
Abstract
Transforming growth factor-β1 (TGF-β1) has been shown unequivocally to enhance neointima formation in carotid and ileo-femoral arteries. In our previous studies, however, TGF-β1 expression in coronary arteries actually reduced neointima formation without affecting luminal loss postangioplasty, while expression of a TGF-β1 antagonist (RIIs) in balloon-injured coronary arteries reduced luminal loss without affecting neointima formation. These observed effects may be a consequence of the mode of coronary artery gene transfer employed, but they may also represent differences in the modes of healing of coronary, carotid, and ileo-femoral arteries after endoluminal injury. To help clarify whether a gene therapy strategy to antagonize TGF-β might have application within the coronary vasculature, we have investigated the effect of high-level periluminal expression of RIIs using stent-based adenovirus-mediated intracoronary gene transfer. Porcine coronary arteries were randomized to receive a custom-made CoverStent preloaded with saline only, or with 1×109 infectious units of adenovirus expressing RIIs or β-galactosidase (lacZ). Vessels were analyzed 28 days poststenting, at which time angiographic in-stent diameter was significantly greater in RIIs-treated arteries, and in-stent luminal loss significantly reduced. Computerized morphometric minimum in-stent lumen area was ∼300% greater in RIIs-exposed vessels than in lacZ or saline-only groups. This was because of significantly reduced neointima formation in the RIIs group. RIIs had no demonstrable effect on cellular proliferation or apoptosis, but greater normalized neointimal/medial collagen content was observed in RIIs-exposed arteries. These data highlight the qualitatively similar effect of TGF-β antagonism on neointima formation in injured coronary and noncoronary arteries, and suggest that since cellular proliferation is unaffected, TGF-β1 antagonism might prevent in-stent restenosis without the delayed healing that is associated with drug-eluting stents in current clinical use.
Introduction
W
All other studies of antagonism of TGF-β in the coronary arteries have failed to demonstrate reduced neointima formation (Paiement et al., 1998; Chung et al., 2002b, 2010). These observations differ from studies of TGF-β1 in noncoronary vessels, where TGF-β1 increased neointima formation within normal (Nabel et al., 1993; Schulick et al., 1998) and injured (Kanzaki et al., 1995) arteries, while antagonists of TGF-β1 reduced neointima formation (Wolf et al., 1994; Smith et al., 1999; Yamamoto et al., 2000; Ando et al., 2004; Ikeda et al., 2004; Yao et al., 2009). The apparent discrepancy between observations made in coronary arteries and those made in other vessels might be the consequence of the location of transgene expression: the Infiltrator catheter (used by us and by Chung et al.) delivers material principally to the adventitia (Kingston et al., 2001, 2003; Chung et al., 2002b), while studies of TGF-β1 antagonism in noncoronary arteries have used endoluminal routes that deliver antagonists to the luminal aspect of the artery (Wolf et al., 1994; Smith et al., 1999; Yamamoto et al., 2000; Ando et al., 2004; Ikeda et al., 2004; Yao et al., 2009). Other possibilities might account for these observations, however. Coronary smooth muscle cells (SMC) are phenotypically distinct from SMC of other arteries (Christen et al., 1999; Patel et al., 2000); they display changes in proliferation, migration, and the expression of differentiation markers on exposure to TGF-β1 that differ from those observed in noncoronary arterial SMC (Christen et al., 1999). This has led to the suggestion that the mechanism of vascular repair may differ between coronary and noncoronary vessels (Patel et al., 2000). Significant differences also exist between the mechanisms of noncoronary balloon injury and of percutaneous transluminal coronary angioplasty (PTCA) (Lafont and Faxon, 1998). In keeping with this, the mean I:M ratio in control vessels in our previous studies (1.9±0.3) was substantially greater than those reported in early studies of injured noncoronary arteries (range 0.3–1.0) (Wolf et al., 1994; Kanzaki et al., 1995; Smith et al., 1999; Yamamoto et al., 2000; Ikeda et al., 2004), raising the possibility that the high-pressure, low-stretch injury of PTCA represents a greater stimulus to neointima formation than the low-pressure, high-stretch forms of injury used in noncoronary vessels. The difference between the reported effects of TGF-β antagonists on neointima formation in coronary and noncoronary arteries might have arisen, therefore, because of a combination of different injury methods and different healing mechanisms within the coronaries by comparison with other arteries.
The large majority of clinical coronary interventions now involve stent deployment. In-stent restenosis (ISR) is characterized by even greater enhancement of extracellular matrix (ECM) accumulation than is seen after simple PTCA, with increased neointimal expression of TGF-β1 (Chung et al., 2002a). Chung et al. studied the effects of delivery of a virus analogous to Ad5-RIIs into pig coronaries immediately before stenting (Chung et al., 2010). No effect of secreted TGF-β type II receptor expression on neointima formation was observed; however, use of the Infiltrator for virus delivery means that low-level periluminal transgene expression may account for the lack of effect in this study too. Fortunately, stents represent a convenient platform for endoluminal coronary gene transfer; we have previously used a collagen-membrane-bound stent (the CoverStent; a custom-made investigational stent produced by Medtronic) to deliver adenovirus to the periluminal coronary media (Salem et al., 2006). In contrast to adventitial fibroblasts, however, vascular SMCs are difficult cells in which to elicit high-level transgene expression (Clesham et al., 1996). In order to maximize periluminal transgene expression, we have developed an expression cassette (the PREP cassette) that elicits 90-fold greater transgene expression in coronary SMCs than the regulatory elements present in Ad5-RIIs (Appleby et al., 2003). We have used CoverStents loaded with Ad5-PREP-RIIs to investigate the effect on in-stent neointima formation of antagonism of TGF-β1 by RIIs expression in the periluminal layers of the coronary artery.
