
Abstract
We have examined the role of a novel cytokine, interleukin-27 (IL-27), in mediating interactions between prostate cancer and bone. IL-27 is the most recently characterized member of the family of heterodimeric IL-12-related cytokines and has shown promise in halting tumor growth and mediating tumor regression in several cancer models, including prostate cancer. Prostate cancer is frequently associated with metastases to the bone, where the tumor induces a vicious cycle of communication with osteoblasts and osteoclasts to induce bone lesions, which are a significant cause of pain and skeletal-related events for patients, including a high fracture risk. We describe our findings in the effects of IL-27 gene delivery on prostate cancer cells, osteoblasts, and osteoclasts at different stages of differentiation. We applied the IL-27 gene delivery protocol in vivo utilizing sonoporation (sonodelivery) with the goal of treating and reducing the growth of prostate cancer at a bone metastatic site in vivo. We used a new model of immune-competent prostate adenocarcinoma and characterized the tumor growth reduction, gene expression, and effector cellular profiles. Our results suggest that IL-27 can be effective in reducing tumor growth, can help normalize bone structure, and can promote enhanced accumulation of effector cells in prostate tumors. These results are promising, because they are relevant to developing a novel IL-27-based strategy that can treat both the tumor and the bone, by using this simple and effective sonodelivery method for treating prostate tumor bone metastases.
Introduction
M
Therefore, since no available therapies have shown to either improve survival or significantly delay tumor progression in advanced prostate cancer patients, an ideal agent likely would have to display the dual function of restoring bone health while impairing interactions between the tumor and the associated bone lesions. An emerging area of research, osteoimmunology, aims to investigate new immune stimulatory strategies with the potential to combat cancer while also stimulating regeneration of bone. We propose the use of a novel osteoimmune cytokine, interleukin-27 (IL-27), for achieving this therapeutic purpose. IL-27 halts tumor growth and mediates tumor regression preclinically in studies that include prostate cancer (Zolochevska et al., 2011), neuroblastoma (Salcedo et al., 2004), colorectal cancer (Engel and Neurath, 2010; Zhu et al., 2010), and breast cancer (Zhu et al., 2010). IL-27 is a heterodimeric cytokine composed of subunit IL-27p28, related to IL-12 p35, and subunit EBI3 (Epstein-Barr virus-induced gene 3), related to IL-12 p40. IL-27 is immunomodulatory, is predominantly produced by antigen-presenting cells in response to microbial or host immune stimuli, and is involved in the regulation of immune responses against tumor development. IL-27 synergizes with IL-12 in interferon gamma (IFNγ) production from naïve T and NK cells. The receptor for IL-27 is composed of WSX1 and gp130 subunits, and is highly expressed in lymphoid organs, particularly in naïve CD4+ T and NK cells. IL-27 signaling induces T-bet, IFNγ, and IL12-Rβ2 expression, promoting initiation of Th1 differentiation (Lucas et al., 2003; Takeda et al., 2003). Moreover, expression of IL-27 in tumor cells inhibits tumor development and induces T cell-dependent antitumor immune memory (Hisada et al., 2004; Salcedo et al., 2004), with either systemic (Zhu et al., 2010) or intratumoral (I.Tu.) (Zolochevska et al., 2011) IL-27 treatments eliminating tumors without any observed toxicity. IL-27 also shows antitumor activity through indirect mechanisms such as induction of natural killer and CTL response or inhibition of angiogenesis through induction of CXCL10 and CXCL9 (Feng et al., 2007; Zhu et al., 2010).
We have reported recently a direct role for IL-27 on reducing the communication between prostate tumor cells and bone cells (Zolochevska et al., 2013). The IL-27 actions involved direct effects on STAT1 and IFNγ signaling in tumor, T cells, osteoblasts (OB), and OC (OC). That report was the first evidence that IL-27 could directly mediate bone cell signaling that enhanced proliferation and differentiation of mature OB and prevented OC formation in coculture with tumor cells. Other direct effects of IL-27 reported on bone cells have included blocking human bone marrow OC formation, through induction of c-Fos and NFATc1, indispensable transcription factors for osteoclastogenesis (Furukawa et al., 2009), as well as IL-27 downregulation of RANK and TREM-2 and blockage of RANKL-mediated ERK, p38, and NF-kB activation in OC precursors (Kalliolias et al., 2010). The role of IL-27 in mediating bone cell phenotypic changes also has been described as indirect, via inhibition of OC differentiation (Kamiya et al., 2007), partially through activated T cell effects on survival and OC cell fusion. In summary, IL-27 inhibits osteoclastogenesis by suppressing multiple pathways that impact the response of OC precursors to RANKL, suggesting that this cytokine plays a strong homeostatic role in restraining bone erosion.
Other supporting studies implicated a dual role for IL-27 as an antitumor agent in multiple myeloma, where it reduced malignant plasma cell proliferation, with beneficial effects on bone cells, including inhibition of osteoclastogenesis and amplification of mature functional OB (Cocco et al., 2010). Moreover, their results indicated that IL-27 could simultaneously treat myeloma and the surrounding bone, without any evidence of inflammation. For prostate cancer treatment, the present work is the first to report the effect of IL-27 on tumor and bone in vivo. We propose that IL-27 represents a novel, promising therapeutic agent for prostate tumors that have metastasized to the bone, based on its multifunctional activity. We show here a role for the cytokine IL-27 in preventing OC formation and in promoting OB differentiation and also that in vivo gene delivery of IL-27 can significantly reduce the rate of tumor growth as well as normalize bone density. In combination, we propose that IL-27 can be used to simultaneouslty treat prostate tumor metastases and the surrounding bone lesions.
