
Abstract
The ability to deliver a large transgene efficiently to photoreceptors using viral vectors remains problematic and yet is critical for the future therapy of inherited retinal diseases such as Stargardt's and Usher's 1B. Herein, we examine the ocular tropism of a HIV-1-based lentivirus vector pseudotyped with Venezuelan equine encephalitis virus-derived glycoprotein (VEEV-G) after intraocular delivery to the posterior and anterior chambers of C57BL/6 wild-type mice. Reporter gene (EGFP) expression was evaluated using in vivo fluorescence imaging followed by postmortem immunohistochemistry and retinal function assessed by electroretinography. Intracameral administration of VEEV-G and vesicular stomatitis virus glycoprotein (VSV-G)-pseudotyped vectors resulted in robust transgene expression in the corneal endothelium and trabecular meshwork. After subretinal administration, onset of transgene expression was observed in the retinal pigment epithelium (RPE) 1 day postinjection with both VEEV-G and control VSV-G pseudotypes, but no significant photoreceptor transduction was apparent. Substantial degeneration of the outer nuclear layer was observed with VEEV-G-pseudotyped vector, which corresponded to ablation of retinal function. Subretinal administration of VSV-G was observed to result in significant suppression of electrophysiological function compared with buffer-injected and uninjected control eyes. Suppression of the c-wave amplitude, in addition to reduced RPE65 expression, indicated potential RPE dysfunction. Ex vivo tropism of VSV-G was assessed using organotypic culture of explanted retina harvested from wild-type mice and human patients undergoing retinal detachment surgery to examine the prevention of transduction by physical barriers and species differences in tropism.
Introduction
I
Lentivirus vectors, typically based on human/simian immunodeficiency virus (HIV/SIV) or equine infectious anemia virus (EIAV), are a viable alternative, having a comparatively large coding capacity and the ability to be pseudotyped, a process whereby the native viral surface glycoproteins are exchanged for those of another enveloped virus, thus altering the tropism. The most commonly employed pseudotype, derived from vesicular stomatitis virus glycoprotein (VSV-G), has broad cellular tropism because of an affinity for binding to ubiquitously expressed surface residues, most likely negatively charged lipids (Lyles and Rupprecht, 2006). VSV-G-pseudotyped vectors have demonstrated efficient transduction of RPE in vivo after delivery to the subretinal space, yet have thus far proven ineffective at targeting photoreceptors in rodent (Auricchio et al., 2001; Ikeda et al., 2003; Bemelmans et al., 2005; Greenberg et al., 2007) and primate models (Ikeda et al., 2009), limiting their utility for delivering large genes necessary for the treatment of several retinal diseases, such as Stargardt's disease or Usher syndrome type Ib. Attempts to improve neuronal targeting by employing pseudotypes derived from neurotropic viruses, such as rabies-G (Balaggan et al., 2006) or mokola (Bemelmans et al., 2005), have met with little success. Similarly, while photoreceptor-specific promoters have been successfully incorporated into both HIV and EIAV vectors, they effectively limit ectopic expression of the transgene, but do not significantly increase the levels of photoreceptor transduction in the adult retina (Miyoshi et al., 1997; Nicoud et al., 2007). Herein, we employ longitudinal in vivo fluorescence imaging to examine the tropism of a further pseudotype derived from Venezuelan equine encephalitis virus (VEEV-G), a zoonotic virus that causes encephalopathy in various equine species and humans.
In addition to evaluating retinal tropism after subretinal administration, we also examine the tropism of the VEEV-G pseudotype in the anterior chamber, an ocular niche where VSV-G-pseudotyped vectors have previously been shown to effectively transduce the corneal endothelium and trabecular meshwork (Balaggan et al., 2006). The tropism of VEEV-G-pseudotyped vector is compared throughout with concurrently produced dose-matched VSV-G vector by histology, and the effect of subretinal delivery on retinal function assessed for both vectors by electroretinography (ERG).
It has previously been proposed (Gruter et al., 2005) that the lack of observed photoreceptor transduction after subretinal delivery of lentivirus vectors may not be caused by the receptor binding of a given pseudotype, but by the presence of physical barriers, such as the outer limiting membrane or outer segment disc shedding, which may impede virion access to the photoreceptor cell bodies. To examine this clinically important question, we utilized a modified explant culture system to assess virus tropism in the absence of physical barriers in the murine retina, in addition to retina biopsied from human patients undergoing surgery for retinal detachment.
Materials and Methods
Viral vectors
Recombinant nonreplicative HIV-1-based lentiviral vectors were produced using a modified transient calcium phosphate transfection protocol based on the standard four-plasmid transfection of 293T cells. Briefly 12×150 mM tissue culture dishes were seeded with 1.4–1.6×107 293T cells per dish. The next day, cells were transfected with 15 μg vector plasmid (pRRLsincppt-CMV-eGFP-WPRE expressing GFP from the CMV promoter), 15 μg of plasmid expressing the HIV-1 gag/pol gene (pMD2-LgpRRE), 3 μg of plasmid expressing HIV-1 Rev (pRSV-Rev), and 5.1 μg of the plasmid expressing the appropriate envelope GP pMD2-VSVg (this and other HIV1 vector plasmids were obtained from Professor James Uney, University of Bristol, United Kingdom) or pcDNA3-VEEV.3908 (obtained from Dr. Robert A. Davey, University of Texas) after the addition of 1 M CaCl2. About 16 hr posttransfection, media were replaced with fresh media supplemented with 10 mM sodium butyrate. About 36 hr after addition of sodium butyrate, vector-containing media were harvested and filtered through a 45 μm filter.
Large-scale preparations were concentrated by centrifugation at 6,000 RPM for 12–16 hr at 4°C (Beckman Coulter Avanti J-E; F500 rotor). The pellet was then resuspended in ice-cold PBS and was further concentrated by ultracentrifugation for 90 min at 20,000 RPM and 10°C (Beckamn Coulter Optima L-80XP). The pellet was resuspended in 50 μl ice-cold TSSM formulation buffer (20 mM Tromethamine, 100 mM NaCl, 10 mg/ml sucrose, and 10 mg/ml mannitol) every 15 min for 3 hr and then centrifuged at 13,000 RPM for 30 sec. The supernatant was stored overnight at 4°C along with the pellet that was resuspended with a further 50 μl ice-cold TSSM. The resuspended pellet was centrifuged at 13,000 RPM for 30 sec, and the two supernatants were pooled, giving a final volume of 100 μl (2,000-fold concentration). Vector preparations were stored at −80°C.