Materials and Methods
Generation of recombinant adenoviruses
Viruses were generated by homologous recombination in 293 cells. Ad5-RIIs contains the cDNA for a secreted human TGF-β type II receptor (RIIs) under transcriptional control of the major immediate-early enhancer/promoter from human cytomegalovirus. Ad5-PREP-RIIs and Ad5-PREP-lacZ contain the cDNA for RIIs and gene for Escherichia coli β-galactosidase, respectively, within the PREP cassette (Appleby et al., 2003). Stocks were purified by cesium-chloride gradient ultracentrifugation and titrated by serial-dilution end-point assay as previously described (Kingston et al., 2001, 2003).
In vitro assay of RIIs activity
Human coronary SMC (TCS Cellworks) at 90% confluence were infected at multiplicity of infection of 100 with one of the three viruses studied. Forty-eight hours postinfection, conditioned media were collected and assayed for TGF-β1 antagonist activity: mink lung epithelial cells stably transfected with the cDNA for luciferase under control of a TGF-β-responsive promoter [MLEC(PAI/L), provided by Dr. D.B. Rifkin, New York University School of Medicine] were cultured as described previously (Kingston et al., 2001, 2003). Conditioned media from infected SMC were supplemented with human TGF-β1 to final concentrations of 0.01, 0.1, and 1.0 ng/ml. Exactly 500 μl of each supplemented medium was added to 24-well plates seeded with MLEC(PAI/L). After overnight incubation, luciferase activity and protein content in cell lysates from each well were assayed.
CoverStent deployment and adenovirus delivery
Forty-four Large White pigs (20–27 kg) underwent coronary stenting. All protocols were approved by the University of Manchester Animal Ethical Review Group and authorized by the UK Home Office. Animals received 150 mg of aspirin daily, commencing 24 hr prestenting and continuing until the vessels were harvested. Thirty-six animals were randomized to receive 1×109 infectious units (iu) of Ad5-PREP-RIIs, 1×109 iu of Ad5-PREP-lacZ, or phosphate buffered saline (PBS) only, loaded onto a CoverStent (Fig. 1a) immediately preprocedure. Anesthesia was induced by inhalation of 4% isoflurane, and an endotracheal tube was inserted. Adequacy of anesthesia was confirmed by continuous ECG and pulse-oximetric monitoring. Coronary angiography was performed in left anterior oblique and right anterior oblique views using an OEC 9600 digital image intensifier as described previously (Kingston et al., 2003). Segments in the left anterior descending artery (diameter 2.1–2.7 mm) were selected for stenting by an operator blinded to the randomization. CoverStents (3.0 mm×15 mm) were deployed by balloon inflation to 12 atmospheres for 30 sec. Postdeployment angiograms were acquired and in-stent angiographic diameter was measured by the blinded operator. Animals received 200 μg of intramuscular buprenorphine before recovery from anesthesia.
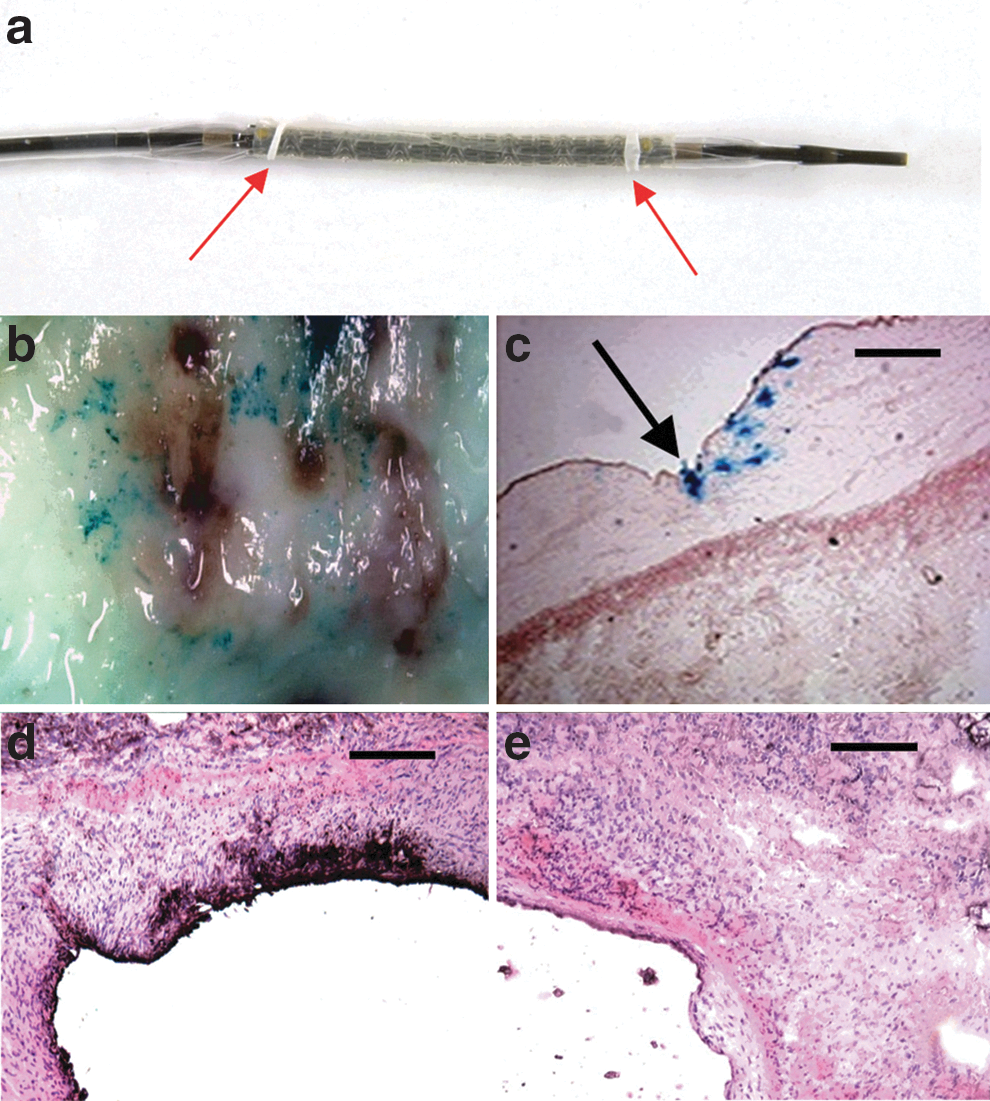
Transgene expression 72 hr after CoverStent-mediated intracoronary gene transfer. The CoverStent
Eight more animals received a CoverStent prepared with 1×109 iu Ad5-PREP-RIIs or 1×109 iu Ad5-PREP-lacZ. Two were euthanized 72 hr postdeployment for assessment of transgene expression; the remaining six were euthanized at 7 days for mechanistic assessment: general anesthesia was induced as previously and animals were euthanized with intravenous pentobarbitone (20 mg/kg). The 36 animals randomized for morphometric analysis were euthanized 28 days poststenting: general anesthesia was induced and angiograms were performed as described above. Minimum in-stent lumen diameter was measured by a blinded operator before animals were euthanized with intravenous pentobarbitone. Angiographic data were analyzed from those animals that survived to 28 days and whose stented vessel segments were suitable for morphometric analysis.
