
Abstract
Asthma exacerbations are caused primarily by viral infections. Antisense and small interfering RNA (siRNA) technologies have gained attention as potential antiasthma and antiviral approaches. In this study we analyzed whether gene silencing of interleukin (IL)-4 expression and respiratory syncytial virus (RSV) replication by RNA interference is able to suppress allergen- and virus-induced responses in a mouse model of virus-induced asthma exacerbation. Knockdown efficacy of IL-4 siRNA molecules was analyzed in the human HEK293T cell line by cotransfection of six different siRNAs with a plasmid carrying mouse IL-4. The most potent siRNA was then used in a mouse model of RSV-induced asthma exacerbation. BALB/c mice were sensitized intraperitoneally with ovalbumin (OVA) and then infected 12 days later intranasally with RSV Long strain (1×106 TCID50/mouse), followed 1 day later by intranasal challenge with OVA for 3 days. Mice were pretreated intranasally three times with either siRNA to IL-4 or GFP control, 2 days before, and on the first two OVA challenge days. siRNAs to RSV or rhinovirus control were inoculated intranasally once, 3 hr before RSV infection. Combined anti-IL-4 and anti-RSV siRNAs were able to significantly reduce total cell counts and eosinophilia in bronchoalveolar lavage fluid, development of airway hyperresponsiveness, and airway inflammation and to downregulate IL-4 mRNA expression and RSV viral RNA, but to upregulate IFN-γ levels in lung tissues. We conclude that anti-helper T cells type 2 and antiviral siRNAs may constitute a new therapeutic approach for treatment of virus induced asthma exacerbations.
Introduction
A
Viral respiratory tract infections are associated with the majority of asthma exacerbations in both children and adults (Pattemore et al., 1992; Johnston, 2005). Several types of viruses, such as rhinoviruses (RVs), respiratory syncytial virus (RSV), human metapneumovirus (hMPV), influenza, parainfluenza, and coronaviruses, have been detected in respiratory tract secretions from patients with asthma exacerbations (Newcomb and Peebles, 2009). RSV is the major cause of bronchiolitis and is the commonest single cause of hospitalization during infancy. Treatment of RSV infection with antiviral drugs is currently of limited efficacy, there is still no vaccine available, and the previous formalin-inactivated vaccine produced enhanced disease in clinical trials (Habibi and Openshaw, 2012).
Antisense and small interfering RNA (siRNA) technologies have gained greater attention in allergy and asthma research. Anti-mRNA agents are used for sequence-specific posttranscriptional inhibition of gene expression (Popescu, 2005). Targeting IL-4 has been one of such strategy, because IL-4 plays a crucial role in Th2 responses and in isotype class switching of B cells to IgE synthesis, and is also involved in mast cell recruitment (Maes et al., 2012). According to data obtained by Brusselle and colleagues in experiments with IL-4-deficient mice, repeated allergen exposure in sensitized mice induces airway inflammation and bronchial hyperresponsiveness, and IL-4 plays a predominant role in the pathogenesis of both phenomena (Brusselle et al., 1995).
In addition, siRNAs are potent in specific viral gene expression inhibition, which reduces viral replication (Dector et al., 2002; Cheng et al., 2003; Ge et al., 2003). In this study we aimed to examine the therapeutic potential of the dual action of anti-IL-4 and anti-RSV siRNAs in a mouse model of RSV-induced asthma exacerbation.
Materials and Methods
Mice, virus, siRNAs, and other reagents
Female 6- to 8-week-old BALB/c mice (Stolbovaya, Moscow, Russia) were kept on an ovalbumin (OVA)-free diet under pathogen-free conditions. The RSV Long strain was grown in MA-104 cells (epithelial monkey kidney cells) and purified by sucrose gradient centrifugation (Ueba, 1980). The final preparations had titers in the range of 106–107 TCID50 (median tissue culture infective dose)/ml. siRNAs targeted to IL-4 were designed using OligoWalk web server for siRNA design (Lu and Mathews, 2008). siRNA against gfp served as a negative control and has been described previously (Martinsson-Ahlzen et al., 2008). siRNAs, primers, probes, and kits for real-time PCR (RT-PCR) were purchased from Syntol (Moscow, Russia). RSV and rhinovirus (RV) siRNAs as well as the TaqMan assay for measuring RSV RNA have been described previously (Akimov et al., 2007). Sequences of the siRNAs used in this study are listed in Table 1. The siRNAs were used unmodified and unconjugated.
Construction of a plasmid carrying mouse il-4 and gfp genes
To clone mouse IL-4, the coding sequence of the mouse il-4 gene was amplified by PCR from concanavalin A-stimulated mouse splenocyte cDNA, verified by sequencing, and cloned into the lentiviral expression vector pUCHR IRES GFP (Mazurov et al., 2010). Expression of the gfp gene, located downstream of the internal ribosome entry site (IRES), and of green fluorescent protein (GFP) was used as an indicator of IL-4 expression level.
