
Abstract
Young Rpe65-deficient dogs have been used as a model for human RPE65 Leber congenital amaurosis (RPE65-LCA) in proof-of-concept trials of recombinant adeno-associated virus (rAAV) gene therapy. However, there are relatively few reports of the outcome of rAAV gene therapy in Rpe65-deficient dogs older than 2 years of age. The purpose of this study was to investigate the success of this therapy in older Rpe65-deficient dogs. Thirteen eyes were treated in dogs between 2 and 6 years old. An rAAV2 vector expressing the human RPE65 cDNA driven by the human RPE65 promoter was delivered by subretinal injection. Twelve of the 13 eyes had improved retinal function as assessed by electroretinography, and all showed improvement in vision at low lighting intensities. Histologic examination of five of the eyes was performed but found no correlation between electroretinogram (ERG) rescue and numbers of remaining photoreceptors. We conclude that functional rescue is still possible in older dogs and that the use of older Rpe65-deficient dogs, rather than young Rpe65-deficient dogs that have very little loss of photoreceptors, more accurately models the situation when treating human RPE65-LCA patients.
Introduction
Loss of function prior to marked degeneration of photoreceptors provides a “window of opportunity” for gene augmentation therapy. The initial proof-of-principle studies in young Rpe65-deficient dogs showed that subretinally delivered recombinant adeno-associated viral (rAAV) vectors expressing either human or canine RPE65 dramatically improved photoreceptor function as assessed by ERG, vision-testing performance, and retinal-mediated visual cortex activity (Acland et al., 2001; Narfström et al., 2003; Acland et al., 2005; Aguirre et al., 2007). Similar dramatic improvements in retinal function were achieved in young Rpe65-/- and Rpe65 rd12/rd12 mice treated by rAAV-RPE65 subretinal injection (Dejneka et al., 2004; Pang et al., 2006; Roman et al., 2007; Bennicelli et al., 2008). However, none of the reported subjects in the human clinical trials have shown an ERG improvement that anywhere near matches that achieved in the dog model (Bainbridge et al., 2008; Cideciyan et al., 2008, 2009, 2013; Hauswirth et al., 2008; Maguire et al., 2008; Banin et al., 2010; Jacobson et al., 2012). The likely reason for this difference is that Rpe65-deficient dogs lose photoreceptors at a relatively slower rate than occurs in human RPE65-LCA patients.
Immunohistochemical studies recently reported from Rpe65-deficient dogs up to 18 months of age showed good preservation of both rod and cone photoreceptors (Hernandez et al., 2010). Our own studies with the Rpe65-deficient dog demonstrates slow loss of rods and M/L cones but a more rapid loss of S-cones (Mowat et al., 2013). Encouragingly, we found that gene therapy was able to preserve S-cones as had previously been reported in the mouse models (Li et al., 2011; Mowat et al., 2013). However, a very recent study looking at gene therapy outcomes in patients and Rpe65-deficient dogs showed that once there is established photoreceptor loss (i.e., in most patients treated and older Rpe65-deficient dogs), even though gene augmentation therapy can improve function, it does not preserve structure in the long-term (Cideciyan et al., 2013). Loss of photoreceptors is obviously a limiting factor for gene augmentation therapy. In addition to the loss of photoreceptors, Rpe65-deficient animals develop lipid inclusions within the retinal pigment epithelium. In the dog model, the inclusions become quite large in size, and it is conceivable that they may impact RPE health and could be another factor that might limit the success of gene augmentation therapy (Wrigstad et al., 1994; Annear et al., 2011a).
The results of gene augmentation therapy in 123 eyes from 85 Rpe65-deficient dogs had been reported in the literature at the time of writing (summarized in Supplementary Table S1). Of these, 106 eyes were treated when the dogs were less than 1 year old, 7 eyes were treated when the dogs were between 1 and 2 years old, and 10 eyes from 9 dogs were treated when dogs were older than 2 years of age at the time of treatment (Rolling et al., 2006; Aguirre et al., 2007; Le Meur et al., 2007; Annear et al., 2011b; Cideciyan et al., 2013). Of the 9 eyes treated in dogs over 2 years old, 8 were reported to show rescue (Rolling et al., 2006; Le Meur et al., 2007; Annear et al., 2011b; Cideciyan et al., 2013), while one eye of a dog 2.5 years old at the time of injection had no evidence of rescue despite the expression of RPE65 transgene in the injected region (Rolling et al., 2006). The failure of the expressed RPE65 to rescue function in this latter dog could indicate that there are changes in the affected dog retina that develop with age and may limit success of therapy. Investigation of larger numbers of older Rpe65-deficient dogs could provide important information about the factors that limit success of this therapeutic approach. The purpose of this study was to complete a larger trial of gene augmentation therapy in Rpe65-deficient dogs older than 2 years of age. We treated 13 eyes of 9 dogs aged between 2 and 6 years old and found rescue of retinal function as assessed by ERG in 12 of 13 eyes and improved vision performance in all eyes. This is particularly relevant for the treatment of RPE65-LCA patients where retinal pathology is more advanced than in the young Rpe65-deficient dogs that have been used in most of the canine gene augmentation studies.