Materials and Methods
Cell culture and transfections
Mouse TRAMP-C2 cells were obtained from American Type Culture Collection (ATCC) and maintained in Dulbecco's modified Eagle's medium–nutrient mixture F-12 (DMEM:F12; Mediatech) with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin (PS). TRAMP-C2 cells were transduced with a lentivirus expressing activated H-rasG12V (Lv-Hras) (Zolochevska et al., 2011) at a multiplicity of infection of 1 and a lentivirus expressing mouse androgen receptor at a multiplicity of infection of 1, resulting in TC2R. The RM1 murine prostate cancer cell line (McCabe et al., 2008) has the ability to aggressively grow in the bone microenvironment upon intratibial implantation. Prostate cancer cells were cultured in DMEM:F12 (Cellgro) with 10% FBS (GeneMate) and 1% PS (Invitrogen), and regularly passaged by trypsinization (0.05% [v/v] trypsin, 0.53 mM EDTA; Cellgro). The conditioned culture medium from prostate cancer cells was obtained as follows: RM1 or TC2R cells were grown to 70% confluence and washed 3× in phosphate-buffered saline, phosphate-buffered saline was replaced with 2% FBS DMEM:F12 medium (Cellgro), and then the conditioned culture medium was collected 48 hr later. MC-3T3-E1 clone 14 mouse pre-OB were obtained from ATCC and cultured in 10% heat inactivated FBS plus alpha-minimum essential medium (a-MEM; Invitrogen) with 1% PS. Heat inactivation of FBS (ATCC) was carried out at 55°C for 30 min followed by storage at 4°C before addition to the medium. Raw264.1 mouse cells were cultured in DMEM (Cellgro)/10% FBS (GeneMate) with 1% PS (Invitrogen) and gently scraped for passaging.
Vectors
The mouse androgen lentivirus was polymerase chain reaction (PCR)-cloned with specific primers using NheI and BglII ends and cloned into a pccl-CMV-ires-EGFP vector. Lentiviruses were prepared as described (Nelson et al., 2011). For therapeutic studies, plasmid DNA vectors were prepared using a pCpGfree backbone (Invivogen) and consisted of either control empty vector (multiple cloning site or MCS) or vector containing humanized renilla luciferase (Rluc). For therapeutic studies, IL-27 vector contained mouse EBI3/p28 gene (hyperIL-27 excised from pORF9-EBI3/p28; Invivogen) in the pCpGfree backbone. Vectors were prepared for all experiments using Endofree kits (Qiagen).
Cell growth and coculture assays
For bone cell proliferation in coculture with prostate cancer cells, assays were performed in 24-well plates using 104–105 prostate cancer cells plated in the top chamber in 0.5 ml DMEM/10% FBS (for coculture with OC) or α-MEM/10% FBS heat inactivated (for OB coculture). The bottom chamber received 0.5 ml of the same medium and 104–105 OC or OB cells, with cells fixed and stained using a crystal violet assay for cell proliferation with absorbance changes read at 550 nm (A550) as described (Zolochevska and Figueiredo, 2011). For each cell line, the fold change over control sample was determined. For bone cell differentiation assays, 1.5×105 cells were seeded in 12-well format in triplicate and cultured with differentiation supplements as per manufacturer's instructions using supplements for differentiation. For inducing pre-OB (MC-3T3) differentiation, 50 μg/ml ascorbic acid and 10 mM beta-glycerol phosphate were used when cells reached confluency in a plate using 0.2% heat-inactivated FBS (ATCC). Cells were assayed at either day 14 or day 21 postsupplement treatment. After culturing in differentiation supplements, cells were fixed in ice-cold 70% ethanol for 1 hr at room temperature and stained for Alizarin S Red stain (Millipore) using manufacturers' protocols. For inducing Raw264.1 cell fusion and differentiation into OC, we cultured cells in 100 ng/ml RANKL for 7 days and stained for detection of tartrate-resistant alkaline phosphatase (TRAP; Sigma-Aldrich). Images of differentiated cells were acquired for 7–10 independent microscopic fields under a 10×lens using an epi-fluorescence microscope (Bioexpress), and the total area fraction covered by cells was determined using NIH ImageJ software.
Flow cytometry and ELISA procedures
Cells for flow cytometry detection were assayed at the University of Texas Medical Branch (UTMB) Flow Cytometry Core facility. Tumor-infiltrating lymphocytes were isolated as described by Zolochevska et al. (2011) following a single sonoporation treatment in vivo delivered either intratumorally or intramuscularly. TC2R tumors were collected 7 days after treatment, and 2–4×106 cells were stained, with 2.5–5×105 cells collected during FACS analysis. Cells were stained in cell staining buffer (BioLegend), and antimouse antibodies (0.25 μg) were used to detect surface marker expression, including antimouse CD8a-APC, CD3-PE, CD19-APC, NK1.1-APC, TCRVβ7-PE, and antirat TCRγ/δ-PE (BioLegend). For T regulatory (Treg) cell detection, a kit was used with FoxP3-A467, CD25-PE, and CD4-FITC antimouse antibodies (BioLegend). The corresponding isotype antibodies were used as controls. Flow cytometry was performed on a Becton Dickinson FACSCanto using Cell Quest Software (BD Biosciences), and data analysis was performed using FlowJo 9.1 (Tree Star Inc.).
