
Abstract
Niemann–Pick type C (NP-C) disease is a neurodegenerative disorder characterized neuropathologically by ballooned neurons distended with lipid storage and widespread neuronal loss. Neural stem cells (NSC) derived from NP-C disease models have decreased ability for self-renewal and neuronal differentiation. Investigation of neurogenesis in the adult brain has suggested that NP-C disease can be overcome, or at least ameliorated, by the generation of new neurons. Bone-marrow-derived mesenchymal stem cells (BM-MSCs) are regarded as potential candidates for use in the treatment of neurodegenerative disorders because of their ability to promote neurogenesis. The underlying mechanisms of BM-MSC-induced promotion of neurogenesis, however, have not been resolved. The aim of the present study was to examine the mechanism of neurogenesis by BM-MSCs in NP-C disease. Coculture of embryonic NSCs from NP-C mice that exhibit impaired ability for self-renewal and decreased rates of neuronal differentiation with BM-MSCs resulted in an enhanced capacity for self-renewal and an increased ability for differentiation into neurons or oligodendrocytes. In addition, results of in vivo studies have demonstrated that transplantation of intracerebral BM-MSCs resulted in stimulated proliferation and neuronal differentiation of NSCs within the subventricular zone. Of particular interest, enhanced proliferation and neuronal differentiation of endogenous NP-C mouse NSCs showed an association with elevated release of the chemokine (C-C motif) ligand 2 (CCL2) from BM-MSCs. These effects suggest that soluble CCL2 derived from BM-MSCs can modulate endogenous NP-C NSCs, resulting in their improved proliferation and neuronal differentiation in mice.
Introduction
Occurrence of neurogenesis in the subventricular zone (SVZ) and the subgranular zone of the mammalian brain throughout adult life, as well as its modulation under various physiologic and pathologic conditions, including Alzheimer's disease, Huntington's disease, and Parkinson's disease, have been well documented (Lie et al., 2004; Abrous et al., 2005; Sailor et al., 2006). Occurrence of injury-induced neurogenesis has been reported in both neurogenic and non-neurogenic regions (cerebral cortex, striatum, and hippocampal CA1) of the adult brain (Dayer et al., 2003; Magnus et al., 2007). However, injury-induced neurogenesis by endogenous neural stem cells (NSCs) cannot fully compensate for the neural loss in patients with central nervous system disorders. Yang et al. (2006) reported significantly decreased ability for self-renewal and differentiation of NSCs in NSCs derived from NP-C mouse models (Npc1 −/− mice). This finding suggests that impaired ability for self-renewal and differentiation of NSCs could be a pathological marker in neuronal degeneration of NP-C disease. These findings indicate that neurodegenerative diseases such as NP-C disease are suitable for stimulation of diseased NSC and that stimulation and regulation of neurogenesis using therapeutic interventions may be useful in treatment of an NP-C diseased brain.
Findings from our previous studies have demonstrated the contribution of intracerebellar transplantation of bone-marrow-derived mesenchymal stem cells (BM-MSCs) toward improving neurological function and survival of Purkinje cells in the brain of NP-C mice (Bae et al., 2005b, 2007; Lee et al., 2010). This therapeutic function of MSC has, in part, been explained by their production of diffusible trophic factors (Zhang et al., 2004; Zhang et al., 2005; Ren et al., 2007; Block et al., 2009). Deng et al. (2006) previously demonstrated the elicitation of neurogenesis and promotion of functional recovery by transplantation of MSCs in rhesus monkeys. The goal of this study was to investigate the neurogenic potential of BM-MSCs and soluble factors released from transplanted BM-MSCs for promotion of neurogenesis of NP-C disease in a mouse model as a paradigm of future cell therapy applications.
This study demonstrated that treatment with BM-MSCs resulted in enhanced capacity for self-renewal, proliferation, and neuronal differentiation of Npc1 mutant NSCs. These effects of BM-MSCs are recapitulated in a mouse model of NP-C disease. Transplantation of BM-MSCs into the SVZ of NP-C mice resulted in stimulated proliferation and differentiation of endogenous NSCs that survived as more mature neural cells. In addition, our observations suggest a role for chemokine (C-C motif) ligand 2 (CCL2) as a key paracrine factor for these neurogenic effects of BM-MSCs in the brain of NP-C mice.
Materials and Methods
Animals
A colony of BALB/c Npc1 nih mice has been maintained for this study by brother–sister mating of heterozygous animals. Polymerase chain reaction (PCR) was performed for determining the genotype of each mouse (Loftus et al., 1997). All procedures were performed in accordance with an animal protocol approved by the Kyungpook National University Institutional Animal Care and Use Committee. Animals were housed in a temperature-controlled room on a 12-hr-light/12-hr-dark circadian cycle.
Isolation and culture of BM-MSCs
Single-cell suspensions were obtained, using a 40-μm cell strainer (BD Biosciences), from bone marrow that was harvested from tibias and femurs dissected from 4- to 6-week-old mice. According to what we learned from a previous study (Bae et al., 2007), approximately 1×106 cells were plated in 25-cm2 flasks containing MesenCult MSC Basal Medium and Mesenchymal Stem Cell Stimulatory Supplements (Stem Cell Technologies) with antibiotics. Cell cultures were grown for 1 week, and the plastic-adherent population (BM-MSC) was used for subsequent experiments.
Knockdown of CCL2 using small interference RNA
Small interference RNA (siRNA) oligonucleotides for CCL2 and scrambled sequence siRNA (siCONTROL), which served as control, were obtained from Dharmacon. BM-MSCs were seeded into T-75 flasks at 1.5×106 cells per flask. On the following day, transfection was performed using Lipofectamine 2000 reagent (Invitrogen). Two days post-transfection, real-time reverse transcriptase–PCR was performed for evaluation of efficiency of knockdown by siRNA.
Isolation and culture of NSCs
Using a previously described method (Chen et al., 2007) with some minor modifications, dissociated cortical and hippocampal neurosphere (NS) cultures were prepared from individual embryonic (E) day 15.5 fetuses, either homozygous NP-C mice or control littermates. The cortex and hippocampi were removed and kept in ice-cold Ca2+/Mg2+-free Hank's balance salt solution. Each cortex and hippocampus was then mechanically dissociated in Dulbecco's modified Eagle's medium (DMEM)/F12 supplemented with 1% N2 supplement (Gibco), epidermal growth factor (20 ng/ml; Peprotech) and basic fibroblastgrowth factor (20 ng/ml; Peprotech). The single-cell suspensions were filtered through a 40 μm cell strainer and seeded in individual wells of a six-well tissue culture plates (BD Biosciences). NSCs proliferated in suspension and formed aggregates referred to as NSs. Every 2 days, half of the medium in each well was replaced with a fresh culture medium.
