HIV-2 RNA (but not vice versa) (Kaye and Lever, 1998). To mediate high-affinity binding to the structural motifs of the lentiviral ψ, NC contains zinc knuckle domains (Muriaux and Darlix, 2010). In comparison to other retroviruses, the lentiviral packaging signal is rather long and requires additional sequences within the trans-activation response element (TAR), U5, the primer binding site (PBS), and the group-specific antigen (Gag) (∼300 bp) for optimal packaging (Fig. 1A) (Harrison et al., 1998). Recent work also shows the necessity of the Rev protein (Brandt et al., 2007; Blissenbach et al., 2010), which enhances the encapsidation of RNA containing the Rev-responsive element (RRE, overlapping with Env coding sequences) by up to 70-fold. The size of the packaging signal (including 300 bp of Gag) and the RRE requirement for efficient packaging explain why the untranslated leader region of currently used lentiviral vectors is rather large (∼1.7 kb; Fig. 1A) (Dull et al., 1998). In an attempt to streamline the necessary components and leader size, minimal lentiviral vectors—which lack parts of Gag, Env, or the RRE (Cui et al., 1999)—have been constructed. However, these have substantially lower vector titers.
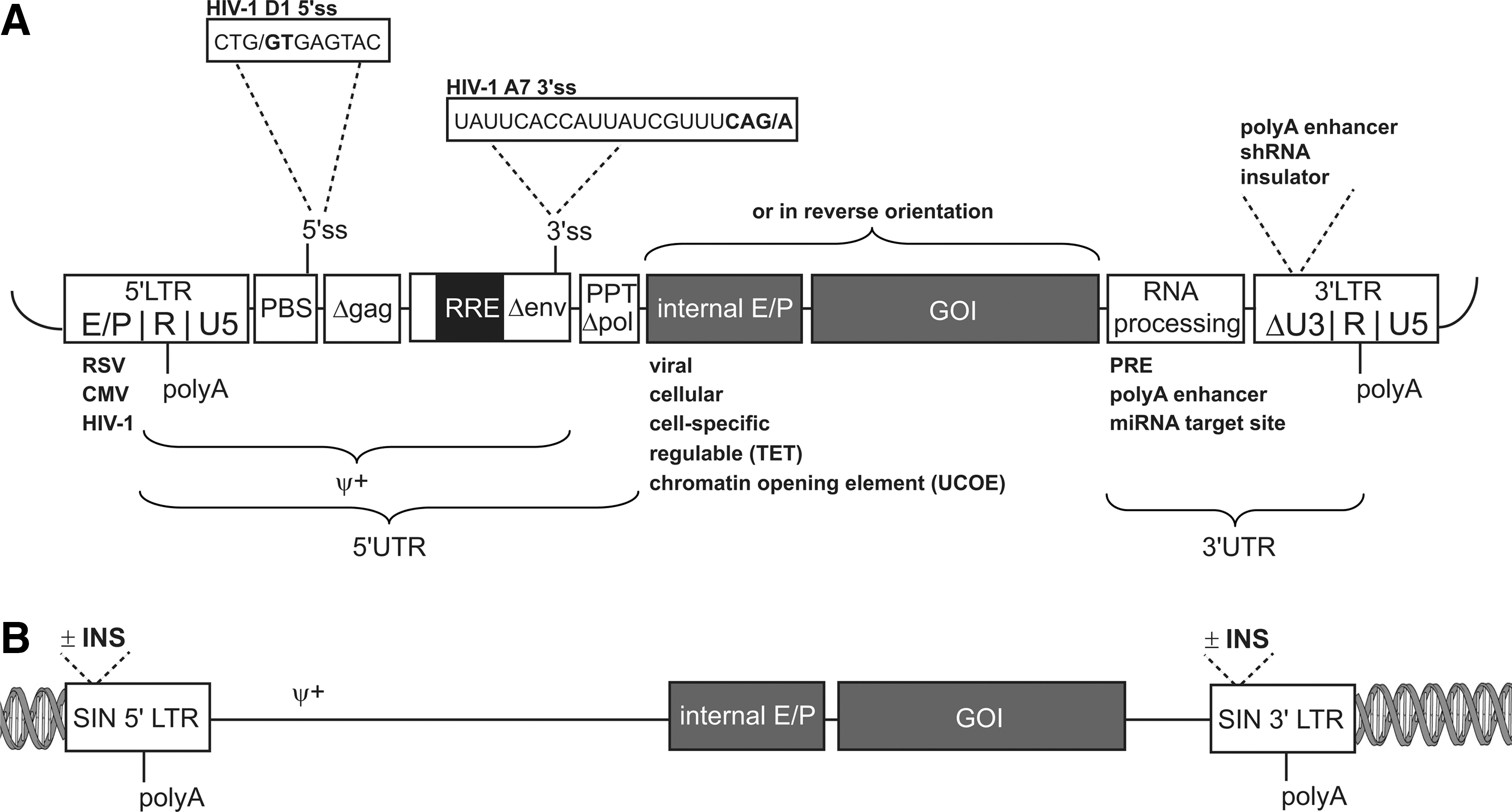
Another sequence that is useful for lentiviral vector design but overlaps with lentiviral coding regions is the central polypurine tract (cPPT, derived from the Pol reading frame). Initially thought to be the mediator of nuclear entry of the PIC (Zennou et al., 2000), a more conservative interpretation of its function is that it increases the efficiency of reverse transcription (RT), which starts within the nascent particles but is typically only completed in the target cells, thus elevating functional vector titers. From the perspective of the sequence fidelity of vector genome, the process of RT is error-prone, resulting in misincorporation in the range of at least 1 in 10,000 processed nucleotides. Furthermore, in a given proportion of transduced cells, the reverse-transcribed sequence may contain larger, sometimes poorly predictable deletions due to cryptic splicing events that occur prior to RNA encapsidation (pre-genome splicing), or because of reverse transcriptase “jumps” between homologous sequence stretches. These errors can be prevented by synthetic sequence design (see below). A more theoretical concern is the chemical editing of the lentiviral RNA sequence by restriction factors such as APOBEC3G (Imahashi et al., 2012), which can be circumvented with the use of appropriate packaging cells and thus does not require co-expression of the lentiviral accessory Vif in the packaging process. Therefore, the lentiviral split-packaging system typically consists of at least four plasmids: one encoding the vector genome, one for Gag-Pol, one for Rev, and one (or two) for Env.
