
Abstract
Adeno-associated virus serotype 8 (AAV8) has been demonstrated to be effective for liver-directed gene therapy in humans. Although hepatocytes are the main target cell for AAV8, there is a loss of the viral vector because of uptake by macrophages and Kupffer cells. Reducing this loss would increase the efficacy of viral gene therapy and allow a dose reduction. The receptor mediating this uptake has not been identified; a potential candidate seems the macrophage scavenger receptor A (SR-A) that is involved in the endocytosis of, for instance, adenovirus. In this study we show that SR-A can mediate scAAV8 endocytosis and that blocking it with polyinosinic acid (poly[i]) reduces endocytosis significantly in vitro. Subsequently, we demonstrate that blocking this receptor improves scAAV-mediated liver-directed gene therapy in a model for inherited hyperbilirubinemia, the uridine diphospho-glucuronyl transferase 1A1–deficient Gunn rat. In male rats, preadministration of poly[i] increases the efficacy of a low dose (1×1011 gc/kg) but not of a higher dose (3×1011 gc/kg) scAAV8-LP1-UT1A1. Administration of poly[i] just before the vector significantly increases the correction of serum bilirubin in female rats. In these, the effect of poly[i] is seen by both doses but is more pronounced in the females receiving the low vector, where it also results in a significant increase of bilirubin glucuronides in bile. In conclusion, this study shows that SR-A mediates the endocytosis of AAV8 in vitro and in vivo and that blocking this receptor can improve the efficacy of AAV-mediated liver-directed gene therapy.
Introduction
Although hepatocytes are the main target cell especially for AAV8, other cells such as macrophages and Kupffer cells are able to take up AAV particles (Zaiss and Muruve, 2008). Overcoming the loss of vector to these nontarget cells may increase its efficacy and thus allow additional dose reduction. It is unknown which receptor is involved in the uptake of AAV uptake by Kupffer cells and other macrophages. Potential candidates are the scavenger receptors expressed on macrophages, Kupffer cells, and certain endothelial cells. This family is involved not only in the endocytosis of apoptotic cells and certain macromolecules such as lipoproteins, but also in that of blood-borne pathogens, including viruses (Peiser et al., 2002; Kzhyshkowska et al., 2012). Previously, we showed that the macrophage scavenger receptor A (SR-A/CD204) present on Kupffer cells can mediate endocytosis and transduction of macrophages and Chinese hamster ovary (CHO) cells by adenoviral vectors in vitro (Haisma et al., 2008). Blocking of SR-A with polyinosinic acid (poly[i]) in mice in vivo inhibited the clearance of adenovirus by Kupffer cells and thereby increased adenovirus-mediated transgene expression in hepatocytes (Haisma et al., 2008). For AAV it has been shown that in human serum the capsid associates with iC3b and that this mediates uptake of this vector by macrophages (Zaiss and Muruve, 2008; Goh et al., 2010). Since iC3b is recognized by SR-A1, upon association with this complement factor AAV might indeed be endocytosed via this receptor. Blocking it may therefore increase the efficacy of AAV-mediated liver-directed gene therapy.
Our goal is to develop gene therapy for Crigler-Najjar syndrome type 1. This inherited liver disorder is caused by a single gene defect resulting in deficiency of uridine diphospho-glucuronyl transferase 1A1 (UGT1A1), which leads, when left untreated, to a fatal unconjugated hyberbilirubinemia (Axelrod et al., 1957; Bosma et al., 1992). By catalyzing the glucuronidation of the hydrophobic unconjugated bilirubin (UCB) in the liver, UGT1A1 is of vital importance in the bilirubin homeostasis. The current standard treatment for Crigler-Najjar type I consists of phototherapy during early childhood. When later in life serum bilirubin levels start to increase, a liver transplantation will be required (Pett and Mowat, 1987). The burden of extensive phototherapy, the lack of sufficient liver donors, and the need for lifelong immune suppression makes the development of new effective treatments for Crigler-Najjar syndrome, such as liver-directed gene therapy, desirable. Using single-stranded and double-stranded AAV, we have demonstrated that AAV-mediated gene therapy is effective in correcting serum bilirubin in the Gunn rat, the appropriate animal model for Crigler-Najjar syndrome type 1 (Seppen et al., 2006; Montenegro-Miranda et al., 2011). The aim of this study was to investigate whether SR-A can mediate the endocytosis of AAV by macrophages and if so whether blocking it can increase the efficacy of AAV-mediated gene therapy for Crigler-Najjar syndrome type 1.