Transgene expression and mechanistic assessment
Stented vessel segments used for mechanistic studies were dissected out and opened longitudinally before the stent was carefully removed. A short length of the 72 hr Ad5-PREP-lacZ-infected artery was reserved for en face examination. The remaining 72 hr Ad5-PREP-lacZ-infected segment and the 72 hr Ad5-RIIs-infected segment were embedded in OCT and frozen in liquid nitrogen before cryostat sectioning. β-Galactosidase expression was assessed by the previously described X-gal staining method (Appleby et al., 2003). RIIs expression was assessed immunohistochemically using antibody to human TGF-β type II receptor. Sections were counterstained with hematoxylin and eosin.
All 7-day segments were embedded in OCT and frozen in liquid nitrogen before cryostat sectioning. Cellular proliferation, apoptosis, and inflammatory infiltration were assessed immunohistochemically using anti-PCNA, anti-activated-caspase-3, and anti-CD45 primary antibodies, respectively. Proportional areas of vessel wall staining with each antibody (corrected for area of staining in controls exposed to secondary antibody only) were quantified using Leica QWin software and a Quantimet 600S digital analysis system as previously described (Kingston et al., 2001). Fibrillar collagen content was assessed as the area of red birefringent staining in picrosirius red-stained segments using QWin and a Quantimet 600S digital analysis system as previously described (Kingston et al., 2001, 2003). Levels of biologically active TGF-β were assessed by modified MLEC(PAI/L) assay (Ranjzad et al., 2009): 20-μm-thick cryostat sections from each Ad5-PREP-RIIs- and Ad5-PREP-lacZ-infected artery were added to the wells of a 24-well plate preseeded with MLEC(PAI/L). Luciferase expression in lysates of the MLEC(PAI/L) monolayer was assessed luminometrically 16 hr later and corrected for the volume of the section added to each well.
Morphometric assessment
Arteries harvested at 28 days poststenting were pressure-fixed in situ with 4% formaldehyde. Stented segments were excised, further fixed overnight, and embedded in LR White resin. Embedded vessels were cut into 100-μm-thick blocks. Five-micrometer sections were cut from each block, stained with Millers elastic stain, and examined using QWin and a Quantimet 600S digital analysis system. Vessels were excluded from analysis if the animal did not survive to 28 days and if no collagen membrane was visible around the stent on microscopic inspection (Fig. 3).
Lumen area was measured in all sections, by an operator blinded to the group to which each vessel had been allocated, to determine minimum luminal area (MLA) within each stent. The following areas were then measured in the segment with MLA from each vessel: (1) area within the internal elastic lamina (IEL); (2) area within the external elastic lamina (EEL); (3) area within the stent struts, and (4) area of the collagen membrane. The mean injury score around the stent struts was calculated using the method of Gunn et al. (2002). Neointimal and medial areas were calculated as stent area minus lumen area, and EEL area minus IEL area, respectively.
Statistical methods
Data from the in vitro assessment of RIIs activity, morphometric data, and angiographic data were analyzed by one-way ANOVA and Newman–Keuls post-test. Mechanistic data in which only two groups were compared (Ad5-PREP-RIIs- and Ad5-PREP-lacZ-infected vessels) were analyzed by two-tailed T-test. Analysis was performed using GraphPad Prism software. Data are presented as mean±SEM.