Biological activity of lentiviral vectors carrying the GFP reporter gene was determined by flow cytometry (Becton Dickinson LSR Benchtop Flow Cytometer). Briefly, HEK 293T cells were seeded in 12-well plates at 5×105 cells per well. About 18 hr after seeding the cells in a single well were quantified using a hemocytometer. Cells were transduced for 6 hr with a serial dilution of the vector to be quantified in the presence of 8 μg/ml polybrene. About 72 hr posttransduction, the percentage of GFP-positive cells was determined by flow cytometry. The number of physical lentiviral particles present in a preparation was estimated using a HIV-1 p24 Antigen ELISA (RETROtek from ZeptoMetrix).
Mice and anesthesia
All animal experiments were performed in compliance with the ARVO statement for the Use of Animals in Ophthalmic and Vision Research and ARRIVE guidelines, and were conducted with U.K. Home Office approval. Adult (6–8 weeks old) C57BL/6 mice were purchased from either Harlan Laboratories (C57BL/6JOlaHsd) or Charles River Laboratories (Jax C57BL/6J) and housed under standard 12:12 light–dark cycle with food and water available ad libitum. For all procedures, general anesthesia was induced by a single intraperitoneal injection of Dormitor medetomidine hydrochloride (Dormitor, 1 mg/kg body weight; Pfizer) and ketamine (Ketaset, 60 mg/kg body weight; Fort Dodge), and their pupils were fully dilated with 1% tropicamide and 2.5% phenylephrine hydrochloride eye drops (both from Bausch & Lomb). Where appropriate, anesthesia was reversed following procedures by intraperitoneal injection of atipamezole hydrochloride (Antisedan, 5 mg/kg body weight; Pfizer).
Intraocular injection
After pupil dilation, a gel lubricant (Viscotears; Novartis) was used to position a 6 mM circular cover slip over the cornea, allowing good visualization of the retina when viewed under a surgical microscope (M620 F20; Leica). Injections were performed by advancing a Hamilton syringe with attached 10 mM 34-guage needle (Hamilton 65N; Hamilton AG) trans-sclerally into the subretinal space (subretinal injection) or further, through the neural retina, into the vitreous chamber (intravitreal injection), or into the anterior chamber (intracamerally). Reflux of vector suspension was limited by allowing the intraocular pressure to normalize for 20–30 sec before rapid removal of the needle. Orientation and stabilization of the eye was maintained throughout by holding the superior or inferior rectus muscle with notched forceps. Separate syringes were used for each vector pseudotype to avoid contamination and the needles flushed with sterile saline between injections.
In vivo fluorescence imaging using a confocal scanning laser ophthalmoscope
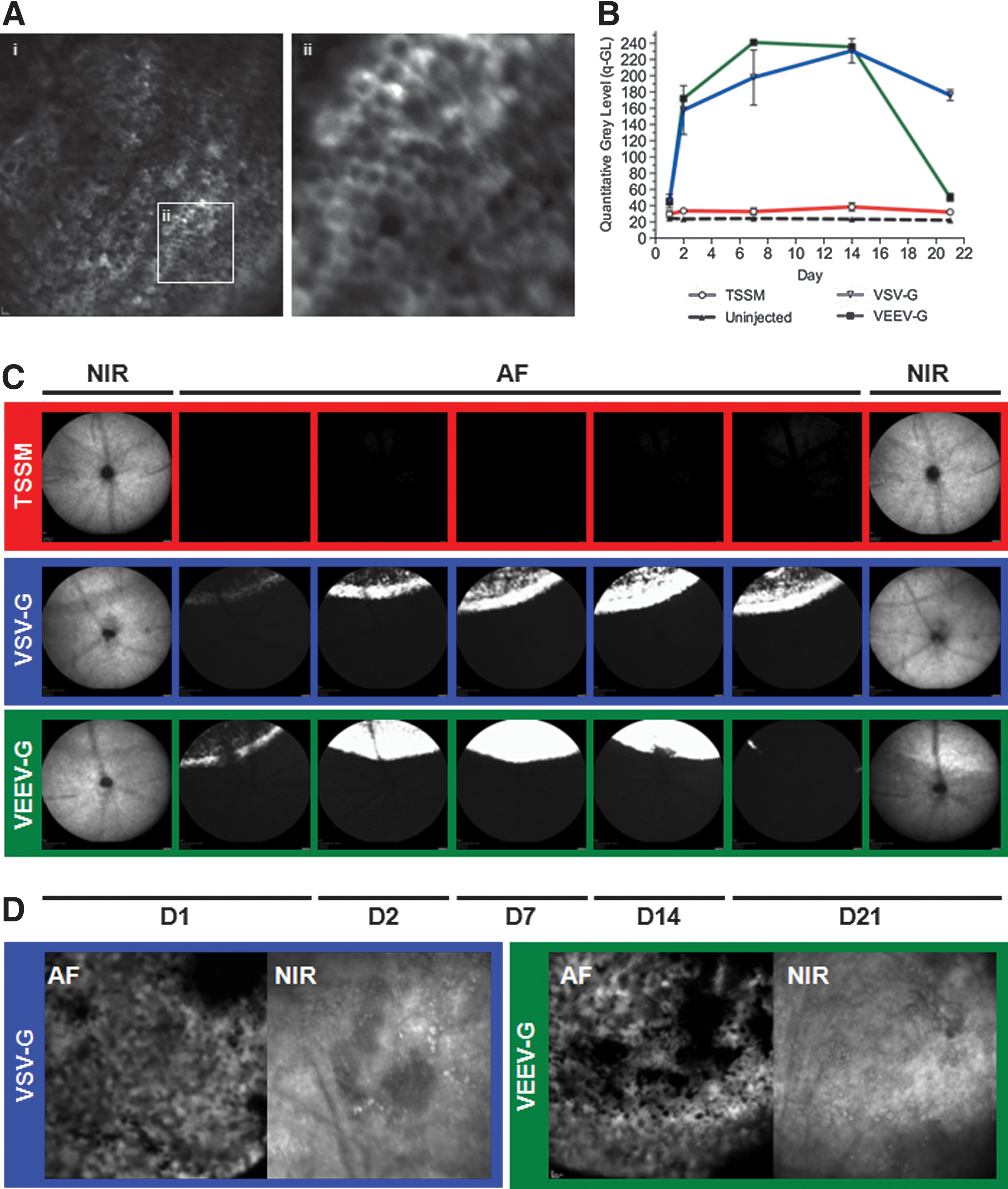
The ocular fundi of each mouse were imaged using a confocal scanning laser ophthalmoscope (Spectralis HRA; Heidelberg Engineering) 1, 2, 7, 14, and 21 days (D) postinjection, as previously described (Charbel Issa et al., 2011). Briefly, after dilation a contact lens was placed on the cornea using a viscous coupling gel (0.3% w/v hypromellose) to prevent cataract formation and to improve and standardize image quality. Each mouse was placed on a raised imaging platform, and the near-infrared (NIR) reflectance mode (820 nm laser) used to achieve camera alignment at the confocal plane of the RPE. Onset of EGFP transgene expression was evaluated using the autofluorescence (AF) imaging mode (480 nm excitation) and high-resolution images (1,536×1,536) recorded at a range of standardized sensitivities using automated real-time averaging.
