
Abstract
Corneal transplantation is the oldest and one of the most successful transplant procedures with a success rate in many studies in excess of 90%. The high success rate is mainly attributable to the relatively immune-privileged status of the eye and the fact that the cornea is largely avascular. However, the success rate in patients with failed grafts is much lower such that regrafting is frequently the top indication for corneal transplantation in many centers. Neovascularization is the most important risk factor for rejection, as it allows access of the immune system to the donor tissue, compromising immune privilege of the graft/eye. We have developed a process to modify donor corneal tissue to prevent rejection by a single exposure to a gene therapy vector before surgery (EncorStat®). The vector used is based on clinically relevant equine infectious anemia virus (EIAV)-derived lentiviral platform and contains genes for two potently angiostatic genes, endostatin and angiostatin. We show that incubation of rabbit, primate, and human corneal tissue with the EIAV vector mediates strong, stable expression in the corneal endothelium. We have optimized this process to maximize transduction and, once this is complete, maximize the removal of free vector before transplant. Rabbit corneas treated with two different antiangiogenic expression vectors (EIAV-EndoAngio and to a lesser extent EIAV-Endo:k5) significantly suppressed neovascularization in a rabbit model of corneal rejection. As a result, corneal opacity, edema, and inflammatory infiltrates were reduced in these corneas. This study demonstrates that angiogenesis is a suitable target to prevent corneal rejection, and provides the first proof-of-concept data for the development of EncorStat, an ex vivo gene therapy treatment to prevent corneal rejection.
Introduction
T
The reported failure rate of corneal transplants varies widely between studies (3–68%), but in “low-risk” patients the rate is low (<10% in the first year). However, there is a constant rate of graft failure every year after surgery, so that 15 years posttransplant over half of all corneas have rejected (Williams et al., 2008) and require regrafting, resulting in more than $55M additional annual health-care-related costs in the United States. The leading cause of graft failure has consistently been found to be irreversible immune rejection (Panda et al., 2007; Patel, 2011). Neovascularization of the recipient corneal bed is a major risk factor for graft failure (Bachmann et al., 2010), because of the abrogation of immune privilege by allowing direct access of the systemic immune system to the graft. Regrafting itself is also a significant risk factor for further rejection (Panda et al., 2007; Williams et al., 2008; Rahman et al., 2010; Anshu et al., 2011; Perera et al., 2011), and this is becoming the main indication for this type of surgery in the United States and Europe (Boimer et al., 2011). Around one-third of all transplants are to such “high-risk” patients, and the failure rate is consequently higher, around 50–90% (Saban et al., 2010). In some of these patients the prognosis is extremely poor, with grafts failing at an accelerating rate to the point where they are no longer considered suitable for further transplants, despite an otherwise normally functioning visual system. There is therefore an unmet medical need in these high-risk patients.
The cornea lends itself to gene therapy for several reasons, such as accessibility (donor tissue is stored before surgery [normally up to 10 days with hypothermic storage, extending to 30 days with hyperthermic storage (Armitage, 2011)], allowing a convenient window for gene therapy intervention; corneal endothelial cells are highly accessible and are nondividing in humans; therapeutic protein can be produced locally and continuously, exactly where it is needed; gene transfer can be completed ex vivo, allowing all the vector to be easily removed by washing before surgery. The eye is also anatomically separated from the rest of the body, and so forms a natural barrier between itself and surrounding tissues; the eye–blood barrier provides another level of anatomic and immunologic isolation to prevent potential transfer of vector to the systemic circulation. Additionally, the short- and long-term safety of the procedure and function of the corneal graft can be readily monitored by noninvasive, quantitative, and standard methods used in eye banks/specialized centers. There are also good experimental animal models of corneal graft rejection to demonstrate efficacy of this treatment. Efficacy has been demonstrated in these models using gene therapy strategies targeting different biological processes such as immune system modulation (Konig Merediz et al., 2000; Pleyer et al., 2000; Klebe et al., 2001, 2005; Comer et al., 2002; Zhang et al., 2003; Beutelspacher et al., 2006), acceleration of wound healing (Gong et al., 2007), preventing apoptosis (Barcia et al., 2007), and suppressing neovascularization (Murthy et al., 2003; Rocher et al., 2011). These studies have employed a variety of vector platforms such as adenovirus, adeno-associated virus, and lentivirus. The latter platform is particularly attractive for gene therapy as lentiviral vectors integrate into the target cell genomes, mediating long-term expression, can transduce nondividing cells, and have shown to be highly effective at targeting the corneal endothelial layer (Wang et al., 2000; Murthy et al., 2003; Beutelspacher et al., 2005; Kampik et al., 2011).
Neovascularization is an ideal target to prevent corneal rejection because it acts on one of the main causes of rejection, the abrogation of corneal immune privilege, rather than down-stream effects, against which strategies such as blocking immune activation, preventing cell death, and increased healing are targeted. Endostatin and angiostatin are two potently angiostatic proteins that are found naturally in the eye to prevent aberrant angiogenesis, and are specifically involved in maintaining corneal avascularity after injury (Ohlmann et al., 2005; Azar, 2006; Ma et al., 2006). Their precise mechanism of action is still not fully understood, but they are known to act on vascular endothelial cells to induce apoptosis and block proliferation and migration, and both proteins have been shown to be potently antiangiogenic in numerous models of cancer (Blezinger et al., 1999a, b; Sauter et al., 2000; Jin et al., 2001; Wen et al., 2001; Shi et al., 2002, 2003; Hong et al., 2004; Noro et al., 2004; Ponnazhagan et al., 2004; Celik et al., 2005; Subramanian et al., 2005; Tjin Tham Sjin et al., 2006; Zhang et al., 2007) and corneal neovascularization (Kim et al., 1999; Shin et al., 2000; Lai et al., 2007).