Materials and Methods
Animals
Nine crossbred Rpe65-deficient dogs from a colony maintained at Michigan State University comparative ophthalmology laboratory were used in this study. All animals were housed under 12:12 hour light–dark cycles and cared for in compliance with the Association for Research in Vision and Ophthalmology (ARVO) statement on the Use of Animals in Ophthalmic and Vision Research. All procedures performed were approved by Michigan State University's Institutional Animal Care and Use Committee.
Recombinant AAV2/2 construct and subretinal injection
A recombinant AAV2/2 vector was used, containing the human RPE65 cDNA coding sequence driven by the human RPE65 promoter (rAAV2/2.hRPE65p.hRPE65) as previously reported (Bainbridge et al., 2008; Annear et al., 2011b). Injections were performed in the tapetal (central and dorsal) fundus consistent with the previously reported finding of greater ERG improvement in dogs treated in this location compared with those injected in the nontapetal (ventral) fundus (Acland et al., 2005). The vector titers were determined by dot blot, and the preparation diluted to provide two viral vector concentrations, 1x1011 and 1x1012 vg ml−1. In 10 eyes, subretinal injections were performed as previously described (Petersen-Jones et al., 2009). In 3 eyes, a standard pars plana three-port 23-gauge vitrectomy was performed (Accurus, Alcon, Irvine, CA). The prior vitrectomy made starting the subretinal bleb easier than if no vitrectomy was performed but was not anticipated to alter the outcome of the gene augmentation therapy. Two different volumes of viral construct were administered, 350 and 500 μl. Details of subretinal injections are provided in Table 1. All dogs were treated with topical, subconjunctival, and oral anti-inflammatory medications pre- and postoperatively as previously described (Annear et al., 2011b).
1, pigmentary spots; 2, regions of tapetal hyper-reflectivity (suggestive of retinal thinning); 3, injection problems; ac, tapetal fundus including the area centralis; ns, no significant changes; t, tapetal fundus; v, had vitrectomy prior to subretinal injection.
Eye O5-069 OD reported as a flat mount in Mowat et al., 2013.
Ophthalmic evaluation and fundus imaging
Ophthalmic examinations including slit-lamp biomicroscopy with aqueous humor flare scoring, indirect ophthalmoscopy and wide-field fundus imaging (RetCam II, Clarity Medical Systems, Pleasanton, CA) were performed pre-operatively, immediately postoperatively and at all subsequent study time points. The size of the subretinal injection bleb was calculated as a proportion of the extent of the tapetum as previously described (Annear et al., 2011b). Images were also examined to divide eyes into those in which the injection bleb encompassed the area centralis, and those where it was not included. The position of the area centralis on postoperative images was determined using the findings of Mowat et al. (2008).
Electroretinography
To assess rod and cone photoreceptor function, ERGs were recorded preoperatively and 4 months postoperatively as previously described (Annear et al., 2011b). Briefly the ERG assessment consisted of a dark-adapted intensity series from below response threshold up to a flash intensity of 2.38 log cdS/m2. This was followed by light adaptation at 30 cd/m2 for 10 min and recording of cone flicker responses to 33Hz of flashes at 0.39 log cdS/m2 superimposed on a background light of 30 cd/m2. For comparison of rod photoreceptor responses, the dark-adapted b-wave amplitude at 0.0 log cdS/m2 was used; a flash intensity that was below the response threshold for all eyes prior to treatment. The first intensity at which an ERG waveform could be recorded was noted, and the amplitude and implicit time of a- and b-wave responses were measured, shape of waveforms assessed, and intensity: response curves plotted. To evaluate cone photoreceptor responses, the amplitudes of the light-adapted 33 Hz flicker responses were measured. Flicker responses were used because it is possible that the standard rod-suppressing background light might not adequately suppress responses from the desensitized rods in Rpe65-deficient dogs, but rod recovery is too slow to allow them to respond to the 33Hz frequency flashes (Kijas et al., 2002).