First, the effect of SR-A expression on the endocytosis of scAAV8 was determined in CHO cells. This revealed that SR-A expression resulted in a significant increase of AAV8 endocytosis. Blocking scavenger receptors with the general ligand poly[i] reduces endocytosis in these cells, in the human monocytic leukemia cell line THP-1, and in primary human interleukin 4 differentiated macrophages. In vivo, administration of poly[i] just before the injection of scAAV8-LP1-UGT1A1 improved the correction of hyperbilirubinemia in the Gunn rat. These results demonstrate that the human SR-A can mediate endocytosis of AAV by macrophages and suggest that specifically blocking this receptor improves AAV-mediated liver-directed gene therapy via the systemic circulation.
Materials and Methods
Cells
The CHO cell line was cultured in F12 medium (Gibco) supplemented with 10% fetal bovine serum (FBS), 2 mM L-glutamine, penicillin (100 IU/ml), and streptomycin (100 IU/ml) at 37°C in a humidified 5% CO2 atmosphere. The human acute monocytic leukemia cell line THP-1 was cultured in RPMI medium (Gibco) supplemented with 10% FBS, 2 mM L-glutamine, penicillin (100 IU/ml), and streptomycin (100 IU/ml) at 37°C in a humidified 5% CO2 atmosphere. THP-1 cells were differentiated and polarized as described by Daigneault et al. (2010). Briefly, THP-1 cells were seeded in a 12-well plate at a density of 1.0×106 cells per well and incubated with 200 nM phorbol-12-myristate-13-acetate (PMA) for 3 days, washed with HBSS, and subsequently cultured for 5 days in RPMI containing 10% FBS, 2 mM L-glutamine, penicillin (100 IU/ml), and streptomycin (100 IU/ml). Human peripheral blood mononuclear cells were isolated and differentiated to type 2a macrophages as described Monocytes were polarized and validated for specific phenotypic markers as described (Ambarus et al., 2012).
Fluorescence-activated cell sorting analysis
CHO, CHO SR-A, differentiated THP1, and differentiated primary human macrophages were seeded in a 12-well plate at a density of 1.0×106 cells per well and cultured overnight. The next day, cells were washed and incubated with Alexa-647-labeled scAAV8-LP1-UGT1A1±poly[i] 25 μg/ml preincubation for 5 min at 4°C or 37°C in medium supplemented with 10% human serum. After incubation for 4 hr, cells were washed, isolated, fixed in 2% formaldehyde, and resuspended in fluorescence-activated cell sorting buffer. Flow cytometry analysis was performed within 30 min after harvesting of the cells.
Animal experiments
All animal experiments were performed in accordance with the Animal Ethics Committee guidelines of the Academic Medical Center of Amsterdam. Gunn rats from our own colony with a weight between 160 and 260 g for males and between 140 and 170 g for females were used. The rats were fed ad libitum and randomly assigned to different treatment groups. Poly[i] 4 mg/kg body weight was injected in the tail vein 5 min before the injection of 1×1011 or 3×1011 of scAAV8-LP1-UGT1A1. Intravenous tail vein injection, blood sampling, and bile collection were performed as described by Montenegro-Miranda et al. (2012).
Bilirubin quantification
Total bilirubin in serum was determined by the routine clinical chemistry department using a standard colorimetric assay. UCB and bilirubin conjugates in bile were analyzed and quantified by high-performance liquid chromatography as described with the modification that a pursuit column (Agilent) was used (Seppen et al., 1996, 2006).
Construction of production of AAV vectors
The recombinant AAV vector was constructed by replacing the factor IX cDNA with the UGT1A1 cDNA, using the EcoR1 and Bsb1 sites of plasmid scAAV-LP1-hFIXco (kindly provided by Dr. A. Nathwani, University College of London, London, United Kingdom). The plasmid was sequenced to confirm proper insertion of UGT1A1. Recombinant AAV was produced with AAV2 Rep and pseudotyped with capsid from AAV8 using the adenovirus-free method. Briefly, AAV8scLP1UGTA1 vectors were produced after triple-transient transfection into adherent HEK293 cells. AAV8 particles were purified using AVB chromatography affinity gel and were finally formulated using diafiltration into D-PBS. Titration of AAV vectors was performed by quantitative PCR. AAV vectors were labeled with an Alexa Fluor 647 following instructions as provided by the manufacturer (Molecular Probes) followed by extensive washing to remove unbound label.
Results
Human SR-A mediates AAV endocytosis that can be blocked by poly[i] in vitro
To evaluate whether the SR-A can mediate endocytosis of scAAV8 vector, stable expression of the human receptor was established using a lentiviral vector as reported previously (Haisma et al., 2008) in CHO cells that do not have endogenous expression (Penman et al., 1991). SR-A expression resulted in a 3-fold induction of scAAV8 endocytosis, 74% versus 24% in the parental cell line after a 4 hr incubation at 37°C. To confirm uptake and to exclude a specific binding of the vector, a cold incubation was performed with these cells for 4 hr at 4°C. The lack of a detectable signal in those cells showed that binding to these cells is minimal, indicating that the signal at 37°C results from internalized vector (Fig. 1).