Cell cultures and cotransfection with plasmid and siRNA
The human embryonic kidney HEK293T cell line was cultured in Dulbecco's modified Eagle's minimal essential medium (DMEM) containing 10% fetal calf serum (FCS), Gibco gentamicin (80 mg/liter; Life Technologies, Grand Island, NY), and 25 mM HEPES. Subconfluent cultures were split with trypsin–ethylenediaminetetraacetic acid (EDTA). All culture media and reagents, unless otherwise stated, were obtained from Paneco (Moscow, Russia). For transfection with plasmid and siRNAs, cells were seeded in 24-well culture plates at 1×105 cells per well in appropriate medium and cultured (37°/5% CO2) for 24 hr before transfection. In all cases, adherent cells were cotransfected with 0.25 μg of plasmid and 0.5 μg of siRNAs, using Lipofectamine 2000 (Life Technologies) as recommended by the manufacturer. The siRNA targeting RV and siRNA targeting GFP were used as negative and positive controls, respectively, in experiments evaluating anti-IL-4 siRNA activity in vitro. Twenty-four hours after transfection cell supernatants, cells for fluorescence-activated cell-sorting (FACS) analysis, and lysates were harvested. The level of GFP protein expression was quantified by FACS analysis, and IL-4 secretion was determined by ELISA (IL-4 ELISA set; BD Biosciences, San Jose, CA). The level of mRNA for IL-4 was measured by quantitative RT-PCR as described below.
Asthma exacerbation model
A schematic presentation of treatment protocols is shown in Fig. 1. Briefly, mice were sensitized with 20 μg of ovalbumin (OVA) from chicken eggs, emulsified in 2 mg of aluminum hydroxide, intraperitoneally on days 1, 14, and 21 (on day 21, OVA at 10 μg). Mice were intranasally challenged with OVA (per mouse: 50 μl of a 10-mg/ml solution in phosphate-buffered saline [PBS]) on three consecutive days (days 35–37) to induce allergic airway inflammation. On day 34 mice were intranasally infected with 50 μl of RSV Long strain (1×106 TCID50/mouse). Mice were intranasally treated with siIL4 or siGFP three times (days 33, 35, and 36), at a total dose of 100 μg/mouse. siRSV or siRV was intranasally inoculated 3 hr before RSV infection, at a dose of 70 μg/mouse. We assessed lung function on day 5 after infection and mice were killed on day 6 after infection (Fig. 1) for end-point analyses. Unchallenged mice did not receive any treatment (neither OVA nor RSV) and served as a negative control.

Experimental protocol of animal sensitization, challenge, RSV infection, and siRNA treatment. OVA, ovalbumin from chicken eggs; RSV, respiratory syncytial virus (Long strain); AHR, airway hyperresponsiveness; BAL, bronchoalveolar lavage; i.n., intranasal; i.p., intraperitoneal; V, volume; siRSV, siRV, siIL4, and siGFP, siRNAs against RSV, rhinovirus (RV), mouse IL-4, and GFP, respectively.
In experiments with IL-4 siRNA alone, the OVA sensitization model was used. Mice were sensitized by intraperitoneal injection of OVA/alum on days 1, 14, and 21, and challenged with OVA via the airways on days 35–37 (on days 33, 35, and 36 mice were treated with IL-4 or GFP siRNAs at a total dose of 100 μg/mouse). Lung function was measured 24 hr after the last OVA challenge. Mice were killed on day 39. IL-4 protein release in the bronchoalveolar lavage (BAL) was measured by ELISA (IL-4 ELISA set; BD Biosciences).
RSV infection model
RSV infection alone (for anti-RSV siRNA testing) was performed with the RSV Long strain.
Four groups of mice were treated intranasally with 60 μg of various specific siRNAs (RSV01, RSV02, and RSV03) targeting the p gene of RSV, unspecific RV siRNA (targeted HRV), and 40 μl of RSV (2×106 TCID50/ml). The fifth group was left untreated (unchallenged mice). Six days later mice were killed for end-point analyses.
TaqMan quantitative real-time PCR
We excised the right lobe of the lung from each mouse and stored the tissue at –80°C until use. Total RNA was isolated from pUCHR IRES GFP/siRNA-transfected HEK293T cells or mouse lung tissue, using RNeasy mini kits (Qiagen, Valencia, CA), and thereafter reverse-transcribed into cDNA, using random hexamers and a RevertAid H minus first strand cDNA synthesis kit (Fermentas; Thermo Fisher Scientific, Waltham, MA). The reverse transcription reaction product was amplified by RT-PCR, using an iCycler iQ real-time PCR detection system (Bio-Rad Laboratories, Hercules, CA). The following primers and probe were used for mouse IL-4 mRNA quantification by RT-PCR: forward, 5′-AGATGGATGTGCCAAACGTC; reverse, 5′-TATGCGAAGCACCTTGGAAG; probe, 5′-AAGAACACCACAGAGAGTGAGCTCGT.
