
Abstract
The epithelial–mesenchymal transition (EMT) has been recognized to occur during embryonic development, fibrosis, and tumor metastasis. Nuclear factor (NF)-κB plays a central role in mediating the inflammation and wound-healing responses during liver fibrogenesis. However, the involvement of NF-κB during EMT in liver cells remains unidentified. To develop a therapeutic approach to EMT during liver fibrosis, we examined the inhibition of transcription factor NF-κB, using a decoy oligodeoxynucleotide (ODN) strategy in liver fibrosis in vitro and in vivo. NF-κB decoy ODN contains consensus binding sequences of the NF-κB-binding site. NF-κB decoy ODN effectively suppresses transforming growth factor-β1-induced EMT in AML12 murine hepatocytes. Liver fibrosis induced by CCl4 administration was suppressed by NF-κB decoy ODN. Furthermore, NF-κB decoy ODN was shown to inhibit the EMT process in fibrotic liver in vivo. This study demonstrates the feasibility of NF-κB decoy ODN treatment for preventing liver fibrosis via EMT processes.
Introduction
L
Most liver injuries (viral, alcoholic, and autoimmune hepatitis) activate acute inflammation and wound-healing responses. When the injury is repetitive or chronic, livertissue is exposed to continuously elevated expression of proinflammatory cytokines and chemokines. Hepatic nuclear factor (NF)-κB is increased by elevated inflammatory changes in various nonparenchymal and parenchymal liver cells. In particular, the induction of NF-κB in hepatocytes (liver parenchymal cells) during liver injury contributes to a chronic inflammatory state (Knittel et al., 1999; Lv et al., 2007). However, whether NF-κB participates in the progression of EMT during liver fibrogenesis still remains unclear.
In general, gene transcription is regulated by two regulatory pathways: chromatin rearrangement and transcription factors. Transcription factors are controlled by direct binding to their binding sites. To inhibit the binding of transcription factors, a decoy strategy is employed to block transcription factor activity via the use of a synthetic double-stranded oligodeoxynucleotide (ODN), which contains a consensus binding sequence. The decoy ODN strategy has been shown to be an effective approach for suppressing specific gene expression both in vitro and in vivo (Morishita et al., 1998; Tomita et al., 2007). In abdominal aortic aneurysms and renal ischemia–reperfusion, the decoy ODN strategy has been reported as a novel therapeutic application for treating these disorders (Cao et al., 2004; Miyake et al., 2006). In addition, we previously demonstrated that Sp1 and chimeric decoy ODN effectively suppressed the expression of fibrosis-related genes in vivo and in vitro (Park et al., 2009; Kim et al., 2013).
In this study, we examined the effect of ring-type NF-κB decoy ODN transforming growth factor (TGF)-β1-induced EMT in vitro and further characterized the inhibitory mechanisms in vivo, using CCl4-induced liver fibrosis.
Materials and Methods
Synthesis of ring-type NF-κB decoy ODN
Decoy ODN was purchased as an HPLC-purified product from Bionics (Seoul, Korea). NF-κB and scrambled decoy ODN sequences were used as follows (the consensus binding sequence is underlined): NF-κB decoy ODN: 5′-GAAATTTCA Scrambled (Scr) decoy ODN: 5′-GAATTCAATTCAGGGTACGGCAAAAAATTGCCGTACCCTGAATT-3′
NF-κB and Scr decoy ODNs were annealed for 8 hr, while the temperature was decreased from 80 to 25°C. These decoy ODNs were predicted to form a stem–loop structure (Fig. 1A). To obtain a covalent ligation for ring-type decoy ODN, each decoy ODN was mixed with T4 ligase (Takara Bio, Otsu, Japan) and incubated for 18 hr at 16°C.