In vivo studies
Animal care and procedures were performed in accordance with the UTMB institutional review board guidelines. Mouse prostate cancer cells (106 TC2R) were delivered intratibially in 10 μl sterile 0.9% saline to 6–8-week-old C57/BL6 male mice (Taconic). The tumor growth was monitored over time using vernier calipers. For micro-computed tomography (μCT), a Siemens Inveon multimodality scanner (Siemens Medical Solutions) was used with a time course of CT imaging of tibia at high resolution (FOV 37×56 mm) with routine acquisition parameters (80 kVp, 500 μA, and voxel size of 56×56×56 μm), exposure time 800 ms per projection, and 520 equidistant steps covering 360 degrees of rotation. Step-and-shoot mode of acquisition was used to provide better stability of images compare with continuous imaging. The scan time is about 20–30 min. All scans were carried out using isoflurane anesthesia and data analyzed using the Siemens Inveon Research Workplace software version 3.0 with 3D Visualization option. Isotropic resolution of the reconstructed images was 0.056 mm. Dark-field, light-field, and flat-field corrections were applied during calibration. Center offset was applied during positioning calibration. Intensity was in linear mode according to histogram calibration. Data were analyzed using the Siemens Inveon Research Workplace software with 3D Visualization option.
For gene delivery studies, we utilized NLS2, a polycyclooctene polymer with pendant tetralysine and NLS (nuclear localization signal) oligopeptide, synthesized as described in (Parelkar et al., 2011). We prepared polymers in low retention Eppendorf tubes, dissolved in nuclease-free water, and sterilized by filtration. The stock solution of NLS2 was diluted to enable complexation with pLuc plasmid DNA at an N/P ratio=6 (i.e., the ratio of protonatable nitrogens in the polymer, N, to DNA phosphates, P). DNA (12.5 μg) was added to nuclease-free water 1:1 to the polymer solution and allowed to equilibrate for a minimum of 35 min under sterile conditions. After polyplex formation, 2.2 μl sterile Micromarker microbubbles (VisualSonics) were added per tube and injected intramuscularly to the hind legs of male mice. After applying ultrasound (US) gel, we irradiated the muscle to mediate sonoporation and thus gene delivery of pRluc plasmid using a Sonigene sonoporator (VisualSonics) using settings of 1 MHz, 20% duty cycle, 3 W/cm2, and 60 sec. In vivo imaging for renilla luciferase expression in muscle was performed on day 6 after sonoporation using previously published procedures by intravenous coelenterazine substrate administration and collection of images within 10 min (Zolochevska and Figueiredo, 2009; Zolochevska et al., 2011).
Real-time quantitative reverse transcription–PCR (qPCR) analyses
Total RNA from 1 mg tissue was extracted using the Trizol method using polytron homogenization (Figueiredo et al., 2006) followed by a SurePrep kit (Fisher Scientific); 2 μg RNA was reverse transcribed using TaqMan (Applied Biosystems); 1 μl template cDNA was used in a real-time qPCR reaction with 2× Sybr green master mix (Applied Biosystems), and 10 μM each forward and reverse primers for experimental or β-actin control housekeeping gene. Reactions were run on an Eppendorf Realplex 2S, using 40×95°C/15 sec, 56°C/30 sec, and 72°C/30 sec, and analyzed using Realplex software. Primer sequences are available upon request. All samples were normalized to the internal β-actin control and expressed as change in C t relative to β-actin.
Histology
Injected tibiae were fixed and decalcified overnight in decalcification (CalEX) solution (Fishersci), cut longitudinally, and incubated overnight again in Decalcifier I. Tissue was then embedded in paraffin and sectioned (5 μm). Sections were stained with toluidine blue, and visualization and imaging were done using a photomicroscope at the Research Histopathology Core.
Statistical analysis
Assays were performed in triplicate, and values provided as mean±SEM or 95% confidence interval. Comparisons were performed using an unpaired t-test, and p<0.05 was considered to indicate a significant difference.
Results
IL-27 enhances proliferation of OB and reduces proliferation of OC
We examined the effect of autocrine or paracrine IL-27 expression on bone cells in vitro by using coculture models. Pre-OB (MC3T3E1 clone 14) or RM1 cells were transfected with control (empty vector) or IL-27-expressing plasmid. Then, cells were cocultured in the absence or presence of osteoblastic differentiation stimuli (ascorbic acid plus beta-glycerol phosphate supplements). The coculture of pre-OB or OB with RM1 resulted in no change in proliferation compared with control on day 14 (Fig. 1A and Supplementary Table S1; Supplementary Data are available online at

The effect of IL-27 on osteoblast (OB) and osteoclast (OC) proliferation.
In OC, the same coculture model was used but with a different differentiation supplement, RANKL, to induce OC formation from RAW264.1 monocytic lineage cells. In all groups, IL-27 exposure inhibited OC proliferation by day 7 relative to day 2 (Fig. 1B). Although there was a trend toward reduced proliferation, expression of IL-27 in OC (autocrine) did not significantly reduce OC proliferation compared with pre-OC control (p=0.2). However, expression of IL-27 in a paracrine manner (in prostate cancer cells cocultured with OC) resulted in a significant reduction of OC proliferation (p<0.03) (Fig. 1B). Expression of IL-27 in both OC and RM1 cells (OC-27+RM1-27) further reduced the OC proliferation compared with control (p<0.02) and this difference was also significantly reduced compared with IL-27 expression in RM1 alone (RM1-27) (p<0.04). Therefore, although providing paracine IL-27 (via tumor cell transfection) might be sufficient to prevent OC proliferation, interestingly, combined expression in both OC and RM1 target cells appears to help achieve higher cytokine efficacy against OC proliferation. This effect would likely suport efforts to halt bone erosion by using IL-27.