Tissue processing and histology
Eyes were enucleated and fixed in 4% paraformaldehyde overnight, during which time the anterior segment and lens were removed. The posterior eye cup was cryoprotected for 48 hr at 4°C in 30% (w/v) sucrose and embedded in optimum cutting temperature media, and 18 μm cryosections were taken coronally from the optic disc toward the iris. Sections were immunostained for either RPE65 (1:1,000 dilution; antibody was a kind gift from Rosalie Crouch, Medical University of South Carolina) or cone arrestin (1:1,000; Millipore AB15282), before being counterstained with Hoechst 33342 and mounted with ProLong Gold antifade reagent (both from Invitrogen).
Electroretinography
ERG recording was carried out using an ERG system as described previously (Espion E2; Diagnosys LLC) (Lipinski et al., 2011b). After dark adaptation overnight, brief (4 msec) white flash stimuli were delivered over a eight-log intensity series (−6 to 1.4 log cd·sec/m2) in otherwise dark conditions as follows: −6 to −5 log cd·sec/m2, 16 responses were averaged with an interstimulus internal (ISI) of 1 sec; −4 to −3 log cd·sec/m2, 9 responses, ISI=1 sec; −2 log cd·sec/m2, 9 responses, ISI=5 sec; −1 log cd·sec/m2, 4 responses, ISI=13 sec; 0 log cd·sec/m2, 4 responses, ISI=29 sec; 1 log cd·sec/m2, 4 responses, ISI=61 sec; 1.4 log cd·sec/m2, 1 response. Light-adapted (30 cd/m2 white background, >10 min) flicker recordings were carried out with continuous 20 Hz, 25 cd·sec/m2 stimuli with 20 response epochs (500 msec) averaged for each condition. The a-, b-, and c-wave amplitudes (flash stimuli) and trough-to-peak amplitudes (flicker stimuli) were quantified with specialized software (Espion; Diagnosys LLC).
Ex vivo culture of human retinal explants
Retinal explant samples were obtained from patients undergoing retinectomy as part of complex retinal detachment surgery with approval from the U.K. Research Ethics Committee (REC 10/H0505). In accordance with the Declaration of Helsinki, patients gave written, informed consent before the procedure's commencement. A routine 23-gauge pars plana vitrectomy was performed for retinal detachment surgery. Discs of the peripheral retina were cut with a vitrectomy system (Ocutome; Alcon Surgical), refluxed into the eye, aspirated with a flute needle, and placed in a balanced salt solution. Within 30 min, retinal samples were transferred into organotypic culture, as describe previously (Lipinski et al., 2011a). Briefly, retinal tissue was transferred using a cut-off 3 ml Pasteur pipette (to reduce crush injury) into 24-well tissue culture inserts containing 700 μl complete culture medium (Johnson and Martin, 2008) consisting of NeurobasalA, L-glutamine (0.08 mM), penicillin (100 U/ml), streptomycin (100 U/ml), Amphotericin B (0.25 μg/ml), B27 supplement (2%), and N2 supplement (1%; all from Invitrogen). One day (D1) postexplantation, the medium was replaced and supplemented with 10 μl virus vector suspension (VEEV-G=1.7×107 TU; VSV-G=2.6×107 TU); the medium was changed every third day thereafter. Transparency of the tissue culture inserts (cat. 353095; BD Falcon) allowed explants to be monitored for onset of transgene expression in culture using an inverted epi-fluorescent microscope (DMIL; Leica). At D14, explants were fixed in 4% paraformaldehyde for 24 hr at 4°C, during which time both outer and inner retinal aspects were imaged. Subsequently, explants were cryoprotected for 48 hr in 30% (w/v) sucrose and embedded in optimum cutting temperature, and 18 μm cryosections were cut.
Statistical analysis
Two-way analyses of variance (ANOVAs) were performed on uncorrected raw data to determine significant differences in group means. The quantitative gray level (q-GL) was compared using vector pseudotype and titer as factors at given time points. Bonferroni or Sidak's post-hoc tests were applied where appropriate. Differences in flicker amplitudes were assessed by paired t-tests. In all tests, a p-value of ≤0.05 was considered statistically significant.
