
Abstract
In the context of motoneuron diseases, gene delivery as an experimental or therapeutic approach is hindered by the challenge to specifically target cell populations that are widely distributed along the spinal cord. Further complicating the task, transgenes often need to be delivered to motoneurons and/or glial cells to address the non-cell-autonomous mechanisms involved in disease pathogenesis. Intracerebroventricular (ICV) injection of recombinant adeno-associated viruses (AAVs) in newborn mice allows distributing viral vectors throughout the central nervous system while limiting undesired transduction of peripheral organs. Here, we show that by combining the appropriate set of AAV serotype and promoter, specific transgene expression can be achieved in either motoneurons or astrocytes along the whole mouse spinal cord. ICV injection of recombinant AAV6 with the cytomegalovirus (cmv) promoter preferentially targets motoneurons, whereas AAV9 particles combined with the astrocyte-specific gfaABC1D promoter lead to significant transgene expression selectively targeted to astrocytes. Importantly, ICV coinjection of both AAV6-cmv and AAV9-gfaABC1D results in segregated expression of two different transgenes in motoneurons and astrocytes, respectively. Relevance of viral vector delivery via the cerebrospinal fluid was further investigated in young nonhuman primates. Intracisternal injection of recombinant AAV6-cmv led to robust cervical transduction of motoneurons, highlighting the potential of this approach for gene therapy and modeling of motoneuron diseases.
Introduction
A
Viral vector technology is an effective tool for gene delivery to the spinal cord, as it allows targeting specific cell populations that are critical for disease pathogenesis. Viral vectors can be used to investigate the role of any protein linked to ALS pathogenesis by either overexpressing or silencing its expression. Finally, recombinant viruses can be used as a delivery system for therapeutic proteins or nucleic acids. Indeed, gene therapy is an attractive approach against some hereditary forms of ALS, but also against other monogenic motoneuron disorders such as spinal muscular atrophy (SMA) (Passini and Cheng, 2011; Federici and Boulis, 2012). As a proof of principle, the motor phenotype of a severe murine SMA model could be rescued after viral vector-induced overexpression of the SMN protein in motoneurons (Passini and Cheng, 2011).
Adeno-associated viruses (AAVs) are attractive vehicles for gene transfer as they can readily infect a wide range of tissues, including nondividing cells within the central nervous system (CNS). Different parameters can be modulated to gain specificity, including the serotype and the promoter used as well as the route of delivery (Wu et al., 2006; Mingozzi and High, 2011). Several AAV serotypes, in combination with different injection paradigms, have been tested for widespread motoneuron and astrocyte targeting. In particular, intravenous injection of recombinant AAV9 led to robust motoneuron transduction in newborn mice (Duque et al., 2009; Foust et al., 2009; Miyake et al., 2011; Zhang et al., 2011). This pattern appeared to shift toward preferential astrocyte transduction when recombinant AAV9 was injected in adult mice (Foust et al., 2009). In nonhuman primates, intravenous injection of recombinant AAV9 led to transgene expression in both neurons and astrocytes (Bevan et al., 2011; Gray et al., 2011).
Gene transfer efficiency via peripheral routes might be restricted by the high prevalence of pre-existing antibodies against most AAV serotypes in humans. Another limiting factor resides in the passage of recombinant viral particles across the blood–brain barrier after intravenous gene delivery. Hence, high viral loads are needed to achieve broad transduction in the CNS, which might lead to undesired effects after widespread transduction of peripheral organs. Injection of recombinant AAVs in the cerebrospinal fluid (CSF) has proven to be an interesting alternative to intravenous delivery for CNS applications. Indeed, direct injection in the CSF bypasses the blood–brain barrier and therefore reduces the injected vector load. Both AAV6 and AAV9 were shown to preferentially infect motoneurons after intrathecal injection in adult mice and newborn pigs (Bevan et al., 2011; Federici et al., 2011; Snyder et al., 2011). However, a study performed in nonhuman primates demonstrated high transduction of astrocytes with AAV9 in examined brain areas (Samaranch et al., 2012).
In the present study, we aim to establish AAV vectors to selectively express a transgene in a high proportion of either motoneurons or astrocytes in the mouse spinal cord. AAV vectors are administered intracerebroventricularly in mouse neonates to maximize transgene spread across the CNS. We demonstrate that AAV6 leads to effective and preferential transduction of motoneurons across the entire spinal cord. To selectively target astrocytes, AAV9 vectors are used in combination with a minimal GFAP promoter to restrict expression of the transgene in glial cells. In addition, we show that injection of a recombinant AAV6 vector in the cisterna magna of nonhuman adult primates leads to motoneuronal transduction consistent with the transduction pattern found in mouse neonates.
Materials and Methods
Preparation of viral vectors
pAAV shuttle plasmids were engineered by cloning cytomegalovirus (cmv):GFP, phosphoglycerate kinase (pgk):GFP, chicken β-actin hybrid (cba):GFP, and cmv:RFP cassettes in the pAAV-MCS plasmid (Stratagene). The minimal GFAP promoter gfaABC1D (kindly provided by Dr. M. Brenner, Department of Neurobiology, University of Alabama) was cloned in pAAV-MCS modified with the bovine growth hormone polyadenylation signal. Self-complementary pAAV9-gfaABC1Ds were designed by removing the terminal resolution site from the 5′ ITR of the pAAV-MCS plasmid as described previously (McCarty et al., 2003; Wang et al., 2003). All viruses used in this experiment are based on a recombinant genome containing the AAV2 ITRs and pseudotyped with an AAV capsid derived from either serotype 6 or 9.
For AAV6 and AAV9 virus production, shuttle plasmids were cotransfected with helper pDP6 or pDF9 plasmids in HEK293-AAV cells (Agilent Technologies). Transfected cells were lysed 48 hr later by freeze–thaw cycles. Recovered viral particles were sequentially purified on iodixanol and heparin affinity columns. Viral genomic copies (VG) were determined by TaqMan quantitative polymerase chain reaction (qPCR) using primers against the viral β-globin intron. Infectivity titers were also measured for AAV6 vectors by determining the amount of transducing units (TU) at 48 hr after injection of HEK293T cells, as described previously (Dusonchet et al., 2009).
Animals and vector administration
C57BL6/J mice (Charles River Laboratories) were housed in a 12 hr light/dark cycle, with ad libitum access to food and water. All animal procedures were conducted in accordance with Swiss legislation and the European Community Council directive (86/609/EEC) for the care and use of laboratory animals. Newborn P2 C57BL/6 pups were anesthetized through intraperitoneal injection of 4 μl of a mix containing medetomidine (0.67 mg/ml; Dorbene; Dr. Graeub AG) and midazolam (1.7 mg/ml; Dormicum; Roche). Anesthetized mice were tape-fixed, and intraventricular injection of 3 μl virus was performed using a 29G insulin syringe; 0.1% Fast Green FCF (Sigma-Aldrich) was added to the virus suspension to visualize spreading of the injected preparation to the ventricles. Pups were returned to their cage after reversal of the anesthesia through subcutaneous injection of 5 μl Atipamezol 1 mg/ml (Alzane; Dr. Graeub AG). Mice were euthanized 21 days postinjection by intracardial perfusion of phosphate buffered saline (PBS) followed by 4% paraformaldehyde (PFA). Spinal cord and brain were recovered and postfixed for 2 hr in 4% PFA.