Coculture of BM-MSCs and NSCs
For the indirect three-dimensional coculture experiments, Millicell Hanging Cell Culture Inserts (Millipore) with a pore size of 1.0 μm were placed on top of previously plated NSCs. BM-MSCs were seeded onto the insert at a density of 3×104 cells per insert. In this system, there was no direct contact between NSCs and BM-MSCs. In addition, we cocultured NIH-3T3 cells with NSCs. To prevent proliferation, we first treated NIH-3T3 cells with mitomycin C (10 μg/ml) for 3 hr. After washing twice in phosphate buffered saline (PBS), mitomycin-C-treated cells were trypsinized and re-suspended in DMEM (Gibco) containing 10% fetal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin and seeded onto the insert at a density of 3×104 cells per insert. During the 7-day-long incubation period, half of the medium was replaced with a fresh medium after 72 hr.
NS formation assay
To examine the effects of BM-MSCs on the formation of NSs, NSs were mechanically dissociated, and the resulting viable individual cells were counted. These cells (1×104 cells per well in uncoated 24-well plates) were also used for assessment of its ability for self-renewal. NS cultures from passage number 3 were used throughout this study. After 7 days of incubation with BM-MSCs, newly formed NSs were counted in each well, using an Olympus IX71 microscope. A minimum cutoff size of 50 μm in diameter was used in defining an NS.
NSC proliferation assays
The proliferative activity of NSCs was evaluated by 5-bromo-2-deoxyuridine (BrdU) immunocytochemistry. The single-cell suspensions from NSs were seeded on glass coverslips coated with poly-L-ornithine (Sigma) at a density of 1×104 cells/cm2. After 7 days of incubation with BM-MSCs, cells were labeled with a 10 μM of BrdU (Sigma) and incubated for an additional 12 hr. After the labeling medium was removed, cells were fixed with phosphate-buffered 4% (w/v) paraformaldehyde (Sigma) for 20 min at room temperature (RT). To denature nuclear DNA into a single strand, cells were incubated in 2 N HCl for 1 hr, and 0.15 M sodium borate for 15 min. Cells were washed by PBS and processed for immunofluorescence analysis of BrdU with DAPI nuclear counterstain.
NSC differentiation assays
For NSC differentiation assays, single-cell suspensions cultured in vitro for 4 days were plated on glass coverslips coated with poly-L-ornithine at a density of 1×104 cells/cm2, followed by incubation in Neurobasal medium (Gibco) supplemented with 100 U/ml penicillin/streptomycin, 2 mM L-glutamine, 10 μg/ml of heparin, 2% B-27 supplement, and 3% FBS. Seven days in vitro after plating, differentiated cultures were processed for immunofluorescence staining.
Immunocytochemistry
Cells were fixed with PBS (0.1 M) containing 4% paraformaldehyde at RT for 15 min and permeabilized with 0.1% Triton X-100 in PBS for 5 min. Cells were preincubated for 10 min with 3% normal goat serum and 2% bovine serum albumin (BSA) in PBS containing 0.4% Triton X-100 to block background immunostaining. For BrdU immunofluorescence staining, cultures were incubated overnight with anti-BrdU (rat, diluted 1:250; Abcam) antibody. For NSC differentiation assays, differentiated cultures were incubated overnight with β-tubulin (mouse, diluted 1:400; Chemicon) as a marker for neurons, GFAP (rabbit, diluted 1:1000; Dako) as a marker for astrocytes, and O4 (mouse, diluted 1:200; Stem cell Technologies) as a marker for oligodendrocyte progenitor cells. For visualization, the primary antibody was developed by incubating with Alexa Fluor 488– or 594–conjugated secondary antibodies for 1 hr at RT against a corresponding species. The cells were analyzed with a laser scanning confocal microscope equipped with Fluoview SV1000 imaging software (Olympus FV1000), or with an Olympus BX51 microscope.
Transplantation of BM-MSCs into NP-C mouse brains
NP-C mice, approximately 4 weeks of age, were anesthetized with a combination of 100 mg/kg ketamine and 10 mg/kg xylazine, and an injection was administered using a stereotaxic injection apparatus (Stoelting Co.), as previously described (Bae et al., 2007). A glass capillary (1.2 mm×0.6 mm) was used for transplantation of BM-MSCs into the cerebrum (te Vruchte et al., 2004). About 3 μl of BM-MSC suspension in PBS (∼1×106 cells) was infused at a rate of 0.3 μl/min at +0.74 anterior/posterior,±1.5 medial/lateral, −2.5 dorsal/ventral relative to bregma into the left striatum. The same volume of PBS was injected at the same rate into the right hemisphere. After transplantation, the scalp was closed by suture and the animals were allowed to recover from the anesthesia before being returned to their cages. Age-matched normal littermates in the NP-C colony were used as controls. For in vivo tracking of transplanted cells, we labeled BM-MSCs with nanoparticle (0.1 mg/ml, NFP-STEM Silanol TMSR-RITC 50; Biterials).
BrdU injection
Mice received intraperitoneal injection with BrdU dissolved in 0.9% NaCl/0.007 M NaOH solution at 12 hr intervals for 7 days (50 mg/kg).
Tissue preparation
Animals were transcardially perfused with 4% paraformaldehyde in PBS at 7 and 30 days after BM-MSC transplantation. After perfusion, brains were removed and postfixed overnight at 4°C, and incubated in 30% sucrose at 4°C until equilibrated. Sequential 30-μm-thick transversal sections were taken on a cryostat (CM3050S; Leica) and stored at −20°C.
Neurogenesis detection
Before performance of BrdU histochemistry, cerebral sections were incubated in 2 N HCl for 1 hr at 37°C, followed by incubation in 0.1 M borate buffer for 10 min. They were then incubated overnight at 4°C in a mixture of anti-BrdU (rat, diluted 1:100; Abcam) and mouse anti-NeuN (mouse, diluted 1:200; Chemicon) antibodies and a mixture of mouse anti-BrdU (mouse, diluted 1:50; BD Biosciences) and rabbit anti-Doublecortin (anti-DCX) (rabbit, diluted 1:1000; Abcam) antibodies; all were visualized using Alexa anti-rat 488, Alexa anti-mouse 488 and 594, and Alexa anti-rabbit 594 as secondary antibodies. Stereological analysis was used for estimation of the number of BrdU+ cells in the striatum and SVZ.
Stereology
Cell proliferation and neuronal differentiation were quantified using stereologic counting of BrdU, DCX, and NeuN+ cells in the SVZ. Systematic random sampling was obtained using every fifth in a series of 30-μm-thick coronal sections through the striatal SVZ. The equidistant sections (five per brain) were used for stereologic counting to assess cell proliferation and neuronal differentiation in the SVZ. All counts were performed through a 60× objective lens on an Olympus BX51 microscope equipped with a DP70 digital camera, and the Visiomorph software (VisioMorph).