We report the development of a gene therapy approach to make donor corneas resistant to rejection (EncorStat®). We show that rabbit corneas transduced with equine infectious anemia virus (EIAV) lentiviral vectors expressing endostatin and angiostatin (either kringles 1–3 or kringle 5) effectively suppressed neovascularization in a rabbit model of corneal rejection. This suppression reduced the infiltration of immune cells after surgery, and subsequently reduced corneal opacification.
Materials and Methods
Plasmids
The EIAV vector genome plasmids, EIAV-EndoAngio, EIAV-Endo:k5, and EIAV-GFP, contain a cytomegalavirus constitutive promoter driving transgene expression, a woodchuck posttranscriptional regulatory element and self-inactivating long-terminal repeats (LTRs) (Fig. 1). EIAV-EndoAngio contains the endostatin and angiostatin genes linked by an internal ribosomal entry site element (described previously in (Binley et al., 2012). EIAV-Endo:k5 contains the endostatin and angiostatin kringle 5 gene linked by an elastin linker motif (described previously by Murthy et al., 2003).

Schematic representation of equine infectious anemia virus (EIAV) vector constructs. Full-length EIAV vector genome transcripts are expressed from a cytomegalovirus (CMV) immediate early/EIAV long-terminal repeat (LTR) RU5 chimeric promoter. The vectors all contain a deletion in the U3 region (ΔU3) of the 3′-LTR resulting in self-inactivation (SIN) of the vector upon transduction. The neomycina (Neo) open reading frame (ORF), the internal CMV promoter, the transgene cDNAs (codon-optimized human endostatin [hEndo] and either codon-optimized human angiostatin [hAngio] kringle 1–3 or nonoptimized human angiostatin kringle-5, or enhanced green fluorescent protein [eGFP]) and woodchuck posttranscriptional regulatory element (WPRE) are indicated. aAn ORF at this position confers rev independence to the vector RNA during production, negating the need for both the Rev-responsive element (RRE) within EIAV vector genome or expression of the REV gene during vector production.
Vector production
EIAV vectors were produced by transient transfection of vector genome plasmid, gag-pol packaging plasmid, and envelope plasmid into HEK293T cells using Lipofectamine 2000 (Invitrogen) in 10-layer cell factories (Nunc) as described previously (Wong et al., 2006). Virus was pseudotyped with the G glycoprotein of vesicular stomatitis virus envelope and concentrated by double centrifugation (6000×g for 18–24 hr, and then 20,000 rpm for 1.5 hr).
Vector characterization
All of the vectors were characterized with respect to integration (strength titer) in HEK293T cells and RNA copy number. To determine the integration titer in HEK293T cells, DNA was extracted from the transduced cells after>12 population doublings to ensure dilution of unintegrated copies to undetectable levels. Integrated vector genomes were quantified by qPCR using a primer/probe set designed to the EIAV ψ sequence. A stock of plasmid DNA was used to prepare standards of known copy number to construct a standard curve. DNA prepared from HEK293T cells containing a single EIAV vector copy was used as an internal reference standard. The predicted number of TU per milliliter of each sample was calculated from the standard curve. Titers were adjusted to account for the number of cells at the time of transduction and the dilution and volume of the vector applied.
To determine the RNA copy number, RNA was extracted from neat virus using the QIAamp viral RNA mini kit (Qiagen). RNA copy number was quantified by qRT-PCR using the same ψ-specific primers and probe set as above.
Demonstration of ex vivo gene transfer using EIAV-GFP
Freshly harvested rabbit (Harlan Laboratories UK Ltd.) and primate corneal–scleral buttons (Centre for Macaques) and stored human corneal–scleral buttons (all human donor corneal tissues was kindly provided by Bristol Eye Bank) were transferred to a 12-well tissue culture plate containing 1 ml sterile MegaCell DME corneal storage medium (Sigma). After brief washing (by transferring to fresh wells three times), the corneas were transferred to a fresh well containing 2.6×106 TU/ml EIAV-GFP in MegaCell DME or medium alone as a control, and cultured for 1 month at 37°C. The medium was replaced twice a week to maintain tissue viability. GFP expression was measured in-well throughout, and in sections at the end of the experiment, by fluorescence microscopy.
Optimization of transduction process
Incubation time
Three matched pairs of human donor corneal–scleral buttons were used to assess the impact of incubation time on tissue transduction. Each matching pair was divided into eight quarters with eye scissors (Altomed) under aseptic conditions, and seven quarters were incubated for different times at 37°C in a 24-well tissue culture plate containing MegaCell DME with the EIAV-EndoAngio vector at a concentration 2.7×106 TU/ml. A negative control containing quarters in MegaCell DME alone was also included. Three corneal quarters (from different donors) were transduced per condition. After transduction, each quarter was washed 5 times by immersion in 0.75 ml fresh medium for 5 min with gentle agitation and cultured for 1 month at 34°C. The medium was replaced twice a week, and sampled 24 hr later for hEndostatin quantitation by ELISA (Quantikine human endostatin ELISA kit; R&D Systems).
Minimizing vector shedding
Donor human corneal–scleral buttons were divided into 4 quarters and transferred to a 24-well tissue culture plate containing MegaCell DME with the EIAV-EndoAngio vector at concentrations between 2.7×106 and 7.4×106 TU/ml and transduced for 18–24 hr at 37°C. Three corneal quarters (from different donors) were transduced per condition. After transduction, each quarter was washed 5 times by immersion in 0.75 ml fresh medium for 5 min with gentle agitation. At each stage, the medium was removed, RNA extracted, and vector copy number assayed by qRT-PCR. These values were used to calculate total transducing units and the percentage of starting dose in each sample.
Ex vivo transduction of rabbit corneas with the EIAV-EndoAngio vector
Freshly harvested corneal–scleral buttons from adult New Zealand white rabbits (Charles River) were transferred to a 24-well tissue culture plate containing 2 ml sterile Optisol-GS corneal storage medium. After brief washing (by transferring to fresh wells three times), the corneas were transferred to a fresh well containing Optisol-GS with or without vector at 2×106 TU/ml and incubated overnight at 37°C. The next day the corneal–scleral buttons were washed once more by transferring between wells containing 1 ml balanced salt solution (BSS; Alcon Laboratories) if before transplant, or 1 ml fresh MegaCell DME medium if culturing ex vivo. Care was taken to remove washes as gently as possible and to only handle the tissue by the scleral margin, to protect the endothelial layer.