Results
In vivo fluorescence imaging reveals rapid onset of transgene expression in the RPE after subretinal administration
Confocal scanning laser ophthalmoscope imaging is employed clinically to noninvasively assess retinal pathology, where the retina can be visualized through the pupil aperture. Observations can be used to inform diagnosis, guide treatment, and allow monitoring of disease progression. The AF mode utilizes a 488 nm laser for excitation that can also be used to examine the ocular fundus in mice (Charbel Issa et al., 2011), and may visualize retinal expression of fluorescent reporter proteins, particularly EGFP, which has an excitation maximum proximate to that of the AF laser stimulus (Beck et al., 2010). Pseudotyped VSV-G (5.20×106 TU in 2 μl) or VEEV-G (3.40×106 TU in 2 μl) lentiviral vectors carrying an EGFP transgene (Supplementary Fig. S1A and B; Supplementary Data are available online at
Robust transgene expression in corneal endothelium and the trabecular meshwork after intracameral injection of pseudotyped lentivirus
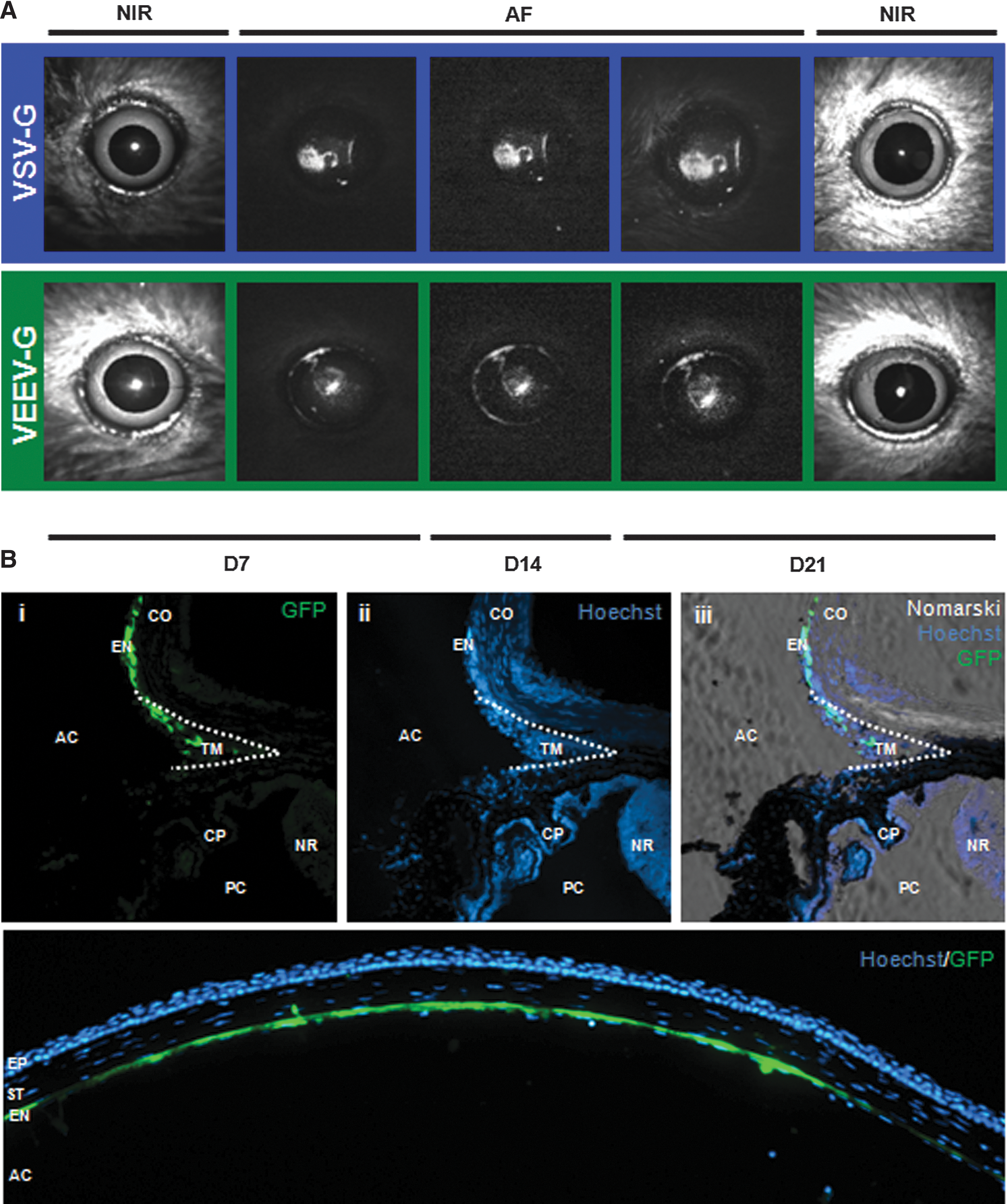
VSV-G-pseudotyped vector has previously been described to transduce cells of the anterior segment, in particular the corneal endothelium and trabecular meshwork (Balaggan et al., 2006). Given the robust RPE expression of the VEEV-G pseudotype observed after subretinal administration, we examined the tropism of this vector after administration into the anterior chamber. VSV-G (5.20×106 TU in 2 μl) or VEEV-G (3.40×106 TU in 2 μl) vector was injected intracamerally in six C57BL/6 mice (n=6 per pseudotype) through a self-sealing corneal tunnel made at the surgical limbus. Three further C57BL/6 mice acted as injection controls receiving 2 μl TSSM injection in one eye, with the contralateral remaining uninjected. The virus suspension was delivered to the center of the anterior chamber, where aqueous convection is most likely to drive virions toward the inner corneal surface, and aqueous outflow to drive virions radially toward the iridocorneal angle. NIR imaging was used to obtain a centered image of the anterior chamber; reflectance artifacts (white glare spots) were observed on the corneal surface only in the NIR mode (Fig. 2A). AF imaging revealed expression of EGFP from both pseudotypes by D7, which was maintained until D21 (Fig. 2A). Fluorescence was predominantly observed centrally at the confocal plane of the cornea with both pseudotypes, indicating robust expression of EGFP in the corneal endothelium, which was later confirmed by histology (Fig. 2B). The pattern of fluorescence was in an atypical “Krukenberg spindle” distribution, a phenomenon observed whereby aqueous convection drives free-floating particles in the anterior chamber (classically, pigmented iris cells) toward the interior corneal surface, leading to their deposition in a central, vertically orientated stripe (Krukenberg, 1899). Fluorescence was observed to have radial symmetry (e.g., Fig. 3A, vi), being most intense centrally and less intense peripherally. The absence of a classic Krukenberg spindle pattern (vertically orientated ovoid) likely results from differences in the size and shape of the mouse lens relative to the cornea (Heys and Barocas, 2002). AF imaging focusing at the plane of the limbus revealed a ring of fluorescence at the irido-corneal angle, which on histology correlated to EGFP expression in the anatomical area of the trabecular meshwork (Fig. 2B), a structure that regulates aqueous outflow and is critical to maintaining proper intraocular pressure.