Accelerating Rota-Rod analysis
The Rota-Rod test assesses animal's balance and coordination by measuring the amount of time the animal is able to remain on a longitudinally rotating rod. The Rota-Rod apparatus (accelerating model 47600; Ugo Basile) has a 3-cm-diameter rod, which is suitably machined to provide grip. In this apparatus, a motor sets the rotor in motion via the gear belt at a selected speed. When the mouse falls off its cylinder section, the plate below trips and the corresponding counter is disconnected, thereby recording the animal's endurance time in seconds. The machine was set to an initial speed of 4 rpm, and the acceleration was increased by 32 rpm every 25–30 sec. BM-MSCs or CCL2-knockdown BM-MSC-transplanted NP-C mice were analyzed along with sham-transplanted NP-C and normal control mice. Scores were registered every 2 days beginning at 72 hr after transplantation, and three independent tests were performed at each measurement. Uniform conditions were carefully maintained for each test, and there was a rest time of 1 hr between trials. Each test was limited to 300 sec.
Antibody-based mouse cytokine arrays
RayBio Mouse custom Cytokine Antibody Arrays (RayBiotech) were employed for assay of cell culture supernatant fluid from coculture experiments. For cytokine assay, we followed the manufacturer's directions resulting in the following process: Membranes were incubated with 2 ml of a 1× blocking buffer at RT for 30 min to block membranes. About 1 ml of the conditioned medium (CM) from samples with or without cocultured BM-MSCs was then added, followed by incubation overnight at 4°C. After decanting the samples, all membranes were washed with a 1× wash buffer I and II at RT with shaking. One milliliter of 1:250 diluted biotin-conjugated antibodies was prepared and incubated overnight at 4°C, and the washing steps were repeated. Two milliliters of 1:1000 diluted horse-radish-peroxidase-conjugated streptavidin was added, and membranes were incubated for 1 hr at RT, followed by additional washing. Detection buffers C and D (Pierce Biotechnology) were used for visualization of spots. Membranes were wrapped in plastic wrap and exposed to radiographic film (Kodak X-Omat; Kodak) for 20 min, and the signal was detected using a film developer. Each film was scanned into an image processor, and densitometric measurements were performed using an imaging densitometer (Bio-Rad), followed by quantification using Bio-Rad analysis software. Density data were exported into a spreadsheet (Excel; Microsoft Corporation), and background intensity was subtracted before analysis. Densitometry and statistical analysis were performed on the immunoblots normalizing to the positive and negative controls on the array.
Quantitative real-time PCR
The RNeasy Lipid Tissue Mini kit or RNeasy Plus Mini Kit (Qiagen, Korea, Ltd) was used for extraction of RNA from brain homogenates (SVZ of 1 month post-transplantation) and cell lysates, and a Nanodrop ND-1000 spectrophotometer was used for determining concentration. About 5 μg of each RNA was converted to cDNA using the sprint RT complete-oligo (dT) (Clontech), and cDNA was quantified by the QuantiTect SYBR Green PCR Kit (Qiagen, Korea, Ltd). For investigation of each transcript, a mixture of the following reaction components was prepared according to the indicated end-concentration: forward primer (10 pM), reverse primer (10 pM), and QuantiTect SYBR Green PCR master mix. About 15 μl of master mix was added to a 0.1 ml tube, and a 5 μl solution containing 100 ng reverse-transcribed total RNA was added as a template for PCR. The tubes were placed into the Corbett research RG-6000 real-time PCR machine. The following primers were used: CCL2 (forward 5′- ATGCAGTTAATGCCCCACTC-3′, reverse 5′- TTCCTTATTGGGGTCAGCAC-3′), interleukin-4 (IL-4; forward 5′-ATCCATTTGCATGATGCTCT-3′, reverse 5′-GAGCTGCAGAGACTCTTTCG-3′).
ELISA measurement of tissue CCL2 concentrations
CCL2/JE/MCP-1 ELISA assays were performed according to the manufacturer's protocol using Quantikine ELISA Kits (R&D Systems) and approved CCL2 standards. The SVZ from one hemisphere was homogenized in RIPA buffer (Cell Signaling) with 1 mM PMSF. Homogenates were mixed at RT for 3–4 hr. After mixing, homogenates were diluted in PBS containing 5% BSA, 0.03% Tween 20, and protease inhibitor cocktail (Calbiochem). Each CCL2 standard and experimental sample was run in duplicate and the results were averaged.
Western blot analysis
The SVZ from one hemisphere was homogenized in RIPA buffer with 1 mM PMSF at 7 days after BM-MSC transplantation. Total proteins (80 μg) were electrophoresed through 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis and blotted. Membranes were incubated with anti-CCL2 (goat, diluted 1:700; R&D Systems) and anti-β-actin (goat, diluted 1:700; Santa Cruz), and visualized by enhanced chemiluminescence (ECL; Amersham). We carried out densitometric quantification using ImageJ software (U.S. National Institutes of Health).
In vitro CCL2 neutralization
To neutralize CCL2 in vitro, goat anti-mouse CCL2-neutralizing antibodies (R&D Systems; 2.5 μg/ml) were added concomitantly during the 7-day BM-MSC coculture. Control goat IgG (Santa Cruz; 2.5 μg/ml) antibodies served as isotype control for the in vitro assays.
Statistical analysis
The Student's t-test was performed to compare the two groups, whereas Tukey's HSD test and Repeated Measures Analysis of Variance test were used for multigroup comparisons using the SAS statistical package (release 9.1; SAS Institute Inc.). Differences were accepted to be statistically significant at p<0.05.