Impact of IL-27 on OB and OC differentiation
The normal differentiation from pre-OB to OB can be observed by utilizing stains such as alizarin S red for detecting OB differentiation and mineralization by day 21 of culture. In Fig. 2A, we show that coculture of RM1 with pre-OB does not seem to stimulate pre-OB differentiation (panel 1). When differentiation supplements are added (+AAB, ascorbic acid, and b-glycerol phosphate), some mineralization can be detected (panel 2). When pre-OB are transfected with IL-27 (autocrine) (Fig. 2A, panel 3), some differentiation can be seen compared with control transfected with empty vector (RM1/pOB). When differentiation suppplements are added, the effect of IL-27 is further enhanced, as evidenced by darker extent of mineralization staining (Fig. 2A, panel 4). When IL-27 is provided in a paracrine manner (RM1-27/OB), differentiation of pOB is enhanced, but only in the presence of supplements (Fig. 2A, panel 6) compared with without (panel 5). Finally, when IL-27 is provided in both RM1 and OB cells (Fig. 2A, panels 7–8), the differentiation effect is magnified further compared with expression in either cell type alone. Analyses of these coculture image data yields a semiquantitative comparison plot that is shown in Fig. 2B, with significant differences at p<0.04.
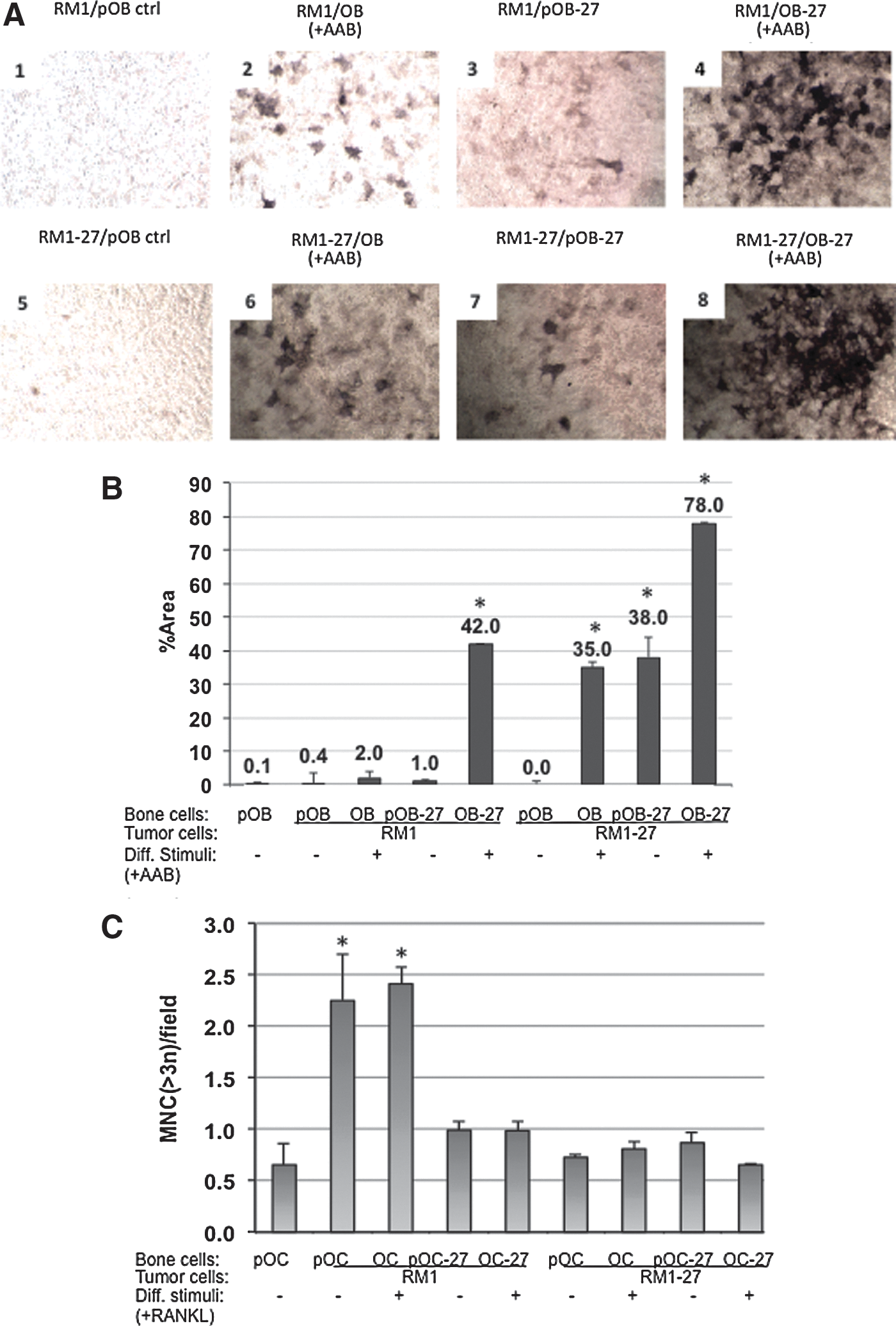
Effect of the conditioned medium from prostate cancer cells on OB differentiation.