Importantly, and in contrast to earlier generations of MLV vector systems, the potential formation of replication-competent viruses by recombination events between the vector and the packaging constructs has not been observed to date for advanced split-genome lentiviral vectors (Donahue et al., 1992; Chong and Vile, 1996; Leath and Cornetta, 2012). Even though there is considerable overlap of the extended lentiviral packaging signal with lentiviral coding sequences (Δgag, Δenv, Δpol) (Fig. 1A), the artificial alignment of these sequences in the lentiviral leader sequence, the incorporation of additional safety features such as the deletion of translational initiation codons from residual reading frames, plus the lack of larger segments of sequences encoding Env and some of the accessory proteins, have all contributed to an improved design that has a convincing safety record.
SIN Design and RNA Processing
The wildtype lentiviral LTR contains the 5′ attachment signal for the integrase and enhancer/promoter sequences in the U3 region, the polyadenylation motif in the R region, and sequences required for completion of the viral life cycle, including the 3′ attachment signal for the integrase, in U5. The terminal repetition of these sequences, as in all other retroviruses, explains a selection for a rather weak polyadenylation motif and also establishes an enhancer/promoter at the end of the provirus. In the case of MLV and other oncogenic retroviruses, the terminal repetition of the enhancer/promoter plays an important role for the risk of insertional mutagenesis (Uren et al., 2005). In the case of HIV-1, which causes cell death when fully activated for replication, the consequences of insertional mutagenesis induced by the wildtype virus are less well investigated. In any case, full activity of the HIV promoter depends upon the accessory protein Tat, which is expressed in the late phase of the replication cycle. In early vector generations, some of which are still used in preclinical studies, Tat must be cotransfected in the packaging cells to achieve maximal expression of the lentiviral vector genome (Demaison et al., 2002).
The complete deletion of the enhancer/promoter sequences from the LTR was an important goal of lentiviral vector design in order to reduce the risk of mobilization and to avoid interference between the lentiviral enhancer sequences and the transcriptional control elements incorporated into the vector. As mentioned above, this SIN design was originally developed for MLV vectors, but only low titers were achieved in the initial versions and the deletion of the enhancer/promoter sequences was incomplete (Yu et al., 1986). The relevant enhancer/promoter sequences are deleted in most of the currently used lentiviral SIN vectors and these vectors are also equipped with a heterologous 5′ promoter (typically derived from the Rous sarcoma virus [RSV] or the human cytomegalovirus [CMV]) to express high levels of genomic RNA in packaging cells, making the additional expression of the lentiviral Tat protein superfluous (Dull et al., 1998). If expression of the vector genome is toxic for the packaging cells, inducible promoters can also be used in the 5′ U3 position of the lentiviral vector plasmid (Xu et al., 2001).