Results
BM-MSCs enhance self-renewal and proliferation of NP-C NSCs
To investigate the neurogenic potential of BM-MSCs, we used self-renewal and proliferation assay. A summary of the protocol is shown in Fig. 1A. The results showed that the number of cortical NSs in NP-C cultures was significantly decreased, compared with wild-type (WT) cells (Fig. 1B and C). In addition, indirect coculture of NP-C cortical NSCs with BM-MSCs resulted in increased formation of NSs (Fig. 1B and C, p<0.001). BM-MSCs also induced an increase in NS formation in WT cultures. To validate whether the improved NS formation is a BM-MSC-specific, cortical NSCs were indirectly cocultured with NIH-3T3. NS formation in cortical NSCs cocultured with NIH 3T3 cells was similar to that of NSs without coculture in both NP-C cells and WT cells (Fig. 1B and C). These results suggest a BM-MSC-specific effect on NSC self-renewal. Similar results were obtained when hippocampal-derived NSC cultures were indirectly cocultured with BM-MSCs (Supplementary Fig. S1; Supplementary Data are available online at
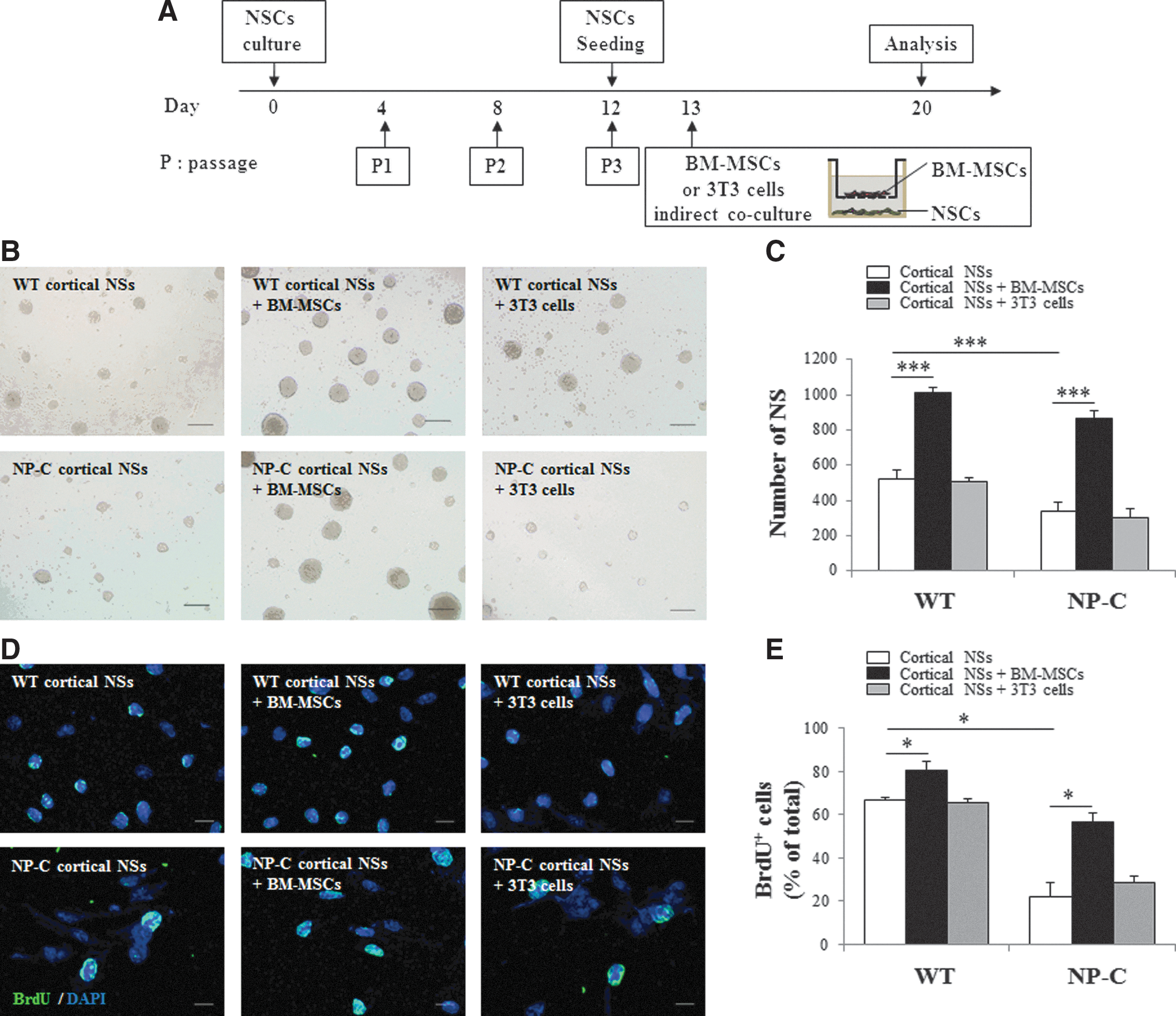
Indirect coculture with BM-MSCs promotes self-renewal and proliferation of NP-C mouse-derived cortical NSCs.
BM-MSCs affect differentiation of NSC into specific cell types
In addition to the ability for self-renewal, the second hallmark of stemness in NS cultures is multilineage differentiation (Reynolds and Weiss, 1996). NSs were dissociated and cocultured with BM-MSCs in differentiation media. After 7 days, immunocytochemistry was performed for analysis of cells in order to investigate the effect of BM-MSCs on capacity for neural multilineage differentiation in NP-C NSC cultures. Expression of markers for the three neural lineages was detected in NSC-derived cells, single-cultured or cocultured with BM-MSCs: β-tubulin for neurons (Fig. 2A and B), O4 for oligodendrocytes (Fig. 2C and D), and GFAP for astrocytes (Fig. 2E and F). Compared with WT cells, we observed diminished differentiation of NP-C cortical NSCs into neurons (Fig. 2A and B, p<0.05). However, indirect coculture of NP-C cortical NSCs with BM-MSCs resulted in significantly enhanced efficiency of neuronal differentiation (Fig. 2A and B, p<0.05). In addition, compared with WT cortical NSCs, we observed a reduced rate of differentiation of O4-expressing cells in NP-C cortical NSCs (Fig. 2C and D). BM-MSCs affected a significant increase in the number of differentiating oligodendrocytes in NP-C cortical NSC cultures (Fig. 2C and D, p<0.05). Compared with WT cells, however, no significant changes in the numbers of GFAP+ cell types were observed in differentiated NP-C cortical NSCs (Fig. 2E and F). Re-evaluation of these results was performed in hippocampal-derived NSCs cultures (Supplementary Fig. S2). Overall, the results indicate that BM-MSCs selectively promote neurogenesis and oligogenesis by NSCs in vitro.

Indirect coculture with BM-MSCs results in enhanced differentiation of NP-C cortical NSC into specific cell types. Differentiation of NSCs from E15.5 cortex was induced for 7 days with or without coculture with BM-MSCs using the indirect three-dimensional system. Seven days after coculture, cells were stained with a specific cell type marker. Representative fluorescence images of β-tubulin/DAPI
BM-MSCs promote SVZ neurogenesis and decrease atrophy of the striatal volume in NP-C mice
Munoz et al. (2005) showed that grafting MSCs into the hippocampus of healthy mice resulted in increased proliferation of endogenous neural cells. This study demonstrated that soluble factors from BM-MSCs promote proliferation and multipotency of NP-C NSCs. We transplanted BM-MSCs labeled with nanoparticles (Fig. 3A) in order to determine whether BM-MSCs can enhance proliferation of endogenous cells in an NP-C mouse model. The treatment protocol is described in Fig. 3A. For assessment of the number of proliferating cells in the SVZ and striatum, injection of mice with BrdU was performed every 12 hr for the first 7 days post-implantation and all actively dividing cells were labeled during the first 7 days (Fig. 3A). Results of immunostaining indicated an increase in the number of BrdU+ cells in the BM-MSC-transplanted NP-C hemisphere within 7 days post-implantation, compared with the PBS-injected, contralateral hemisphere (Fig. 3B and C, p<0.01). In addition, an increase in the number of BrdU+ cells was also observed in the BM-MSC-treated WT hemisphere, compared with the untreated hemisphere (Fig. 3B and C, p<0.05). To confirm differentiation of BrdU+ cells, sections were colabeled with markers of differentiation. DCX is a marker of early neural differentiation (Brown et al., 2003). We quantified the new neuron by counting DCX and BrdU-double-positive cells at 1 week after BM-MSC transplantation (Fig. 3D and Supplementary Fig. S3A). At 4 weeks after transplantation, using unbiased stereological methods, we also performed double-immunostaining using an anti-BrdU antibody and an anti-NeuN antibody in order to estimate the number of proliferating cells that differentiated into mature neurons (Fig. 3F and Supplementary Fig. S3B). When BM-MSCs were grafted in 4-week-old NP-C mice, the number of new neurons (BrdU+ DCX+ cells) showed an increase in the SVZ of NP-C mice (Fig. 3D, p<0.01). In addition, many BrdU+ cells colabeled with NeuN were observed in the transplanted hemisphere, compared with the contralateral hemisphere in NP-C (p<0.001) and WT (p<0.01) mice (Fig. 3F). To examine possible neuron-specific effects of treatment, we analyzed the percentage of DCX+ cells/BrdU+ cells and NeuN+ cells/BrdU+ cells. Almost BrdU+ cells were neuron-specific in all groups (Fig. 3E and G).