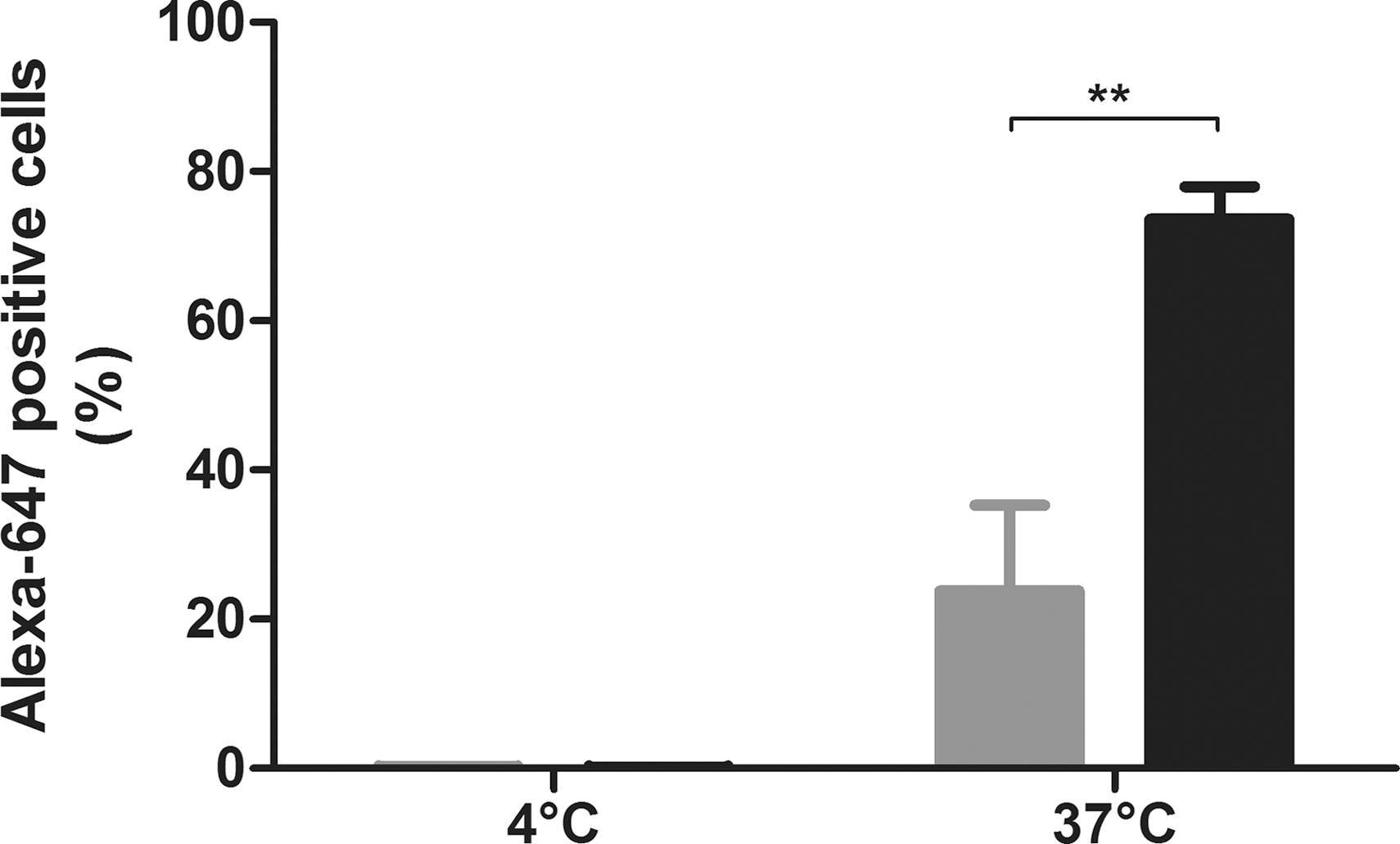
Scavenger receptor A1 can mediate AAV vector endocytosis. CHO and CHO-expressing SR-A1 were incubated for 4 hr with Alexa-647-labeled scAAV8 (300 gc/cell) in Dulbecco's modified Eagle's medium plus 10% HS at 4°C or at 37°C and were subsequently analyzed for scAAV8 endocytosis using flow cytometry. The percentage of CHO (gray bar n=3) and of CHO-expressing SR-A (black bar n=3) containing Alexa-647-labeled scAAV8 is presented±SD; **p<0.01. AAV, adeno-associated virus; CHO, Chinese hamster ovary; HS, human serum; SR-A1, scavenger receptor A1; SD, standard deviation.