Studies addressing posttranscriptional RNA processing play an increasingly important role for the biosafety and potency of lentiviral vectors. On the one hand, alternative splicing from the vector cargo into neighboring genes, or vice versa, presents a safety risk potentially leading to truncations and abnormal transcripts. On the other hand, defined interventions in RNA processing offer increased fidelity of the vector genome, improved gene expression features, and overall improved biosafety.
Alternative splicing is a major mechanism through which lentiviruses ensure expression of a variety of viral proteins from their limited genomic space. The full-length HIV-1 transcript contains various splice donors and acceptors leading to a variety of alternative mRNAs, including unspliced (9 kb genomic RNA-encoding Gag/Pol), incompletely spliced (e.g., encoding Env, Vif, Vpr), and fully spliced mRNAs (e.g., for Rev, Tat) (Stoltzfus and Madsen, 2006). The majority of splice signals are removed in SIN lentiviral vectors. However, some splice sites (SS) are retained, especially in the 5′ UTR, including the major splice donor (D1) and a splice acceptor in the RRE/Env fragment (A7) (Fig. 1A). Almarza et al. (2011) reported fusion transcripts between cryptic SS of the lentiviral vector and cellular sequences in keratinocyte clones containing lentiviral vector integration events. Although this occurred in half of the targeted genes, most aberrant transcripts accumulated only at low levels, possibly due to nonsense-mediated decay effects or nuclear retention. In line with this general observation, Montini and colleagues used a whole transcriptome methodology to detect aberrant splicing events induced by lentiviral integrations and demonstrated the presence of chimeric lentiviral-cellular fusion transcripts (Cesana et al., 2012). Noteworthy, the authors reduced residual transcriptional read-through by recoding identified SS, indicating that incorporation of splicing knowledge into vector design may lead to the generation of safer vectors. Underlining the relevance of this goal, changes of mRNA processing following intragenic lentiviral vector integration have been associated with clonal expansion in two cases (Cavazzana-Calvo et al., 2010; Heckl et al., 2012).
Therefore, in addition to the vector backbone, precautions have to be taken to avoid cryptic SS in the vector's transgene expression cassette. “Codon optimization” can be used to remove cryptic splice sites, simultaneously destroying unwanted RNA secondary structures and instability motifs and introducing optimal codons for eukaryotic cells of Homo sapiens. Optimal codon use results in increased mRNA stability, mRNA export, and translation efficiency of the transgene cassette (Moreno-Carranza et al., 2009; Ng et al., 2010; Zhou et al., 2010; Pike-Overzet et al., 2011; Warlich et al., 2011). Although the many point mutations introduced by codon optimization may theoretically cause novel unwanted properties, to the best of our knowledge, no issues of relevance for the biosafety assessment of lentiviral vectors have been reported. Still, we would recommend a careful side-by-side evaluation of wildtype and synthetic sequences with appropriate endpoints of efficiency and toxicity when employing codon-optimization of lentiviral-vector cargo for therapeutic applications.
In contrast to the sequences introduced to improve expression in the target cells of the lentiviral vector, the 5′ UTR sequences of the vector backbone (Fig. 1A) tend to be poorly expressed even if a suitable promoter is present to initiate transcription. Indeed, the wildtype lentiviral Gag-Pol sequences are typically retained in the nucleus unless the transcript is recognized by the lentiviral accessory protein Rev, which binds to its cognate RRE. In wildtype HIV, RRE is located in the Env reading frame and thus in the 3′ UTR of the Gag-Pol message. In the vector, RRE is typically placed upstream of the cPPT (Dull et al., 1998). Thus, unless the vector-transduced cell is infected by HIV, 5′ UTR sequences of lentiviral vectors are not very permissive for expression in the target cells. While the presence of nuclear retention signals in the 5′ UTR might thus contribute to the biosafety of the lentiviral vector, the addition of the RRE has two important functions in the packaging cell. The RRE allows high expression and packaging of the genomic message of the vector in this particular context and suppresses aberrant splice events. The latter function presumably contributes to the observation that lentiviral vectors tend to allow the incorporation of more complex transgene cassettes than other types of retroviral vectors. The most prominent example for this is the successful transduction of complex globin expression cassettes, which contain larger pieces of regulatory DNA and introns (May et al., 2000; Pawliuk et al., 2001; Roselli et al., 2010). However, as the suppression of aberrant splicing by Rev is not complete, cassettes containing introns should be inserted in the inverse orientation (with respect to the orientation of the genomic vector transcript), in a way that does not form new aberrant splice sites.