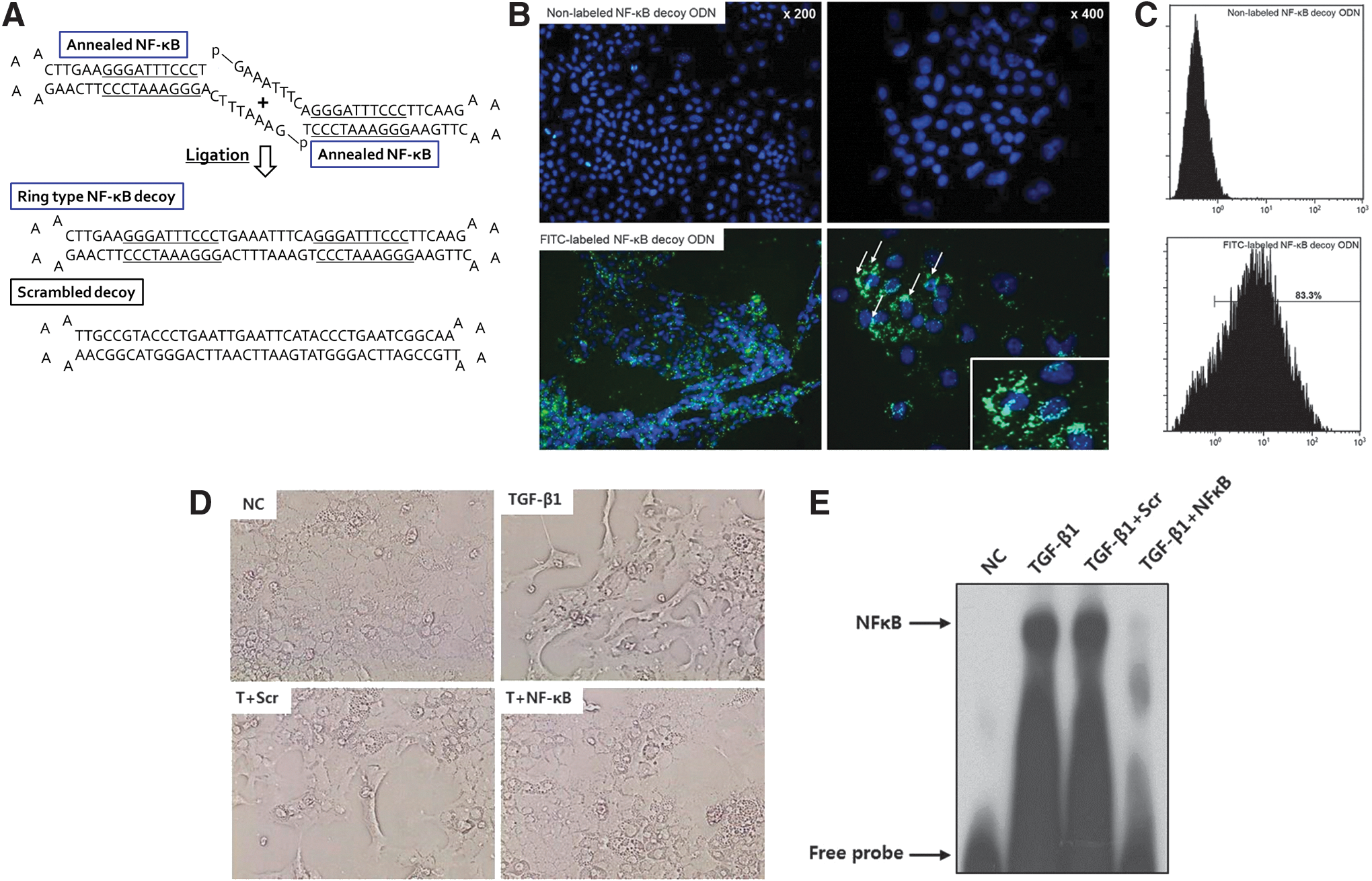
Structure of ring-type nuclear factor (NF)-κB decoy oligodeoxynucleotide (ODN) and effect of NF-κB decoy ODN on transforming growth factor (TGF)-β1-induced hepatocyte differentiation.
Cell culture
AML12 (CRL-2254), the murine hepatocyte cell line, was purchased from the American Type Culture Collection (ATCC, Manassas, VA). AML12 cells were grown in Dulbecco's modified Eagle's medium (DMEM)/F-12 medium containing 10% fetal bovine serum supplemented with insulin–transferrin–selenium-X, dexamethasone (40 ng/ml), penicillin (100 U/ml), and streptomycin (100 μg/ml) (Gibco BRL; Life Technologies, Bethesda, MD).