For mouse interferon (IFN)-γ quantification: forward, 5′-AAATCCTGCAGAGCCAGATTAT; reverse, 5′- GCTGTTGCTGAAGAAGGTAGTA; probe, 5′-ACGCTTATGTTGTTGCTGATGGCC. We normalized real-time PCR assays to human β-actin mRNA (in vitro experiments) and to mouse hypoxanthine–guanine phosphoribosyltransferase (HPRT; in vivo experiments).
Airway hyperresponsiveness
Twenty-four hours after the final allergen challenge (on day 38) AHR was measured in mice, using a FinePointe NAM (noninvasive airway mechanics) system (Buxco Electronics, Wilmington, NC). Briefly, mice were placed in a double-chamber plethysmograph, which consists of a nasal (head) chamber and a thoracic (body) chamber. The nasal chamber served as a means for delivering aerosolized methacholine (Sigma-Aldrich, St. Louis, MO). Each mouse received nebulized PBS, followed by increasing concentrations of methacholine (6.25, 12.5, 25, and 50 mg/ml) to induce bronchoconstriction. Increasing doses of aerosolized methacholine were delivered for 2 min, and the response to each dose was measured for the next 3 min. Lung function was recorded and calculated by FinePointe software as sRaw (specific airway resistance, cmH2O×sec–1×ml–1).
Bronchoalveolar lavage
Mouse lungs were cannulated through the trachea and then the lungs were lavaged. Cellular fractions were processed for differential azure–eosin staining by cytospinning. Cell counting was performed by light microscopy (at least 200 cells per slide) in a blinded manner.
Histology
The left lobe of each lung was fixed in 4% formalin. Lung sections (4 μm) were stained with hematoxylin and eosin for histological evaluation and monitoring of inflammatory cells. Slides were rehydrated through a series of decreasing ethanol washes and stained with a periodic acid–Schiff kit (Sigma-Aldrich) to visualize goblet cells. The histopathological changes (cell infiltration around bronchioles, peribronchial arteries, and veins; and proliferation of goblet cells in bronchial epithelium) were graded according to a modified semiquantitative scoring system in a blinded manner (none, 0; mild, 1.0; moderate, 2.0; or severe, 3.0) (Ennis et al., 2005).
OVA-specific IgE assay
Serum anti-OVA IgE antibody titers were determined by ELISA, using a mouse IgE ELISA set (BD OptEIA; BD Biosciences) as recommended by the manufacturer, with only one exception (plates were coated with OVA at 5 μg/well).
Statistical analysis
All data were normally distributed and are expressed as means±SEM. Data were analyzed by one-way analysis of variance (ANOVA) followed by Bonferroni's post-test, with 95% confidence levels (Prism 4; GraphPad Software, San Diego, CA). Data were accepted as significantly different at p<0.05.
Results
Evaluation of IL-4 siRNA activity in vitro
To evaluate the six various IL-4 siRNA activities in vitro, we constructed a plasmid simultaneously expressing mouse il-4 and gfp genes (pUCHR IRES GFP). We cotransfected HEK293T cells with vector pUCHR IRES GFP and six different siRNA variants to evaluate their knockdown activity. Expression of GFP, measured by flow cytometry, was used as a positive control of transfection (data not shown). siRNA against RV was used as a negative control. Five of six designed siIL4s reduced IL-4 expression in vitro with various levels of efficacy. Two siIL4s (siIL4-267 and siIL4-408) significantly inhibited expression of mouse IL-4, based on data obtained by RT-PCR (Fig. 2A) and ELISA (Fig. 2B). These siRNAs decreased the level of IL-4 mRNA up to 73 and 75%, respectively (compared with cells transfected with unspecific siRV). The level of IL-4 protein secreted into the medium from the producer HEK293T cells, measured by ELISA, was downregulated by 67% for siIL4-267 and by 83% for siIL4-408. According to these findings, siIL4-408 was chosen as the most potent IL-4-silencing siRNA and was used for the following experiments in vivo. Data were obtained in five independent experiments.