Demonstration of endostatin and angiostatin expression after ex vivo transduction
Freshly harvested rabbit corneal–scleral buttons were transduced overnight with EIAV-EndoAngio in MegaCell DME (or medium alone as a control) and cultured on for 1 month at 37°C. The medium was replaced on days 1, 2, 5, 6, 12, 13, 19, 20, 27, and 28. Endostatin and angiostatin in the medium from days 2, 6, 13, 20, and 28 were measured by ELISA and represented as protein secretion/24 hr over time.
Suppression of neovascularization in the rabbit model of corneal rejection
Adult New Zealand white rabbits were handled according to procedures in accordance with the U.S. Animal Welfare Act and performed under Institutional Animal Care and Use Committee (IACUC) approval, which also conforms to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research standards for humane animal care.
Rabbits were anesthetized with isofluorane and the anterior ocular chamber was instilled with heparin and viscoelastic by paracentesis. A 7 mm corneal button was removed from the host eye using a trephine and replaced with a transduced or control donor buttons grafted into the host eye with 16 interrupted 7–0 nylon sutures, and the anterior chamber was reconstructed with the sterile BSS (Alcon Laboratories). Trephined corneal tissue from these animals was used as donor tissue for subsequent surgeries to reduce the number of animals required. Immediately after the corneal transplantation, the host eyes were administered with 0.1 ml enrofloxacin (Baytril; Bayer Animal Health; 23 mg/ml) and 0.1 ml triamcinolone (Kenalog; Westwood Squibb Pharmaceuticals; 40 mg/ml), as well as topical atropine (1%) and a single injection of carprofen (Rimadyl; Pfizer Animal Health; 2.5 mg/kg). Seven animals received control (buffer)-treated and EIAV-EndoAngio-transduced tissue, and four animals received EIAV-Endo:k5-treated tissue. Grafts were monitored twice weekly by slit-lamp microscopy and neovascularization was measured weekly by fluorescent angiography. Corneal thickness measurements were attempted using a PachyPen (Accutome Inc.). However, pachymetry testing was found to be unsuitable for analysis as pachymetry values were remarkably variable and were out of the range of the testing device soon after transplant. Quantitation of blood vessels and immune infiltrates from hematoxylin and eosin-stained corneal sections was also performed at the end of the study (see Supplementary Materials and Methods).
Corneal angiography