Modifications of the 3′ UTR of the lentiviral vector message have also made an important contribution to vector design. The Woodchuck hepatitis virus (WHV) posttranscriptional regulatory element (PRE), originally identified in a Hepatitis B–like virus, is a promising tool for improvement of 3′ end processing (Zufferey et al., 1999; Hope, 2002). Introduction of the PRE into the 3′ UTR (Fig. 1A) improves titer and transgene expression in a context-dependent fashion. The extent to which the PRE enhances transgene expression (up to 10-fold) depends on the sequence, the size, and the region of the PRE used and the specific vector and target-cell context (Schambach et al., 2000; Schambach et al., 2006; Higashimoto et al., 2007; Schambach et al., 2007; Zanta-Boussif et al., 2009). Mechanistically, the PRE links the RNA, on which it is located, to CRM-1–dependent nuclear export pathways and also improves mRNA stability and translation efficiency. The PRE partially overlaps with the X protein open reading frame (ORF), which might play a role in WHV-associated hepatocellular carcinoma (Bouchard and Schneider, 2004). Although the role of the PRE in tumorigenesis is only partially substantiated (Bouchard and Schneider, 2004), we and others have designed PRE elements that lack the X protein coding sequence, the X ORF promoter, and potential ATGs for ORFs, while retaining their improved vector titer production and expression (Fig. 2) (Schambach et al., 2006; Schambach et al., 2007; Zanta-Boussif et al., 2009). While one report suggested that the WHV PRE might contribute to the risk of tumor induction after lentiviral vector transduction of hepatocytes (Themis et al., 2005), we found no evidence that it enhances insertional transformation of hematopoietic cells (Modlich et al., 2009).
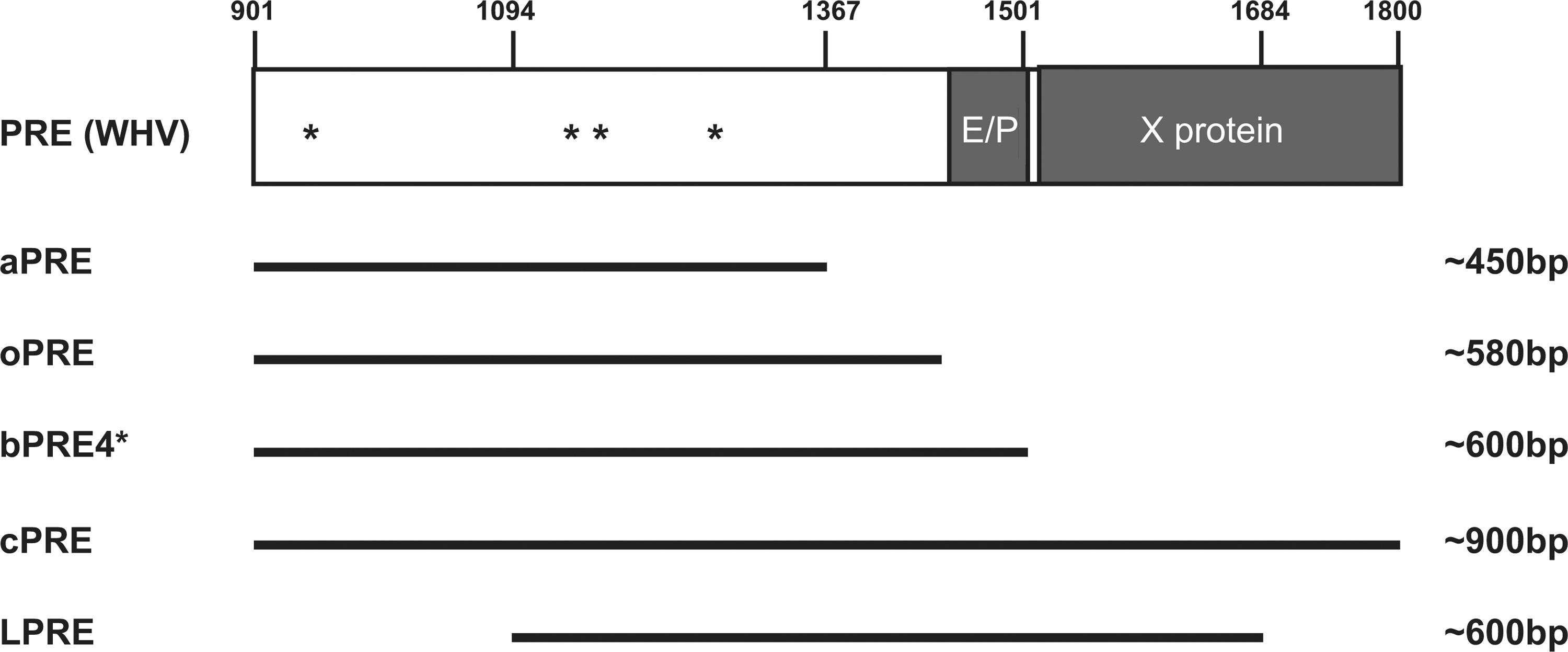
Schematic depiction of the post-transcriptional regulatory element (PRE) from Woodchuck hepatitis virus B (WHV); nucleotide numbering is according to Genbank accession no. J02442. Five different PRE fragments have been designed and tested in different studies. Based on secondary structure predictions, three different PRE fragments were developed: aPRE (450 bp), bPRE (600 bp), and cPRE (900 bp). The PRE fragment by Hope and coworkers is frequently used in lentiviral vectors (nt. 1093–1684) and was termed LPRE for simplicity. The original wPRE harbors enhancer/promoter sequences followed by a partially overlapping open reading frame (ORF) for the WHV X protein. In the bPRE4*, asterisks mark the four mutated ATGs that are followed by an ORF encoding more than 25 amino acids. The oPRE is derived from bPRE4* and is completely devoid of the X-protein promoter.