Representative histology demonstrating the expression pattern of EGFP after subretinal injection in C57Bl/6J mice of either VEEV-G or VSV-G vectors. Both pseudotypes show some co-localization of the EGFP signal with RPE65
VEEV-G-pseudotyped lentiviral vector exhibits toxicity in the outer retina
After fixation and embedding at D21 (Supplementary Fig. S1C), injected eyes were sectioned for histology and immunohistochemistry to determine the cellular tropism. In both groups, robust EGFP expression was observed to co-localize with areas of heavy pigmentation (Fig. 3), strongly indicating transduction of the RPE within the area of injection. EGFP expression was observed to co-localize with RPE65, an RPE-specific retinal isomerase, although levels appeared to be reduced in areas of greatest EGFP expression (Fig. 3, i–iii, vii–ix). Downregulation of RPE65 has previously been observed to result from cellular stress potentially preceding apoptosis (Alizadeh et al., 2001; Alge et al., 2003; Lee et al., 2010). However, no morphological evidence of RPE thinning, cell migration, or death was observed compared with surrounding uninjected areas with either vector. EGFP expression was not observed in the outer nuclear layer (ONL) after injection with either pseudotype, in accordance with findings described previously (Auricchio et al., 2001; Bainbridge et al., 2006; Balaggan et al., 2006). Immunohistochemistry with a marker of cone photoreceptor (c-arrestin) revealed no co-localization (Fig. 3, iv–vi, x–xii), strongly indicating that neither pseudotype successfully targets murine photoreceptors in vivo. Thinning of the ONL within the area of injection was observed in eyes administered the VEEV-G-pseudotyped vector (Fig. 3, vii–xii). No retinal thinning was observed with the VSV-G vector (Fig. 3, i–vi), or in TSSM-injected control eyes (data not shown). Disruptions in cellular pathology and decreased electrophysiological responses have previously been noted in rodents after administration of high-titer lentivirus vector suspensions, and are likely caused by a sustained inflammatory reaction (Ikeda et al., 2009). Areas where EGFP expression was absent were visible within the area of injection (e.g., Fig. 3, i and vii), which likely correspond to areas of decreased AF observed in vivo (Fig. 1B and C). However, the RPE appeared normal in morphology with no evidence of cellular infiltration into the subretinal space (Doi et al., 2002), limited RPE migration (Fig. 3, vi), and no depigmentation, which would indicate significant cellular pathology.
To address the possibility of toxicity being associated with vector production (e.g., residual chemical contaminants), injections were repeated with multiple vector preparations and ONL thinning was observed in each instance with the VEEV-G pseudotype, but not with concurrently produced VSV-G vector batches of comparable titer, indicating a pseudotype-specific effect. TSSM-injected eyes additionally showed no indicators of toxicity. While expression of various fluorescent proteins has been shown to result in cytotoxicity both in vitro (Liu et al., 1999) and in vivo (Huang et al., 2000), photoreceptor toxicity arising from intracellular expression of EGFP is unlikely, given the lack of observed ONL transduction. Furthermore, EGFP has been shown to be well tolerated in the retina, with no significant cell death observed when EGFP is expressed after transduction of photoreceptors using alternative vectors (e.g., rAAV), or when EGFP is expressed intrinsically, as in various transgenic mouse models (Dreyer et al., 1999; Daniels et al., 2003; Nour et al., 2004; Lipinski et al., 2011b). The rapid time course of ONL degeneration (<3 weeks) makes it unlikely that cell death arises from RPE dysfunction (e.g., down regulation of RPE65), where photoreceptor loss in models of RPE dysfunction typically occurs over several months (Pang et al., 2005; Fan et al., 2008). While the mechanism of degeneration remains unclear in this instance, together the data suggest that photoreceptor loss is not the result of EGFP toxicity, does not occur secondary to RPE dysfunction, and is specific to the VEEV-G pseudotype.
Subretinal injection of VEEV-G and VSV-G leads to a significant reduction in electrophysiological function
Because of the observation of reduced in vivo fluorescence and evidence of ONL thinning in VEEV-G-injected eyes, we examined retinal function by ERG. ERG measures changes in corneal field potential that are propagated from the retina after stimulation with light, and can be used to assess the function of distinct cell types within the retina. For safety assessment, each eye received an injection under both the dorsal and ventral retina (1.5 μl volume for each), which resulted in a detachment of 70–100% of the total retinal area. ERGs were then performed at D21 in C57BL/6 mice that had received VEEV-G (4.14×106 TU in 3 μl; n=6 mice), VSV-G (8.75×106 TU in 3 μl; n=6 mice), or TSSM buffer (3 μl; n=6 mice) in one eye; the contralateral eye in each animal remained uninjected to allow pairwise assessment of vector safety. Recordings were initially made in dark-adapted (scotopic) conditions to facilitate the assessment of rod photoreceptor and RPE function (Fig. 4A–N). Scotopic recordings were carried out over an 8-log intensity series (10−6–25 cd·sec/m2) with a-, b-, and c-waves measured at each intensity (Fig. 4P). A major ERG component is the a-wave—a large negative deflection in corneal potential that reflects activation of the photoreceptors (Fig. 4P). The dark-adapted a-wave is predominantly an assessment of rod function, primarily because rods comprise ∼97% of the ONL in most mammals, including mice. In all treatment groups, the a-wave was found to be intensity dependent, appearing and becoming larger as the brightness of the stimulus was increased. However, subretinal injection of TSSM (Fig. 4A and B) resulted in a significant reduction in a-wave at the highest intensities versus uninjected control eyes (10 and 25 cd·sec/m2; p<0.0001, paired 2-way ANOVA with Sidak's test for multiple comparison); this reduction reflects reduced function as a result of surgically detaching the retina from the RPE. The reduction in a-wave amplitude was >2-fold greater in VSV-G-injected eyes (Fig. 4F and G) than was observed in the TSSM-injected group (10 and 25 cd·sec/m2; p<0.0001). These data suggest that subretinal injection of VSV-G-pseudotyped lentivirus has a greater negative effect on rod function than can be explained by surgical detachment alone. The a-wave was absent in VEEV-G-injected eyes (Fig. 4K and L) at all intensities (10−4–25 cd·sec/m2; p<0.0001 vs. contralateral controls), indicating substantial photoreceptor toxicity when VEEV-G-pseudotyped virions are injected subretinally, in accordance with the ONL thinning observed on histology.