The macromolecule complex poly[i] is shown to be an SR-A ligand (Platt and Gordon, 2001). Preincubation of the CHO SR-A cell line with poly[i] (25 μg/ml for 5 min) resulted in a significant 58% reduction of scAAV8 endocytosis (Fig. 2B). In the parental control CHO cell line, no effect of poly[i] on the uptake of scAAV8 was seen (Fig. 2A). Since these data indicate that SR-A mediates uptake of scAAV8, it was investigated subsequently if it could play a role in the endocytosis of this vector by human macrophages. First, the uptake was determined in the human monocytic cell line THP-1, which upon PMA differentiation resembles the type 2 macrophage (MΦ2) and exhibits significant expression of SR-A/CD204 (Tjiu et al., 2009). Upon incubation for 4 hr at 37°C, 26% of the differentiated THP-1 cells were positive for scAAV8 endocytosis. This percentage was dramatically reduced to 2% after preincubation of these cells with poly[i], indicating the important role of SR-A in this process (Fig. 2B). In the cells incubated with the vector at 4°C, the signals were very low, indicating that the fluorescence represents internalized vector.
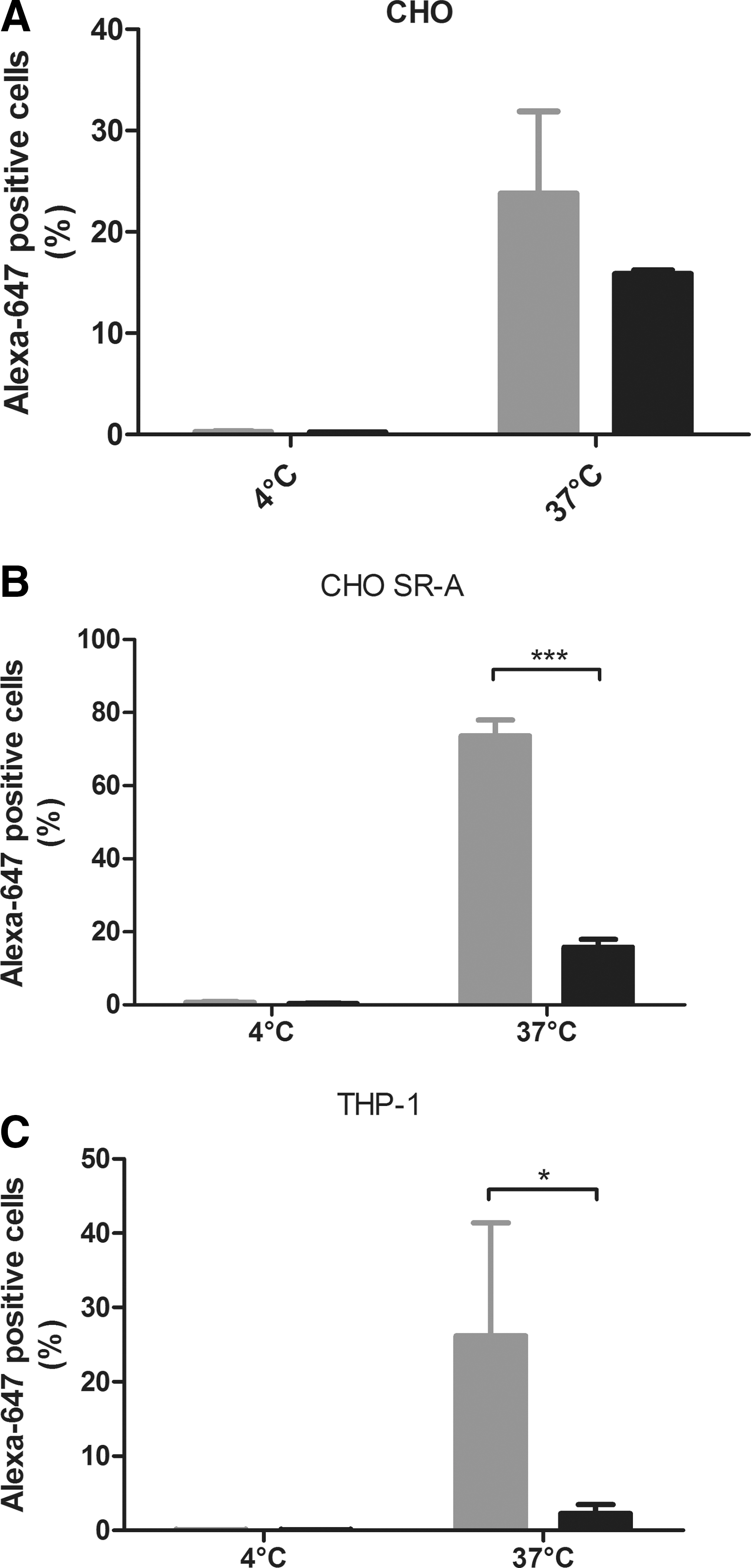
The scavenger receptor ligand poly[i] reduces endocytosis of the scAAV8.