Transfection of NF-κB decoy ODN and transfection efficiency
The transfection efficiency of decoy ODN was examined by fluorescence microscopy (Eclipse 80i; Nikon, Tokyo, Japan) and flow cytometry (Navios; Beckman Coulter, Miami, FL). Before the transfection, NF-κB decoy ODN was labeled with fluorescein isothiocyanate (FITC), using a Label IT nucleic acid-labeling kit (Mirus Bio, Madison, WI). AML12 cells were cultured in 6-well plates (0.8×105 cells/ml) and transfected with 2 μg of FITC-labeled NF-κB or nonlabeled NF-κB decoy ODN, using FuGENE HD (DNA:lipid molar ratio, 2:3) (Roche, Mannheim, Germany). For flow cytometric analysis, cells were washed, trypsinized, dispersed, and then transferred into 500 μl of phosphate-buffered saline (PBS). Samples were analyzed with Navios (Beckman Coulter) and calculations were done with Kaluza software (Beckman Coulter).
For transfection analysis, AML12 cells were incubated with serum starvation for 24 hr before the experiment. Cells were then transfected with 2 μg of NF-κB or Scr decoy ODN, using FuGENE HD. After incubation at 37°C for 8 hr, AML12 cells were cultured in serum-free medium containing TGF-β1 (2.5 ng/ml; Santa Cruz Biotechnology, Santa Cruz, CA) for 48 hr.
Immunofluorescence staining
AML12 cells were collected by Cytospin centrifugation (Thermo Fisher Scientific, Waltham, MA) and fixed with 3.7% paraformaldehyde in PBS for 30 min at room temperature. Cells were permeated with 0.5% Triton for 15 min and incubated with primary antibodies against vimentin (BD Biosciences, San Jose, CA) and E-cadherin (Cell Signaling Technology, Beverly, MA) for 1 hr at room temperature. After washing with the PBS, cells were incubated with secondary antibodies (Alexa Fluor 488 and/or Alexa Fluor 594) for 30 min at room temperature. Cells were then counterstained with Hoechst 33342 for 10 min at 37°C. Immunofluorescence labeling was examined with an Eclipse 80i microscope (Nikon).
Animal model and NF-κB decoy ODN transfer
Animal protocols were approved by the Institutional Animal Care and Use Committee of the Catholic University of Daegu (Daegu, South Korea). Male C57BL/6 mice (6 weeks old, 20–22 g; Orient Bio, Seongnam, South Korea) were housed in a room with controlled humidity and temperature, and a 12-hr light–dark cycle. To examine the in vivo transfection efficiency of NF-κB decoy ODN, FITC-labeled NF-κB decoy ODN was injected into mice via the tail vein. The mice were killed 24 hr after injection. Liver tissues were frozen with O.C.T. compound (Sakura Finetek Japan, Tokyo, Japan). Cryosections of liver, which were transfected with FITC-labeled NF-κB decoy ODN, were examined by fluorescence microscopy.
Mice were randomly divided into three groups as follows: (1) untreated group (normal control, NC), (2) group treated with CCl4 and Scr decoy ODN (CCl4+Scr), and (3) group treated with CCl4 and NF-κB decoy ODN (CCl4+NF-κB). Chronic liver injuries were induced by intraperitoneal injection of CCl4 (2 ml/kg, dissolved in corn oil [1:3 ratio]) three times a week.
One week after the first CCl4 injection, Scr or NF-κB decoy ODN (10 μg) was transferred biweekly via the mouse tail vein, using an in vivo gene delivery system (Mirus Bio). Mice were killed 4 and 8 weeks after the first CCl4 injection.
Histopathology and immunohistochemistry
Paraffin-embedded liver tissues were sectioned and stained with Masson's trichrome according to standard protocol. Immunohistochemical staining of liver sections was performed as described previously (Park et al., 2009). Primary antibodies used were as follows: type I collagen and fibronectin (Santa Cruz Biotechnology), TGF-β1 (R&D Systems, Minneapolis, MN), and α-smooth muscle actin (SMA; Sigma-Aldrich, St. Louis). For immunofluorescence staining, the sections were incubated with primary antibodies against E-cadherin and vimentin. The slides were examined with an Eclipse 80i microscope (Nikon).