Three young adult male African green monkeys (Chlorocebus sabaeus, 1.12–2.35 kg) were used for intracisternal injection in nonhuman primates. The research protocol was in accordance with the U.S. Public Health Service Guide for Humane Care and Use of Laboratory Animals and was approved by the Institutional Animal Care and Use Committee. Animals were maintained in the St. Kitts Biomedical Research Foundation colony (St. Kitts, West Indies). Animals were anesthetized with ketamine (10 mg/kg) and injected into the cisterna magna with 7.5E8 TU of AAV6-cmv:GFP diluted in 250 μl saline using a 25G needle. Monkeys were euthanized 1 month later by overdose with intravenous pentobarbital and transcardially perfused with PBS containing sodium heparin followed by 4% PFA in PBS. The spinal cord was dissected and stored in 4% PFA in PBS for 24 hr and then moved to 30% sucrose for shipping and storage.
Histological analysis
Brains and spinal cords were soaked in a 30% sucrose solution to prevent freezing artifacts. Tissues were then embedded in Cryomatrix (Thermo Scientific) and frozen on dry ice. Spinal cords were cut free-floating at 25 μm, whereas brains were cut at 30 μm and mounted on superfrost slides. Free-floating spinal cord sections were conserved at 4°C in PBS-azide-filled 48-well plates. Brain sections were stored at −20°C.
Mouse spinal cord and brain sections were either analyzed for native GFP expression or labeled with 1/500 rabbit anti-GFP (Invitrogen, Molecular Probes), 1/500 mouse anti-GFP (Invitrogen, Molecular Probes), 1/500 rabbit anti-RFP (Abcam), 1/100 goat anti-ChAT (Chemicon Millipore), or 1/500 rabbit anti-GFAP (DakoCytomation) primary antibodies. Immunostaining was revealed using 1/500 donkey anti-goat Cy3 (Jackson ImmunoResearch), 1/500 donkey anti-rabbit AF488 (Invitrogen, Molecular Probes), 1/500 goat anti-rabbit Cy3 (Jackson ImmunoResearch), or 1/500 goat anti-mouse AF488 (Invitrogen, Molecular Probes) secondary antibodies. Sections were first incubated for 1 hr in blocking solution, containing either 5% bovine serum albumin, 3% goat or donkey serum. Sections were then incubated with primary antibodies diluted in the blocking solution plus 0.15% Triton X-100 for 24 hr at 4°C. Sections were rinsed in PBS three times and incubated with the secondary antibody for 2 hr at room temperature. Tissues were rinsed three times in PBS and mounted with Mowiol (Sigma-Aldrich).
To analyze the transduction pattern in monkeys, the length of the spinal cord was sectioned at 40 μm. Transgene expression was analyzed using an antibody against GFP (Invitrogen, Molecular Probes) followed by amplification with the 3,3′-diaminobenzidine (DAB)-peroxidase substrate kit (Vector Labs) as per manufacturer's instructions.
Quantification of transgene-expressing cells
For quantification of GFP-expressing motoneurons, serial 25 μm sections separated by 500 μm were immunostained with anti-GFP and anti-ChAT antibodies. Sections were separated in cervical, thoracic, lumbar, and sacral categories and assessed using an Olympus AX70 microscope (Olympus Corporation). The total number of ChAT-positive motoneurons was counted under a 20× objective using a Cy3 filter, whereas GFP coexpressing motoneurons were visualized under a fluorescein isothiocyanate filter. An average of 660±180 ChAT-positive motoneurons were counted in the cervical spinal cord per animal. An average of 510±150, 490±120, and 110±50 ChAT-expressing motoneurons were counted in the thoracic, lumbar, and sacral spinal cord, respectively.
Percentage of GFP and GFAP coexpressing astrocytes was evaluated in the ventral horn of 25 μm spinal cord sections. Z-stacks were taken using a Zeiss LSM 700 inverted confocal microscope (Carl Zeiss) endowed with a 64× objective. Lasers of 488 and 555 nm were used to obtain z-stacks of the GFP and GFAP signals, respectively. In order to identify astrocytes, 3D reconstructions were made using ImageJ. Ten GFAP-positive astrocytes were visualized on average per field of view. An average of 120, 130, 130, and 90 astrocytes were counted in the cervical, thoracic, lumbar, and sacral spinal cord of each animal, respectively. Merging of 3D reconstructions obtained with the 488 or 555 nm lasers allowed identification and counting of astrocytes positive for GFAP and GFP.
Measurements of native GFP intensity were made on focal plane pictures obtained with an LSM 700 inverted confocal microscope (Carl Zeiss) under a 20× objective and 488 nm laser beam. The mean intensity values for individual GFP-expressing motoneurons were obtained using MetaMorph (Universal Imaging). Background mean intensity was subtracted to the values obtained for each GFP-expressing motoneuron.
In monkey sections, the percentage of total motoneurons transduced was calculated by comparing GFP-stained spinal cord slices with serial sections stained by the Nissl method. Motoneurons were identified as the large Nissl-stained cell bodies in the ventral horn.
Determination of transgene copy numbers by real-time PCR
Intracerebroventricular (ICV) injections were performed in newborn mice as described above. Animals were sacrificed by injection of sodium pentobarbital and perfused with PBS. Lumbar, thoracic and cervical spinal cord, brainstem, cerebellum, cerebral cortex, heart, lung, liver, and kidney tissues were recovered and fresh-frozen on dry ice. Tissues were kept at −80°C awaiting further handling.
Tissues were defrosted and cut into small pieces, which were resuspended in PBS containing 4 mg/ml RNAse A. NucleoSpin tissue kit (Macherey-Nagel) was used for extraction of genomic DNA.
Vector DNA was amplified using primers against the human β-globin intron (fw primer, 5′ CGT GCC AAG AGT GAC GTA AG 3′; rv primer, 5′ TGG TGC AAA GAG GCA TGA TA 3′; probe, 5′ FAM-TTG CCC TGA AAG AAA GAG ATT AGG GAA-BHQ1 3′). Mouse β-tubulin was used as control for genomic DNA (fw primer, 5′-GCC AGA GTG GTG CAG GAA ATA-3′; rv primer, 5′-TCA CCA CGT CCA GGA CAG AGT-3′; probe, 5′-FAM-CTG GGC AAA GGG CCA CTA CAC AGA GG-BHQ1-3′). The Rotor Gene Probe PCR kit (Qiagen GmbH) was used for amplification on a Rotor-Gene cycler (Corbett Life Science). Cycling parameters were as follows: 95°C for 180 sec, and then 40 cycles at 95°C for 3 sec and 60°C for 10 sec.