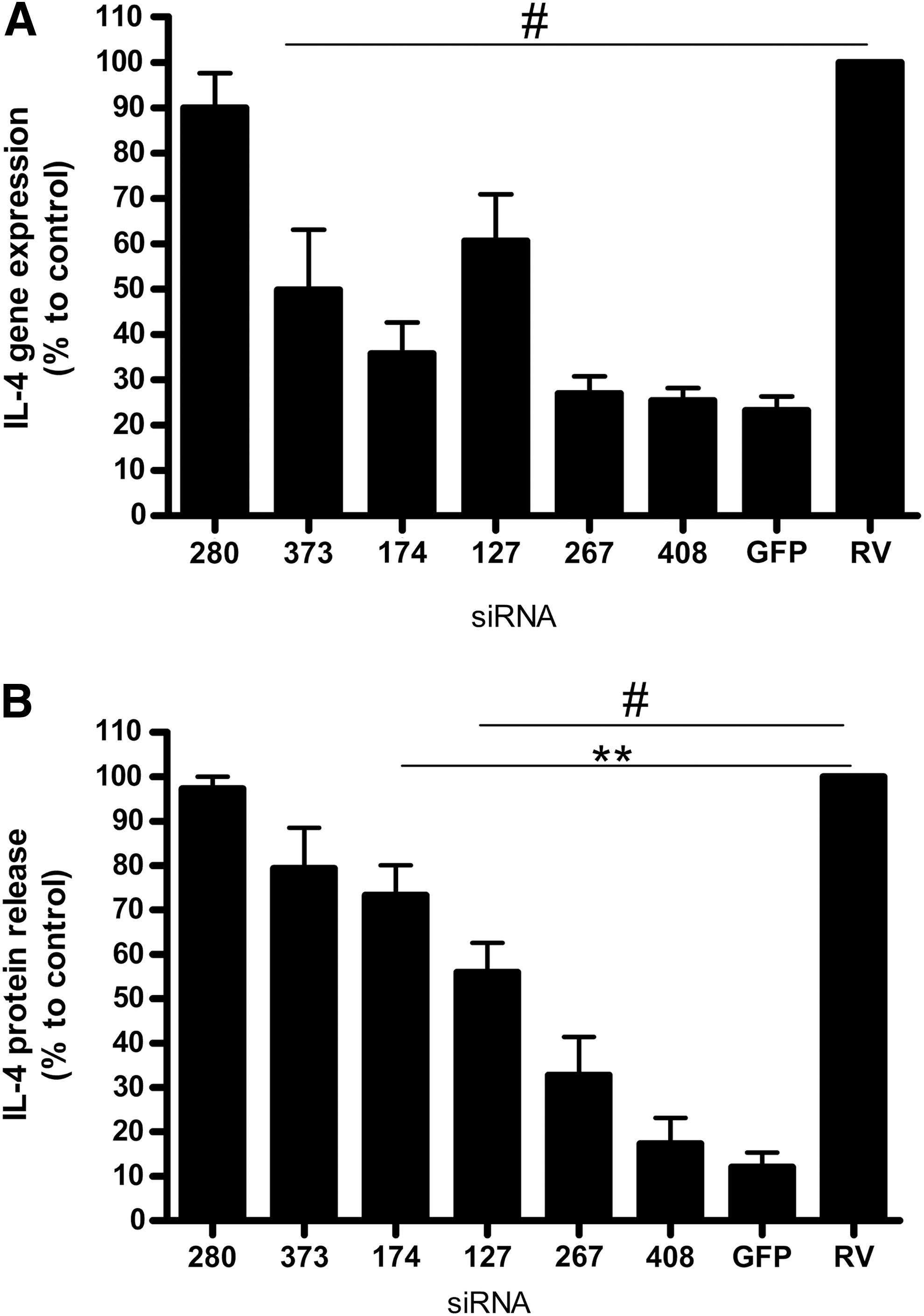
Identification of most potent siRNA inhibiting IL-4 expression in human cell line HEK293T by RT-PCR
Anti-RSV and anti-IL-4 siRNAs reduce IL-4 expression, RSV replication, and development of AHR, but increase IFN-γ expression in the lung in vivo
We used a mouse RSV-induced asthma exacerbation model to verify siRNA activity in vivo. Sensitized mice were infected with the RSV Long strain, challenged with OVA, and treated intranasally with siRNAs (siRSV and siIL4 [or siRV and siGFP as negative controls]) as described in Materials and Methods and Fig. 1. We analyzed the expression levels of IL-4 mRNA (Fig. 3A), IFN-γ mRNA (Fig. 3C), and RSV viral RNA (vRNA) (Fig. 3B) in lung tissue by RT-PCR. OVA sensitization/challenge and RSV infection (OVA+RSV group) resulted in increased levels of IL-4 mRNA expression and decreased levels of IFN-γ expression compared with unchallenged mice. The intranasal application of siIL4/RSV to sensitized/challenged and infected animals reduced IL-4 mRNA expression by 69% and the level of RSV vRNA by 71% compared with negative control siRNA-treated mice, but increased IFN-γ mRNA expression up to 50%. OVA sensitization/challenge and RSV infection enhanced airway reactivity compared with untreated control animals (Fig. 3D). In contrast to mice treated with negative control siRNAs, siIL4/RSV-treated mice developed reduced levels of AHR. IgE levels, measured in mouse serum, were upregulated in OVA-sensitized animals and did not significantly change after IL4/RSV siRNA treatment (data not shown), which can be explained by local, rather than systemic actions of siRNA. Others have observed similar findings after anti-IL-4 and IL-13 treatment with short hairpin RNA (shRNA) (Lee et al., 2011). We found no significant difference between IgE levels in sensitized animals infected with RSV compared with mice treated only with OVA (data not shown).