The PRE reduces, but does not fully prevent, read-through transcription into cellular sequences (Higashimoto et al., 2007; Schambach et al., 2007). Transcriptional read-through is a notorious problem of retroviruses, which—possibly due to their LTRs—have evolved rather weak polyA signals in the R regions (Zaiss et al., 2002). To improve 3′ end processing and transcriptional termination, upstream sequence elements (USE), derived from viral or cellular genes, have been introduced into the 3′ U3 region of lentiviral vectors. In particular, a recombinant direct repeat of the USE derived from simian virus 40 (2xSV USE) effectively blocked transcriptional read-through into adjacent sequences, thereby improving vector efficiency and potentially also biosafety (Schambach et al., 2007).
Another important principle to improve lentiviral-vector performance at the level of RNA processing is the incorporation of target sites for differentiation-specific cellular miRNAs into the 3′ UTR of the therapeutic cassette. This principle has been useful to avoid antigen presentation in dendritic cells, thus promoting tolerance to the transgene product, or to avoid toxic effects of ectopic transgene expression in hematopoietic stem cells (Brown et al., 2006; Brown et al., 2007a; Brown et al., 2007b; Brown and Naldini, 2009; Gentner et al., 2010). Dose-response studies suggest that this approach has little, if any, negative consequences for the regulation of cellular targets of the miRNA recognizing the vector sequence, unless the vector has a very strong enhancer/promoter and is introduced at a high copy number per cell (Gentner et al., 2009).
Integration Pattern, Insertional Mutagenesis, Insulators
The cellular response to lentiviral transduction includes early events that activate innate immunity through various mechanisms (Agudo et al., 2012, and refs. therein), and even global epigenetic changes have been reported (Yamagata et al., 2012), with yet unknown functional consequences. As the efficiency of lentiviral gene transfer can be increased by transiently blocking the innate immune response with immunosuppressants (Agudo et al., 2012), genotoxic lesions remain the most problematic cellular adverse reaction induced by lentiviral gene transfer.