Vision testing
Vision testing was performed using a previously validated four-choice vision-testing device consisting of a light-proof box with four exit tunnels, one of which is randomly selected to be open per run (Gearhart et al., 2008; Annear et al., 2012). Evaluation of individual eyes was performed by placement of an eye mask over the contralateral eye. Vision was assessed at three different light intensities, full room light, and two dim intensities as previously described (Annear et al., 2011b). The number of correct exit choices and mean time to exit the device were recorded and averaged over seven runs at each light intensity. Vision testing was performed preoperatively and 4 months postoperatively.
Retinal histology
Five eyes from 5 dogs (Table 1) that were treated at an older age (mean age 5.04 years) were analyzed following humane euthanasia at a mean of 4.6 months following injection. Injected eyes from these animals were fixed in 4% paraformaldehyde and embedded for collection of sagittal sections as previously described (Mowat et al., 2013). Immunohistochemistry was performed on 14 μm central retinal sections for the cone-specific marker human cone arrestin (hCAR LUMif [Li et al., 2003]). Sections were counterstained with the nuclear stain DAPI (Invitrogen Molecular Probes, Carlsbad, CA). Sections were imaged using a fluorescent microscope (Nikon Eclipse 80i; Nikon Instruments Inc., Melville, NY) using commercial image capture software (MetaVue; Molecular Devices, Sunnyvale, CA) using a 40× objective. Three consecutive images were taken spanning a region measuring 337.5 μm from both treated and untreated regions of each eye. Images were masked before quantification so the observer was unaware of the injection status or identity of the animal. The total number of outer nuclear layer (ONL) nuclei (4′,6-diamidine-2-phenylindole [DAPI]-positive cells in the ONL), and the number of cone nuclei (human cone arrestin [hCAR]-positive cell bodies in the ONL) per unit length of retina were counted. The number of rod photoreceptors per unit length was calculated by subtracting the number of cone nuclei from the total number of nuclei in the ONL as previously described (Mowat et al., 2013).
Statistical analysis
ERG and vision-testing outcomes were analyzed using independent samples t-tests to evaluate for differences between average preoperative and 4-month postoperative outcomes. The 4-month postoperative time period was chosen as this was the time-point of peak ERG responses after subretinal gene augmentation in a previous study (Annear et al., 2011b). Analysis was performed on b-wave amplitudes at 0.0 log cdS/m2 and 33Hz cone flickers for ERG responses, mean number of correct exit choices, and mean time to exit the device for vision-testing outcomes. To assess for correlation between age, bleb size, viral dose, and ERG and vision-testing outcomes, two-tailed Pearson analysis was performed. The ERG and vision-testing results were further compared using a one-way ANOVA between three age groups at the time of treatment; 2–3 years (n=4), 3–5 years (n=5), and 5–6 years (n=4). Photoreceptor cell body counts were assessed by a linear regression analysis using commercial statistical software (Graphpad Prism 5.0a; Graphpad, LaJolla, CA) to test whether the variables tested (rod photoreceptors per unit length, cone photoreceptors per unit length, and rod/cone ERG amplitudes) were significantly correlated. Differences in photoreceptor counts from the treated and untreated areas were analyzed using a paired t-test. Data were considered significant at p<0.05.
Results
Subretinal injection of rAAV2/2.hRPE65.hRPE65
Details of the subretinal injections are shown in Table 1. At the time of treatment, the dogs ranged in age from 2 to 6 years. Thirteen eyes of nine Rpe65-deficient dogs were given subretinal injections of the vector. All injection blebs were created in the superior fundus, and the mean proportion of the tapetal fundus occupied by the blebs was 44% (range 7–84%). Twelve of 13 eyes had blebs occupying between 32 and 84% of the tapetal fundus. Technical difficulties in establishment of an initial retinal detachment in the left eye of dog no. 05-067 resulted in a small bleb occupying only 7% of the tapetal fundus. In 5 of 13 eyes, the area centralis was included in the injected area, and in 8 of 13 eyes, the area centralis was not included within the bleb boundary; there was no significant difference in bleb size between these groups (p=0.061). All injection blebs had resolved within 1 week postoperatively.
Evaluation for ocular inflammation and fundus changes
Mild aqueous humor flare (graded as 1 on a scale of 1–4), was detected by slit-lamp biomicroscopy postoperatively in all treated eyes for up to one week. With indirect ophthalmoscopy, a small pigmented scar at the subretinal injection site and a “highwater” mark indicating the bleb boundary were observed in all treated eyes. Additionally, within the bleb boundary, 4 of 13 eyes developed small pigment spots (Fig. 1, B2 and D2) and 3 of 13 eyes developed patchy areas of tapetal hyperreflectivity suggestive of retinal thinning (Fig. 1, B2). Summary findings for treated eyes are recorded in Table 1 with representative fundus images shown in Figure 1.