IL-4 and RSV siRNAs reduced IL-4 expression, RSV replication, and development of AHR in the mouse lung and influenced IFN-γ production. OVA-sensitized mice were infected with RSV (OVA+RSV) and challenged intranasally with specific IL-4 and RSV siRNAs (OVA+RSV+IL-4/RSV siRNA) or with GFP/RV siRNAs (OVA+RSV+GFP/RV siRNA). Quantification of IL-4
Anti-RSV and IL-4 anti-siRNAs potently suppress lung inflammation and pulmonary histological alterations in vivo
Inflammatory eosinophils are thought to be crucial mediators for the development of asthma, and RSV infection induces rapid neutrophilic inflammation. The combination of OVA challenge and RSV infection (OVA+RSV) resulted in a significant increase in the number of total leukocytes (data not shown) and especially eosinophils (Fig. 4A) and neutrophils (Fig. 4B) in the BAL. Administration of siIL4/RSV reduced airway eosinophilia and neutrophilia. Treatment with negative control siRNAs did not reduce the percentage of eosinophils or neutrophils recovered from the airways. Next, we performed histopathological analysis of inflammation in the lungs of mice. Cell infiltration around bronchioles, peribronchial arteries, and veins, and proliferation of goblet cells in bronchial epithelium, were scored. Total inflammation, neutrophil, eosinophil, lymphocyte, and goblet cell infiltration scores (Fig. 5I) were significantly lower in the animals treated with anti-siIL4/RSV siRNAs compared with the OVA+RSV group and negative control siRNA-treated mice. Lung sections of each mouse were analyzed with regard to pathologic alterations resembling the histopathology of allergic asthma and viral infection, such as goblet cell hyperplasia and peribronchial infiltration (Fig. 5A–H). Control unchallenged mice did not present obstruction of the bronchoalveolar system or alterations of ciliated epithelium or eosinophil infiltration (Fig. 5G and H). OVA-sensitized and RSV-infected mice (OVA+RSV group) and the negative control siRNA group exhibited a large number of goblet cells, peribronchial infiltrates, and eosinophils (Fig. 5A, B, E, and F). In contrast, in anti-IL-4/RSV siRNA-treated animals only marginal inflammation was observed (Fig. 5C and D). Data were obtained in three independent experiments.