Measurement of alanine aminotransferase and aspartate aminotransferase
The blood of each mouse was collected and serum was separated to analyze the activity of alanine aminotransferase (ALT) and aspartate aminotransferase (AST), using an automated analyzer.
ELISA
The blood of each mouse was collected and serum was separated. Serum tumor necrosis factor (TNF)-α and interleukin (IL)-1β concentrations (pg/ml) were measured with Quantikine mouse TNF-α and IL-1β kits (R&D Systems).
Western blot analysis
Cells and liver tissue were homogenized in radioimmunoprecipitation assay (RIPA) buffer (Cell Signaling Technology) for 15 min on ice and centrifuged at 6000×g for 15 min. The supernatant was collected and analyzed by Western blot as described previously (Park et al., 2009). Primary antibodies used were against E-cadherin, vimentin, TGF-β1, α-SMA, fibronectin, type I collagen, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Signal intensity was quantified with an image analyzer (LAS-3000; Fuji, Tokyo, Japan).
Electrophoretic mobility shift assay
Nuclear extracts of cells and liver tissues were made with an NE-PER nuclear and cytoplasmic extraction kit (Thermo Fisher Scientific). A DIG (digoxigenin) gel shift kit (Roche) was used for electrophoretic mobility shift assay (EMSA) analysis, according to the manufacturer's protocol. The probe for NF-κB was as follows: 5′-CTT GAA
Statistical analyses
Results were analyzed with Duncan's test and expressed as means±SD. All experiments were performed at least three times.
Results
Transfection efficiency of NF-κB decoy ODN in AML12 cells
To assess the successful transfer of NF-κB decoy ODN, the transfection efficiency of the FITC-labeled ODN was initially analyzed by fluorescence microscopy. Cells transfected with nonlabeled NF-κB decoy ODN did not show any specific fluorescence (Fig. 1B, top). Twenty-four hours after transfection of the FITC-labeled NF-κB decoy ODN, fluorescence was detected in both the cytoplasm and nuclei of AML12 cells (Fig. 1B, bottom). To quantify the proportion of FITC-labeled NF-κB decoy ODN in AML12 cells, flow cytometric analysis was performed. Flow cytometric analysis showed a high positive rate (83.3%) of fluorescence activity compared with nonlabeled NF-κB decoy ODN in AML12 cells (Fig. 1C). These results indicated that NF-κB decoy ODN was effectively transfected into AML12 cells.
Effect of NF-κB decoy ODN in AML12 cells treated with TGF-β1
To induce EMT, AML12 cells were incubated for 48 hr with TGF-β1 (2.5 ng/ml), which is the most important profibrogenic cytokine in the process of liver EMT. In response to TGF-β1, the epithelial cobblestone appearance of the AML12 cells changed to a fibroblastic spindle-shaped morphology after 48 hr of treatment (Fig. 1D). To examine the effect of NF-κB decoy ODN in TGF-β1-treated hepatocytes, NF-κB DNA-binding activity was analyzed by EMSA. The DNA-binding activity of NF-κB was increased after treatment with TGF-β1 and transfection with Scr decoy ODN, whereas the increase was greatly suppressed by the NF-κB decoy ODN (Fig. 1E). E-cadherin is a prototypical epithelial cell marker that plays an important role in the maintenance of cellular integrity at the cell–cell junction. In addition, vimentin is an intermediate filament used to identify mesenchymal cells in EMT (Zeisberg and Kalluri, 2004). The changes in AML12 cellular morphology were accompanied by an increase in the labeling intensity of vimentin after TGF-β1 treatment. NF-κB decoy ODN treatment attenuated the expression of vimentin compared with Scr decoy ODN treatment. In contrast, the decreased labeling of E-cadherin after TGF-β1 treatment was reversed by treatment with NF-κB decoy ODN (Fig. 2A). Consistent with the immunofluorescence staining, NF-κB decoy ODN treatment significantly inhibited the upregulation of vimentin and fibronectin as well as the downregulation of E-cadherin (Fig. 2B and C). These results indicate that NF-κB decoy ODN effectively blocks mesenchymal changes and maintains the epithelial phenotype in TGF-β1-induced hepatocyte EMT.