Next we investigated the relevance of this receptor in endocytosis in primary human macrophages. Human peripheral blood mononuclear cells can be polarized into a wide range of different macrophages (MΦ) (Sica et al., 2006; Daigneault et al., 2010). Monocytes were polarized into MΦ1, MΦ2a, and MΦ2b as described (Ambarus et al., 2012). Extensive profiling revealed that monocytes that were differentiated into MΦ2a with interleukin 4 (20 ng/ml) have higher expression of SR-A in comparison to MΦ1 differentiated macrophages (100 ng/ml LPS plus 20 ng/ml IFN-γ) (Supplementary Fig. S1; Supplementary Data are available online at
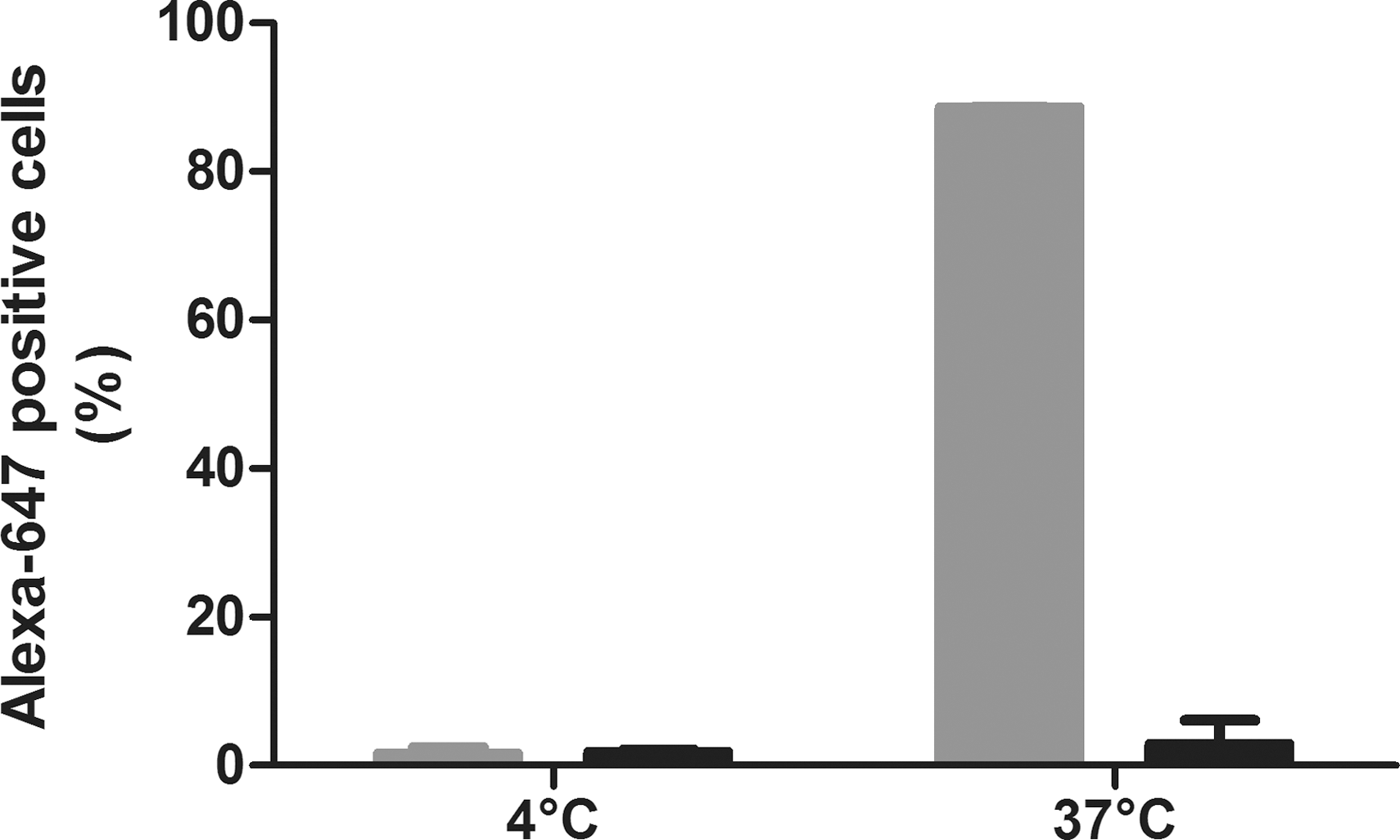
Blocking of the scavenger receptor reduces scAAV8 uptake in differentiated primary human macrophages. Human monocytes were isolated from blood and differentiated into type 2a macrophages (MΦ2a) with interleukin 4. After 4 days, cells were incubated with Alexa-647-labeled scAAV8 (300 gc/cell) in RPMI plus 10% HS in the presence (black bars) or absence (gray bar) of poly[i] (25 μg/ml) added 5 min before the vector. After 4 hr incubation at 4°C or 37°C, cells were subsequently analyzed for scAAV8 endocytosis using flow cytometry. Data are presented as average (n=2).