Malignant cell transformation has been observed in animal models and clinical trials after retroviral vector-mediated transfer of sequences, which are not oncogenic by themselves. Insertional mutagenesis—that is, the upregulation or disruption of cellular genes by the retroviral integration event—has been identified as a crucial initiation event in these cases (Li et al., 2002; Hacein-Bey-Abina et al., 2003; Hacein-Bey-Abina et al., 2008; Howe et al., 2008; Stein et al., 2010). A first manifestation of insertional mutagenesis is the outgrowth of distinct clones from a polyclonal population (clonal dominance) (Kustikova et al., 2005; Ott et al., 2006). However, clonal dominance can also develop in the absence of insertional alterations of proto-oncogenes or tumor-suppressor genes, probably due to systemic forces that drive clonal selection on the basis of other events (Schmidt et al., 2003; Cornils et al., 2009). Therefore, mapping a vector integration event in the vicinity of a cellular gene, even if this is involved in growth regulation, is not sufficient to conclude that aberrant cell proliferation was induced by the integration event. Clonal dominance as a premalignant manifestation of insertional mutagenesis can be difficult to detect if the cell clone does not yet contribute to the sample studied in the clinical follow-up. This became evident in the cases of acute lymphoblastic T-cell leukemias (T-ALL) observed in the gene therapy trials for X-linked severe combined immunodeficiency (SCID-X1) (Hacein-Bey-Abina et al., 2003; Hacein-Bey-Abina et al., 2008; Howe et al., 2008). If clonal dominance is associated with the formation of mature, functionally intact cells, it can even contribute to a therapeutic effect, as observed in gene therapy trials for chronic granulomatous disease (CGD) and thalassemia (Ott et al., 2006; Cavazzana-Calvo et al., 2010). Therefore, only malignant cell transformation, which typically requires the acquisition of secondary mutations in addition to the insertional event (Hacein-Bey-Abina et al., 2003, 2008; Howe et al., 2008; Stein et al., 2010), but not clonal dominance per se, represents a serious adverse event.
So far, malignant cell transformation after vector-mediated insertional mutagenesis has only been observed in three clinical entities (SCID-X1, CGD, and Wiskott-Aldrich syndrome [WAS]), all of which occurred in conjunction with the use of first-generation gammaretroviral vectors harboring LTRs with strong enhancer/promoter sequences (Hacein-Bey-Abina et al., 2003, 2008; Howe et al., 2008; Boztug et al., 2010; Stein et al., 2010; Persons and Baum, 2011). Data available to date, from clinical trials conducted with lentiviral vectors, have demonstrated more polyclonal patterns (Cartier et al., 2009; Biffi et al., 2011), with the exception of the first patient reported from a thalassemia trial (Cavazzana-Calvo et al., 2010). In this case, the lentiviral vector was found to disrupt the 3′ regulatory region of the HMGA2 transcript, which encodes a transcription factor involved in aging-dependent stem cell self-renewal (Hammond and Sharpless, 2008). An enhancer effect mediated by regulatory sequences of the therapeutic cassette contained in the lentiviral vector might also have contributed to HMGA2 upregulation (Cavazzana-Calvo et al., 2010). Additional studies conducted in preclinical models underline the concern that intragenic insertion events of lentiviral vectors can have functional consequences (Table 2) (Arumugam et al., 2009a; Bokhoven et al., 2009; Modlich et al., 2009; Montini et al., 2009; Knight et al., 2010; Cesana et al., 2012; Heckl et al., 2012; Knight et al., 2012; Moiani et al., 2012).
However, despite a very high transduction efficiency achieved in recent clinical trials using lentiviral vectors, molecular clonality studies have not indicated any reasons for concern. Several hypotheses have been put forward to explain why relatively stable polyclonal hematopoiesis can occur despite the expected presence of multiple insertional events affecting growth-regulatory genes. First, vectors designed to lack strong enhancer sequences greatly reduce the risk of proto-oncogene activation (Montini et al., 2006; Ramezani et al., 2008; Ryu et al., 2008; Zychlinski et al., 2008; Zhou et al., 2010). Second, the remaining risk of gene truncation events may have a lower power (as compared to proto-oncogene upregulation) for the generation of dominant clones. Third, the presence of multiple competitor clones that do not harbor functionally relevant mutagenic events might suppress insertional mutants (Maetzig et al., 2011). Fourth, some of the current trials using lentiviral vectors might be conducted in clinical settings that have a relatively low risk to select transformation events.
While the latter two hypotheses point to disease-specific and still poorly understood cofactors involved in the selection of malignant cell clones, convincing evidence has been obtained that the vector cargo is the most important variable contributing to the risk of insertional mutagenesis. The principles of SIN vector design have been explained above. Quantitative assays have demonstrated that the risk of insertional upregulation of proto-oncogenes is directly related to the strength of the enhancer sequences contained in the vector (Montini et al., 2006; Ramezani et al., 2008; Ryu et al., 2008; Zychlinski et al., 2008; Zhou et al., 2010). The internal promoter and all other components of the transgene should be designed in a way that renders the inclusion of strong enhancers superfluous. Following the concept that the risk of transformation is higher in stem cells compared to mature cells, enhancers that are only active in differentiating cells may be relatively safe. Improvement of mRNA processing, as discussed above, can thus make an important contribution to the safety of lentiviral gene transfer. Furthermore, the space generated by the U3 deletion in the 3′ LTR can be used to introduce so-called insulator sequences.