NF-κB decoy ODN suppresses TGF-β1-induced epithelial–mesenchymal transition (EMT) in AML12 murine hepatocytes. AML12 hepatocytes were treated with TGF-β1 (2.5 ng/ml) in the presence of Scr or NF-κB decoy ODN as indicated.
Effect of NF-κB decoy ODN in CCl4-induced liver fibrosis
On the basis of the results of NF-κB decoy ODN administration in AML12 hepatocytes, we determined the effect of NF-κB decoy ODN on CCl4-induced liver fibrosis in vivo. To confirm the successful transfer of NF-κB decoy ODN in vivo, we examined the distribution of FITC-labeled NF-κB decoy ODN in mouse liver. The FITC-labeled NF-κB decoy ODN exhibited apparent fluorescence in mouse liver (Fig. 3A).
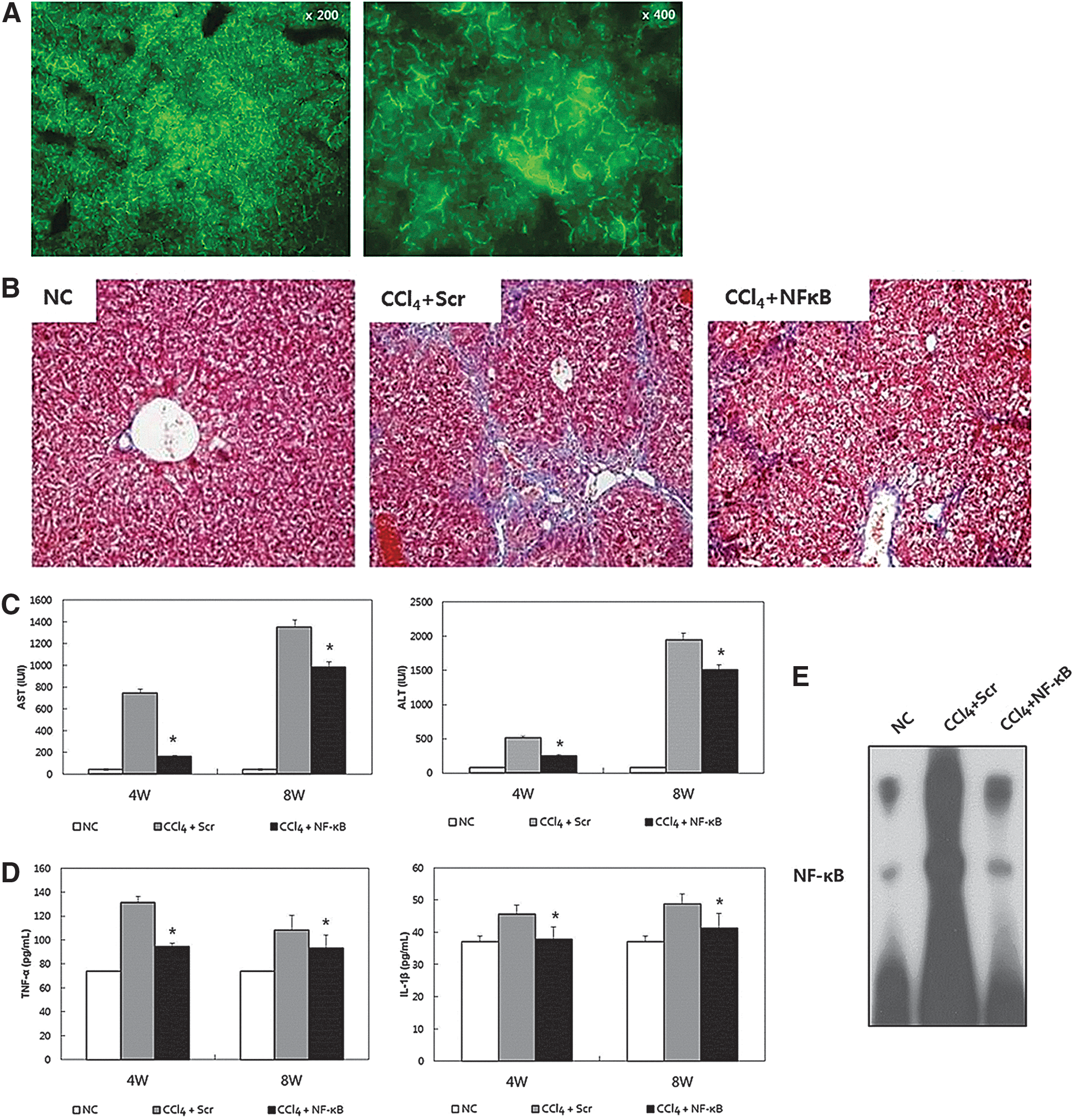
NF-κB decoy ODN effectively suppresses CCl4-induced liver fibrosis through inhibition of inflammatory changes and NF-κB-binding activity.