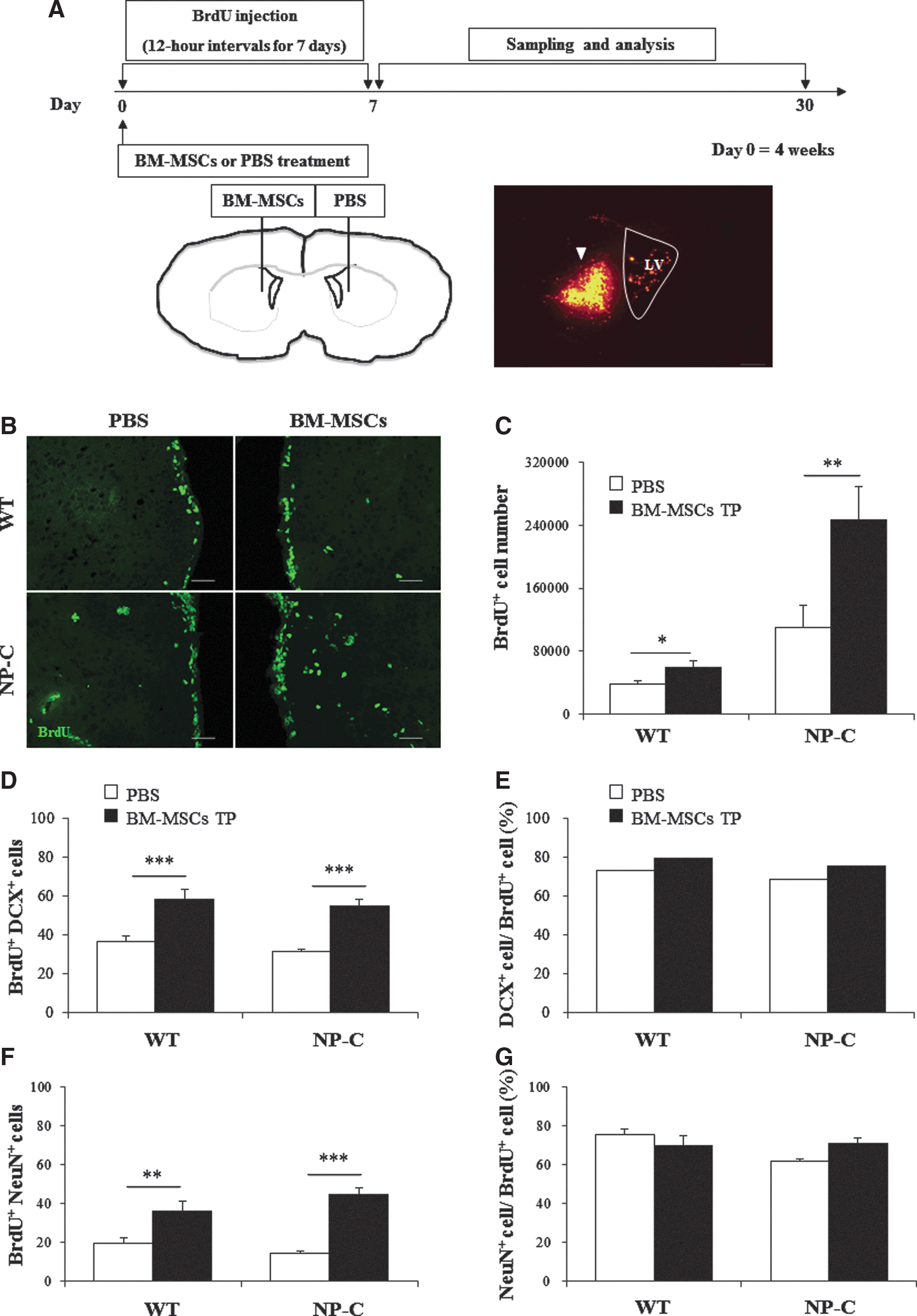
Grafting BM-MSCs increase proliferation and neural differentiation of endogenous NSCs in a mouse model of NP-C. Transplantation of BM-MSCs was performed in NP-C mice at 4 weeks of age.
To assess the effects of BM-MSCs on NP-C pathophysiology, BM-MSCs were transplanted into one hemisphere of the striatum and PBS was injected into the contralateral striatum in 4-week-old NP-C mice. Four weeks post-transplantation, BM-MSC-transplanted striatum appeared larger than the contralateral, PBS-injected striatum (Supplementary Fig. S4A and B). Stereology was used for quantification of total striatal volume of each experimental and control hemisphere. The size of the striatum in PBS-transplanted NP-C mice was approximately 3.9×109 μm3 and was significantly larger in BM-MSC-transplanted striatum (∼5.3×109 μm3, Supplementary Fig. S4B, p<0.05). Therefore, BM-MSCs transplantation resulted in significantly decreased atrophy of the striatum in NP-C mice (p<0.05).
BM-MSCs increase neurotrophic factor signaling
The previously described observations indicated that soluble bioactive factors secreted from BM-MSCs could promote proliferation of NP-C NSCs. To identify factors that were upregulated in BM-MSC-cocultured CM, we conducted screening and comparison of the CM of NP-C NSCs cocultured with and without BM-MSCs for 20 different secreted cytokines using an antibody-based mouse cytokine array (Supplementary Table S1). Notably, the intensities of IL-4 and CCL2 (also known as monocyte chemotactic protein-1, MCP-1) showed greater expression in CM of NP-C NSCs cocultured with BM-MSCs than in CM of NSCs without coculture (Fig. 4A and B). Other candidate factors did not differ significantly between the two groups. These results suggest involvement of CCL2 and IL-4 in the ability for self-renewal of NP-C NSCs by BM-MSCs.