Rabbits were sedated with a ketamine injection (35 mg/kg i.m.) and one drop of proparacaine (0.5% Alcon) was placed on the cornea to minimize any discomfort before injection of fluorescein solution (10%; 3.5 mg/1 lb [0.45 kg]) via an ear vein. Corneal blood vessels were visualized and recorded with the fundus camera (FF 450 Zeiss). Images were taken every 2–10 sec for approximately 3 min, at which time the fluorescein solution will be cleaned from the blood stream (Supplementary Fig. S1; Supplementary Data are available online at
Statistical analysis
Statistical differences in protein expression from transduced corneas in the ex vivo experiment or in vivo neovascularization compared with the control group were determined using the Student's t-test (unpaired, two-tailed).
Results
Identification of transduced cells after transduction of donor tissue ex vivo
The aim of this study was to evaluate gene expression to corneas after incubation with EIAV vectors (Fig. 1), and to evaluate efficacy in a rabbit model of corneal rejection. All vector genomes were constructed using identical configurations (cytomegalovirus promoter, eGFP reporter gene, woodchuck hepatitis virus posttranscriptional regulatory element, self-inactivating LTRs). To identify the cell type transduced after incubation of donor corneal tissue, the EIAV-GFP vector was used. GFP expression was detectable by fluorescence microscopy from 3–4 days posttransduction, reaching peak expression by around 7–14 days and remained stable thereafter. Expression analysis indicated that epithelial and endothelial layers were transduced in human, primate, and rabbit corneas (Fig. 2).
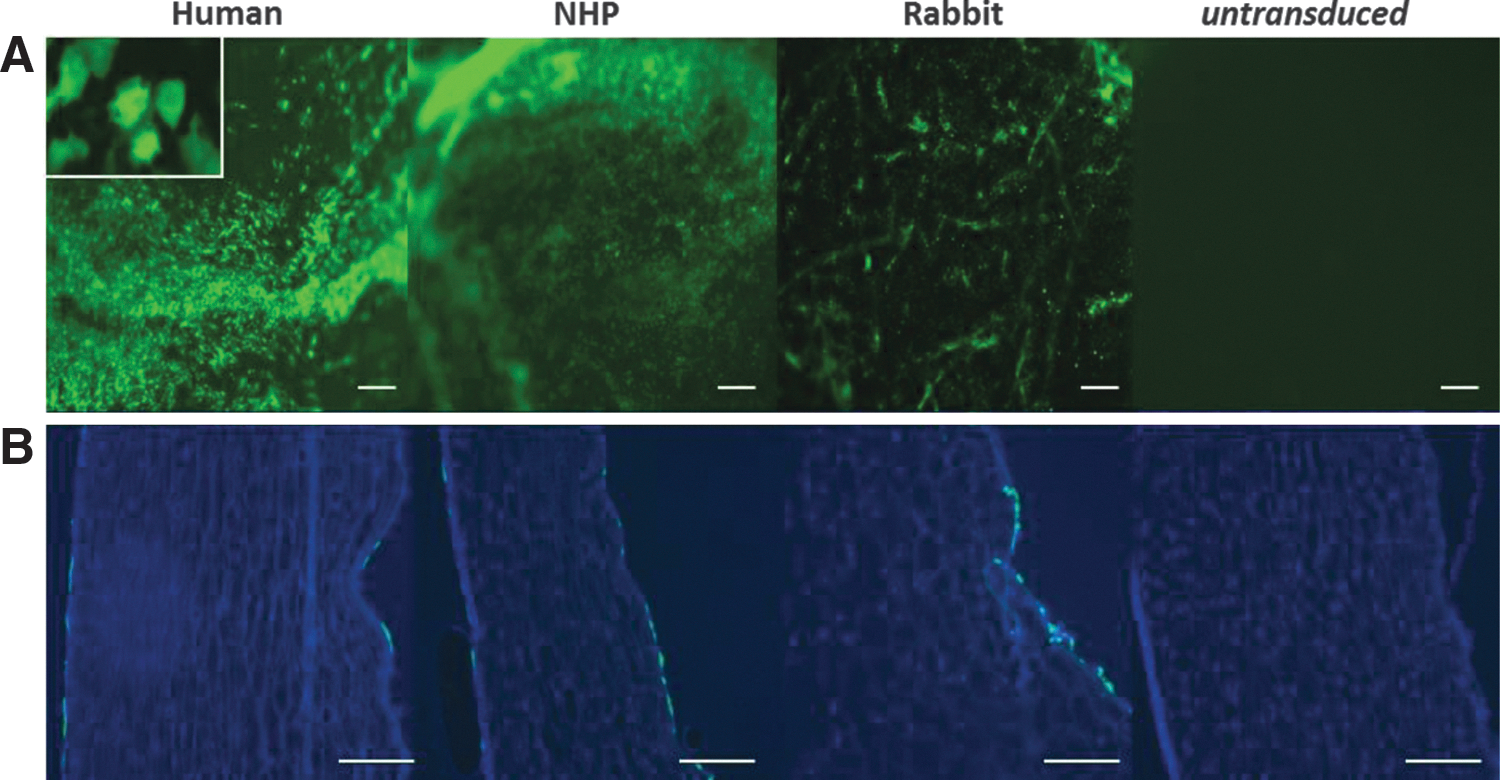
GFP expression in human, primate, and rabbit corneas. EIAV CMV eGFP vector (matched dose) was used to transduce donor corneal–scleral buttons ex vivo overnight and cultured for ∼1 month.
Optimization of transduction of human donor corneas
Human donor corneas incubated for 0–8 hr resulted in very low levels of endostatin production, whereas incubation times of 12–24 hr resulted in high levels of endostatin (Fig. 3). Angiostatin levels showed a similar pattern (data not shown). Protein production peaked at around 9 days and then began to drop off, presumably because of changes in the viability of the cornea ex vivo.
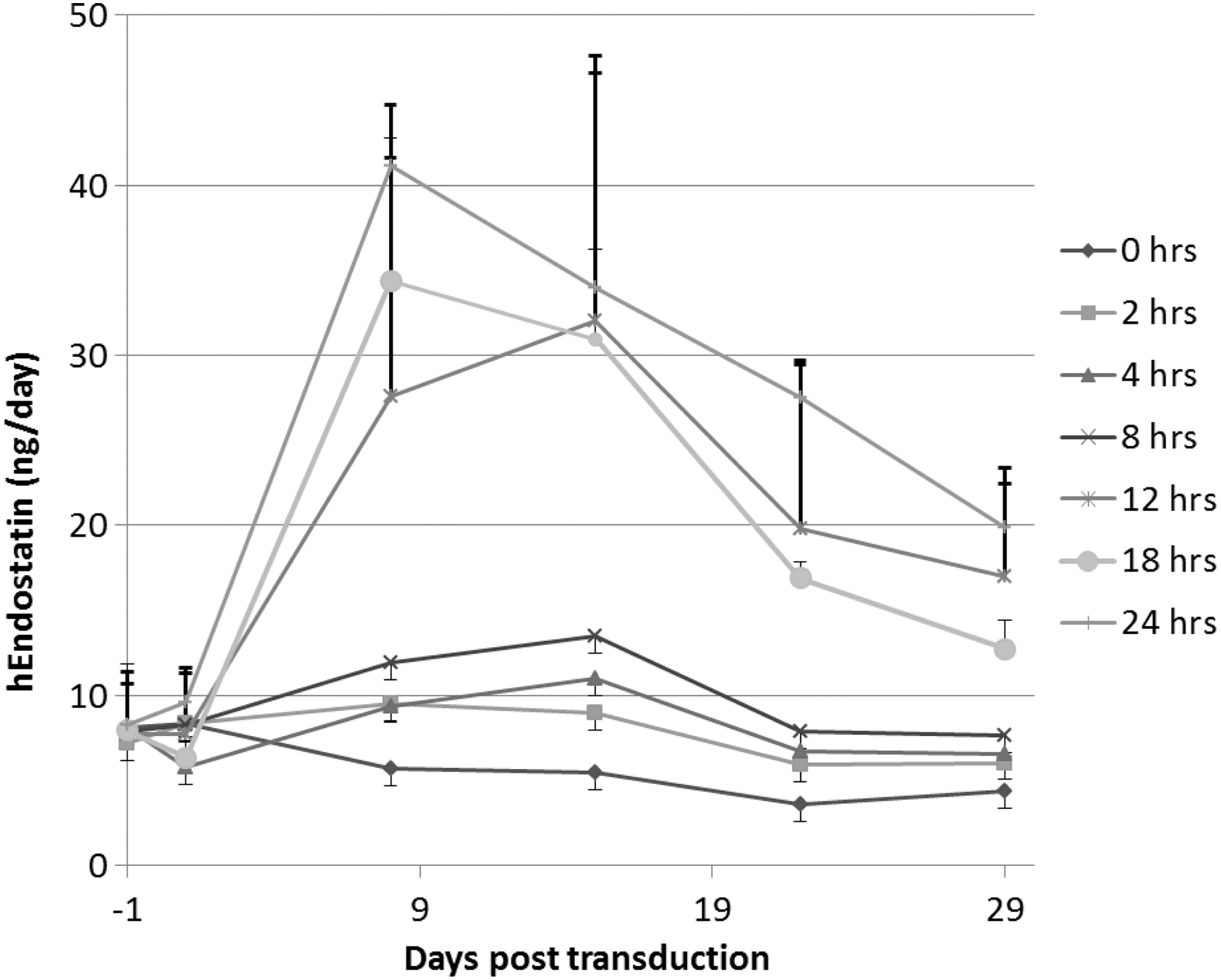
hEndostatin production over time from human corneas exposed to vector for different times. Human corneal–scleral buttons were dissected into quarters and transduced for 0–24 hr with the EIAV-EndoAngio vector at the same concentration. Tissue was washed and cultured on for 1 month after transduction, replacing the medium twice weekly and endostatin quantified 24 hr after medium change.