Statistical analysis
Data are expressed as mean±standard error of the mean. Statistical significance was analyzed either with one-way ANOVA followed by Newman–Keuls post-hoc test or with Student's t-test, as indicated in the figure legends. The alpha level of significance was set at p=0.05.
Results
ICV injection of recombinant AAV6 leads to widespread infection of spinal motoneurons
In the first part of this study, we aimed to maximize transduction of motoneurons throughout the spinal cord. AAV6 virus can be very efficiently delivered to motoneurons through retrograde transport after intramuscular injection (Towne et al., 2010, 2011). Because ventral roots are in close contact with CSF, we hypothesized that ICV delivery of AAV6 would allow viral particles to infect motoneurons at the ventral roots. Recombinant particles would then be transported retrogradely to the cell soma. This hypothesis was hence tested by ICV injection of 4E10 VG of AAV6 encoding GFP under the cmv immediate-early promoter in five P2 mouse pups. Injected animals were euthanized 3 weeks after injection and assessed for GFP expression.
In the brain, GFP expression was observed in neurons and astrocytes in the cortex, particularly around the ventricles and close to the needle track. We also found infection of neurons in the granular layer of the hippocampus (Fig. 1a and b). In regions of the CNS that are more distant to the injection site, the AAV6 vector mainly transduced long projection neuronal populations. For instance, we found consistent transduction of Purkinje neurons in the cerebellum (Fig. 1c).

Widespread transduction of motoneurons after ICV injection of AAV6-cmv:GFP.
Expression of GFP was predominantly observed in motoneurons throughout the spinal cord gray matter. Indeed, GFP-positive astrocytes were only very rarely observed. GFP expression was also present in the posterior columns, suggesting transduction of dorsal root ganglia (Fig. 1d). The amount of transduced motoneurons varied depending on spinal cord level with more cells transduced in the cervical, lumbar, and sacral regions compared with the thoracic level.
Recombinant AAV9 vectors have been extensively studied for motoneuron transduction following different routes of delivery. To compare transduction efficiency of AAV6 and AAV9, 2E11 VG of AAV6-cmv:GFP or AAV9-cmv:GFP were injected in P2 pups (n=5 per group). Both vectors transduced motoneurons, although transduction efficiency was 4–10 times higher for the AAV6 vector (Supplementary Fig. S1; Supplementary Data are available online at
Neonatal ICV injection using an AAV6-cmv vector therefore leads to efficient infection of neuronal subpopulations in the CNS, with a selective motoneuronal transduction pattern in the spinal cord gray matter. Next, we sought to optimize the promoter used for transgene expression and AAV6 vector dose in order to improve motoneuron targeting in the spinal cord.
The constitutive cmv promoter induces efficient transgene expression in spinal cord motoneurons
Because the association of AAV6 serotype and ICV injection already provides a high specificity for motoneurons in the spinal cord, we went on testing three strong constitutive promoters in order to optimize GFP expression in these cells. To compare promoter efficacy in similar conditions, we determined the infectious titer for each AAV6 vector preparation, by measuring the amount of TU at 48 hr postinfection in HEK293T cells. For the AAV6-cmv:GFP vector, 670 VG corresponded to 1 TU detected in HEK293T cells. A total of 6E7 TU of recombinant AAV6 particles expressing GFP under either the cmv (n=5), pgk (n=5), or cba (n=5) promoters were injected intracerebroventricularly in P2 mouse pups. Tissues were processed for immunodetection of GFP expression 3 weeks postinjection. All three promoters led to expression of GFP in motoneurons throughout the spinal cord (Fig. 2a). Specificity for motoneurons was highest for cmv and pgk promoters, whereas some astrocytes were also shown to express GFP under the cba promoter (Fig. 2a).
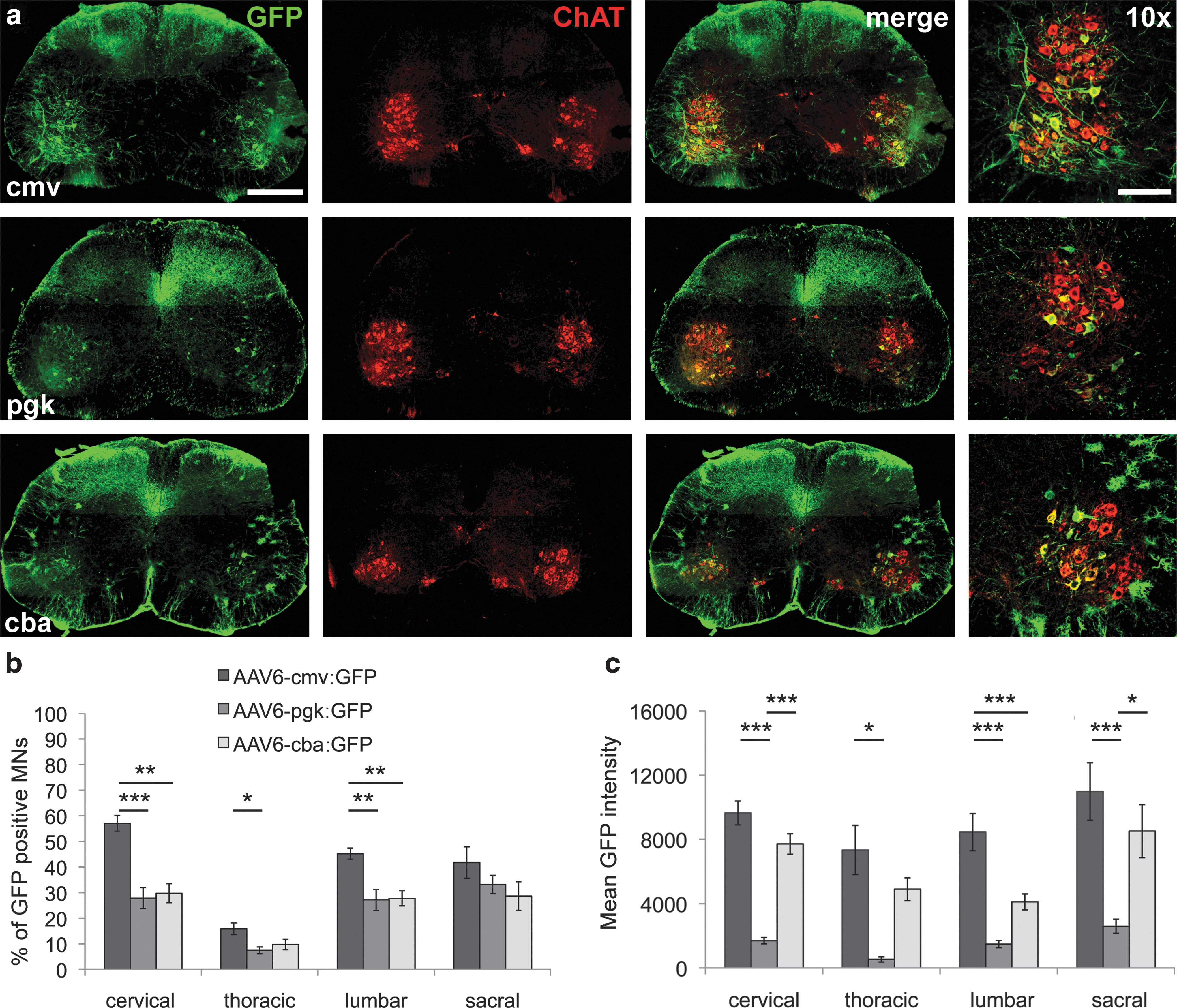
Comparison of cmv, pgk, and cba constitutive promoters for transgene expression in motoneurons.