Insulators are genetic sequences that establish boundaries between regulatory gene regions. They may increase the autonomy of a given transcription unit by acting as a barrier element that suppresses the spread of heterochromatin and by blocking the interaction between an enhancer and a promoter. The latter function has been connected to the binding of CTCF, an architectural transcription factor that may stabilize epigenetic interactions even when occurring across long genetic distances (Gaszner and Felsenfeld, 2006). Various forms and configurations of insulators, mostly but not exclusively derived from the archetypical HS4 element of the chicken β-globin gene (Gaszner and Felsenfeld 2006), have been tested in lentiviral vectors in an attempt to increase the probability and precision of gene expression from the semi-randomly integrated transgene. Blocking epigenetic cross-talk between the integrated vector and the neighboring chromatin may also reduce the risk of insertional mutagenesis, at least in the cases induced by enhancer-related interactions.
However, the effects caused by the inclusion of insulators into the U3 of lentiviral SIN vectors are quite controversial. While some evidence suggests that insulators decrease the risk of activating proto-oncogenes located proximal to the integration site (Ryu et al., 2008), thus reducing the risk of clonal imbalance in various experimental platforms employing lentiviral or gammaretroviral gene transfer (Evans-Galea et al., 2007; Ramezani et al., 2008; Zychlinski et al., 2008; Arumugam et al., 2009a; Li et al., 2009), the exact mechanism of this “genoprotective” action is still unclear. Genetic instability of large or repeated insulator sequences and interference with post-transcriptional RNA processing or lentiviral reverse transcription complicate the analysis (Arumugam et al., 2009b; Hanawa et al., 2009). This issue is illustrated by the paradoxical observation of a case of clonal dominance in the first successful human application of a lentiviral globin vector. Here, a rearranged insulator element had formed a splice site that contributed to the truncation of the HMGA2 transcript, in association with (yet nonmalignant) clonal expansion (Cavazzana-Calvo et al., 2010). Thus, the optimal setting and the best sequences to employ insulators for the improvement of lentiviral transgene expression and biosafety still remain to be defined.
Next to the genetic cargo of the vector, the second most important risk factor for insertional mutagenesis is the precise pattern of semi-random integration, which is rather unique depending on the nature of the integrase and its viral and cellular cofactors (Schroder et al., 2002; Mitchell et al., 2004; Lewinski and Bushman, 2005). Lentiviral vectors based on HIV-1 or other lentiviruses analyzed to date have the canonical feature of preferential integration within transcription units in gene-dense regions and are associated with the selection of a few preferred loci (“hotspots”), which are largely determined by epigenetic factors (Schroder et al., 2002; Biffi et al., 2011). This pattern appears to be related to at least two mechanisms: the function of the cellular protein LEDGF/p75 as a tethering factor of the integrase, and the “injection” of the PIC through nuclear baskets, thus preferentially targeting genomic regions of active chromatin located next to these sites (Ocwieja et al., 2011; Schaller et al., 2011). Attempts to redirect the lentiviral pattern involve the introduction of LEDGF mutants into the target cells (Gijsbers et al., 2010) and the direct modification of the integrase (Lewinski et al., 2006). Directly swapping the integrase of a lentiviral vector with versions derived from retroviruses that have a more neutral and even less genotoxic integration pattern, such as alpharetroviruses (Mitchell et al., 2004; Hu et al., 2008; Suerth et al., 2012), has not been possible to date without severely reducing infectivity. Attempts made to modify the integration pattern should be studied in models that can detect cell transformation events with high sensitivity. It is also important to maintain the major advantages of lentiviral vectors, such as the high transduction efficiency, including resting cells, achieved with minimal manipulation.