Results
Antagonism of TGF-β1 by Ad5-PREP-RIIs
Conditioned medium from Ad5-RIIs- and Ad5-PREP-RIIs-infected SMC inhibited TGF-β1-mediated induction of luciferase in MLEC(PAI/L) at all concentrations by comparison with Ad5-PREP-lacZ (see Supplementary Fig. S1; Supplementary Data are available online at
Transgene expression after CoverStent-mediated adenovirus delivery
A CoverStent is shown in Fig. 1a, immediately before virus loading. En face examination of the X-gal-stained segment of Ad5-PREP-lacZ-infected coronary artery revealed punctate blue staining on the luminal surface (Fig. 1b) confined to the stented segment of vessel. X-gal staining of cryostat sections confirmed β-galactosidase expression on the luminal surface and in the periluminal media, where staining was most marked in the region of the stent struts (Fig. 1c). No staining with X-gal was seen in the Ad5-PREP-RIIs-infected vessel. Immunohistochemistry revealed intense staining for RIIs within the periluminal media and at the luminal surface of the Ad5-PREP-RIIs-infected vessel (Fig. 1d). No periluminal staining for RIIs was observed in Ad5-PREP-lacZ-infected vessels (Fig. 1e).
Effect of Ad5-PREP-RIIs on luminal loss and in-stent neointima formation
Angiographic diameters pre- and immediately poststent deployment, and balloon:artery ratios did not differ significantly between groups (Table 1). Minimum angiographic in-stent luminal diameter at 28 days was greater (p=0.002) in RIIs-treated vessels than in control groups (Fig. 2a). Concomitantly, mean angiographic luminal loss in the RIIs group was reduced by 41% (p<0.001) compared with the control groups (Fig. 2b).
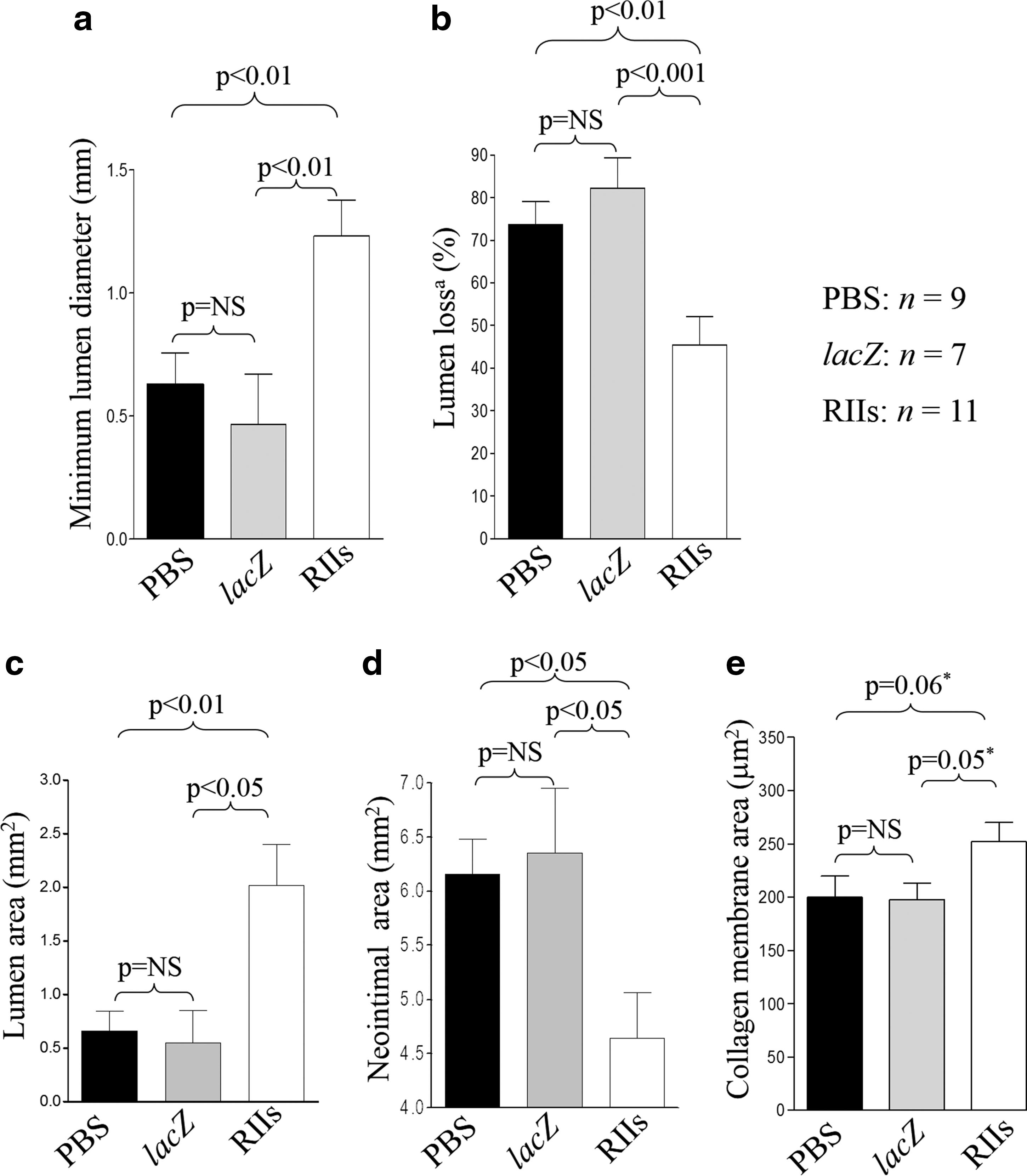
Angiographic in-stent parameters and morphometric parameters at the site of minimum lumen area 28 days poststenting.
PBS, phosphate buffered saline; RIIs, a TGF-β1 antagonist.
Values are presented as mean±SEM.