Most of the initial vector dose used to transduce human donor corneal–scleral tissue remained in the transduction medium after transduction (shown by comparable starting dose and posttransduction values in Fig. 4A). Of the vector remaining on the tissue, amounts shed at each wash step was very small (W1-5 in Fig. 4B). Most of this residual vector was removed after the first three washes, after which point <0.02% vector was shed at each subsequent wash. Similar results were also obtained with primate corneas (data not shown).

Measurement of vector “shed” from transduced corneas.
Rabbit corneas transduced with the EIAV-EndoAngio vector secrete human endostatin and angiostatin ex vivo
Rabbit donor corneas were transduced overnight in 2×106 TU/ml therapeutic vector (EIAV-EndoAngio) and were assessed for human endostatin and angiostatin production (Fig. 5). A steady-state level was reached within 1–2 weeks. A similar pattern of expression was obtained with EIAV-Endo:k5 (data not shown).
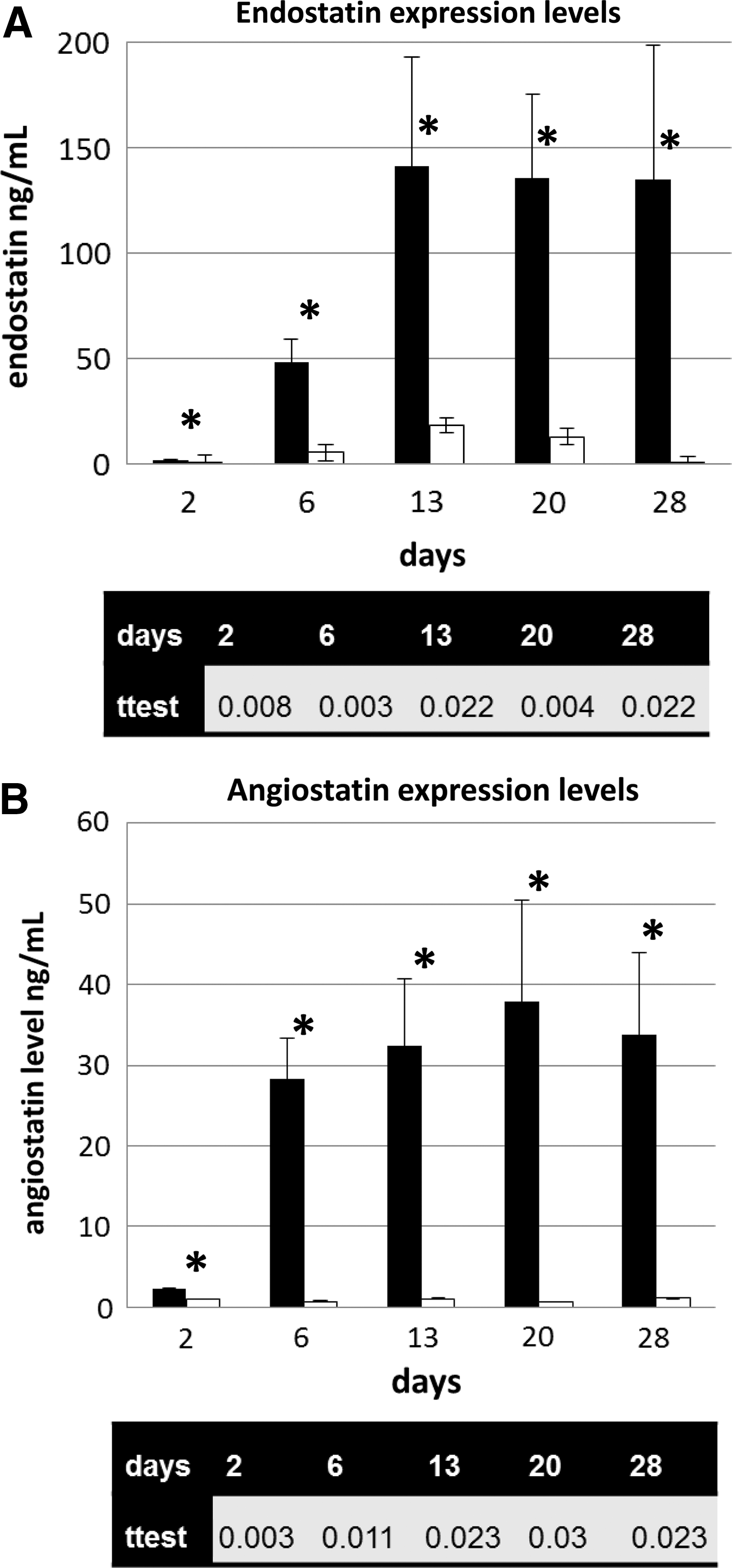
Expression of endostatin and angiostatin proteins by rabbit corneas transduced with the EIAV-EndoAngio vector ex vivo. Mean levels of expression (±standard deviation) in a 24 hr period of endostatin
Rabbit corneas transduced with EIAV-EndoAngio and EIAV-Endo:k5 vectors reduce neovascularization, immune infiltration, and corneal opacity in vivo
Rabbit donor corneas were transduced overnight with either EIAV-EndoAngio or EIAV-Endo:k5. The following day these were washed and transplanted into recipient rabbits. Corneal neovascularization (measured by fluorescent angiography; Fig. 6; see Supplementary Fig. S1 for representative images) and opacity (measured by masked scoring of ocular images; Fig. 7) were measured over the course of 1 month, after which time the animals were euthanized and the corneas removed for histological analysis. Neovascularization was also measured in histological sections (Supplementary Fig. S2) and shown to correlate with the fluorescent angiography measurements. Of the seven animals receiving control and EIAV-EndoAngio-treated corneas, one animal from each group was removed because of glaucoma and infection, respectively. Of the four animals receiving EIAV-Endo:k5-treated corneas, one animal developed an infection from day 15 so measurements after this time were discounted for this animal.