We then investigated the effect of NF-κB decoy ODN in CCl4-induced liver fibrosis. NF-κB decoy ODN was transferred biweekly during CCl4 administration. According to Masson's trichrome staining, 8 weeks of treatment with CCl4 established bridging fibrosis in the CCl4+Scr mice, whereas NF-κB decoy ODN was shown to markedly suppress fibrosis in CCl4+NF-κB mice (Fig. 3B). At 4 and 8 weeks after CCl4 administration, AST and ALT were increased in CCl4+Scr mice, whereas the increase was significantly smaller in CCl4+NF-κB mice (Fig. 3C). CCl4 administration increased the serum concentrations of TNF-α and IL-1β in CCl4+Scr mice. However, these increases were significantly inhibited in CCl4+NF-κB mice (Fig. 3D). To determine the effect of NF-κB decoy ODN on NF-κB DNA-binding activity in liver fibrosis, EMSA was performed to analyze the transcriptional activity of NF-κB. NF-κB was significantly increased in CCl4+Scr mice. In contrast, transfection of NF-κB decoy ODN significantly suppressed the activation of NF-κB in CCl4+NF-κB mice (Fig. 3E).
Effect of NF-κB decoy ODN during EMT in liver fibrogenesis
To assess the molecular mechanism of NF-κB decoy ODN, we examined its effects on liver fibrosis and matrix accumulation. An immunohistochemical study demonstrated that accumulation of cells positive for TGF-β1, fibronectin, α-SMA, and type I collagen within the liver sinusoid was increased in CCl4+Scr mice compared with NC mice. However, NF-κB decoy ODN effectively suppressed the expression of TGF-β1, fibronectin, α-SMA, and type I collagen in CCl4+NF-κB mice (Fig. 4A). Consistent with the immunohistochemical staining, Western blotting results showed that NF-κB decoy ODN significantly inhibited the expression of TGF-β1, fibronectin, α-SMA, and type I collagen (Fig. 4B). These results collectively suggest that inhibition of NF-κB is able to ameliorate liver fibrotic lesions through the suppression of TGF-β1 and ECM-related proteins in liver fibrosis.

Inhibition of fibrosis-related gene expression by NF-κB decoy ODN treatment in CCl4-induced liver fibrosis.
During liver fibrogenesis, hepatocytes lose their epithelial phenotype and acquire features of mesenchyme via EMT processes. We next examined the effect of NF-κB decoy ODN on the expression of E-cadherin and vimentin, which are essential markers for epithelial cells and mesenchymal cells, respectively. Consistent with a previous report (Zeisberg et al., 2007), immunofluorescence staining revealed that administration of CCl4 induced suppression of E-cadherin as well as dramatic induction of vimentin, a shift that is in agreement with hepatocyte EMT. Interestingly, NF-κB decoy ODN treatment resulted in restoration of E-cadherin and reduction of vimentin expression in fibrotic liver (Fig. 5A). In addition, Western blotting results showed that NF-κB decoy ODN treatment preserved E-cadherin expression and inhibited vimentin induction in CCl4+NF-κB mice (Fig. 5B). Therefore, blocking NF-κB activity effectively inhibits EMT during fibrotic changes in the liver.