Nine arteries from the PBS-only group, 7 Ad5-PREP-lacZ-infected vessels, and 11 from the Ad5-PREP-RIIs group were suitable for morphometric analysis. The area bounded by the stent struts, the IEL area, EEL area, and the injury score at the site of MLA did not differ significantly between groups (Table 1). The MLA was ∼300% greater (p=0.004) in the Ad5-PREP-RIIs group than in either control group (Figs. 2c and 3a–c). The greater MLA in Ad5-PREP-RIIs-treated vessels was associated with a significantly smaller neointimal area (p=0.018; Fig. 2d).
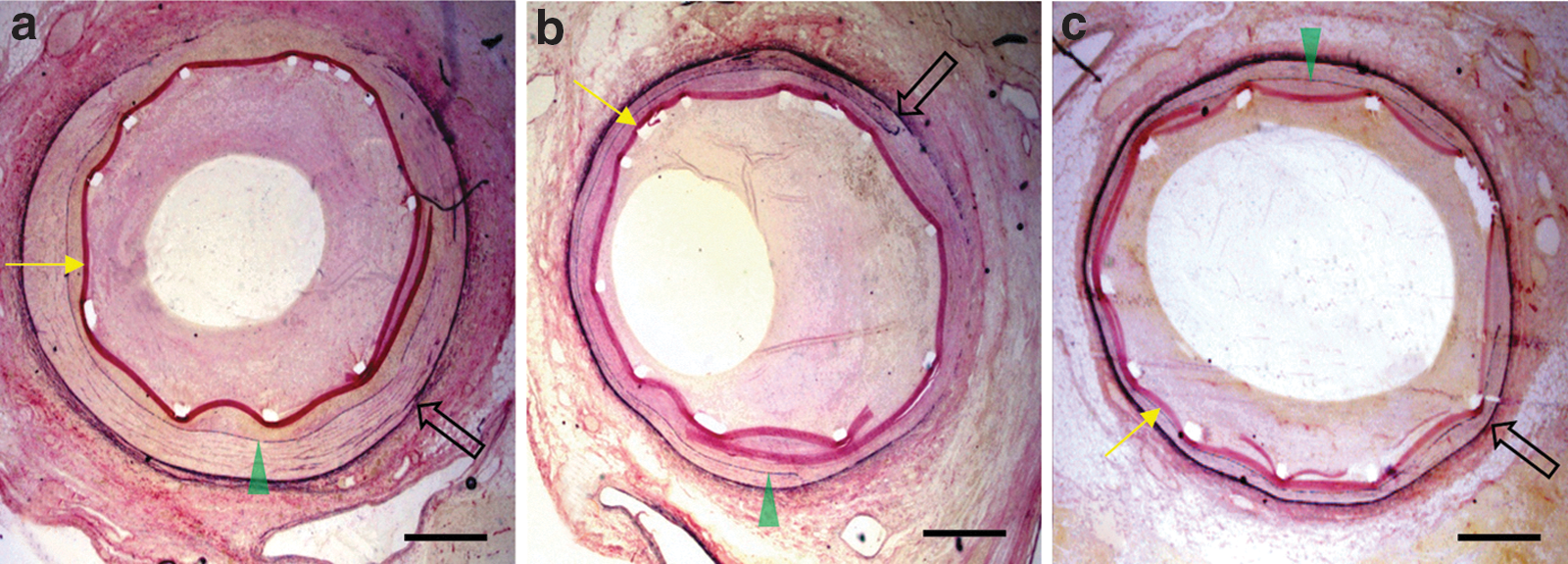
Coronary arteries 28 days after CoverStent deployment. Representative Millers Elastic–stained sections of coronaries at the site of minimum luminal area 28 days post-CoverStent deployment:
The mean cross-sectional area of the collagen membrane in the Ad5-PREP-RIIs group (252±18 μm2) was greater than that of the PBS (200±20 μm2) and Ad5-PREP-lacZ (197±15 μm2) groups (Fig. 2e). This difference was not quite statistically significant by one-way ANOVA (p=0.065). Comparison of the membrane area in the Ad5-PREP-RIIs group with those in the PBS and Ad5-PREP-lacZ groups by t-test revealed p-values of 0.06 and 0.05, respectively.
The Ad5-PREP-lacZ and PBS groups did not differ significantly from each other with respect to any of the angiographic or morphometric parameters assessed.
Mechanism of Ad5-PREP-RIIs-mediated reduction of in-stent neointima formation
Luciferase expression in MLEC(PAI/L) exposed to sections from Ad5-PREP-RIIs-infected coronaries was 63% lower (p=0.04) than that in cells exposed to Ad5-PREP-lacZ-infected sections (Fig. 4a), confirming a reduction in biologically active TGF-β in Ad5-PREP-RIIs-infected arteries. Immunohistochemical staining for CD45 was 30% less (p=0.016) in Ad5-PREP-RIIs-infected arteries compared with Ad5-PREP-lacZ-infected arteries (Fig. 4b–d). No difference was observed in PCNA staining or activated caspase-3 staining between groups (data not shown).