Altogether, these in vitro data demonstrate that enhancing the expression of the SR-A leads to increased uptake of AAV by CHO cells, differentiated THP1 cells, and primary human MΦ2a. Blocking this receptor by the potent ligand poly[i] leads to a large reduction of AAV uptake in all these cell types. This suggests that SR-A present on macrophages and Kupffer cells can mediate endocytosis of scAAV8 and therefore seems relevant in the loss of this vector to these cells upon systemic injection.
Preadministration of poly[i] enhances serum bilirubin correction and glucuronidated bilirubin secretion in Gunn rats treated with self-complementary AAV vector
To assess whether poly[i] affects scAAV-mediated liver-directed gene therapy, its effect on the correction of serum bilirubin level was studied in the Gunn rat (Montenegro-Miranda et al., 2011). Efficacy of scAAV8-LP1-UGT1A1 as a single agent or in combination with poly[i] was studied by determining the correction of serum bilirubin level and the secretion of bilirubin glucuronides into bile 16 weeks after viral injection.
Male and female Gunn rats were injected via the tail vein with a low dose (1×1011 gc/kg) or high dose (3×1011 gc/kg) of scAAV8-LP1-UGT1A1. Blocking of SR-A was established by injecting poly[i] (4 mg/kg) 5 min before vector administration. In male Gunn rats, tail vein injection of only scAAV8-LP1-UGT1A1 resulted in a significant reduction of serum bilirubin level by approximately 55%. Although the effect seemed somewhat more pronounced with the threefold higher dose, the difference between both treatment groups did not reach significance. Preadministration of poly[i] resulted in an increased correction of serum bilirubin in the male rats treated with the low viral vector dose (p<0.05). In the rats treated with the higher vector dose, the effect of poly[i] was minimal and not significant (Fig. 4A). Correction of serum bilirubin level in female rats upon tail vein injection of scAAV8-LP1-UGT1A1 in comparison to male rats was small. With the low vector dose, the effect only reached significance 1 week after injection, while the higher dose did provide a significant correction (p<0.01). Preadministration of poly[i] improved the efficacy of this vector in female rats and especially in the group treated with the low dose. In this group the reduction of serum bilirubin was significantly increased by poly[i] preadministration (p<0.05).
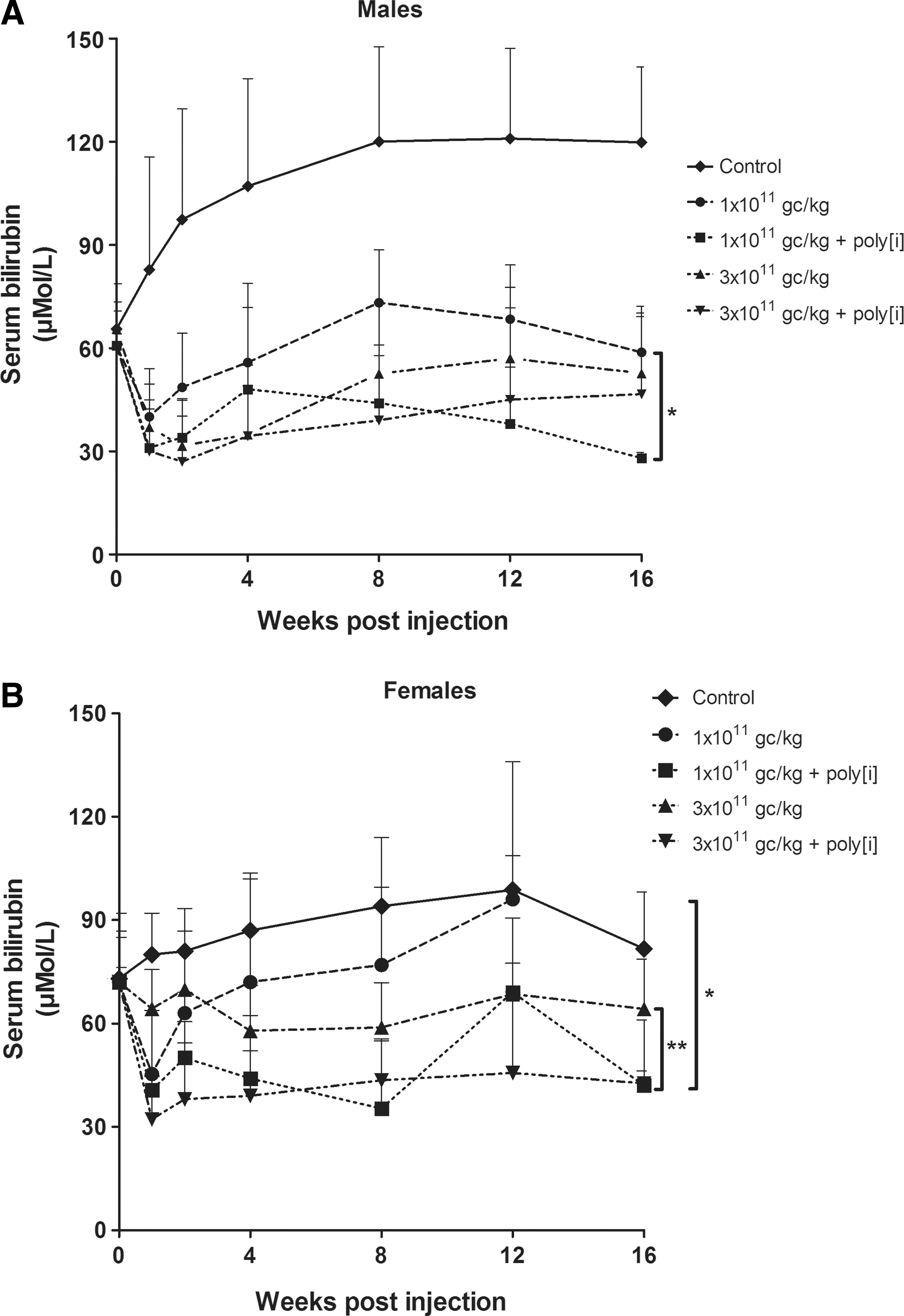
Poly[i] increases efficiency of scAAV8 viral gene therapy in rats. Gunn rats were injected in the tail vein with low- or high-dose scAAV8-LP1-UGT1A1 (1×1011 resp. 3×1011 gc/kg) with or without preadministration of poly[i].