Digital fundus images (Retcam II, Clarity Medical Systems) immediately postoperatively showing the extent of the subretinal bleb (1), and the fundus appearance 4 months following treatment (2).
Evaluation of retinal function
Retinal function as assessed by dark-adapted full-field flash and light-adapted cone flicker ERG showed rescue of rod and cone photoreceptor function in 12 of 13 eyes (Figs. 2 and 3 and Table 2). There was significant lowering of dark-adapted a-and b-wave response thresholds in the 12 eyes with ERG rescue (p<0.001) (Figs. 2 and 3). The lowering of threshold ranged in magnitude from approximately 1 to 3 log units (Fig. 3). The shape and timing of the waveforms at the lower light intensities following treatment were typical for canine rod responses (Fig. 2). Untreated REP65-deficient dogs never show ERG waveforms of this shape. The postoperative dark-adapted a- and b-wave amplitudes at 0.0 log cdS/m2 were significantly increased (p<0.001) (Table 2). This flash intensity was chosen as an indicator of rod rescue. Light-adapted 33Hz flicker responses, a measure of cone function, were recordable in the 12 eyes with ERG rescue (mean amplitude of 9.2 μv; range 3–18 μv), a significant change having been unrecordable in any of the eyes preoperatively (p<0.001).

Dark-adapted rod electroretinogram (ERG) montages
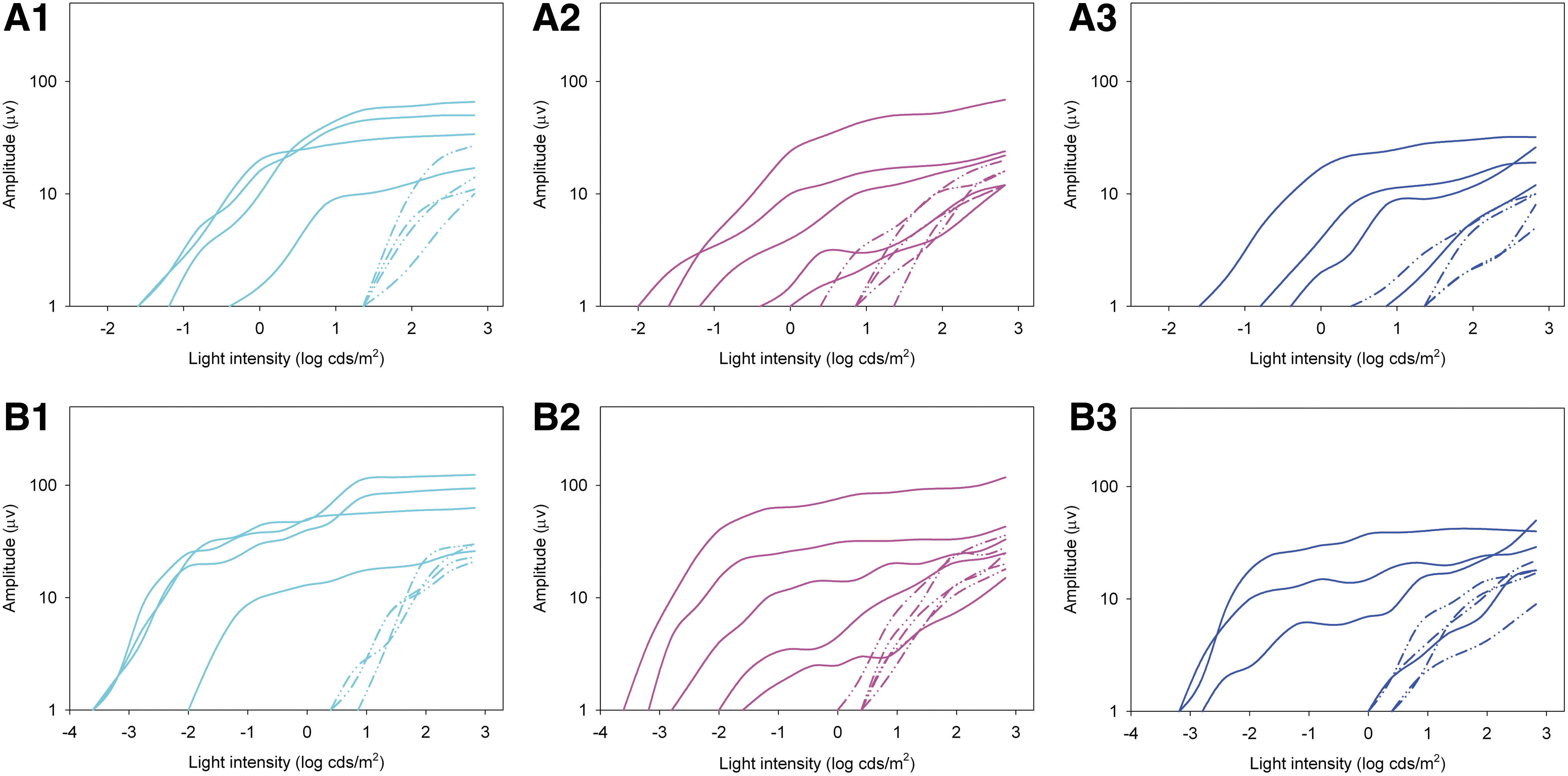
Dark-adapted intensity:response series of the a-waves
Nr, not recordable.