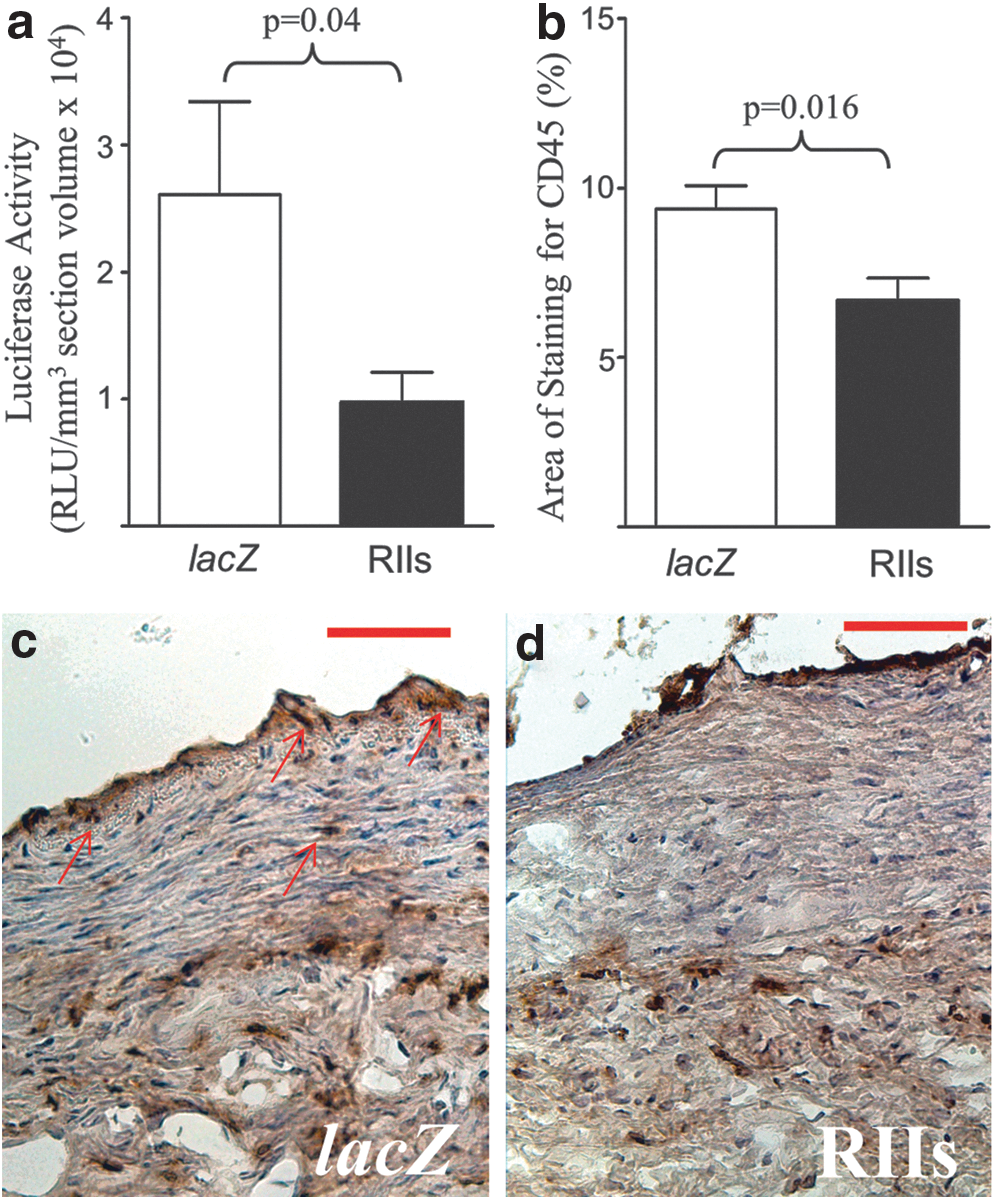
Vessel wall TGF-β activity and inflammatory infiltration 7 days poststenting.
The total area of picrosirius red staining (a direct measure of fibrillar collagen content) in the combined neointima+media did not differ between groups. However, the combined area of neointima+media at 7 days poststenting was greater in Ad5-PREP-lacZ-infected vessels (1877±87 μm2) than in Ad5-PREP-RIIs-infected vessels (1087±107 μm2; p<0.0001). When the area of picrosirius red staining was represented as a proportion of the total area of the neointima+media, the normalized picrosirius red content in neointima+media was found to be 46% lower (p=0.03) in Ad5-PREP-lacZ-infected arteries than in the Ad5-PREP-RIIs group (Fig. 5a–d).