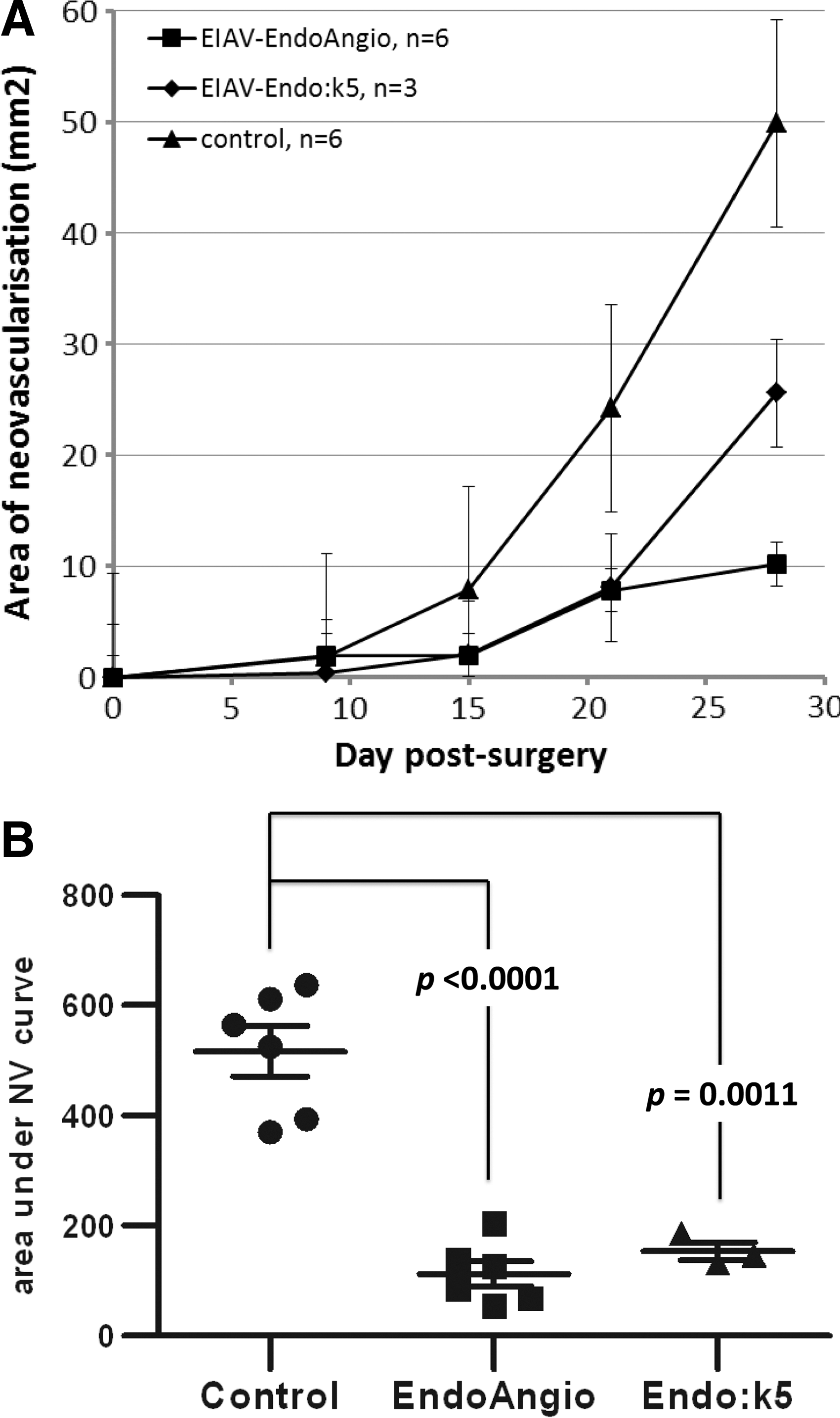
Area of neovascularization in a rabbit keratoplasty model. Corneas treated with EIAV-EndoAngio and EIAV-Endo:k5 vectors have significantly less postoperative neovascularization compared with control corneas.

Representative photographs of postoperative corneal clarity.
Total corneal neovascularization was widespread in all remaining six control corneas by day 28 but absent or significantly less severe in animals that received corneas treated with either therapeutic vector compared with untreated controls (Fig. 6) from day 9 (for the EIAV-Endo:k5 group) or day 15 (for the EIAV-EndoAngio group) for the duration of the experiment. Additionally, the vessels in all control corneas were found throughout the donor tissue, whereas 50% (3 out of 6) corneas within the EIAV-EndoAngio-treated group had neovascularization that failed to cross the donor recipient margin by postoperative day 28. However, within the EIAV-Endo:k5 group, only one rabbit out of three exhibited corneal neovascularization that did not cross the donor recipient margin by postoperative day 28.
Area under the curve analysis of the neovascularization in both groups over the duration of the experiment showed that EIAV-EndoAngio was more potent than EIAV-Endo:k5 (p<0.0001 compared with p=0.0011; Student's t-test, unpaired, two-tailed). Neovascular suppression was confirmed in histological sections of corneas at the termination of the study, which also showed a corresponding marked reduction in the infiltration of immune cells and an increase in corneal thickness (Fig. 8). Quantitation of these immune cells showed that the infiltrates were reduced by ∼50% in the EIAV-EndoAngio-treated corneas compared with either the untreated control corneas or the EIAV-Endo:k5-treated corneas (Supplementary Fig. S3); however, this reduction did not reach statistical significance (p=0.07). Additional histological staining with CD45+ antibodies confirmed that these infiltrates were inflammatory immune cells (Supplementary Fig. S4). Corneal opacity was consequently reduced in both (Supplementary Fig. S5); however, this also did not reach significance (p=0.11 and 0.09 for EIAV-EndoAngio and EIAV-Endo:k5 groups, respectively).