Pseudotypes, Targeting, and Shedding
Multiple principles have been introduced to equip lentiviral vectors with glycoproteins that increase the stability of the infectious particles and/or mediate improved targeting of the cell population of interest. Substantial evidence exists that novel pseudotypes can improve the cell-specific targeting in mixed populations of cultured cells or tissues in vivo and greatly support the ability of lentiviral vectors to transduce various forms of unstimulated hematopoietic cells (monocytes, plasma cells, B cells, T cells, hematopoietic stem cells [HSC]). While reviews of these principles are available elsewhere (Frecha et al., 2008; Buchholz et al., 2009), we would like to focus on a few biosafety considerations. First, it has recently been shown that preformed antibodies that may neutralize lentiviral particles can be circumvented depending on the source of the foreign glycoprotein, as exemplified with measles viruses and related paramyxoviral principles (Enkirch et al., 2012; Kneissl et al., 2012; Levy et al., 2012). Second, while in vivo targeting appears to be in reach, even in rare populations such as primitive hematopoietic cells (Frecha et al., 2012), a substantial amount of the particles can still be found in cells that do not express the receptor mediating cytoplasmic entry (Voelkel et al., 2012). Although unspecific uptake inactivates lentiviral particles in the endolysosomes, it may still contribute to antigenicity, especially in the case of repeated applications. Third, improved cell targeting establishes an important principle by which accidental transduction of germ cells can be avoided. Although this constitutes a rather theoretical risk, every form of a preventive action is certainly welcome (Billings et al., 1999). Fourth, cell-specific targeting, or the use of envelopes that are recognized by preformed antibodies, should increase biosafety in the case of vector shedding. Several investigators have found that lentiviral particles pseudotyped with the glycoprotein of vesicular stomatitis virus (VSVg), by far the most widely used lentiviral pseudotype, can be released from transduced cells for many hours after exposure, thus allowing the transmission of replication-defective yet infectious vectors to secondary target cells. This has been demonstrated after in vivo administration of lentivirally transduced cells (Blomer et al., 2005) and was also recapitulated in cell culture systems (Pan et al., 2007; O'Neill et al., 2010). Washing cells with simple saline solutions, even when adding anti-infectious principles such as heparin, did not fully abrogate transmission to secondary targets (Blomer et al., 2005; Pan et al., 2007; O'Neill et al., 2010). A direct transcytoplasmic journey of macropinocytosed, intact particles may be the underlying principle (O'Neill et al., 2010). We could easily reproduce the transmission of VSVg-pseudotyped lentiviral, gammaretroviral, and alpharetroviral vectors from exposed suspension cells (human K562 and primary CD34+) to co-cultured adherent cells (293T). Only the addition of AZT, an inhibitor of reverse transcription, was able to fully block this transmission, demonstrating that it depends on the shedding of intact retroviral particles (Ehrnstroem, Schambach, and Baum, data not shown). To date, functional consequences other than the transmission of reporter cassettes, or true adverse events caused by such transmission events, have not been described (Scaramuzza et al., 2012). Nevertheless, this issue should continue to be addressed in future studies, if possible, in the context of therapeutic or toxicological models, which examine the kinetics and efficiency of this process in greater detail, as well as its dependence on the type of retroviral particle and envelope.
Summary and Outlook
Current generations of lentiviral vectors have a convincing safety record, offering a number of features that promise both increased potency and safety of therapeutic gene transfer into long-lived and replicating target cells, transduced either ex vivo or in vivo. However, residual imprecision typical for viral gene transfer systems and some undesirable functional consequences of the lentiviral life cycle remain subjects of intense investigation. Currently, the residual genotoxic risk associated with the preferential intragenic integration events represents the most obvious concern. The substantial flexibility of the lentiviral vector genome offers opportunities for redesigning sequences of the backbone as well as the therapeutic cargo to further reduce this risk. When more routine medical use of lentiviral vectors is in sight, or repeated use is indicated in a single patient, issues related to the antigenicity of the particles may receive even greater attention. The most critical bottleneck of the lentiviral technology is still associated with the difficulties to generate large vector stocks in a single production run. It will be interesting to see which modifications will overcome this limitation, and whether these modifications have an impact on the risk of vector mobilization, shedding, the innate and adaptive cellular response, or the genotoxic profile.
Footnotes
Acknowledgments
We thank Michael Morgan for proofreading the manuscript. This work was supported by grants from the German Academic Exchange Service and BMBF (DAAD [0315187, PIDNET], the Deutsche Forschungsgemeinschaft [SFB738, SPP1230], and Cluster of Excellence REBIRTH [EXC 62/1]) and the European Union (Integrated Projects PERSIST and CELL-PID).
Author Disclosure Statement
A.S. and C.B. are inventors of patents on retroviral vector technology. No competing financial interests exist for the other coauthors.