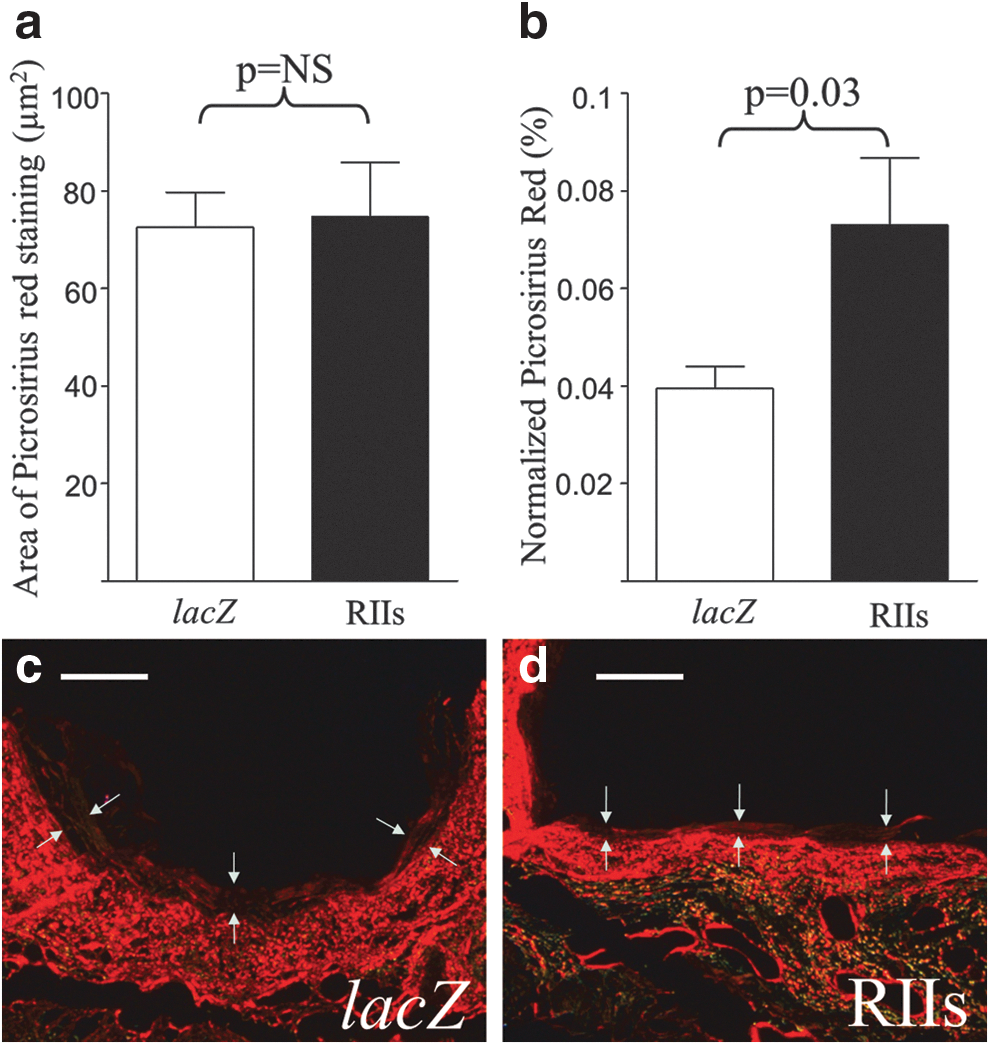
Collagen content 7 days poststenting.
Discussion
Previous studies of the effects of exogenous TGF-β1, or of antagonism of endogenous TGF-β on neointima formation following endoluminal vascular injury, have shown some discrepancy between outcomes in coronary arteries and other noncoronary injury models. In the present study we demonstrate that a transgenic antagonist of TGF-β1 expressed in the periluminal wall of stented coronary arteries suppresses neointima formation and sustains luminal patency 28 days poststenting. In doing so, we confirm that the effect of RIIs on the response of coronary arteries to endoluminal injury is qualitatively similar to the previously reported effects of other antagonists of TGF-β in injured carotid and ileo-femoral arteries (Wolf et al., 1994; Smith et al., 1999; Yamamoto et al., 2000; Ando et al., 2004; Ikeda et al., 2004; Yao et al., 2009).
ECM constitutes the majority of neointima in vessels that have undergone balloon angioplasty or stenting (Christen et al., 2001), and TGF-β1 is the principal mediator of ECM synthesis by SMC and fibroblasts. It is not entirely surprising that antagonism of TGF-β1 inhibits neointima formation in injured vessels. Nonetheless, although most in vivo studies of the role of TGF-β in the vascular response to injury have suggested that TGF-β1 enhances neointima formation, two studies have shown the opposite effect. As mentioned previously, we noted that adventitial expression of TGF-β1 reduced neointima formation after PTCA (Kingston et al., 2003). It seems probable that the mechanism underlying this effect is disruption of the migration of adventitial myofibroblasts to the luminal aspect of the coronary artery. Unfortunately, Infiltrator catheters have been unavailable for acquisition for several years, which means that we are unlikely ever to be able to test this hypothesis.
In addition to our observations, Smad3-knockout mice, in which TGF-β signaling is impaired (but not completely disrupted), demonstrated enhanced neointima formation compared with wild-type mice after vascular injury (Kobayashi et al., 2005). These studies suggest that, under some specific circumstances, TGF-β may have an inhibitory effect on neointima formation. In coronary arteries, the evidence presented here supports a stimulatory role for TGF-β in in-stent neointima formation. This could be further ratified by stent-mediated delivery of TGF-β1-expressing adenovirus to the coronary vasculature; however, as demonstrated by both angiographic and morphometric data, the CoverStent elicits substantial neointima formation. It is likely that delivery of a proneointimogenic transgene from the CoverStent would result in severe compromise of vessel patency and significantly reduced survival to 28 days. An alternative stent-based system would be required for Ad5-TGF-β1 delivery. It is also clear on the basis of the results from our studies and those of Chung et al. (2010) that, even in a structure as insubstantial as the balloon-injured coronary artery, transgene expression must be directed accurately to the appropriate location within the target tissue to achieve the desired therapeutic effect.