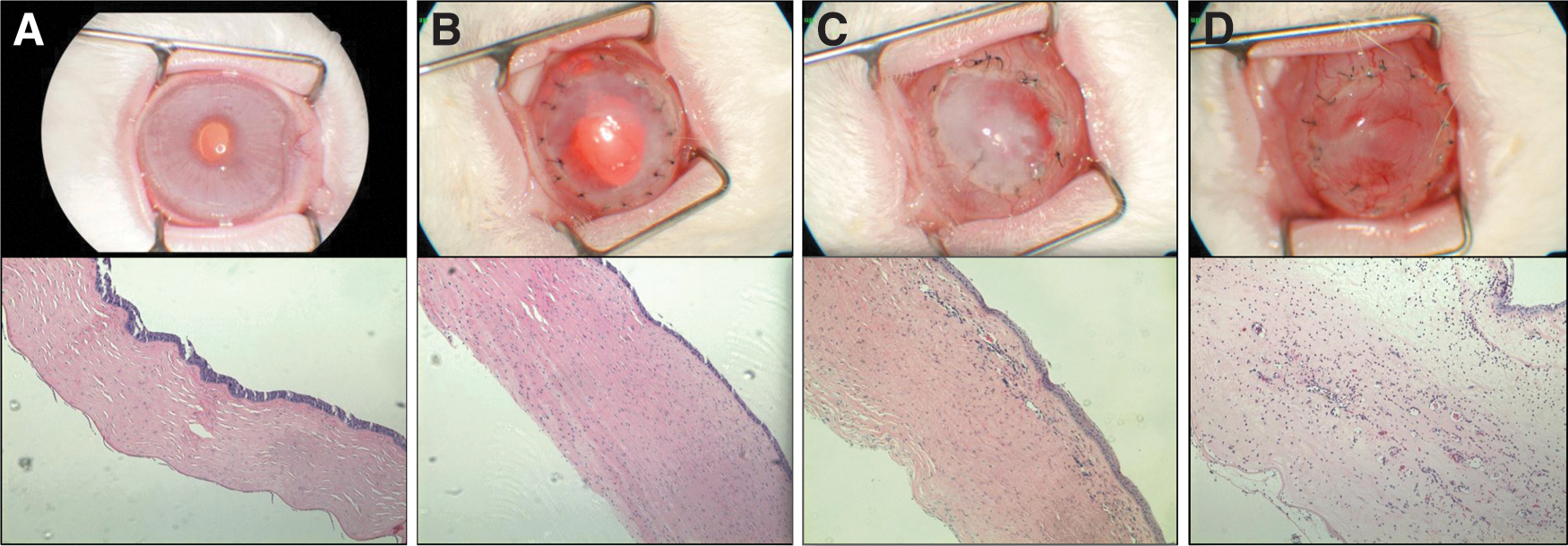
Representative photographs of postoperative corneal histopathology.
Discussion
Penetrating keratoplasty is the most successful tissue transplant worldwide partly because the normally avascular cornea is isolated from the immune surveillance machinery of the body. Pathologic, posttransplantation corneal neovascularization abrogates immune privilege by allowing direct access of the immune system to the graft. Suppressing neovascularization is therefore a valid therapeutic strategy to prevent corneal graft rejection. Moreover, even in the vast majority of successful grafts (usually described as unrejected at 12 months), a third will have had at least one episode of rejection that was successfully treated with aggressive, protracted, and expensive regimes of anti-inflammatory drugs and (less commonly) immunosuppressants, both of which have serious side effects. As these episodes are likely driven by the neovascularization associated with surgery, it is quite possible that these can also be reduced or prevented by an antiangiogenic therapy. In addition, the high level of success in year 1 for this type of surgery belies a constant, year-on-year rate of graft failure, such that by year 15 more than half of the successful grafts have rejected (Williams et al., 2008); although the cause of rejection in most instances is unknown, undiagnosed neovascularization, either transient or occult, cannot be ruled out. There is therefore a real need for a treatment to reduce corneal neovascularization after penetrating keratoplasty. High-risk patients are the obvious group to benefit the most from such therapy, but as highlighted it is also likely that prevention of neovascularization could prevent or reduce the need of aggressive anti-inflammatory/immunosuppressant regimes required to treat the transient episodes of rejection for a much larger proportion of patients, and it remains possible that, if the long-term failure rate has a neovascular cause, this treatment may have long-term benefit for all transplants.
Previously, we have demonstrated that rabbit corneas modified to secrete human endostatin and angiostatin (kringle 5) significantly reduced rejection in a rabbit model of corneal rejection (Murthy et al., 2003). In that study, an HIV-1 vector containing the Endo:k5 expression cassette was used to establish proof of concept for this therapeutic approach. However, to facilitate the clinical development of a therapy, we sought to (1) switch from this research-suitable HIV vector platform to the EIAV-based LentiVector platform, a clinically proven gene therapy vector that has shown good patient safety (with over 60 years of patient data to date in four ongoing clinical trials in Europe and the United States), that has shown to mediate robust and persistent gene expression in the human eye (Ellis et al., 2012) and has also been shown to be superior for the transduction of target corneal endothelial cells (Beutelspacher et al., 2005); (2) optimize the transduction process to maximize protein production and minimize vector shedding from human donor tissue; and (3) change the original Endo:k5 expression cassette to a similar cassette (EndoAngio) that has demonstrated good safety both preclinically (Binley et al., 2012) and clinically in an ongoing clinical phase I study (for wet age-related macular degeneration; trial number NCT01301443). Although these two expression cassettes are expected to have similar (but possibly not identical) angiostatic activities, using the EndoAngio cassette also reduces the potential for immunogenicity caused by novel epitope generation in the fused protein encoded by the EndoAngio cassette. The ultimate aim of this study therefore was to evaluate the EIAV-EndoAngio vector in the same rabbit model of rejection used previously, to ascertain comparability or superiority to the EIAV-Endo:k5 vector.