At the same vector dose, efficiency in terms of number of GFP-expressing motoneurons was significantly higher for the cmv promoter than for both cba and pgk promoters. To establish the percentage of GFP-positive motoneurons, neurons depicting both ChAT and GFP immunoreactivity were counted in the ventral horns at different levels of the spinal cord. Up to 57%±3% of cervical motoneurons were expressing GFP under the cmv promoter, whereas pgk and cba promoters led to expression of GFP in, respectively, 28%±4% and 30%±4% of cervical motoneurons (Fig. 2b). The cmv promoter also led to significantly more GFP-positive motoneurons in the lumbar spinal cord with an average of 45%±2% infected cells against 27%±4% for the pgk promoter and 28%±3% for the cba promoter (Fig. 2b).
The level of GFP expression per neuron was also assessed. Mean native GFP intensity was determined on confocal images taken from spinal cord sections. Motoneurons were identified in the ventral horn based on their morphology and size, they were then individually outlined, and mean GFP intensity was determined using MetaMorph software. The cmv promoter was again the most efficient, leading to higher levels of GFP expression per cell at all examined levels of the spinal cord (Fig. 2c). Whereas the number of GFP-expressing motoneurons was found to be similar for both pgk and cba promoters, the pgk promoter appeared to induce much lower levels of GFP expression per cell (Fig. 2b and c).
AAV6-cmv:GFP leads to dose-dependent expression of GFP in motoneurons
Having established the most effective promoter system, we went on determining vector concentrations needed to obtain high percentage of motoneuron infection. Hence, 1.3E10, 4E10, or 1.2E11 VG of recombinant AAV6-cmv:GFP were injected intracerebroventricularly in five P2 pups per group. A significant dose-dependent effect could be observed, as 1.2E11 VG led to a clearly higher number of infected motoneurons as well as higher level of GFP expression per cell (Fig. 3a).
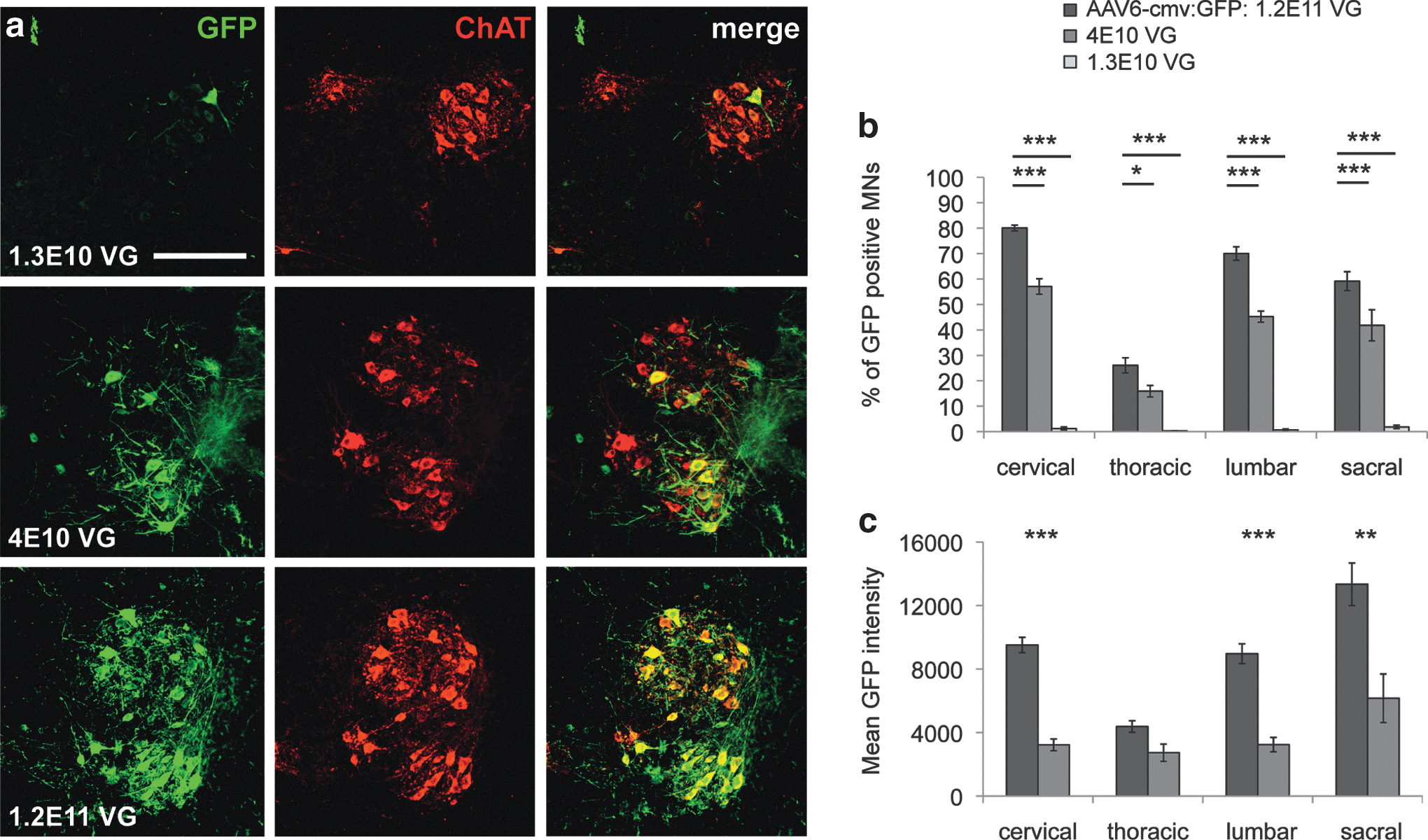
Dose-dependent transgene delivery in motoneurons after ICV injection of AAV6-cmv.