Assessment of retinal function by ERG. Representative dark-adapted, flash ERG traces for TSSM-
Another major ERG component is the b-wave, a large positive deflection immediately following the a-wave, that relates to activation of ON-type bipolar cells; the b-wave consequently represents signal propagation to second-order neurons from both rod and cone photoreceptors (Fig. 4P). Consequently, the b-wave was affected in a similar way to the a-wave overall. Specifically, a slight decrease in b-wave amplitude was observed after TSSM injection compared with uninjected control eyes (Fig. 4A and B) at higher intensities (1–25 cd·sec/m2; p<0.05, paired 2-way ANOVA with Sidak's test for multiple comparison). In addition, a highly significant decrease in b-wave amplitude was observed in VSV-G-injected eyes (Fig. 4F and G) compared with uninjected controls (10 cd·sec/m2, p<0.01; 25 cd·sec/m2, p<0.001). VEEV-G-injected eyes (Fig. 4K–M) had no observable b-wave at any intensity (10−5–25 cd·sec/m2; p<0.0001 vs. contralateral controls), indicting the absence of photoreceptor input to second-order neurons.
A final important component of the dark-adapted ERG is the c-wave, a slow positive deflection that originates from the RPE, but depends on the integrity of the overlying photoreceptors for initiation, and as such is a measure of photoreceptor–RPE interaction. The c-wave was significantly reduced in TSSM-injected eyes (Fig. 4A and C) at higher intensities (10 and 25 cd·sec/m2; p<0.001, paired 2-way ANOVA with Sidak's test for multiple comparison), indicating disruption of the photoreceptor–RPE interface by the surgical detachment. The c-wave is reduced (1–25 cd·sec/m2; p<0.0001) to a greater extent in VSV-G-injected eyes (Fig. 4F and H), indicating that the presence of VSV-G-pseudotyped virions has a negative impact on the photoreceptor–RPE interaction when delivered subretinally. The c-wave is unrecordable in VEEV-G-injected eyes at all intensities (10−4–25 cd·sec/m2; p<0.0001 vs. contralateral controls), again indicting that photoreceptors are no longer light sensitive or are absent.
Cone function was directly assessed by light-adapted flicker ERG, where a rapidly flickering (20 Hz) high-intensity light stimulus (25 cd·sec/m2) was superimposed on a rod-saturating background (30 cd/m2); several spikes were measured (trough-to-peak) per mouse and the amplitude averaged. Photopic flicker amplitude was not significantly reduced in TSSM-injected eyes (Fig. 4D and E) compared with uninjected controls (p=2.894, paired t-test), indicating no significant reduction in cone function as a result of surgical detachment. On the other hand, cone function was significantly reduced in VSV-G-injected eyes (Fig. 4I and J) compared with uninjected controls (p=0.0469, paired t-test). Cone responses were not recordable above background noise in VEEV-G-injected eyes (p=0.0059), indicating significant cone loss or dysfunction.
Ex vivo transduction of human retinal explants reveals limited transduction of photoreceptors in human tissue
Despite observing suppression of photoreceptor and RPE function in VSV-G-injected eyes after in vivo delivery, we aimed to explore whether the absence of observed photoreceptor transduction was caused by the presence of physical barriers. Using a modified explant culture system (Lipinski et al., 2011a) to maintain small samples (2–3 mm2) of murine retina, we were able to examine VSV-G tropism in the absence of physical barriers, such as the outer segments and outer limiting membrane, which are compromised during the process of explantation. Additionally, the RPE is absent from the tissue culture environment, and the rate of outer segment disc shedding is greatly reduced, further reducing impediments to virion access. One day postexplantation, retinal samples taken from C57BL/6 mice were transduced by the addition of 7.29×108 TU VSV-G diluted in 700 μl culture medium; an ∼100-fold higher titer was utilized for ex vivo transduction to correct for the larger culture volume (700 μl ex vivo vs. 2–3 μl in vivo) and the loss of virions in culture, for example, through adherence to plasticware. Histology of explanted murine tissue at D14 revealed no evidence of photoreceptor transduction, with no co-localization of EGFP with recoverin, a photoreceptor-specific marker (Fig. 5A, i and ii). Long EGFP-positive processes were observed in some tissue, but no intracellular EGFP expression was observed in cells of the ONL (Fig. 5A, iii–v).