TGF-β1 typically promotes apoptosis of cultured SMC (Hishikawa et al., 1999), but under certain conditions protects arterial SMC from apoptosis (Pollman et al., 1999). Likewise, TGF-β1 can stimulate or inhibit proliferation of cultured SMC depending upon culture conditions (Hneino et al., 2009). Despite this, most investigators have reported no effect of TGF-β-antagonism on cellular proliferation or apoptosis at the site of vascular injury (Wolf et al., 1994; Smith et al., 1999; Ando et al., 2004); only one study noted a modest decrease in PCNA expression in treated animals (Yamamoto et al., 2000). In keeping with these observations, we found no evidence that reduced TGF-β activity in Ad5-PREP-RIIs-infected arteries was associated with any effect on cellular proliferation or apoptosis 7 days poststenting. Instead, the effects of Ad5-PREP-RIIs appear to be mediated by suppression of ECM expansion. Most studies that have examined the effect of TGF-β antagonism on vessel collagen content have reported reduced content and/or expression in treated vessels (Smith et al., 1999; Yamamoto et al., 2000; Ikeda et al., 2004; Yao et al., 2009). In contrast, we observed greater normalized collagen content within the neointima and media of Ad5-PREP-RIIs-infected vessels. It seems likely that this observation arises from a decrease in expansion of noncollagenous matrix components in RIIs-treated vessels compared with controls. Limitations upon the quantities of material available for analysis, and cessation of CoverStent development mean that we are unable to do more than speculate upon the alteration in ECM composition in the wall of CoverStented coronary arteries. Studies are ongoing in an attempt to define the changes in ECM expression and content in other models of vascular injury after antagonism of endogenous TGF-β; however, the differences in mode of injury and methods of gene transfer that have been imposed by the unavailability of CoverStents mean that the results of these investigations are not assuredly applicable to the model described herein, and it may never be possible to define the underlying ECM changes in CoverStented coronaries with certainty.
The observed effects upon collagen content are similar in CoverStented arteries exposed to Ad5-PREP- RIIs to our findings following Infiltrator-mediated delivery of Ad5-RIIs post-PTCA, when decreased constrictive remodeling was accompanied by increased adventitial collagen content in RIIs-treated vessels (Kingston et al., 2001). As TGF-β1 has been shown to stimulate expression of MMP-2 and MMP-9, which include collagens I and III as substrates (Overall et al., 1989; Sehgal and Thompson, 1999), increased normalized collagen content in RIIs-exposed vessels might be the consequence of abrogation of TGF-β1-induced expression of MMP-2 and MMP-9 within the neointima of stented arteries. In keeping with this possibility, the mean cross-sectional area of collagen membrane in Ad5-PREP-RIIs-infected vessels was greater than that in the control groups. Admittedly, this difference was not quite statistically significant, but the p-value of 0.065 by ANOVA suggests a trend toward significance in support of the notion that less degradation of fibrillar collagen occurs in the Ad5-PREP-RIIs group.
The CoverStent comprised a standard Medtronic BeStent bound in a membrane of cross-linked type I collagen with covalently linked heparin. We had demonstrated previously that the CoverStent allows vector delivery to the periluminal region of the coronary arterial wall (Salem et al., 2006) and have now demonstrated that subsequent transgene expression is at a therapeutically pertinent level. As demonstrated by both angiographic and morphometric data, however, the CoverStent elicits abundant neointima formation. The BeStent has seen widespread clinical use historically, and did not itself invoke ISR of this magnitude. The accentuated neointimal response is clearly caused by the collagen membrane. In its polymeric form, type I collagen suppresses SMC migration and proliferation (Pickering, 2001); however, type I collagen monomers do not have this effect (Ichii et al., 2001), and degradation products of monomeric type I collagen enhance SMC migration (Stringa et al., 2000). It seems likely that products of degradation of the CoverStent collagen membrane stimulate migration and proliferation of SMC and thereby enhance neointima formation. As noted previously, the mean cross-sectional area of collagen membrane in Ad5-PREP-RIIs-infected vessels was greater than that in the control groups, which supports this hypothesis. In light of the intense neointima formation induced by the CoverStent, the extent to which Ad5-PREP-RIIs reduces neointima formation seems even more impressive. It does mean, however, that the CoverStent is entirely unsuitable for clinical application and further development of the device has ceased.
Inhibition of in-stent neointima formation by RIIs is pertinent to restenosis in the clinical setting: over 80% of SMC derived from coronary endarterectomy specimens displayed altered levels of TGF-β signaling receptors and an enhanced fibrogenic response on exposure to TGF-β1 (McCaffrey et al., 1999). Blockade of TGF-β1 signaling in the presence of such SMC is likely to be of therapeutic benefit. Drug-eluting stents (DES) have not entirely abolished the problem of ISR, and have been associated with increased risk of late thrombotic occlusion. Studies of potential mechanisms of late thrombotic occlusion have revealed delayed healing in DES (Kotani et al., 2006). Stent-based gene transfer offers the prospect of a wide range of therapeutic agents to study as potential means of prevention of ISR. We have achieved “therapeutic” effects without demonstrable suppression of cellular proliferation or enhancement of apoptosis at the site of stenting and these are desirable features if delayed healing in stented vessel segments is to be avoided.
To conclude, we have proved that localized antagonism of TGF-β1 by therapeutic gene transfer represents a means for prevention of ISR. In doing so we have also established that, in qualitative terms, the role of TGF-β in the coronary vascular response to injury is the same as its role in repair of noncoronary vessels. Our findings refute, to some degree, the hypothesis that mechanisms of vascular repair differ between the coronary vasculature and noncoronary vessels. It is interesting, however, that we have consistently demonstrated an increase in normalized collagen content at the site of RIIs expression within coronary arteries, while adenovirus-mediated antagonism of TGF-β in vein segments was accompanied by reduced collagen content (Ranjzad et al., 2009). There is still reason to believe that there are some differences between healing in the coronary arteries and in vessels elsewhere.
Footnotes
Acknowledgments
This work was supported by the British Heart Foundation (PG/02/107/14436 and FS/04/026 to P.A.K.). P.R. was supported by a PhD studentship from the Iranian Ministry of Health and Medical Education. S.J.K. was supported by a scholarship from the National University of Sciences and Technology, Islamabad, Pakistan.
Author Disclosure Statement
All authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.