Up to 80%±1% of cervical and 70%±3% of lumbar motoneurons were shown to express GFP under this condition. These percentages reached 57%±3% and 45%±2%, respectively, at one-third of the dose, whereas they drastically dropped when only 1.3E10 VG were injected (Fig. 3b). Except for the thoracic spinal cord, significantly higher GFP intensity was measured at the cervical, lumbar, and sacral levels of mice injected with the full dose of recombinant AAV6-cmv:GFP (Fig. 3c). Thus, it appears that ICV injection of the AAV6-cmv vector in newborn mice is an efficient way to specifically express a transgene in motoneurons in the spinal cord.
ICV injection of recombinant AAV9-gfaABC1D:GFP induces GFP expression specifically in astrocytes
Specific transduction of high numbers of spinal astrocytes is of interest for investigating and potentially treating non-cell-autonomous motoneuron diseases such as ALS. Indeed, it has been shown that astrocytes expressing mutated SOD1 actively participate in the disease process and influence disease progression. No astrocytes were shown to express GFP after ICV delivery of AAV6-cmv:GFP despite the strong ubiquitous cmv promoter. On the other hand, astrocyte transduction was observed when AAV9-cmv:GFP was injected intracerebroventricularly. Moreover, AAV9 has already been shown to efficiently infect astrocytes in mice, in particular after intravenous injection (Duque et al., 2009; Foust et al., 2009; Gray et al., 2011; Zhang et al., 2011). Hence, the AAV9 serotype was chosen to optimize targeting of spinal cord astrocytes. As AAV9 vectors can also transduce other cell types, including motoneurons, the gfaABC1D promoter was used to drive transgene expression in astroglial cells only (Lee et al., 2008). ICV injection of P2 mice with 2.3E11 (n=5) or 9.2E10 (n=5) VG of AAV9-gfaABC1D:GFP led to specific expression of GFP in a high number of astrocytes in the whole CNS 3 weeks postinjection.
We observed very consistent infection of astrocytes throughout the spinal cord (Fig. 4a and b). High specificity was achieved as neither motoneurons nor other cell types were found to express GFP. Moreover, intense astrocyte-specific expression of GFP was observed in the brain, indicating widespread diffusion of the virus in the whole parenchyma (Fig. 4c–f).
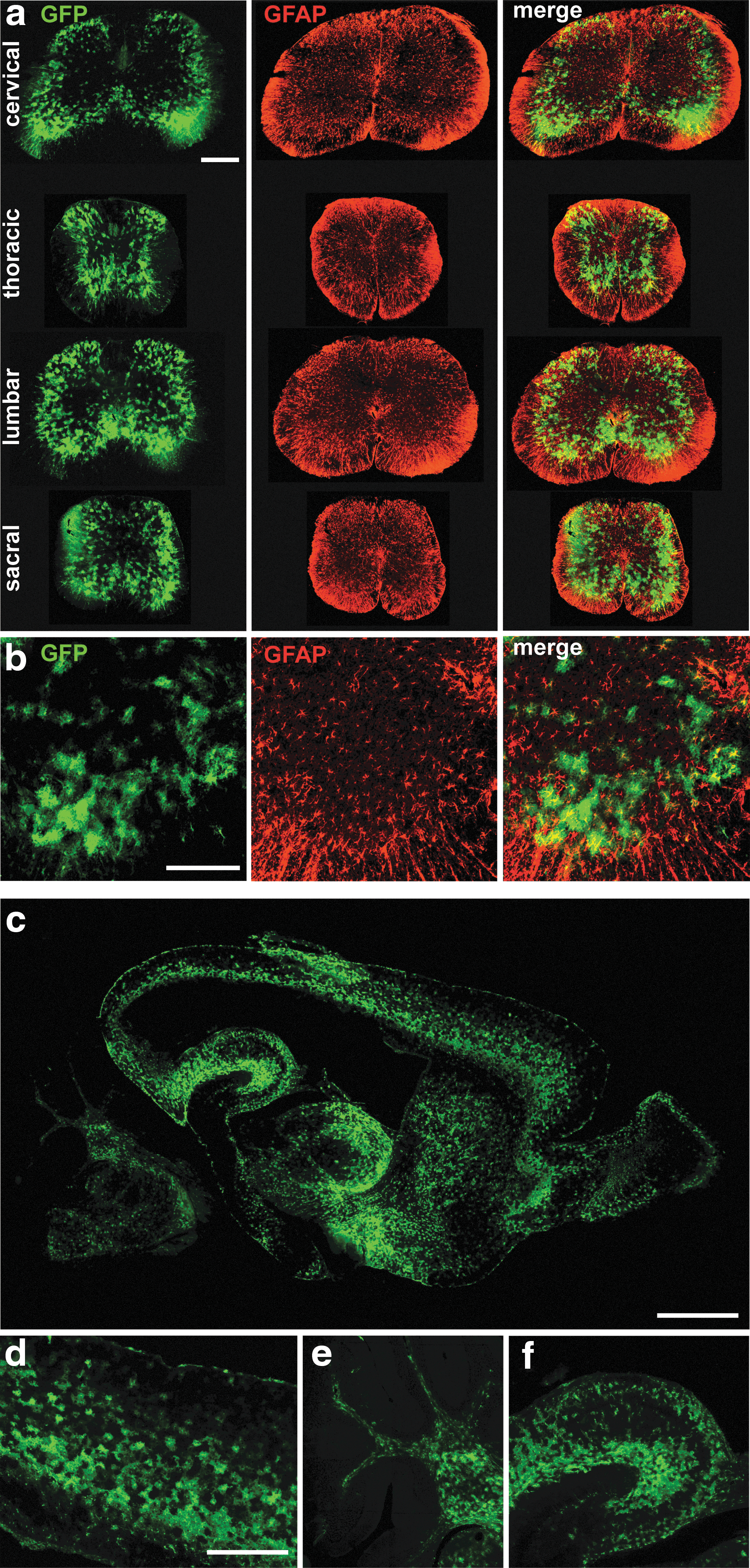
Widespread and specific transgene expression in astrocytes after ICV injection of AAV9-gfaABC1D:GFP.
The percentage of infected astrocytes in the ventral horn was determined using 3D reconstruction of confocal images of spinal cord sections colabeled against GFP and GFAP. Here again, the percentage of infected cells was dose dependent (Fig. 5a and b). About 49%±1% of ventral horn astrocytes were shown to express GFP at the highest viral dose tested, as compared with 29%±0.3% when 2.5-fold less virus was injected (Fig. 5d).

GFP expression in astrocytes increases with AAV9 dose and self-complementary vectors.