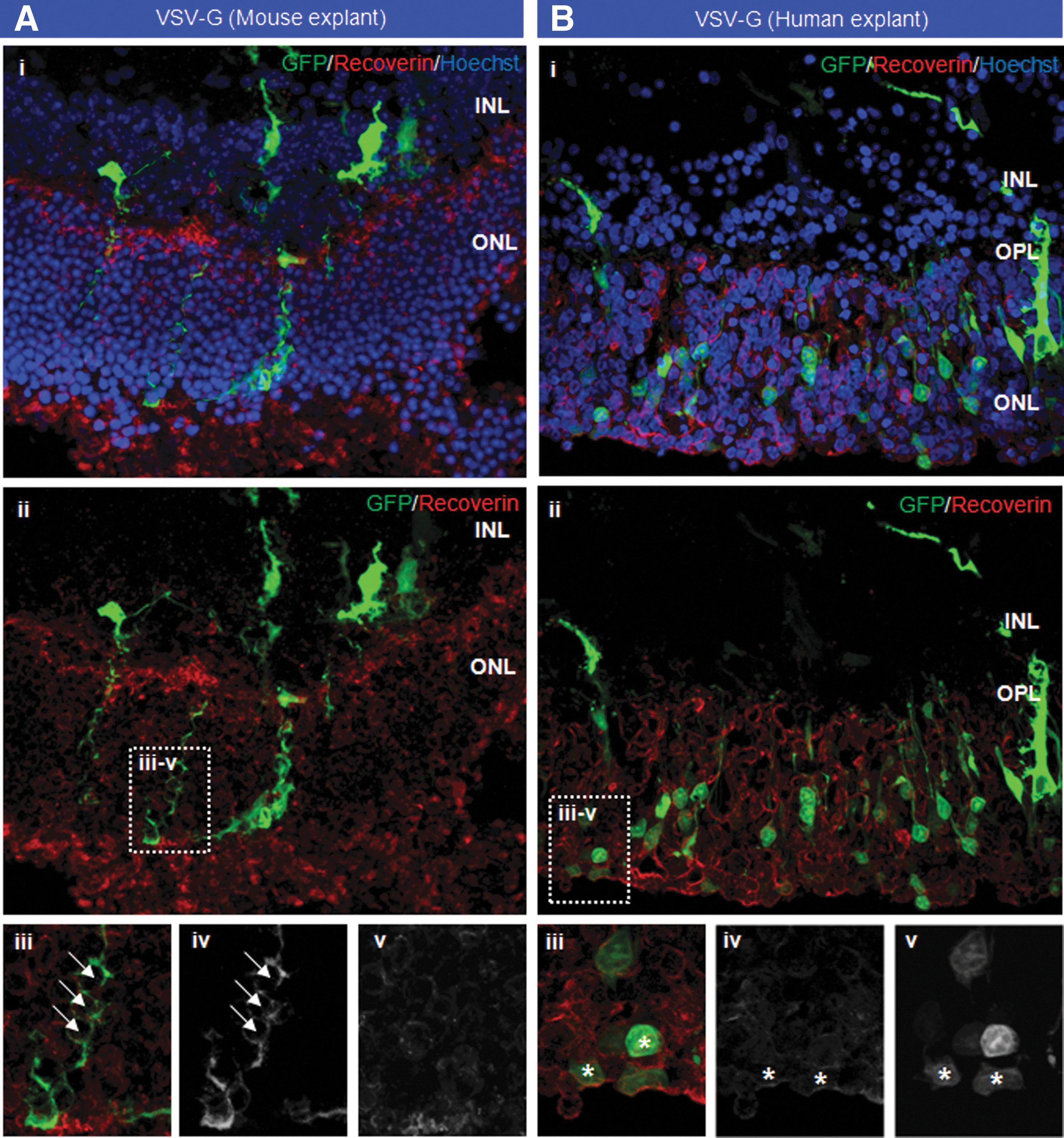
Transduction of mouse retina explanted from C57BL/6 mice
We investigated whether a similar lack of tropism was observed in human retina explanted from patients undergoing retinectomy surgery to treat complex retinal detachment, where the retina in such patients is often diseased and the outer limiting membrane compromised. One day postexplantation, retinal samples taken from a single patient with a retinal detachment were transduced ex vivo by the addition of 2.6×108 TU VSV-G diluted in 700 μl culture medium. Imaging of the retinal explants in culture showed EGFP expression by D3, which increased in intensity until D14 (data not shown), at which time the tissue was fixed. Examination of the retinal tissue while still in culture after delivery of VSV-G vector resulted in a punctuate pattern of EGFP expression visible in culture only when viewed from the outer retinal aspect (Supplementary Fig. S1D). That the observed dot-like pattern was not clearly visible from the inner retinal aspect indicates that the transduced cells were located in the ONL. Histology showed some transduction of cells throughout the ONL (Fig. 5B, i and ii) that co-localized with recoverin (Fig. 5, iii–v), confirming transduction of photoreceptors. The absence of transduction in murine tissue indicates that transduction may not be substantially limited by the presence of physical barriers. Indeed, that limited transduction was observed in retina explanted from a human patient indicates that transduction may be to some extent dependent on species.
Discussion
Herein, we evaluated the ocular tropism of a lentivirus vector pseudotyped with VEEV-G, the neurotropic nature of which we hypothesized would increase levels of photoreceptor transduction. To determine the short-term transduction and safety profile of VEEV-G-pseudotyped vector, tropism and function were assessed through in vivo fluorescence imaging, immunohistochemistry, and ERG compared with concurrently manufactured VSV-G vector of equivalent titer.
In vivo fluorescence imaging revealed similar expression profiles of VEEV-G and VSV-G vectors, with both demonstrating robust EGFP expression within 2 days of transduction. High-magnification imaging (30° lens) demonstrated a cobblestone pattern of fluorescence, indicating transduction of the RPE, a monolayer of hexagonally arrange low columnar or cuboidal epithelial cells underlying the neural retina. Areas of reduced fluorescence were observed on AF imaging at D21 in both VEEV-G- and VSV-G-injected eyes that corresponded with increased near-infrared reflectance, which clinically is commonly associated with RPE atrophy. No evidence of RPE degeneration was observed on histological examination of VEEV-G- or VSV-G-injected eyes; however, expression of RPE65 was frequently observed to be reduced or absent in areas where EGFP expression was highest. Absence of RPE65 staining may indicate increased RPE cell stress as a result of transduction, where RPE cells are known to de-differentiate under stress conditions leading to downregulation of cell-specific genes (Alizadeh et al., 2001; Alge et al., 2003). The downregulation of RPE-specific genes after transduction may have implications for the spatial control of gene expression to limit ectopic gene expression in a clinical environment, where this is typically achieved through use of cell-specific promoters, which are unlikely to be normally regulated in de-differentiated/stressed cells.
No photoreceptor transduction was observed with either vector pseudotype, yet substantial ONL thinning was noted in VEEV-G-injected eyes within the area of injection. Widespread cell death was observed in the ONL with three separately manufactured batches of VEEV-G vector, but not with concurrently produced VSV-G vectors, indicating a pseudotype-specific effect. While this finding strongly contraindicates the use of VEEV-G-pseudotyped vector for gene delivery to photoreceptors, it highlights the requirement for novel vector technologies to be rigorously evaluated in a laboratory environment before clinical application, particularly with respect to toxicity.