The extent of osteoclastogenesis also can be assessed by staining OC cultures for the expression of TRAP and scoring the extent of multinucleated cells (MNC) or mature OC per 20× field. Raw264.1 cells pre-OC can differentiate and fuse into MNC in the presence of 100 ng/ml RANKL differentiation supplements (Hsu et al., 1999; Zolochevska et al., 2013). Prostate cancer cells also can induce osteoclastogenesis in coculture with pre-OC in the absence of RANKL (Fig. 2C). Interestingly, RANKL addition did not enhance the effect of coculturing with RM1 prostate cancer cells in inducing TRAP+ MNC cells (p<0.04). We examined the effect of expressing IL-27 on MNC formation as a measure of the impact of the cytokine therapy on osteoclastogenesis. IL-27 or empty plasmid control was expressed in either prostate cancer cells or bone cells and induced to differentiate by RANKL addition. First, OC displayed an ∼2.5-fold increase in MNC formation during differentiation with either RANKL treatment or during coculture with tumor cells, indicating osteoclastogenesis compared with untreated control (p<0.03). When IL-27 was expressed in OC (autocrine), this effect was suppressed and RANKL could not mediate osteoclastogenesis (Fig. 2C). When IL-27 was expressed in a paracrine manner (RM1-27), RANKL could not exert its osteoclastogenic effects either, suggesting that IL-27 expression can inhibit osteoclastogenesis in the presence of both strong prostate tumor cell and/or RANKL signals. There were no additive effects observed in the coculture when IL-27 was expressed in a combined paracrine and autocrine manner (RM1-27+OC-27), suggesting no added benefit to administering IL-27 to both cell compartments for halting osteoclastogenesis.
Development of an IL-27 gene delivery system in vivo
With the objective of developing a gene delivery system that would enable us to treat distant bone metastases of prostate cancer, we developed an intramuscular (I.M.) US-mediated gene transfer method utilizing plasmid DNA in complex with a novel polymer, oligolysine, and sonoporated in the presence of microbubbles (Fig. 3A). We first developed a protocol for Renilla Luc reporter plasmid DNA delivery to the hind leg muscle using sonoporation gene delivery (sonodelivery). For efficient sonodelivery, we used polyplexes of the novel comb-type oligolysine polymer, termed NLS2 complexed with plasmid DNA (Parelkar et al., 2011). The polymer consists of a polyolefin backbone with multiple copies of pendent tetralysine and NLS (nuclear localization signal) groups strung along the backbone. The NLS group consists of the PKKKRKV amino acid sequence, and the molar ratio of the two pendent groups (tetralysine and NLS) was approximately 1:1. This enhanced sonodelivery method resulted in ∼4-fold higher levels of reporter gene expression in muscle as compared with controls (Fig. 3B). The most effective gene delivery formulations with transfection exceeding that of naked DNA control contained the NLS2 polymer complexed either with sonovue microbubbles (SV+NLS2) or with micromarker microbubbles (MM+NLS2) (Fig. 3B, p<0.03). Representative images of naked DNA or MM+NLS2 are shown in the presence (+US) or absence (−US) of US stimulus (Fig. 3C).
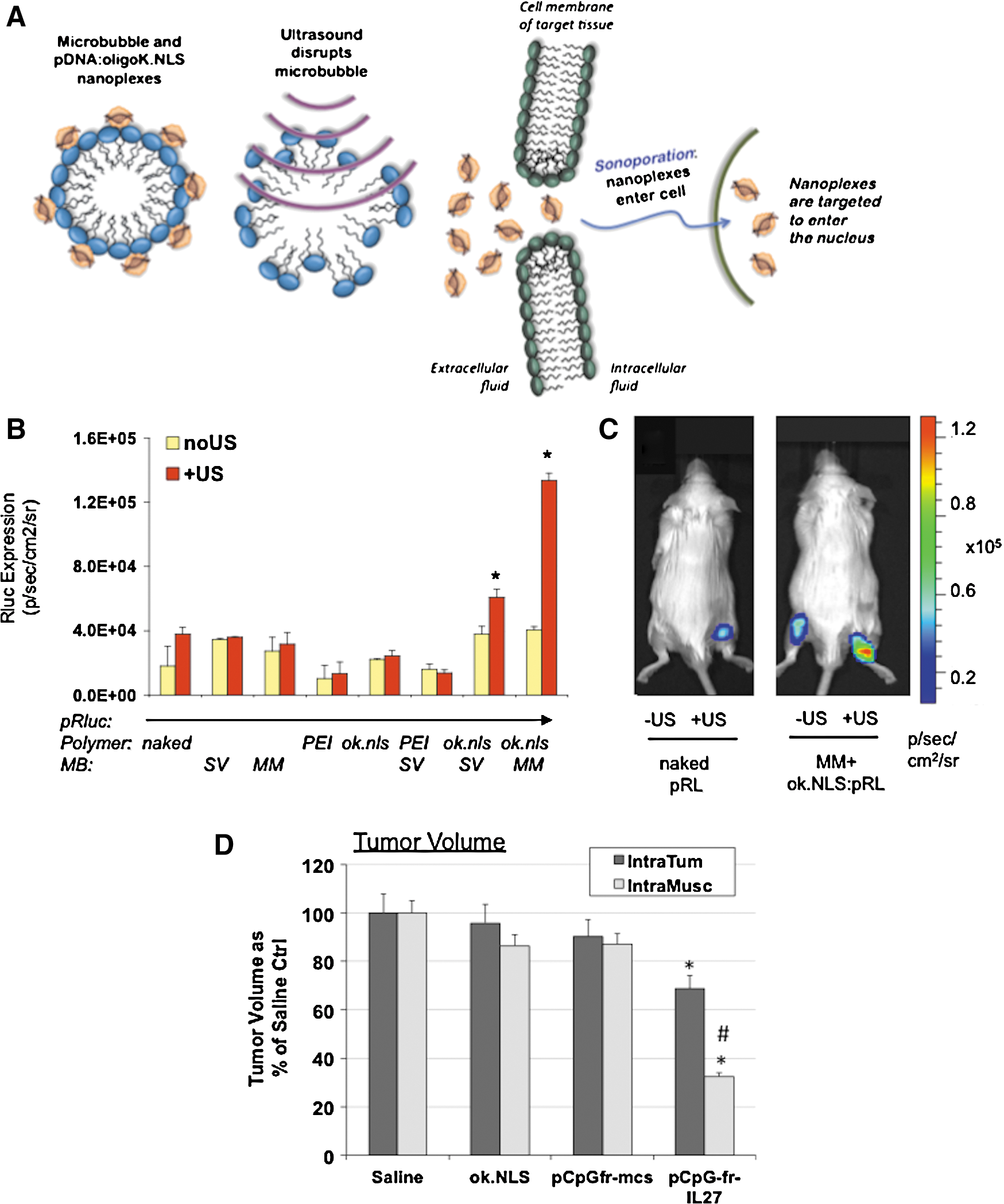
Route of administration of IL-27 gene delivery impacts efficacy in vivo.