IL-4 and RSV siRNAs reduced eosinophil and neutrophil infiltration in BAL fluid. Mice were sensitized by intraperitoneal injection of OVA/alum on days 1, 14, and 21, and challenged with OVA via the airways on days 35–37 (on days 33, 35, and 36 the mice were treated with IL-4 or GFP siRNA). On day 31 mice were treated with siRSV or RV siRNA and 3 hr later infected with RSV. On day 39 (2 days after the last allergen airway challenge and 6 day after RSV infection) lungs were lavaged, and differential cell counts in BAL fluid were assessed. In addition, histological evaluation of the left lobe of the lungs was done. In contrast to OVA+RSV+IL-4/RSV siRNA and unchallenged mouse groups, OVA+RSV and negative control groups showed a strong increase in the number of total leukocytes (data not shown) and especially eosinophils
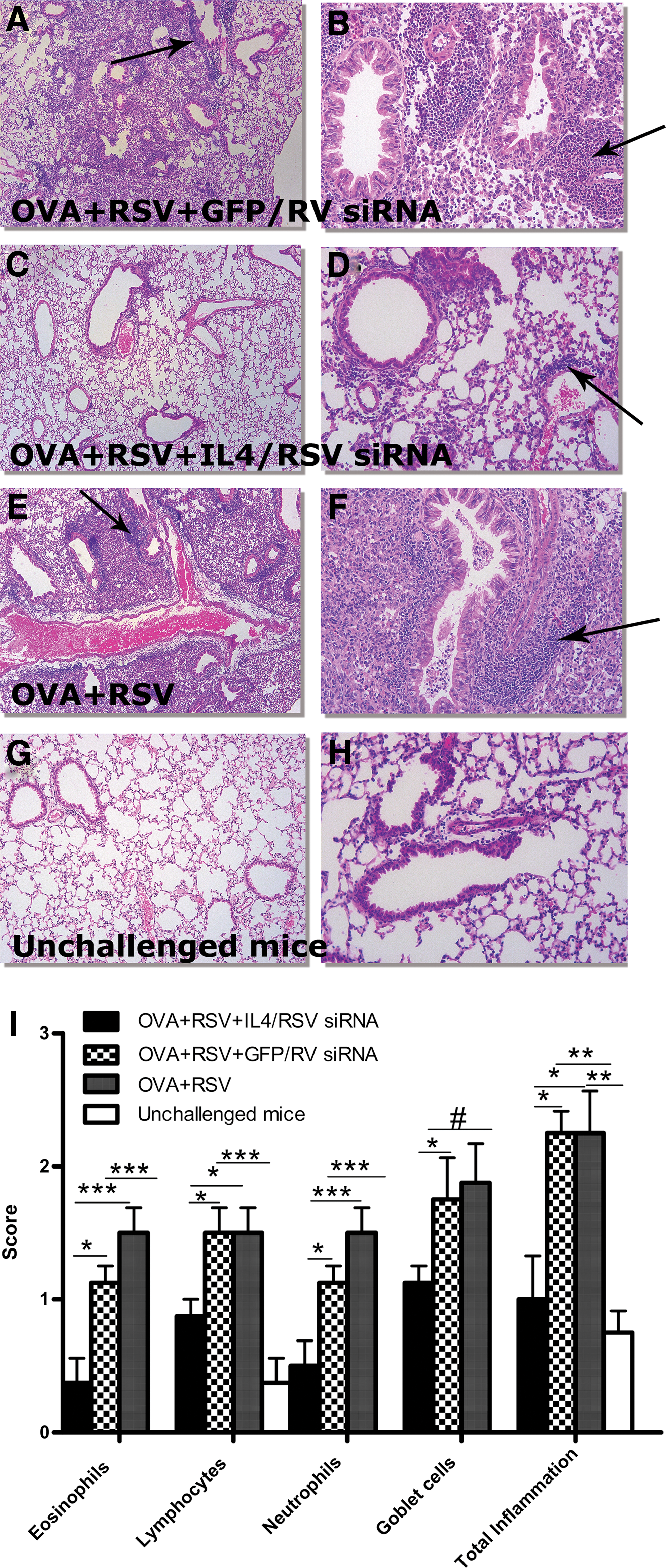
IL-4 and RSV siRNAs prevent histological alterations in lung tissues.
Effects of siIL-4 and siRSV when used alone
siRNAs against the p gene of RSV were tested in an RSV infection model. siRNAs RSV01 and RSV03 demonstrated the highest efficacy in vivo. They significantly reduced the vRNA level in mouse lung (Fig. 6A) as well as the total inflammation score (Fig. 6B). These data were published previously (Khaitov et al., 2010).

Anti-RSV siRNAs reduced viral load and total inflammation score in mice infected with RSV. Infection with RSV Long strain was done for anti-RSV siRNA testing. RSV vRNA levels
In the OVA-induced asthma model, treatment of mice with IL-4 siRNA significantly reduced AHR at a methacholine concentration of 12.5 mg/ml (Fig. 7A), IL-4 mRNA expression in lung tissue (Fig. 7B), and the IL-4 protein level in BAL (Fig. 7C) compared with mice treated with nonspecific GFP siRNA. Data were obtained in two independent experiments.
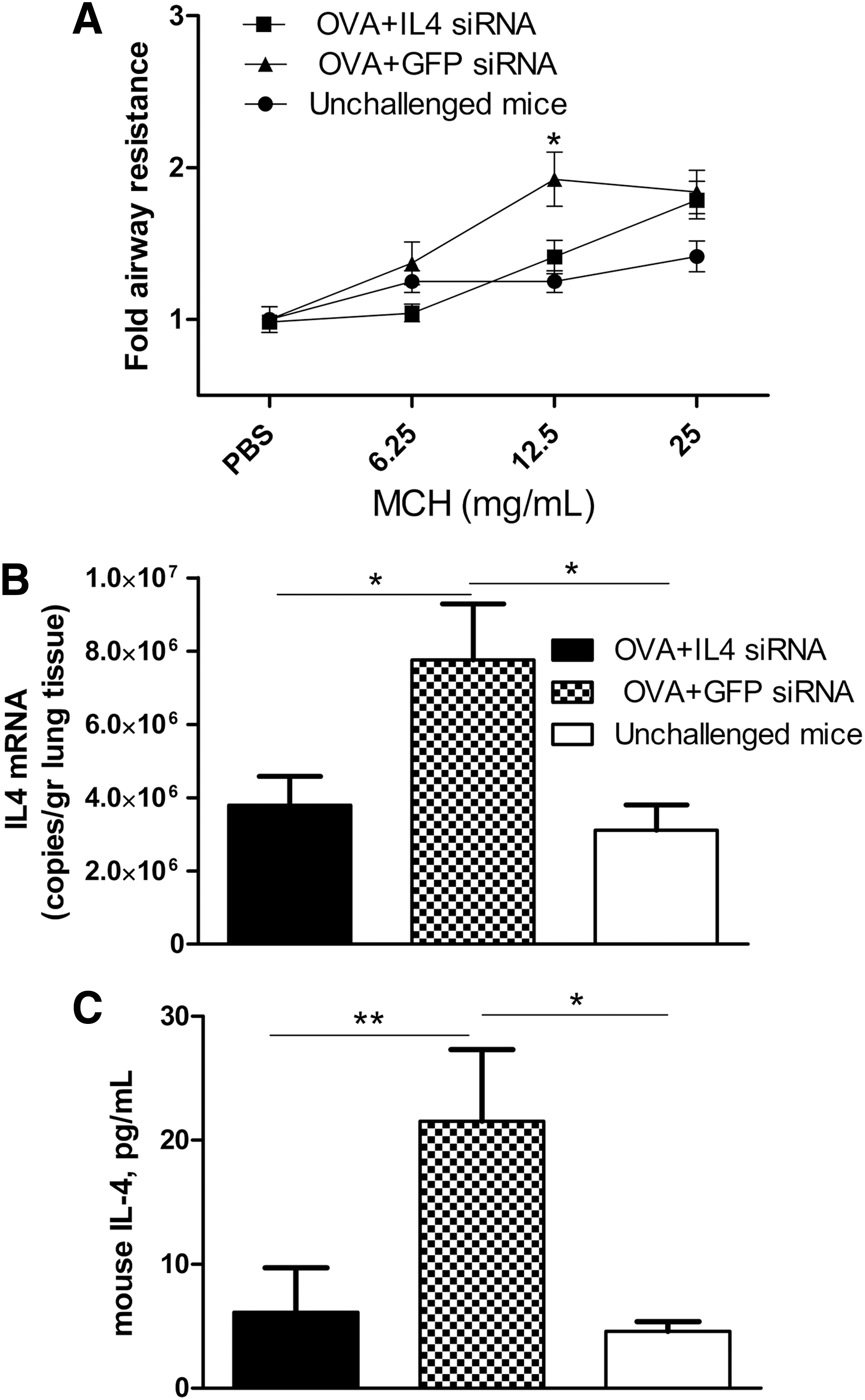
Anti-IL-4 siRNA diminished IL-4 expression and development of airway hyperresponsiveness (AHR) in OVA-sensitized mice.
Discussion
Gene silencing by RNA interference (RNAi) is a potent, selective, and widely applicable method used for specific blocking of targeted gene expression. Current antiasthma therapy is effective for many patients with asthma; however, there is still a considerable number of patients with poor quality of life due to uncontrolled or partially controlled asthma, and who could benefit from additional treatment options (Bateman et al., 2004). Several attempts were made to investigate the feasibility of siRNA-mediated gene-specific silencing of multiple genes as a means of controlling allergic disease in vivo (Darcan-Nicolaisen et al., 2009; Suzuki et al., 2009; Walker et al., 2009; Hosoya et al., 2011). ZaBeCor Pharmaceuticals (Bala Cynwyd, PA) has developed an siRNA drug for the treatment of asthma. This siRNA therapeutic, known as Excellair, targets the mRNA of spleen tyrosine kinase (Syk) (Burnett et al., 2011). This therapeutic has proven its efficacy in preclinical trials and was approved for clinical trials.