Functional assessment of lentivirus-injected eyes by ERG revealed the near-ablation of electrophysiological function in VEEV-G-injected eyes in both dark- and light-adapted conditions, indicating a substantial toxic effect. This magnitude of the functional deficit appears more severe than was expected because of the widespread ONL thinning observed on histology. This may be caused by a critical compromise of RPE function leading to a lack of physiological support for photoreceptors and/or ablation of the visual cycle. A significant reduction in dark-adapted a-wave amplitude was observed in VSV-G-injected eyes at D21, indicating impaired rod photoreceptor function after subretinal administration. A reduction in a-wave amplitude was also observed in TSSM-injected eyes relative to paired uninjected controls, likely as a result of retinal detachment during surgical intervention. However, the a-wave reduction was >2-fold greater in VSV-G-injected eyes than that was observed in TSSM-injected eyes compared with paired uninjected controls, strongly suggesting that VSV-G administration has a greater effect on photoreceptor function. The scotopic c-wave, a combined measure of photoreceptor–RPE function, was also reduced in TSSM-injected eyes, likely because of the transient detachment of the photoreceptors from the RPE; however, the c-wave was reduced to a significantly greater extent in eyes receiving VSV-G. The ERG a- and c-waves are closely associated, where the a-wave reflects a suppression of the photoreceptor “dark current,” which is relative to the level of phototransduction pathway activation in the photoreceptors, and the resulting depletion of extracellular potassium triggers the generation of the c-wave from the RPE cells. Consequently, dysfunction of either cell type typically leads to a reduction in the amplitude of both waveforms. However, the observed reduction in RPE65 staining in VSV-G-injected areas, as well as the absence of noticeable photoreceptor loss, would indicate that reduced ERG amplitudes might arise from RPE dysfunction. Flicker ERGs demonstrated significantly greater reduction in VSV-G-injected eyes than in TSSM-injected eyes (both compared with paired controls). This indicates that cone function (like that of rods) is affected to a greater extent by subretinal injection of VSV-G than by retinal detachment with buffer alone. Past studies have demonstrated that subretinal injection of VSV-G-pseudotyped vectors results in a greater reduction in a- and b-wave ERG response than buffer-only injection, and that amplitudes generally recover to normal levels within several months (Ikeda et al., 2003, 2009). Our findings indicate that the c-wave is similarly affected by subretinal injection. A recent study conducted by a commercial group concluded that the mean amplitude of “certain scotopic” flash intensities was reduced at 81 and 174 days postsubretinal injection of VSV-G-pseudotyped EIAV vector in rabbits (Binley et al., 2013). Unfortunately, Binley et al. (2013) did not specify which ERG components were affected in the injected rabbits, and did not present ERG data relating to concurrently injected primates, making the long-term assessment of safety from this study difficult. Consequently, while it is clear from the data presented herein that VEEV-G-pseudotyped vectors exhibit significant toxic effects in the retina, it remains to be addressed whether VSV-G-pseudotyped vectors are safe and efficacious for use in a clinical environment.
Given the lack of observed photoreceptor transduction in vivo with VSV-G-pseudotyped vector, we aimed to examine two questions: first, whether the presence of physical barriers prevents the transduction of photoreceptors in the murine retina, and, second, whether the tropism of VSV-G is different in the human retina compared with the mouse. Using a modified organotypic culture system, we maintained retina explanted from C57BL/6 mice, in addition to retina biopsied from a human patient undergoing retinectomy as part of surgery for a complex retinal detachment. Retina was maintained over a period of 14 days, during which time little evidence was observed for photoreceptor transduction in the mouse retina explants. This finding indicates that the presence of physical barriers, such as the outer limiting membrane and outer segment discs, which are largely compromised or absent in explanted tissue, is not the sole factor limiting photoreceptor transduction. Importantly, photoreceptor transduction was observed in human retina explants; while limited photoreceptor transduction has been previously observed in animal models (Nicoud et al., 2007), to the authors' knowledge this represents the first time that a lentivirus vector has been demonstrated successfully to transduce human photoreceptors. That transduction was observed only in human retinal explants implies that the tropism of lentivirus vectors may differ between species, as observed with rAAV vectors (Calame et al., 2011; Yin et al., 2011; Boye et al., 2012), possibly because of interspecies differences in the expression of cellular receptors/co-receptors. Photoreceptor transduction in this instance was observed only in a small number of cells, indicating that while lentivirus can transduce human photoreceptors the process is inefficient. On the basis of our current observations, it appears unlikely that lentivirus vectors could be used to achieve the level of transduction required for therapeutic correction of a retinal disease, such as Stargardt's, which would require high-level gene expression in the majority of photoreceptors.
In contrast, both lentivirus vectors were well tolerated in the anterior chamber and led to widespread transduction of the corneal endothelium and the trabecular meshwork. The extent of transduction using VEEV-G was comparable to that of VSV-G, indicating that either might be used successfully to deliver large or multiple gene expression cassettes to these tissues, potentially for the treatment of glaucoma and corneal endothelial dystrophies. Interestingly, AF imaging revealed that transduction of the corneal endothelium is not uniform after intracameral delivery. There are two factors that likely have a role in determining the extent of endothelial cell transduction. The first is the dynamics of aqueous flow within the anterior chamber, which in humans is driven by a number of complex factors resulting in suspended particles being driven toward the corneal endothelium, in addition to the trabecular meshwork and iris (Heys and Barocas, 2002). The second is the injection itself, whereby the angle, force, and volume of the vector suspension injected will likely determine the distribution of the virus. Together, this indicates that VSV-G and VEEV-G lentivirus vectors can transduce the corneal endothelium and trabecular meshwork, but that uniform transduction of the corneal endothelium, as would likely be required for the treatment of endothelial dystrophies, may be difficult to achieve.
In conclusion, we have demonstrated that VEEV-G vector has significant toxic effects when administered subretinally leading to photoreceptor degeneration and loss of retinal function, contraindicating its use as a photoreceptor gene delivery vector, but that VEEV-G is well tolerated in the anterior chamber. While photoreceptors were not transduced by VSV-G-pseudotyped lentivirus vector in vivo, limited transduction of photoreceptors was observed in explanted human tissue, suggesting that species differences may influence tropism, as reported with AAV vectors. However, photoreceptor transduction was not wide spread, and in vivo administration of VSV-G resulted in greater short-term reduction of electrophysiological function that could be accounted for by surgical detachment alone. The reduction in photopic c-wave amplitude, coupled with reduced histological observation of RPE65, may indicate RPE dysfunction. Together, these findings suggest that long-term assessment of RPE function after subretinal lentivirus administration would be necessary before clinical applications being considered, and that given the observed species-dependent differences in transduction, such safety assessments should be independently conducted on species more closely related to humans.
Footnotes
Acknowledgments
We would like to thank Dr. Rosalie Crouch (Medical University of South Carolina) and Dr. Steven Hughes (University of Oxford) for providing the RPE65 antibody and technical assistance, and Dr. Robert A. Davey (University of Texas) for providing the 3908 VEEV-G plasmid. We would also like to thank Prof. Alistair Fitt (Oxford Brookes University) for useful discussions on the aqueous flow dynamics of mice.
This research was funded by Fight for Sight, the Wellcome Trust, the Health Foundation, the Medical Research Council, the Royal College of Surgeons of Edinburgh, the Oxford Stem Cell Institute, the NIHR Ophthalmology and Oxford Biomedical Research Centres, ProRetina, and the European Research Council.
Author Disclosure Statement
The authors declare no conflict of interest or commercial associations.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.