Although UGT1A1 is the major role determinant, other processes such as bilirubin production from heme, its consumption by oxidative processes, and its re-uptake from the intestine do also affect serum bilirubin levels. The presence of bilirubin glucuronides in bile seems another possibly more sensitive parameter for the UGT1A1 activity re-established in the liver by scAAV8-LP1-UGT1A1-mediated gene therapy. At the time of sacrifice, 16 weeks after injection of the vector, bile was obtained and analyzed for the presence of bilirubin and bilirubin glucuronides. In both male and female rats, the amount of conjugated bilirubin secreted in bile was increased in the rats pretreated with poly[i] (Table 1). However, this only reached significance (p<0.05) in female Gunn rats treated with a low vector dose. Also, the amount of bilirubin diglucuronide in bile was significantly increased in the female rats pretreated with poly[i]. In this group 19% of total bilirubin in bile was bilirubin diglucuronide, while in the female rats receiving the low dose of the scAAV8-LP1-UGT1A1 but no poly[i], this was only 5% (Table 1).
The percentage of unconjugated (%UCB), monoglucuronidated (%BMG), and diglucuronidated (%BDG) bilirubin in bile was determined by high-performance liquid chromatography. All studies were performed at 16 weeks after vector administration. * p<0.05 and ** p<0.01 compared between female low-dose (1×1011) vector and female low dose plus poly[i] preadministration. # p<0.01 compared between female high-dose (3×1011) vector and female high dose plus poly[i] preadministration. Poly[i], polyinosinic acid.
Discussion
Crigler-Najjar syndrome is a rare but severe congenital disease that, when left untreated, leads to the development of kernicterus and can eventually be lethal. At this moment liver transplantation is the only curative option, but because of the lack of available donors and the burden of life-long immunosuppression development, a novel therapy is warranted. Liver-directed gene therapy with an AAV vector has shown to be a feasible and effective option for the treatment of monogenetic liver diseases such as Crigler-Najjar (Miranda and Bosma, 2009). A recent trial demonstrated that scAAV8-mediated liver-directed gene therapy is safe and effective, providing sustained correction of factor IX deficiency in hemophilia B, an x-linked bleeding disorder (Nathwani et al., 2011).
The healthy liver has an enormous capacity for the glucuronidation of bilirubin. Restoring only 5% of the normal UGT1A1 activity in Crigler-Najjar patients will reduce serum bilirubin levels effectively and will minimize the risk of the development of kernicterus (Fox et al., 1998). In six hemophilia B patients, a dose of 2×1012 gc/kg scAAV8-LP1-hFIX restored 8–12% of the normal expression levels of FIX. Although this dose was well tolerated and did not cause any acute toxicity, after 4–6 weeks elevated aminotransferase levels in the high-dose cohort indicated that this vector dose did induce an immune response in one patient. A short treatment with prednisone rapidly reduced these levels and prevented loss of efficacy (Nathwani et al., 2011). Although there was not a clear correlation between the vector dose and immune responses in this small group of patients, the dose required for correction was high enough to induce a CTL response at least in some of them. This indicates that there is room for optimizing vector delivery to the liver to lower the therapeutic vector dose.