Next, we compared the efficiency of I.Tu. and I.M. sonodelivery and observed that the I.M. route was ∼2× as efficient in vivo in reducing tumor volume using a subcutaneous prostate tumor model as proof of concept (Fig. 3D). Because of the higher efficacy of the gene delivery observed, we selected the I.M. route to further examine efficacy of IL-27 gene delivery in treating distant bone metastases of prostate cancer. For the following studies, we developed a new cell line that could better mimic the interaction of bone cells with prostate tumors. The cell line we have developed (TC2R) can form bone lesions representative of the human disease, including osteoblastic and osteolytic reactions in vivo after intratibial implantation into syngeneic C57/BL6 mice. TC2R allows us to explore the full effects of immune therapies for skeletal metastasis of prostate cancer and might be better suited for testing immunotherapy efficacy compared with other available immunecompetent models (RM1, RM1.BM), which are predominantly osteolytic (McCabe et al., 2008; Hung et al., 2011). Also, TC2R may be useful for testing prostate cancer therapies targeting the androgen receptor via drugs or transcriptional-targeted gene therapy (Figueiredo et al., 2007).
Using I.M. sonodelivery, we proceeded to examine whether IL-27 could affect the interaction of tumor and bone compared with delivery of control Rluc vector to mice bearing intratibial 1.5-week TC2R tumors. IL-27 treatment reduced tumor weight by ∼40% and reduced tumor volume by ∼75% (Fig. 4A). We monitored the effect of IL-27 treatment on bone structure and density using two-dimensional μCT (Fig. 4B). IL-27-treated mice had significantly less bone degradation over time compared with control mice treated with Rluc plasmid. Also, quantification of the bone density over time showed that IL-27 prevented bone loss (p<0.02, Fig. 4C). The bone density loss in controls was sharp, going from ∼1400 to ∼100 HU within 4 weeks. In contrast, the IL-27 treatment prevented this density decline, with an ∼4-fold slower rate of density loss. Three-dimensional μCT analyses showed that the extent of bone degradation as well as uneven ossification patterns (rounded protrusions) was very high in control animals bearing TC2R tumors by week 3 compared with control baseline (Fig. 4B). In contrast, IL-27-treated animals displayed a significant improvement in bone surface appearance (Fig. 4B), which was more uniform compared with controls, and bone density (Fig. 4C). μCT thus allowed us to qualitatively and quantitatively examine bone changes in response to the IL-27 sonodelivery strategy in vivo. Histological examination showed that IL-27 therapy improved osteoblastic immature bone lesions compared with Rluc control (Fig. 4D), with a significant normalization of bone toward control levels (Fig. 4E).
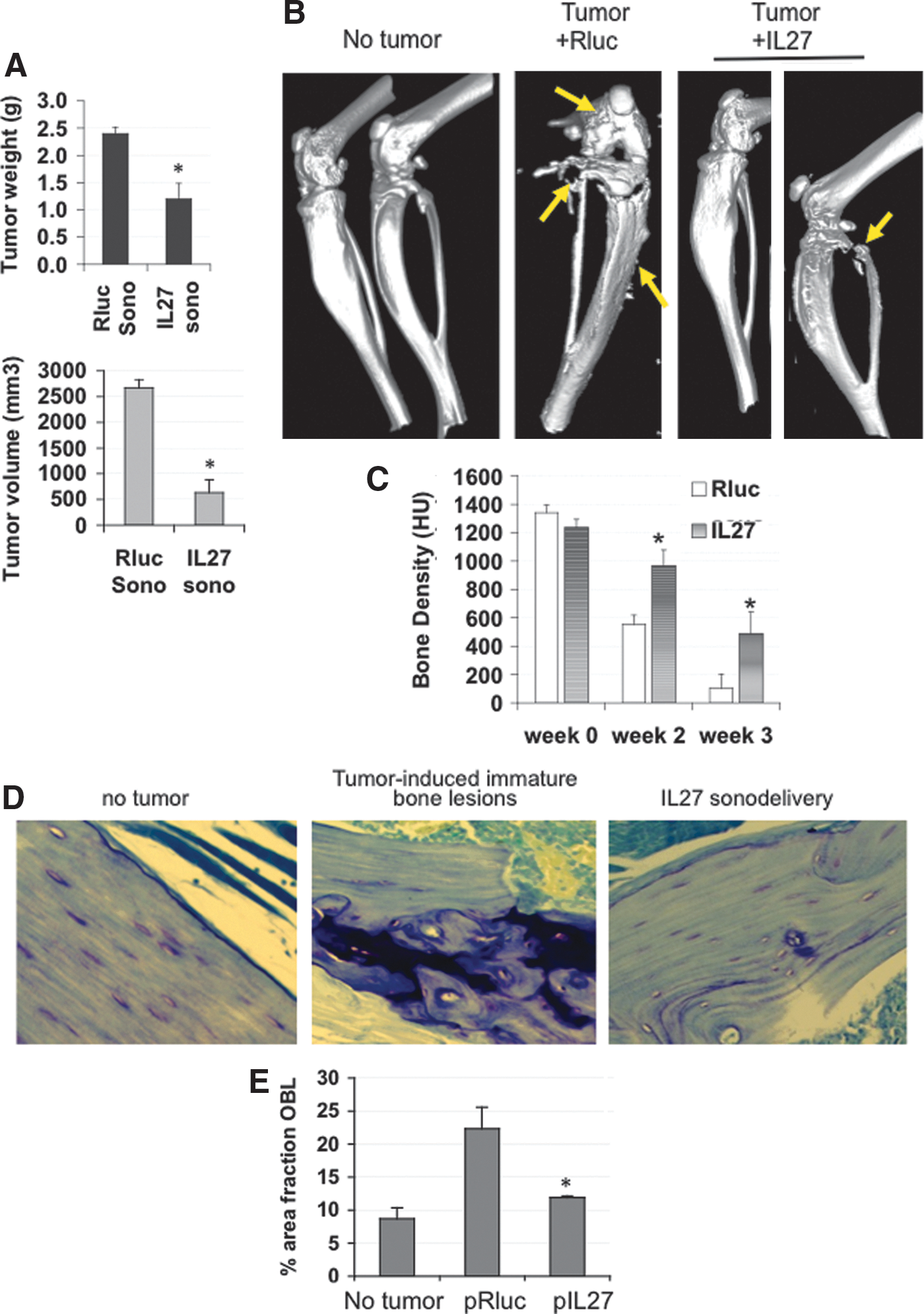
In vivo intramuscular IL-27 gene delivery reduces tumor size and bone resorption. Shown are three-dimensional micro-computed tomography (μCT) images for the tibial bones of C57/BL6 mice implanted with TC2Ras cells (106).