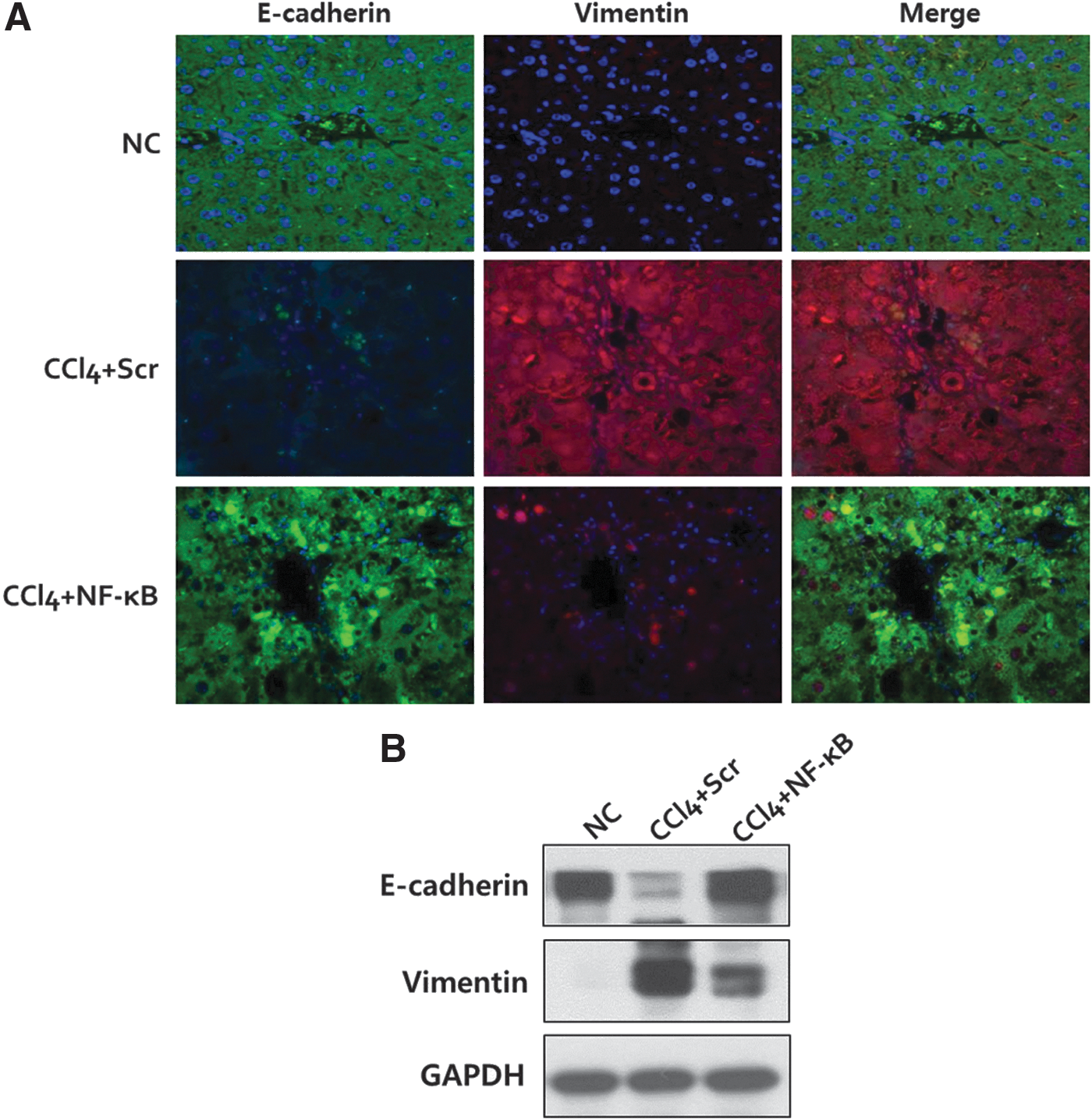
Inhibition of epithelial–mesenchymal transition by NF-κB decoy ODN treatment in CCl4-induced liver fibrosis.