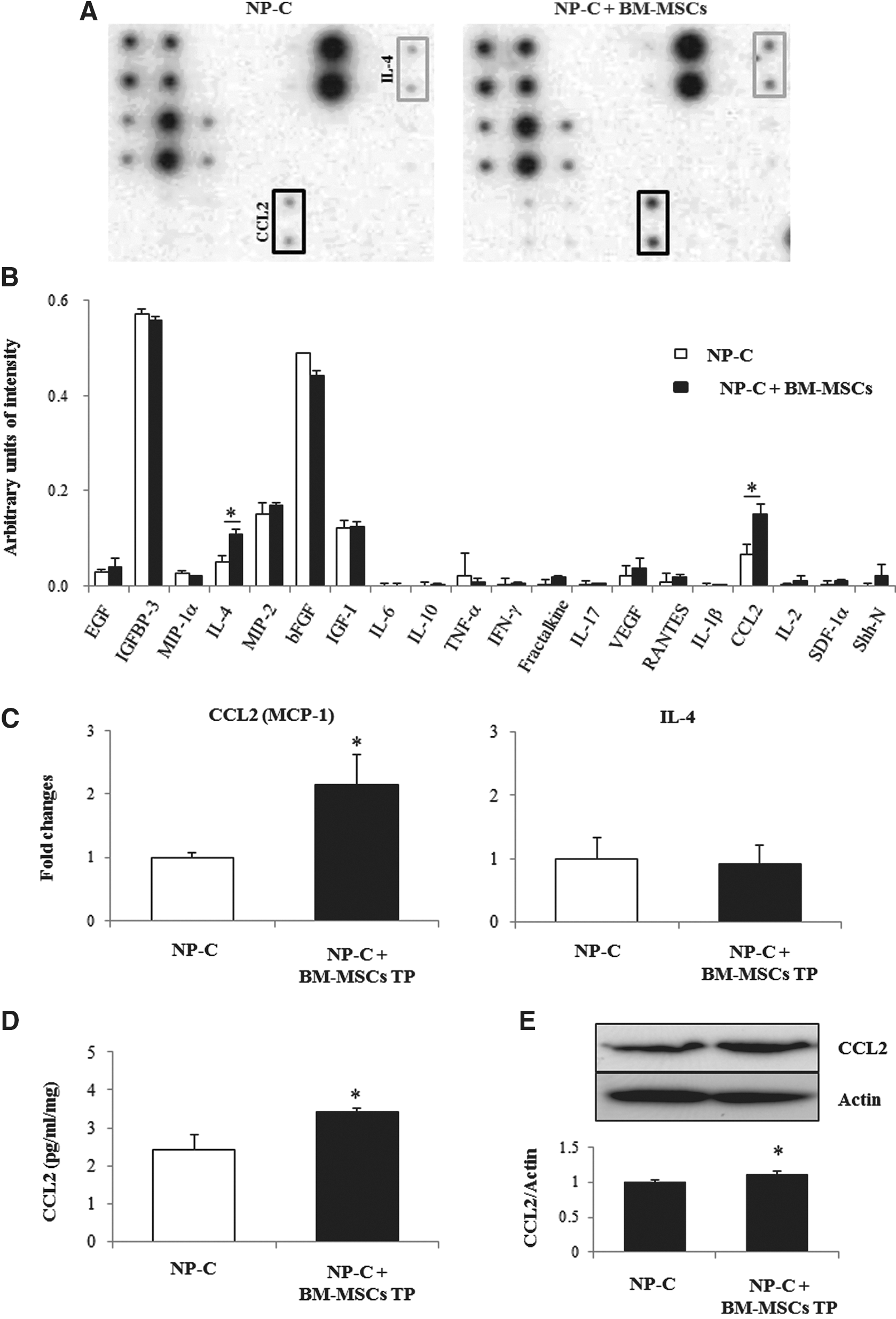
BM-MSCs increased neurotrophic factor signaling. A RayBio custom mouse cytokine Antibody Array (RayBiotech) was incubated with CM recovered from three-dimensional cultures of BM-MSC-treated Npc1 mutant NSCs or untreated Npc1 mutant NSCs.
To confirm our in vitro data, quantitative real-time PCR, ELISA, and Western blot analysis were performed in brains of NP-C mice. Using quantitative real-time PCR, we examined mRNA expression levels of CCL2 and IL-4 in brains of animals belonging to each of our experimental groups (Fig. 4C). Compared with the controls, expression of CCL2 showed a significant increase in the BM-MSC-transplanted NP-C group (p<0.05). However, IL-4 cytokine levels did not differ between the BM-MSC-treated NP-C group and the PBS-treated NP-C group. We also analyzed cerebral CCL2 by CCL2/MCP-1 sandwich ELISA and Western blot analysis. More CCL2 was observed in the SVZ of BM-MSC-treated mice (Fig. 4D and E). Quantitatively, the levels were increased by 41% in the SVZ of BM-MSC-treated mice. The mean value of CCL2 for the PBS-injected NP-C group was 2.42±0.5, and the mean value of CCL2 for the BM-MSC-transplanted NP-C group was 3.43±0.1 (pg/ml/mg, p<0.05).
CCL2 derived from BM-MSCs is a critical factor in increased proliferation and neural differentiation of NP-C NSCs
To study the role of soluble CCL2, we used siRNA for knockdown of CCL2 expression in BM-MSCs (Supplementary Fig. S5A). To elucidate the source of CCL2 in BM-MSC-transplanted SVZ, we transplanted CCL2-knockdown BM-MSC and analyzed the level of CCL2 in SVZ. As shown in Supplementary Fig. S5B, CCL2-knockdown BM-MSC-transplanted NP-C mice showed significantly decreased expression of CCL2 in the SVZ, compared with BM-MSC-transplanted NP-C mice (p<0.05). PBS-treated NP-C mice and CCL2-knockdown BM-MSC-transplanted NP-C mice showed similar levels of CCL2 expression in the SVZ. These results may indicate that increased CCL2 was expressed and secreted from transplanted BM-MSCs.
We next investigated the effect of BM-MSC-derived CCL2 on self-renewal and proliferation of NSCs via in vitro-expanded NP-C cortical NSCs that were indirectly cocultured with BM-MSCs or CCL2-siRNA-treated BM-MSCs. Examination of the efficiency of NS formation was performed 7 days after plating (Fig. 5A and B). Indirect coculture of NP-C cortical NSCs with BM-MSCs resulted in increased formation of NSs (Fig. 5A and B, p<0.001). This effect, however, was significantly lower (p<0.001) after coculture with CCL2-knockdown BM-MSCs (Fig. 5A and B). In addition, the percentage of BrdU+ cells in NP-C NSCs after cocultured with BM-MSCs was increased, as compared with control (Fig. 5C and D, p<0.01). However, the percentage of BrdU+ cells in NP-C NSCs was significantly decreased after coculture with CCL2-knockdown BM-MSCs (Fig. 5C and D, p<0.05). Together, these results indicate that BM-MSCs stimulate proliferation and self-renewal of NP-C NSCs and these responses were mainly mediated by CCL2.

CCL2 derived from BM-MSCs is a critical factor to increase proliferation and neural differentiation of NP-C cortical NSCs. For knockdown of CCL2 in BM-MSCs, cells were pretreated with CCL2 siRNA for 48 hr.