IL-27 sonodelivery enhances the accumulation of effector cells in prostate tumors
We had compared two routes of sonodelivery (Fig. 3D) and had shown that the I.M. IL-27 delivery was more effective than the I.Tu. route in inhibiting tumor growth. To continue to characterize and determine potential mechanisms for this observation, we examined levels of IL-27 and its target genes in the target tissue (tumors) or the site of sonodelivery (muscle). In tumors, I.M. IL-27 sonodelivery induced 2–4-fold higher levels of IL-27 subunits (p28 and EBI3), and the cytokine's target genes T-bet and IFNγ, as compared with I.Tu. as assessed by qPCR (Fig. 5A). At the site of sonodelivery (muscle), I.M. IL-27 sonodelivery also induced IL-27p28, EBI3, and T-bet (although at lower levels than in tumor), but interestingly, IFNγ levels were unchanged. We hypothesized that these differences might indicate a mechanism by which I.M. IL-27 promotes infiltration of immune effector cells into tumors. These effectors would act to further augment IL-27 signaling through a feedforward loop of transcriptional activation of p28 and EBI3 via IFNγ upregulation of IRF1/4, as we have reported (Zolochevska et al., 2011). When we compared the levels of IL-27 in serum between groups treated with IL-27 I.M. and I.Tu., we observed that IL-27 levels were higher following the I.M. sonodelivery route (Fig. 5C). FACS data utilizing immune cells isolated from TC2R tumors suggest that efficacy of IL-27 sonodelivery might be related to an enhanced accumulation of NKT and/or γδT cells, and this is observed with either sonodelivery route used (Fig. 5D). Interestingly, γδT cells were significantly upregulated (∼3-fold) in tumors treated with I.M. IL-27 sonodelivery (Fig. 5D) as compared with the I.Tu. route. These results suggest a role for γδT effectors in mediating the effects of I.M. IL-27 sonodelivery in vivo.

Modulation of immune effector cell accumulation in TC2Ras subcutaneous prostate tumors by IL-27 intramuscular sonoporation gene delivery in vivo.
Discussion
The application of a single “osteoimmune” cytokine that could simultaneously treat metastatic lesions while restoring bone homeostasis makes IL-27 a very attractive therapeutic candidate for prostate cancer. We have previously shown that IL-27 has antitumor effects on prostate cancer in vivo (Zolochevska et al., 2011), an effect supported by other tumor models (Yoshimoto et al., 2008; Cocco et al., 2010; Zhu et al., 2010). In this context, the present work extends the use of IL-27 in reducing osteoclastogenesis and in enhancing OB differentiation when present in coculture with prostate tumor cells. This type of experimental design was chosen as a suitable model for mimicking in vivo applications, where the tumor interacts actively with the bone microenvironment. For preclinical testing, the tumor is typically the primary target of gene delivery/therapy, and the cytokine expressed is provided to the bone microenvironment in a paracrine manner. We thus examined the effect of paracrine IL-27 on bone cells of different differentiation stages. This design is relevant to assessing the different effects of IL-27 on immature (Roudier et al., 2004, 2008) or mature bone cells. We also wanted to assess whether autocrine IL-27 (expressed directly in bone cells) could modify proliferation or differentiation of OB. Paracrine IL-27 enhanced the proliferation of maturing OB, augmented OB differentiation, and inhibited OC proliferation. Autocrine IL-27 expression further promoted the proliferation of mature OB. For OC, paracrine IL-27 reduced OC proliferation. Autocrine IL-27 further reduced the OC proliferation, suggesting that combined IL-27 delivery to both OC and prostate cancer cells appears to help achieve higher cytokine efficacy against OC proliferation. Regarding effects on bone cell differentiation, IL-27 also was able to promote pre-OB differentiation even in the absence of differentiation supplements, and augmented differentiating OB mineralization, suggesting that IL-27 can directly induce and augment osteogenesis. Both autocrine and paracrine IL-27 were able to inhibit osteoclastogenesis by the potent inducing agent RANKL, suggesting promising translational applications of this molecule.