In this study we have shown efficient gene transfer to the target cell layer, the relatively amitotic corneal endothelium, as well as the rapidly dividing epithelium, after transduction of rabbit, primate and human corneal tissue in-well (i.e., ex vivo). This is consistent with previous studies by us and other groups (Wang et al., 2000; Murthy et al., 2003; Beutelspacher et al., 2005; Balaggan et al., 2006; Kampik et al., 2011). This cell layer is ideal for therapeutic transduction as in humans it does not divide, so transduced endothelial cells should remain for the life time of the graft. GFP expression was relatively rapid, detectable from day 3 to day 4, and persistent for the duration of the experiment in all three species. We were able to optimize the transduction process: we showed that optimal transduction required a minimum of 8–12 hr incubation, after which there was no improvement in protein production. We showed that most of the vector used for transduction is efficiently removed by a simple washing regime, resulting in <0.07% vector dose shed during additional washing and subsequent 9 days in cell culture combined. Moreover, this is an overestimation as the majority of this vector is inactive (we have shown that calculating vector dose based on an RNA assay leads to a threefold overestimation of active vector; data not shown). As with human corneas, transduction of rabbit corneas with both therapeutic vector configurations led to significant levels of therapeutic proteins. Again, transduction of donor corneal tissue led to rapid secretion of therapeutic proteins that reached significant and persistent levels.
We evaluated donor rabbit corneas transduced with either vector in a rabbit model of neovascularization. Corneal allograft rejection rates in the rabbit are typically less than 50% without steroid or immunosuppressant treatment, so inducing corneal neovascularization in the recipient before transplant (e.g., by damaging the cornea surface with heat or chemicals, or presuturing the recipient bed) is frequently employed to increase this rejection level (Gebhardt and Shi, 2002). In the model used here, the use of 7–0 nylon sutures that are retained induces neovascularization and rejection of the donor graft in almost all transplanted corneas. This is reflected in the wide-spread level of neovascularization observed in all untreated corneas. In the antiangiogenic vector-treated groups, however, neovascularization on day 28 was greatly reduced in all corneas compared with the control group, consistent with data from our previous study (Murthy et al., 2003). In this previous study the control group consisted of corneas treated with a lentiviral vector expressing GFP; in the current study, we used untreated corneas as our controls as these more closely mirror the clinical setting. Moreover, neovascular suppression was greater with EIAV-EndoAngio compared with EIAV-Endo:k5. Cornea opacification in addition to iris vessel background decreases accurate evaluation of cornea neovascularization in a New Zealand white rabbit model of cornea rejection. For this reason fluorescent angiography was used instead of slit lamp photography described in the previous studies. This method provides more accurate representation of posttransplant cornea neovascularization. Importantly, neovascularization into the graft was markedly reduced in the vector-treated corneas and was restricted to the recipient bed in half of the EIAV-EndoAngio-treated corneas (and a third of the EIAV-Endo:k5-treated corneas), unlike in the control corneas, where neovascularization grew from the surrounding recipient bed into the graft tissue. This finding shows that corneas secreting antiangiogenic proteins not only reduce total neovascularization but also reduce vascularization of the donor tissue specifically, thereby denying the immune system direct access to the graft. This was confirmed by histological analysis of corneal sections, which showed substantially less immune infiltration in the EIAV-EndoAngio vector-treated corneas compared with the controls, although this did not reach statistical significance. Lymphangiogenesis, also shown to be suppressed by both endostatin and angiostatin (Shao et al., 2005; Zhuo et al., 2011), has been shown increasingly to play a role in corneal graft rejection (Nakao et al., 2012) and we attempted to stain for these using antibodies to the lymphatic endothelial cell marker LYVE-1, but this was unsuccessful in these rabbit corneal sections (data not shown). Correlation to therapeutic protein expression in the aqueous was not possible because of a high background in the ELISA (Western blotting of these samples identified abundant high-molecular-weight bands that cross-reacted with endostatin and angiostatin antibodies—as these were not present in naïve animal aqueous, these were probably collagen XVIII and plasminogen products produced as a result of the wound healing process; data not shown). Concentrations in sera between groups were not statistically different (data not shown), presumably because of the relatively low-level production localized to the anterior chamber. Corneal edema on day 28 as determined from histology was also reduced in the vector-treated groups, as was corneal opacity although this also did not reach significance. We believe that failure to reach significance with these endpoints may be because of the nature of this model, as the sutures continue to drive inflammation throughout the in-life phase regardless of neovascularization. This is therefore an aggressive model of rejection, and as such it is very encouraging to see such a high degree of efficacy.
We have shown that corneas transduced with EIAV-based vectors containing antiangiogenic genes produce stable, high-level protein production, which prevent neovascularization of the donor tissue after surgery. By preventing neovascularization, the main driver of corneal rejection in this model, clinical markers of corneal rejection (corneal opacity and edema) were also reduced. These data provide good evidence of therapeutic benefit in pretreatment of donor corneal tissue with EIAV-EndoAngio before penetrating keratoplasty, by a process that would in many respects involve little alteration in the logistics of donor cornea preparation and storage employed by eye banks, and supports the clinical evaluation of EncorStat.
Footnotes
Acknowledgments
The authors would like to thank Dr. Val Smith and Bristol Eye Bank for kindly providing human donor corneas for research use.
Author Disclosure Statement
Competing financial interests exist for J.B., V.S., K.A.M., and S.E. No competing financial interests exist for M.P., T.M., M.H., A.Y., and T.H. Authors T.S. and B.A. are the inventors of technology that is used in this research and that has been licensed to Oxford BioMedica. This potential conflict of interest has been reviewed and managed by Oregon Health & Sciences University.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.