Self-complementary AAV9-gfaABC1D:GFP leads to higher infection efficiency than single-stranded vectors
Since AAV9 particles appear to diffuse through tissues to infect cells, only few infection events are expected to occur deep in the brain and spinal cord parenchyma. In these conditions, second-DNA-strand synthesis is a limiting factor for transgene expression after infection with AAV vectors. Therefore, self-complementary double-stranded vectors may increase expression efficiency in the situation of low infection frequency. For this reason, a self-complementary double-stranded version of the AAV9-gfaABC1D:GFP vector was produced. Five mice were injected with 9.2E10 VG-containing viral particles of single- or double-stranded vectors and euthanized 3 weeks postinjection. Animals injected with the self-complementary vector depicted significantly more GFP-expressing astrocytes than mice injected with the single-stranded version (Fig. 5c and e).
Hence, ICV injection of AAV9-gfaABC1D:GFP allows infection of high numbers of astrocytes throughout the spinal cord. Moreover, for the same vector dose, the use of double-stranded, self-complementary AAV9 vector significantly increases the number of astrocytes expressing the transgene.
Coinjection of AAV6-cmv and AAV9-gfaABC1D particles leads to a cell-specific pattern of transgene expression in the spinal cord
To assess the feasibility of specifically overexpressing two different proteins in motoneurons and astrocytes, we performed ICV coinjections of recombinant AAV6-cmv:RFP (3E10 VG) and AAV9-gfaABC1D:GFP (1.15E11 VG) in five mouse neonates. A cell type-specific pattern of transgene expression was found 3 weeks postinjection, with RFP expressed in motoneurons and GFP in astrocytes of the spinal cord (Fig. 6a and b). The amount of infected cells was in both cases congruent with the injected titers, meaning that no detrimental interaction between AAV6 and AAV9 viruses occurred in vivo. Coinjection of AAV6-cmv and AAV9-gfaABC1D vectors is therefore a powerful tool to specifically target motoneurons or astrocytes, respectively.
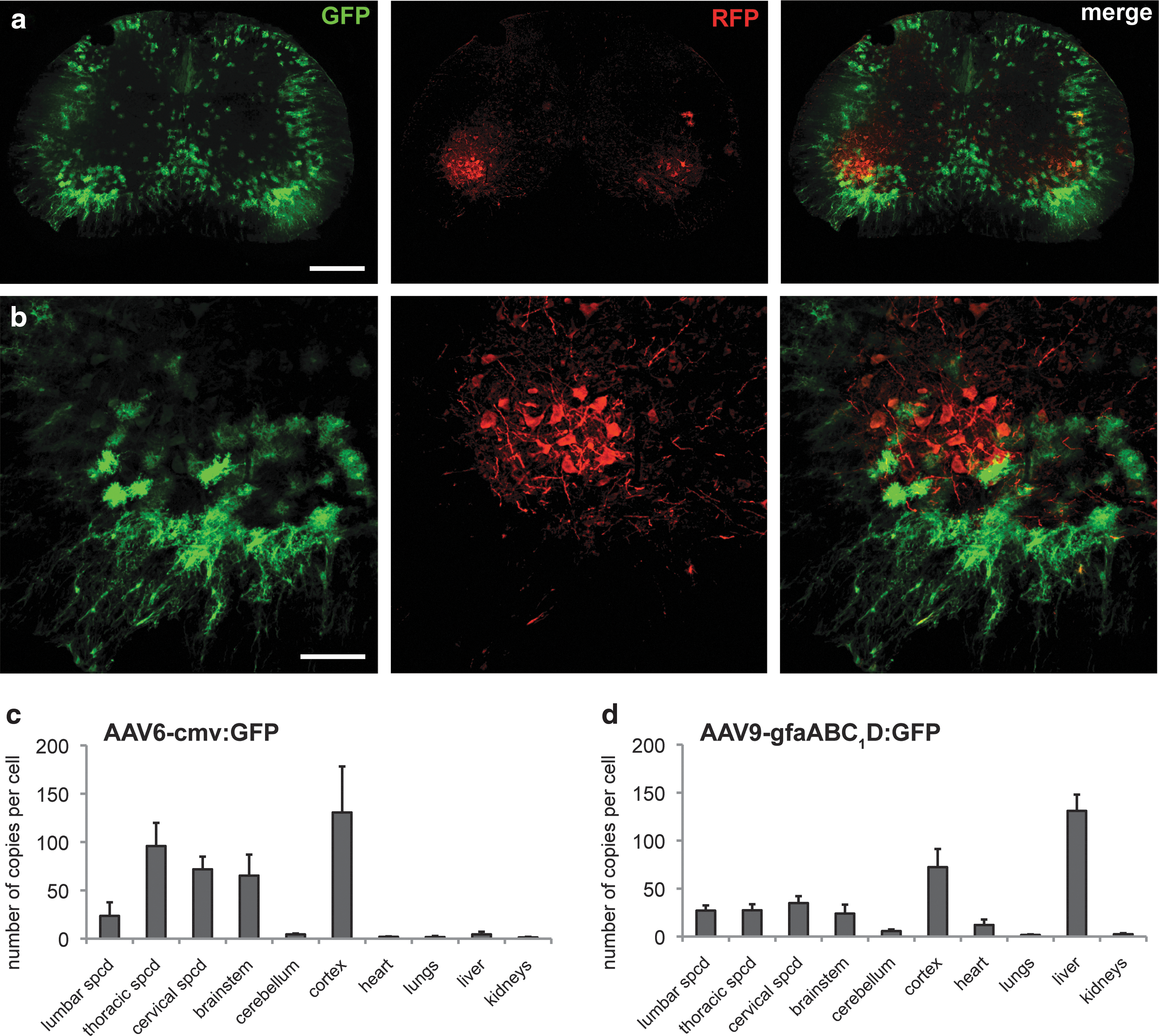
Coexpression of GFP in astrocytes and RFP in motoneurons after a single ICV injection of AAV6 and AAV9 vectors.
Spread of transgene copies is more restricted to the CNS with AAV6 than with AAV9 vectors
To determine the overall pattern of transgene delivery across the CNS and peripheral organs after ICV injection of AAV6 and AAV9 vectors, we determined by real-time qPCR the amount of transgene DNA copies present in individual tissues. AAV6-cmv:GFP (4E10 VG, n=3) and AAV9-gfaABC1D:GFP (9.2E10 VG, n=3) vectors were injected in separate groups of P2 mouse pups. Animals were euthanized 3 weeks postinjection, and total DNA was extracted from the CNS and peripheral tissues.