In patients with allergic asthma, CD4+ Th2 cells are believed to initiate and perpetuate the disease (Cohn et al., 2004). IL-4, IL-5, and IL-13 mRNA and protein levels are increased in BAL fluid, BAL cells, and airway biopsies of patients with asthma (Robinson et al., 1992; Huang et al., 1995). Thus, targeting IL-4 is a potential treatment strategy. Successful blocking of IL-4 has been achieved by administering soluble IL-4 receptor (IL-4R), by monoclonal antibodies (mAbs) and by antisense therapeutics (Maes et al., 2012). A human mAb (AMG-317) developed by Amgen (Thousand Oaks, CA) is reported to block the IL-4/IL-13 pathway and is currently in a phase II clinical trial in patients with moderate-to-severe asthma (Corren et al., 2010).
Viral infections with RV, influenza viruses, RSV, and so on, are known to be the main cause of asthma exacerbations in children and adults and often provoke wheezing in patients with asthma (Nicholson et al., 1993; Johnston et al., 1995). RSV is a major respiratory pathogen infecting almost all children by the age of 3 years; it causes various respiratory tract illnesses ranging from a common cold to severe bronchiolitis and pneumonia in patients of all age groups (Habibi and Openshaw, 2012). Until recently, no efficient antiviral drugs were available against RSV. An siRNA therapeutic (ALN-RSV01) targeting the nucleocapsid n gene of the RSV genome has been developed by Alnylam Pharmaceuticals (Cambridge, MA) and is now in phase IIb clinical trials (DeVincenzo et al., 2010).
The influence of RSV infection on allergic sensitization and the influence of allergic sensitization on RSV bronchiolitis have been examined in several studies (Kalina and Gershwin, 2004). Among the Th2 cytokines released after RSV infection in “allergic” animals, IL-4, IL-5, and IL-13 are involved in the development of airway eosinophilia and AHR (Makela et al., 2003; Park et al., 2003). As previously demonstrated in mouse models, increased AHR was achieved when allergen was administered before RSV infection and also when allergen challenge was performed during the infection (Peebles et al., 2001). These findings appear to contradict the report by Aeffner and Davis, who demonstrated that intranasal infection with replication-competent RSV strain A2 for 2–8 days reduced BAL eosinophil counts and reversed airway hyperresponsiveness in a pertussis toxin-sensitive manner (Aeffner and Davis, 2012).