It has been shown that cells of the innate immune system, such as macrophages and Kupffer cells, are capable of taking up virus particles, thereby providing protection toward viral infections (Smith et al., 2008). When aiming to treat inherited liver disorders with viral vectors, this reduces the amount of vector that reaches the hepatocyte and thus impairs the efficacy. Indeed, depletion of Kupffer cells increased hepatocyte transduction with adenovirus (Kuzmin et al., 1997; Shayakhmetov et al., 2004). We demonstrated previously that uptake of adenoviral vectors in these cell types could be reduced by the scavenger receptor ligand poly[i], showing that the scavenger receptors are involved in vector clearance (Haisma et al., 2008). Preventing the loss of viral vector to nontarget cells such as Kupffer cells by pretreatment with poly[i] indeed increased adenoviral transduction of hepatocytes, thereby enhancing vector efficacy. Although for AAV Kupffer cell depletion did reduce the innate immune response toward AAV (Zaiss et al., 2002; Martino et al., 2011), the potential beneficial effect on vector efficacy has not yet reported. The fact that Kupffer cells are directly involved in the induction of an innate immune response toward AAV indicates that there will be loss of the viral vector to these cells. The uptake of AAV by macrophages has been shown to depend on binding to the complement factor iC3b. Since this factor is recognized by SR-A, endocytosis of AAV may be mediated via this receptor (Goh et al., 2010).
Data presented in this study indeed demonstrate that like adenovirus, endocytosis of AAV8 can also be mediated by SR-A. Furthermore, these data show that AAV uptake in human macrophages in vitro can be effectively reduced by the SR-A ligand poly[i]. Previously, we showed that poly[i] by reducing the sequestering of adenoviral vectors by Kupffer cells enhances transgene expression in hepatocyte in mice and rats almost 10-fold (Haisma et al., 2008). In mice a single dose of poly[i] appeared safe and did not induce hepatocellular damage. Now our data show that intravenous administration of poly[i] just before vector injection also significantly increased the efficacy of scAAV8-LP1-UGT1A1 in the Gunn rat, although to a lesser extent as previously observed with adenoviral vectors. Although the increased efficacy was seen in male and female rats, poly[i] established the most pronounced effect in female rats treated with a low vector dose. As seen in female rats, also in males, poly[i] had the largest effect on the group injected with a low viral dose. One could envision that injection of a vector dose high enough to saturate the Kupffer cell system will allow the vector to disseminate and to transduce other cell types, especially the hepatocytes. Blocking of the scavenger receptor will therefore have a smaller effect on liver transduction when injecting a high saturating viral vector dose. In addition to vector dose, the effect of poly[i] also seems to depend on sex. The most pronounced effect of SR-A blocking is seen in female Gunn rats. This suggests a larger uptake of AAV by Kupffer cells and other macrophages, resulting in a higher threshold requiring a higher vector dose to saturate this system in females. A more active uptake by these innate immune cells may explain the some what reduced efficacy of double-stranded scAAV8 as observed in female mice (Paneda et al., 2009; Dane et al., 2012). Our data in rats now seem to indicate that in part this difference may result from an increased loss of vector to the macrophages. It is known that sex does influence the activity of the immune system in processes such as inflammation and cancer development (Csaba and Pallinger, 2009; Melgert et al., 2010). In addition, it has been shown that in males and ovariectomized and aged females, there is a reduced immune function (Angele et al., 2000). Furthermore, in vivo estrogen administration does induce Kupffer cell phagocytosis activity in rats (Yang et al., 1999; Hsieh et al., 2009). This hormonal effect is reflected in a greater than 1-log increase in AAV-mediated transgene expression in ovariectomized females compared with normal females, which then returned to normal levels upon addition with 17-β-estradiol (Lonning et al., 2002). All these data do suggest that in female rats the macrophages clearance of AAV may indeed be higher in comparison to male rats. Such an increased phagocytosis may explain why the effect of SR-A blocking by poly[i] may be more effective in female rats. Since most inherited liver disorders are inherited autosomally, this sex effect on the efficacy is very relevant for the clinical use scAAV liver-directed gene therapy in both sexes.
In conclusion, our data demonstrate that macrophage scavenger receptor can mediate the endocytosis of AAV serotype 8. In addition, we showed that in vivo administration of poly[i] before the viral vector improved outcome of liver-directed gene therapy in the Gunn rat, especially in females, suggesting that the lower efficacy in this sex seems to caused by an increased uptake by the macrophages. Therefore, these results seem of relevance for scAAV8-mediated liver-directed gene therapy when aiming to treat autosomally inherited liver disorders.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.