Interestingly, a different pattern of transgene distribution was observed for AAV6 and AAV9 vectors. AAV6 particles mostly remained in the CNS. For instance, up to 130 copies of viral DNA per cell were present in the cerebral cortex. In the periphery, only few copies (<4.5 copies per cell) were found in the heart, lungs, liver, and kidneys (Fig. 6c). In contrast, AAV9 showed strikingly high levels of transgene copies in the liver, with an average of 131±17 copies per cell. This indicates consequent leakage of AAV9 virus out of the CNS with a preferential liver tropism. This, however, did not preclude the presence of many AAV9 viral copies in the CNS (Fig. 6d). In conclusion, it appears that AAV6 particles remain mainly clustered in the CNS, whereas AAV9 vectors significantly diffuse outside of the CNS with favored liver tropism.
Intracisternal injection of AAV6-cmv leads to motoneuron transduction in nonhuman primates
We next sought to determine whether CSF delivery of AAV6 would similarly target motoneurons in the spinal cord of a nonhuman primate species. Three young male African green monkeys of St. Kitts origin (C. sabaeus) were injected in the cisterna magna with 5E11 VG of recombinant AAV6-cmv:GFP. Animals were euthanized 1 month later and tissues analyzed. Robust GFP expression was observed in the cervical spinal cord close to the injection site (Fig. 7a). An extensive array of GFP-positive fibers was observed in the deep lamina of the dorsal spinal cord likely corresponding to transduction of large neurons in the dorsal root ganglia. As expected from the rodent experiments, the only soma found to express GFP were located in the ventral horns of the spinal cord. Closer inspection revealed that these large-diameter cells had GFP-positive axonal process projecting into the ventral roots and were therefore putative motoneurons.
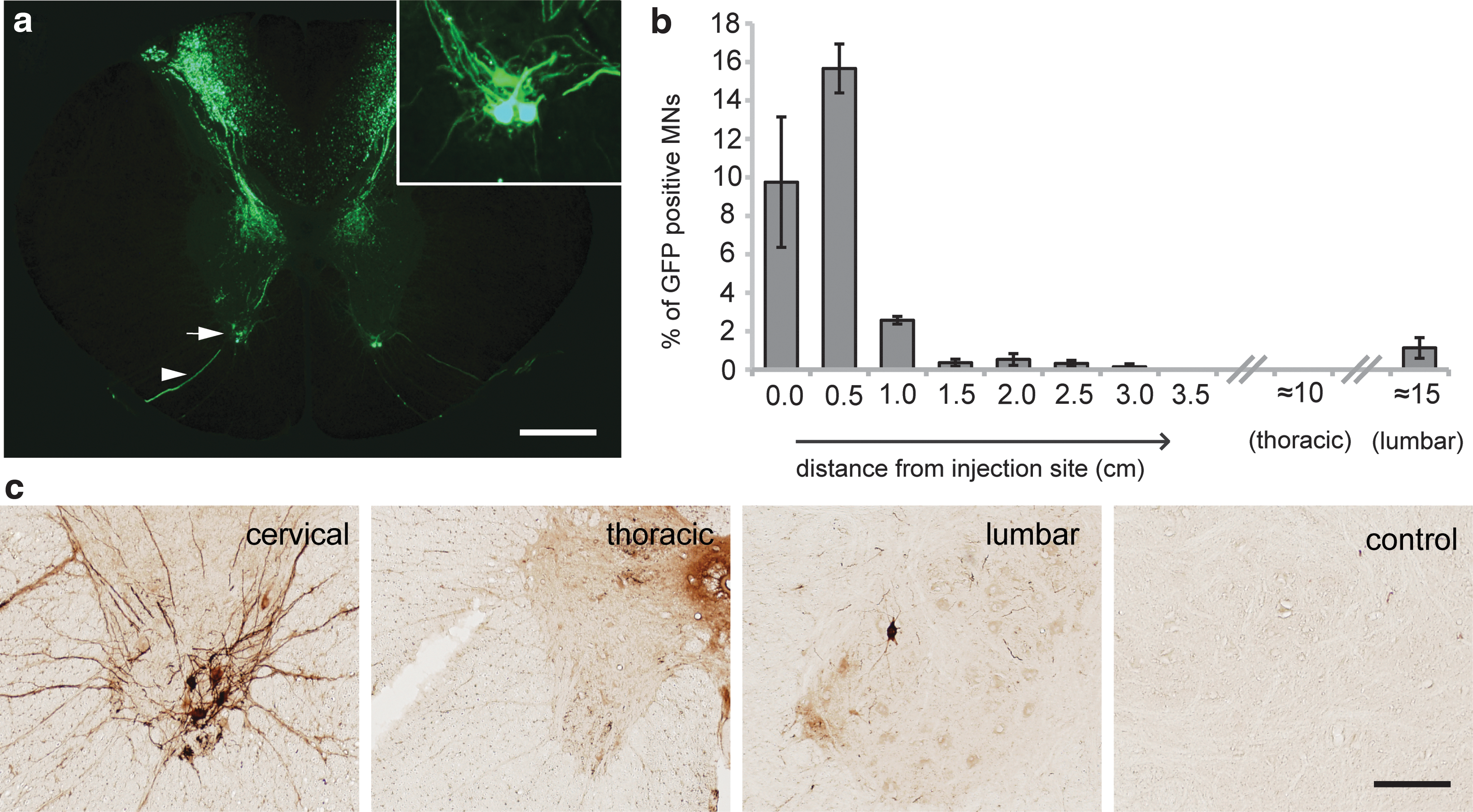
AAV6-cmv delivery into the cerebrospinal fluid transduces motoneurons in nonhuman primates.
GFP expression was analyzed across the entire 20 cm of cord using DAB-based amplification against the fluorescent protein antigen (Fig. 7b and c). Motoneuron transduction was highest close to the injection site (averaging up to 15% at some cervical levels) and reduced moving caudally along the cord. Interestingly, while no GFP-positive motoneurons were observed in the thoracic spinal cord, infected cells were observed in the lumbar enlargement (Fig. 7c). This is consistent with the rodent observation of higher transduction rates in lumbar versus thoracic level after CSF delivery rostral to these zones.
Discussion
In the present study, we established a dual-AAV vector system to specifically overexpress a transgene in motoneurons and/or astrocytes. We combined AAV6 and AAV9 serotypes, used the ICV route of vector delivery, and assessed several promoters to achieve a cell type-specific pattern of transgene expression. In particular, we determined the conditions needed to transduce motoneurons and astrocytes along the entire spinal cord. By targeting cell types that play a critical role in motoneuron disorders, we address specific requirements for effective gene delivery to the spinal cord.