We set up an RSV-induced asthma exacerbation model (OVA sensitization followed by RSV Long strain infection) characterized by significantly increased level of neutrophils and eosinophils in BAL, but invariable airway reactivity compared with OVA-sensitized/challenged animals (data not shown).
In our previous study we showed the antiviral effect of anti-RSV siRNAs in vitro (Akimov et al., 2007) and in vivo in a mouse model of RSV infection (Khaitov et al., 2010). As published previously in a Russian journal, and reproduced here for the non-Russian reader (Fig. 6), we reported that siRNA targeted to the p gene of RSV significantly reduced viral load and total inflammation score in mouse lung (Khaitov et al., 2010). In addition, in a mouse model of OVA-induced asthma (Fig. 7), we found that mice intranasally treated with anti-IL-4 siRNA (100 μg /mouse) had significantly reduced levels of IL-4 expression in the lung, significantly decreased levels of AHR (at a methacholine concentration of 12.5 mg/ml), and a tendency toward reduced levels of eosinophils (data not shown) compared with mice treated with the same amount of unspecific siRNA (siGFP).
The aim of this study was to evaluate the therapeutic effect of dual antiinflammatory (anti-IL-4) and antiviral (anti-RSV) siRNAs in a mouse model of RSV-induced asthma exacerbation. We demonstrated that anti-IL-4/RSV siRNAs (100 and 70 μg/mouse, respectively), but not control siRNAs, were able to (1) inhibit pathological signs of asthma (airway inflammation and AHR), (2) diminish eosinophilia and neutrophilia in the lung, (3) suppress expression of IL-4 associated with allergic response; (4) inhibit RSV replication; and (5) increase IFN-γ expression. We believe that in the model of RSV-induced asthma exacerbation, anti-IL-4 siRNA reduced the level of eosinophils and AHR, while anti-RSV siRNA reduced the viral load and level of neutrophils. In addition, both siRNAs prevented histological alteration and total inflammation in the lung. A combination of anti-IL4 and RSV siRNAs showed a significantly higher inhibitory effect on AHR and reduced the amount of eosinophils and neutrophils compared with IL-4 and RSV siRNAs alone.
The most important bottlenecks in the development of effective RNA interference-based therapeutics are (1) effective and save delivery and (2) the optimal sequence of siRNAs (including the specificity of the siRNA and minimizing its potential off-target effects). In this study we delivered naked siRNAs to mouse lung intranasally. The systemic delivery of naked siRNA generally fails to produce a significant gene-silencing effect, but local delivery of unmodified siRNA to the lung has already been reported to be successful (Lam et al., 2012). The question of location of activity of naked siRNA in the lung remains to be answered. It is known that, in humans, RSV replicates almost exclusively in the single outermost layer of cells of the respiratory epithelium (Johnson et al., 2007). These types of cells (ciliated pseudostratified columnar epithelial cells) form the lining of the nasal passages, the trachea, and the deeper branches of the bronchial tree. We did not determine which cells took up the siRNA in our experiments, but because applying the anti-RSV siRNA had significant antiviral effects, it is likely that the siRNAs were taken up by the same type of cells. In addition, Karras and colleagues have shown that chemically modified IL-4Rα antisense oligonucleotide specifically inhibits IL-4Rα protein expression in lung eosinophils, macrophages, dendritic cells, and airway epithelium after inhalation in allergen-challenged mice (Karras et al., 2007).
To address the second issue, we designed and tested in vitro various siRNA sequences to choose the most effective one (unspecific siRNA was used as a control).
The mechanisms involved in the synergistic interaction between virus infection and allergen exposure that increases the risk of asthma exacerbations are not well characterized (Green et al., 2002; Murray et al., 2006). Thus data obtained in the current study taken together with previously published findings indicate that anti-IL-4/RSV siRNAs might represent an effective therapeutic approach to inhibit the production of IL-4 and replication of RSV in virus-induced asthma exacerbation, leading to shift of the immune response from Th2 to Th1 that results in significant reduction of airway inflammation and AHR. This is in the agreement with a review by Maes and colleagues, that targeting the IL-4/IL-13 pathway is a required supplement for standard inhaled corticosteroid-based therapies (Maes et al., 2012).
Development of novel antiviral and antiinflammatory therapies is required for the treatment and prevention of asthma exacerbations induced by respiratory viral infections. The advancement of more than 20 therapeutic siRNAs into clinical trials illustrates that RNAi-based medicines hold a pivotal place in the near-future treatment of human diseases. The therapeutic approach proposed in this study is a prospective add-on therapy for individuals with virus-induced exacerbations of severe uncontrolled asthma.
Footnotes
Acknowledgments
This work was supported by the Ministry of Education and Science of Russian Federation, contract 16.512.11.2257, from 10/08/2011. S.L.J. is supported by a Chair from Asthma UK (CH11SJ).
Author Disclosure Statement
The authors declare that no competing financial interests exist.