We next investigated the effect of BM-MSC-derived CCL2 on differentiation of NP-C NSCs. Induction of NSC differentiation was performed for 7 days with coculture with BM-MSCs or CCL2-knockdown BM-MSCs using the indirect system to investigate the effect of BM-MSC-derived CCL2 on capacity for neural multilineage differentiation in NP-C NSC cultures. Seven days after coculture, the cells were stained with anti-β-tubulin and anti-O4; according to our observations, a significant increase of the neurogenic (p<0.001) and oligodendrogenic (p<0.01) activities of NP-C NSCs by BM-MSCs was reduced by BM-MSCs expressing CCL2 siRNA (p<0.001, Fig. 5E–H). To confirm these effects of CCL2, cells were treated with a neutralizing antibody to CCL2 (IgG) for a period of BM-MSC cocultures (Supplementary Fig. S6). The number of NSs (Supplementary Fig. S6A and B, p<0.001) and BrdU incorporation (Supplementary Fig. S6C and D, p<0.05) increased in NP-C cultures cocultured with BM-MSCs, compared with control, in the presence IgG control. However, these effects were lower after coculture with BM-MSCs in the presence of a CCL2-neutralizing antibody (Supplementary Fig. S6A–D). The increased neurogenic and oligodendrogenic effects of BM-MSCs in NP-C NSC cultures were reduced by addition of the CCL2-neutralizing antibody (Supplementary Fig. S6E–H).
BM-MSCs or CCL2-knockdown BM-MSCs were transplanted into striatum of NP-C mice in order to determine whether CCL2 released from BM-MSCs is a critical factor in enhancing proliferation of endogenous cells in an NP-C mouse model. Seven days post-implantation, an increase in the number of BrdU+ cells was observed in the BM-MSC-treated NP-C hemisphere when compared with the untreated contralateral hemisphere (p<0.01). This effect, however, was diminished by CCL2 knockdown (Fig. 6A and B). We also quantified new neurons by counting DCX and BrdU-double-positive cells. The number of newborn neurons showed a significant increase (p<0.001) in the BM-MSC-treated NP-C group, compared with the untreated group; however, this effect was abolished in the CCL2-knockdown BM-MSC-transplanted NP-C group (Fig. 6C). At 4 weeks after transplantation, we also performed double-immunostaining using anti-BrdU and NeuN in order to estimate the number of proliferating cells that differentiated into mature neurons (Fig. 6D). The number of new neurons (BrdU+ NeuN + cells) showed an increase in BM-MSC-treated NP-C mice. However, results of CCL2 knockdown in the BM-MSC-transplanted NP-C group did not differ significantly from those of the untreated control group (Fig. 6D). These findings suggest that CCL2 released by BM-MSCs is a critical factor to increased proliferation and neural differentiation of NP-C NSCs.
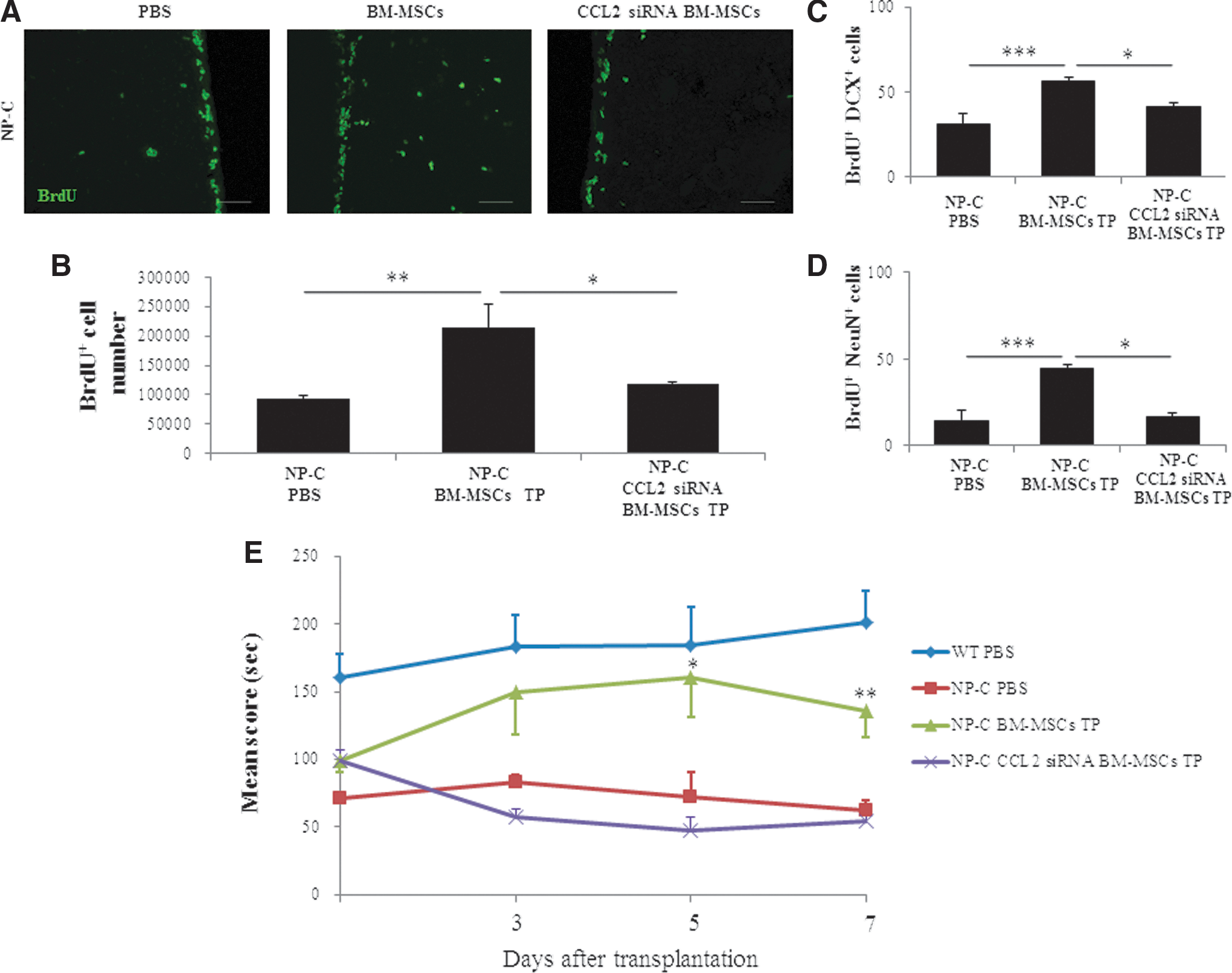
NP-C mice underwent transplantation with BM-MSCs or CCL2-knockdown BM-MSCs at 4 weeks of age. One week and 4 weeks after transplantation, mice were sacrificed and analysis was performed.