Discussion
NF-κB regulates the transcription of many genes, such as interleukins and adhesion molecules, that are primarily related to inflammation. The present study has investigated how NF-κB acts on the EMT process during liver fibrogenesis.
TGF-β1-induced EMT has been implicated in tissue and organ fibrosis (Zeisberg and Kalluri, 2004; Willis and Borok, 2007). Inflammation is closely related to the progression of organ fibrosis and cancer (Cordon-Cardo and Prives, 1999; Mantovani et al., 2008). However, the detailed molecular mechanisms of inflammation for organ fibrosis have only begun to be elucidated. Most liver injuries activate acute inflammatory and wound-healing responses. NF-κB has been closely linked to the induction of proinflammatory gene expression and has been focused on as a therapeutic target for inflammatory diseases. Persistent hepatic damage activates the inflammatory cascade through the activation of NF-κB in various nonparenchymal and parenchymal liver cells (Elsharkawy and Mann, 2007). In particular, it has been reported that the exposure of various types of injury (alcohol, endotoxin, TNF-α, and cholestasis) induces the expression of NF-κB in hepatocytes (Miyoshi et al., 2001). However, whether the inhibition of NF-κB plays a role in TGF-β1-induced EMT in hepatocytes remains unanswered. Here, we provide evidence that the inhibition of NF-κB, using a decoy ODN strategy, attenuates liver fibrogenesis through inhibition of the EMT process.
In this study, we demonstrated that NF-κB inactivation works to attenuate hepatocyte EMT during liver fibrogenesis. To assess the effectiveness of NF-κB inactivation, we designed a ring-type NF-κB decoy ODN, which has two consensus NF-κB-binding sites in a single decoy molecule without an open end. In our previous reports, we developed a ring-type NF-κB decoy ODN constructed by enzymatic ligation with two hairpin overhangs at both ends to prevent degradation by nucleases. This ring-type NF-κB decoy ODN was more stable than conventional phosphorothiolated double-strand ODNs in the presence of serum or nucleases (Kim et al., 2009). We observed that NF-κB decoy ODN treatment significantly inhibited TGF-β1-induced hepatocyte EMT in vitro. Direct inhibition of NF-κB suppresses chronic inflammation and extracellular matrix expression in CCl4-induced liver fibrogenesis. Treatment with NF-κB decoy ODN was also shown to effectively inhibit the EMT processes in vivo in the current study. Disruption of tight junctions and adherens junctions is the critical process of EMT. Downregulation of E-cadherin, which is the main component of adherens junctions, is widely known to be a marker of EMT in epithelial cells. This study demonstrated that NF-κB decoy ODN treatment restored the expression of E-cadherin in vivo and in vitro. Several studies have reported that NF-κB is a potent activator of Snail, Twist, and Zeb, which directly activate EMT at the transcriptional level (Peinado et al., 2004). In addition, one study has shown that NF-κB stabilizes the expression of Snail, which is involved in inflammation-induced cell migration and invasion (Wu et al., 2009). These findings indicate that NF-κB decoy ODN treatment regulates not only inflammation-related gene expression, but also the expression of EMT-related genes, which leads to potent therapeutic effects on the degeneration of hepatocyte integrity during liver fibrogenesis. Therefore, a possible advantage of NF-κB decoy ODN is the inhibition of transcription of several genes including EMT-related genes through the regulation of NF-κB function.
In summary, our data collectively show that NF-κB plays an important role in EMT during liver fibrogenesis. Inhibition of NF-κB by the decoy ODN strategy led to attenuation of liver fibrosis and improvement of liver function. Our findings may open new avenues for the suppression of EMT in liver fibrogenesis, thus leading to new therapeutic options for liver fibrosis.
Footnotes
Acknowledgments
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government [NRF-2009-353-E00022]. This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government [2012R1A1A401015639].
Author Disclosure Statement
There are no conflicts of interest to disclose for any of the authors.