Delivering viral particles directly into the CSF represents an attractive method to transduce cells that are dispatched along the whole spinal cord. Functional relevance of this injection technique has been demonstrated in a severe SMA mouse model. The phenotype could indeed be rescued by ICV and intrathoracic injection of AAV8 encoding the SMN protein in newborn mice (Passini et al., 2010). Here, vector injection in the ventricles of mouse pups is shown to effectively spread AAV6 and AAV9 particles across the CNS, including the spinal cord. However, leakage of viral particles outside the CNS does occur as vector genomes could be detected in peripheral tissues. Interestingly, the extent of leakage depends on the serotype used. Indeed, the infection profile after ICV injection of AAV6 indicates that most of the virus remains in the CNS. This profile appears different when AAV9 particles are injected ICV. Indeed a significant amount of viral AAV9 DNA per cell was found in the liver 3 weeks postinjection. The liver tropism is not surprising, as AAV9 has been described to efficiently infect liver cells when injected intravenously (Inagaki et al., 2006). Some virus leakage out of the CNS most likely occurs via spreading in blood at the time of injection. The differences observed between AAV6 and AAV9 suggest, however, a contribution of unidentified serotype-specific mechanisms. Moreover, it was shown that AAV9 presents slow blood clearance (Kotchey et al., 2011). Functional relevance of virus leakage and liver transduction by AAV9 is not known. The astrocyte-specific gfaABC1D promoter used here should, however, likely limit expression of a transgene in liver cells.
To target motoneurons, we combined ICV injections with the AAV6 serotype and the constitutive cmv promoter. The pattern of transduction we observed after recombinant AAV6 particles were injected intracerebroventricularly in newborn mice was highly reminiscent of retrograde transport. Indeed, the overall specific targeting of motoneurons suggests preferential infection of the ventral roots and subsequent transport of the particles to the soma. We have shown in mice and nonhuman primates that AAV6 is efficiently transported retrogradely in motoneurons after intramuscular injection (Towne et al., 2010, 2011). It is hence likely that AAV6 particles are similarly transported retrogradely after infection of the ventral roots. When the same serotype and injection paradigm were tested with a cba promoter, few astrocytes were also shown to express GFP. They were, however, clustered in the spinal white matter, hinting toward limited diffusion of viral particles in the spinal cord parenchyma. Restricted diffusion might be related to the heparin-binding properties of AAV6. Sequestration of AAV6 particles by heparan sulfate of the extracellular matrix might indeed limit vector mobility. Hence, it appears that some motoneuron specificity is conferred by the combination of AAV6 serotype and ICV injection, which allows for the use of constitutive promoters with high-level transgene expression. The pattern of transgene expression in the spinal cord gray matter was most specific to motoneurons by using the cmv promoter. Indeed, while GFP expression is observed in some astrocytes of the spinal cord after ICV injection of AAV6-cba:GFP vectors, this is rarely the case with AAV6-cmv:GFP, even when immunohistochemical detection of GFP is performed. Higher specificity for neuronal cells or even motoneurons could be achieved by combining AAV6 with neuron-specific promoters, such as a minimal Hb9 promoter (Peviani et al., 2012). Depending on the envisaged application, the level of transgene expression in motoneurons could be a decisive factor between cell type-specific promoters and constitutive promoters such as cmv.
Beside motoneurons, targeting of astrocytes is of particular interest for non-cell-autonomous diseases such as ALS. Astrocytes have indeed been implicated in disease pathogenesis as they have been shown to modify disease progression (Yamanaka et al., 2008). Most AAV serotypes display a preferential neuronal tropism. For instance, only a limited number of astrocytes were observed to express GFP using serotype 6 AAV particles. Capsid engineering has allowed to shift AAV tropism toward glial cells (Koerber et al., 2009). Among natural serotypes, the rh43 AAV has been shown to infect astrocytes after direct intracerebral injections (Lawlor et al., 2009). Extensive transduction of astrocytes has also been demonstrated with AAV9 in particular when delivered intravenously (Foust et al., 2009). We show here that ICV injection of recombinant AAV9-gfaABC1D:GFP leads to extensive and highly specific expression of GFP in astrocytes of the brain and spinal cord. AAV9 particles appeared to broadly diffuse in the spinal cord parenchyma as astrocytes were transduced in the whole depth of the tissue. AAV9 particles do not bind heparin, which might allow better tissue penetration than AAV6. Interestingly, intrathecal injections of AAV9-cmv:GFP mostly led to expression of GFP in neurons in adult mice (Snyder et al., 2011). Our results suggest that AAV9 particles have a broad infection profile and that the promoter plays an important role at directing specificity.
Here, we show that coinjection of AAV6-cmv:RFP and AAV9-gfaABC1D:GFP allows differential transduction of both cell types along the whole spinal cord. Specific targeting of either motoneurons or astrocytes broadens modeling and therapeutic options for ALS, as different transgenes can be overexpressed in one cell type or the other. It also offers a tool for investigation of motoneuron–astrocyte interactions in physiological conditions. Patterns of transduction when AAV vector injections are performed in adults remain to be determined in the sight of therapeutic interventions. Injection in newborn mice is, however, relevant for pediatric diseases such as SMA. In addition, the use of AAV vectors to genetically modify motoneurons and astrocytes at birth could facilitate in vivo investigation of interactions between these two cell types in physiological conditions and along the whole pathogenic process of diseases such as ALS.
Relevance of ICV injections to target motoneurons in nonhuman primates is of importance for future potential clinical applications. Intrathecal injection of recombinant AAV9 has led to transduction of motoneurons in pigs (Bevan et al., 2011; Federici et al., 2011). Injection of AAV9 in the cisterna magna of nonhuman primates led to preferential transduction of astrocytes in the analyzed brain regions (Samaranch et al., 2012). We report here successful targeting of motoneurons after intracisternal injection of recombinant AAV6 in young African green monkeys. The pattern of transduction was reminiscent of retrograde transport as observed after intramuscular injections of AAV6 in nonhuman primates (Towne et al., 2010). Motoneurons transduced after intracisternal injection were mainly localized in proximity to the site of injection. Expression of the transgene was, however, also detected in some lumbar motoneurons. This encouraging observation suggests that increased viral titer and optimization of the injection technique might ensure broad transduction of motoneurons in large, young adult animals.
In conclusion, appropriate combinations of AAV serotypes and cell type-specific promoters can be used to target widespread distributed motoneurons and astrocytes via ICV vector delivery. AAV6 and AAV9 vectors can be combined for in vivo modification of gene expression in one cell type or the other, opening new avenues for the investigation of motoneuron–astrocyte interactions in physiological and disease conditions such as ALS.
Footnotes
Acknowledgments
The authors would like to thank Vivianne Padrun, Fabienne Pidoux, Christel Sadeghi, and Philippe Colin for their expert technical assistance, and Michael Brenner (Department of Neurobiology, University of Alabama) for kindly providing the truncated version of the GFAP promoter. The mouse work was supported by a grant from the Thierry Latran Foundation, by the ALS Association, and by the Merck-Serono EPFL Alliance. The primate studies were supported in part by Project ALS of New York, NY, the Axion Research Foundation, and the St. Kitts Biomedical Research Foundation. E.D. is supported by a fellowship of the Swiss Foundation for Research on Muscle Diseases.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.