Regarding IL-27 therapy delivery in vivo, we selected a method that utilizes clinically safe US frequencies to induce cellular cavitation and deliver plasmid DNA via sonoporation for gene delivery (sonodelivery) (Zolochevska and Figueiredo, 2012). We have observed that this is an efficient method for gene delivery that approximates the efficiency of adenovirus (Zolochevska et al., 2011). In the present report, we devised a strategy for I.M. sonodelivery since we needed to treat the bone tumor lesions and these are difficult to reach via I.Tu. injections. The muscle is an excellent site of gene administration because of long-term gene expression and was shown to successfully deliver IL-27 plasmid in vivo by electroporation in colon and breast tumors (Zhu et al., 2010). In the current report, we optimized several I.M. sonodelivery conditions using reporter gene plasmids, finding that the best approach consisted of complexing pDNA with a novel cationic polymer, NLS2, in the presence of sonoporation assisted by microbubbles (Zolochevska et al., 2011). This novel polymer leads to a transfection efficiency higher or comparable to JetPEI and in some cases, Lipofectamine 2000 (Parelkar et al., 2011). Ours is the first use of the novel DNA:NLS2 polyplex in vivo. We observed efficient gene delivery in muscle and a reduced rate of bone loss in mice bearing TC2R intratibial tumors. These results are promising, since IL-27 was able to slow the rate of bone destruction and inhibit tumor growth within bone. However, one limitation lies in the partial efficacy, whereby some tumors still grew, although at a ∼75% slower rate. The IL-27 I.M. delivery appears to reduce tumor-induced immature osteoblastic lesions, suggesting that IL-27 normalizes bone tissue and restores its homeostasis. Our data indicate that both I.Tu. and I.M. IL-27 sonodelivery mediate significant recruitment of infiltrating γδT and NKT cells to subcutaneous TC2R tumors. Interestingly, I.M. IL-27 sonodelivery recruited ∼3-fold as many γδT cells to the subcutaneous tumor, suggesting that this could be one mechanism for the different efficacy observed. This difference may be related to the higher expression levels of IL-27 found in tumors of animals treated with I.M. sonoporation, although the mechanism for this elevated expression pattern remains uncharacterized at this time. Regarding effector cells recruited to tumors, we have previously reported the antitumor effects of IL-27 on prostate cancer through enhancement of CD8+ T cells, reduction in Treg, and myeloid-derived suppressor cells in tumors in vivo after gene delivery (Zolochevska et al., 2011). rIL-27 has been recently shown to enhance γδT cytotoxicity (Morandi et al., 2012). γδT cells recognize and kill tumor cells in a tumor antigen-independent manner, therefore, these are an interesting and relatively novel effector cell type that may provide protective immunosurveillance against metastatic tumors (Dieli et al., 2007, 2008). Future studies will include examination of the potential infiltration of other organs by γδT effector cells, although we have not observed any significant lymphocytic infiltration in organs other than tumors, even when deliverying IL-27 at a higher DNA dose (45 μg) (Zolochevska et al., 2011). However, such studies will be critical to determining toxicity potential for this therapeutic modality.
We also characterized the potential of IL-27 therapy and its impact on tumor growth as well as bone density. We chose μCT for examining structural bone changes since this technique has several advantages, including high spatial resolution and contrast for imaging mineralized tissues in small animals (Freeman et al., 2009; Morgan et al., 2009; Campbell et al., 2011; Johnson et al., 2011). μCT allows longitudinal monitoring of tumor-associated bone destruction in mouse models and also detection of newly ossified bone in prostate cancer models. However, although promising, the IL-27 therapeutic response was partial; therefore, we propose that strategies still must be devised to enhance IL-27 expression and secretion after I.M. sonodelivery, as well as strategies to enhance retention of IL-27 at bone metastases in order to achieve an efficient and complete therapeutic effect. We have observed that a variety of normal cell types appear to tolerate treatment with IL-27 at relatively high doses in vitro without significantly losing viability (Supplementary Fig. S1), suggesting that the dose delivered can be increased further, and future experiments in vivo will assess the exposure limit of persistent IL-27 after sonodelivery to help characterize any potential toxicity. Other strategies by which the therapy efficacy can be augmented are by utilizing cytokine combinations proven successful for other preclinical studies, such as IL-27+IL-12 (Zhu et al., 2010) or IL-27+IL-2 (Salcedo et al., 2009), and ligand-mediated targeting strategies in order to enhance cytokine efficacy via increased retention within tumors [reviewed by Corti et al. (2011)].
Conclusions
We describe a novel gene delivery strategy that is simple to perform, is cost-effective, and is clinically applicable, since US-mediated gene delivery or sonodelivery can be accomplished using any conventional clinical US apparatus. Our findings are promising for I.M. delivery of a novel osteoimmune cytokine, IL-27, which shows great promise for simultaneously treating tumor and bone. Our results are promising since there are no current therapies that can successfully achieve these outcomes for treating patients with bone metastatic prostate cancer.
Footnotes
Acknowledgments
We thank Dr. Lawrence Sowers and the Department of Pharmacology & Toxicology for support of a bioluminescence optical imaging core and the Center for Bioengineering for support of the Small Animal Imaging core. We thank the UTMB Histopathology Core for assistance with tissue processing and microscopy access and the UTMB Flow Cytometry Core for assistance with cell-staining procedures. We thank Dr. Christopher Suckow from Animal Resources Center at UTMB for expert assistance with fasting and animal care and Dr. Shulin Li from UTMD Anderson Cancer Center for scientific discussions relating to this work. This work was funded in part by award NSF-DMR-BMAT-1207775 to T.E.
Author Disclosure Statement
No competing interests exist for the authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.