We next analyzed whether the CCL2 derived from BM-MSCs elicited functional improvement in NP-C mice. Motor dysfunction and ataxia were evident in the course of degeneration of neuron in NP-C disease (Fink et al., 1989; Imrie et al., 2002). Therefore, Rota-Rod testing was conducted every 48 hr beginning 72 hr after transplantation to assess animal's motor coordination and motor learning. There were improvements in the Rota-Rod score in BM-MSC-treated NP-C mice at 5 and 7 days after transplantation compared with the sham-transplanted NP-C group (Fig. 6E). However, Rota-Rod scores did not differ significantly between CCL2-knockdown BM-MSC-transplanted NP-C mice and control NP-C mice (Fig. 6E). Our results indicate that CCL2 derived from BM-MSCs in SVZ contributes to functional outcome in NP-C mice.
Discussion
The present study is a part of the search for novel therapeutic strategies for treatment of NP-C disease based on the concept of adult neuronal regeneration. Neuropathologically, both human and murine NP-Cs are characterized by ballooned neurons distended with lipid storage, axonal spheroid formation, demyelination, and widespread neuronal loss (Bu et al., 2002). These histological changes, in turn, result in the clinical features observed in patients with psychiatric illness, ataxia, dystonia, and speech difficulties (Sevin et al., 2007; Sedel, 2010). The principal goal of therapy for treatment of NP-C is to restore function to malfunctioning cells, particularly neurons. In practice, cell loss in most patients is sufficient to cause impaired function by the time of diagnosis, implying the necessity for cell replacement to cure the disease (Patterson and Platt, 2004). In the present study, we observed a significant increase of BrdU+ cells in the SVZ of NP-C mice, compared with WT mice (Fig. 3). However, neurogenesis of finally differentiated neurons (NeuN+BrdU+ cells) in NP-C mice remained lower than that in WT mice (Fig. 3). Therefore, NP-C diseases are suitable for stimulation of neurogenesis for treatment of patients with disease.
Several neurogenic activities associated with BM-MSCs have been discussed (Zhang et al., 2004; Deng et al., 2006). Our findings are in agreement with previous investigations on the stimulatory effect of BM-MSCs on neurogenesis (Munoz et al., 2005; Yoo et al., 2008; Croft and Przyborski, 2009). In vitro observations have demonstrated that indirect coculture of BM-MSCs could result in alleviation of the self-renewal and proliferation defect of Npc1 mutant NSCs and enhancement of differentiation into neurons or oligodendrocytes in vitro (Figs. 1 and 2). These findings suggest that the effects of BM-MSCs on self-renewal, proliferation, and differentiation can be attributed to the indirect effect of soluble factors secreted from BM-MSCs and that these cells selectively promote neurogenesis and oligogenesis from NSCs. In studies using in vivo BrdU exposure, we have also demonstrated that transplantation of BM-MSCs into the SVZ of NP-C mice can result in stimulated proliferation and maturation of endogenous progenitors toward the neuronal phenotype (Fig. 3). The finding of an increase in proportion of newborn cells (DCX+BrdU+ cells and NeuN+BrdU+ cells) within the lateral walls of the LVs may indicate selective stimulation of neuronal differentiation. In addition, transplanted BM-MSCs (labeled with nanoparticles) remained near the transplantation site and were never observed along the walls of the LV. Results of this analysis confirmed that newborn neurons were derived from endogenous neuronal progenitors and were not the transplanted cells. It is possible that soluble factors secreted by the transplanted BM-MSCs may mediate regulation of endogenous SVZ neurogenesis in NP-C disease.
The neurogenic effect of BM-MSCs is mediated by a variety of soluble factors (Croft and Przyborski, 2009). Although the differentiation potential of BM-MSCs for tissue repair has been repeatedly demonstrated, one of the constant finding from several experiments was that these cells frequently produced functional improvements without significant evidence of either engraftment or differentiation (Munoz et al., 2005; Parr et al., 2007; Prockop, 2007). These studies suggest that the repair of tissue by BM-MSCs does not occur solely as a result of their stem-cell-like ability to differentiate or fuse with existing cells, but may also be attributed to release of growth factors and other molecules that may elicit therapeutic effects (Prockop et al., 2003; Iso et al., 2007). To assess for paracrine factors that mediate MSC effects on proliferation of NP-C NSCs, we performed cytokine array for analysis of cocultured media samples (e.g., neurotrophins, growth factors). Our results demonstrated increased expression of CCL2 and IL-4 in BM-MSC-cocultured CM, compared with nontreated CM (Fig. 4). We further demonstrated a significant increase in levels of CCL2 expression in the BM-MSC-transplanted NP-C brain (Fig. 4). In addition, we also observed that the effects of BM-MSCs on proliferation and differentiation were considerably decreased in the CCL2-knockdown BM-MSC-treated NP-C group (Figs. 5 and 6). Previous research has shown that, independent of neuropathology, chemokines and their receptors are not only expressed in the developing brain, but also have a role in orienting cell migration, trophic support, proliferation, and/or differentiation (Bajetto et al., 2001). As such, MCP-1/CCL2 may act as a modulator of neuronal activity and neuroendocrine functions (Banisadr et al., 2005). Recent data have indicated an important role for CCL2 in migration and SVZ differentiation of neural progenitor cells after stroke (Liu et al., 2007). Therefore, these results suggest CCL2 as an effective adjuvant for improvement of neurogenic effects and that CCL2 derived from BM-MSCs may be a critical factor leading to increased proliferation and neural differentiation of NP-C NSCs.
In summary, our in vitro and in vivo data suggest that use of BM-MSCs can result in improvement of the developmental deficit of NP-C NSCs by augmentation of decreased proliferation and differentiation, restoration of neurogenesis impairment, as demonstrated by significantly increased levels of BrdU and colocalization of DCX and NeuN, and, ultimately, induction of SVZ neurogenesis and recovery of cerebral atrophy.
In conclusion, findings of the present study have provided evidence for a role for BM-MSCs as a key mediator of adult neurogenesis in adult SVZ, which may in turn have implications for management of NP-C disease. Stem cell transplantation therapies for treatment of central nervous system pathologies are promising therapeutic strategies for management of neurodegenerative diseases, including NP-C disease (Bae et al., 2002, 2005a; Christensen et al., 2002). Regenerative effects of stem cell transplantation might involve both replacements of damaged cells by exogenous cells as well as improvement of endogenous repair processes by release of trophic factors (Qu et al., 2007; van Velthoven et al., 2009).
Authors' Contributions
H.L. and J.E.K. performed the experiments and wrote the article. J.K.L. performed the experiments. J.-S.B. and H.K.J. supervised the project, designed the research, wrote the article, and reviewed the article.
Footnotes
Acknowledgments
This work was supported by the Bio & Medical Technology Development Program (2011-0019356) and Basic Science Research Program (2010-0009421) of the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology, Republic of Korea. Additional support for this work was provided by Kyungpook National University Research Fund, 2012.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.