For further analysis, treated eyes were divided into three groups (2–3 years [n=4], 3–5 years [n=5] and 5–6 years [n=4]) and ERG outcomes compared. There was no significant difference in rod or cone ERG measurements between age groups (p-values for amplitude and threshold ranged from 0.18 to 0.47). ERG outcomes were analyzed for correlation with a number of variables including age at injection, size of the injection bleb, area centralis included within bleb boundary, viral genomes administered, volume of injection, and titer injected. Amplitude of the cone flicker responses significantly correlated with both area centralis included within bleb boundary (p=0.02) and larger bleb size (p=0.008) (Table 3). Although not reaching significance, a trend is suggested for correlation of b-wave amplitude at 0.0 log cdS/m2 with both area centralis included within the bleb boundary (p=0.053) and larger bleb size (p=0.068). No other significant correlations or trends were appreciated (Table 3).
Significant correlation.
One of 13 treated eyes (04-245 OS) failed to show evidence of ERG rescue. There was no obvious reason for failed rescue as there was a relatively large injection bleb and only small pigment foci within the bleb boundary noted on ophthalmoscopy (Fig. 1, D2). This dog was 5.2 years old at time of injection; the other eye had been previously treated (at 3.1 years old) and had shown moderate improvement in ERG measures (Table 2). It is notable that despite the lack of ERG rescue, this eye had improved dim-light vision (see below).
Evaluation of vision
Using an objective testing device, vision was assessed using two outcome parameters—exit choice and time to exit—under standard room light and at two dim-light levels. Preoperatively under standard room lighting all dogs correctly and rapidly exited the device, but at both dim-lighting intensities all dogs were effectively choosing exits randomly and were slower at exiting the device. Four months postoperatively, the vision testing under room lighting remained unchanged, but at both dim-lighting intensities all dogs showed significant improvement in both correct exit choice (p<0.001) and time to exit (p<0.001) (Fig. 4 and Table 2).

Vision-testing results at three light intensities showing the two outcome measures, number of correct exit choices of seven trials
There was no significant difference between the three age groups: 2–3 years (n=4), 3–5 years (n=5), and 5–6 years (n=4) for either correct exit choice (p=0.83) or time to exit (p=0.73) (Table 3). However, area centralis included within the bleb boundary correlated significantly with both correct exit choice (p=0.01) and time to exit (p=0.02).
Histological analysis
RPE65 expression was present in all treated eyes examined and was used to identify the treated area histologically (data not shown). We evaluated whether the degree of rod and cone ERG rescue correlated with the numbers of rod and cone photoreceptors per unit length present in the treated areas of retina in the five injected eyes analyzed (Fig. 5A and Supplementary Table S2). There was no correlation between the extent of rod b-wave rescue and rod photoreceptor number per unit retinal length (Fig. 5B; r2=0.21, p=0.43) nor between the extent of cone 33Hz flicker amplitude post-treatment and cone photoreceptor number per unit retinal length (Fig. 5C; r2=0.46, p=0.21). Comparisons of both rod and cone photoreceptor number per unit length between the treated and untreated areas of the same eye revealed no significant differences (Fig. 5D; p=0.22 for rod nuclei per unit length and p=0.10 for cone nuclei per unit length, paired t-test). The eye (04-245 OS) that had no detectable ERG rescue, but evidence of an improvement in dim-light vision, was amongst those examined (Fig. 5, “poor rescue” eye). This eye had a comparable number of photoreceptors per unit length to the eyes with successful ERG rescue (Fig. 5B and C).
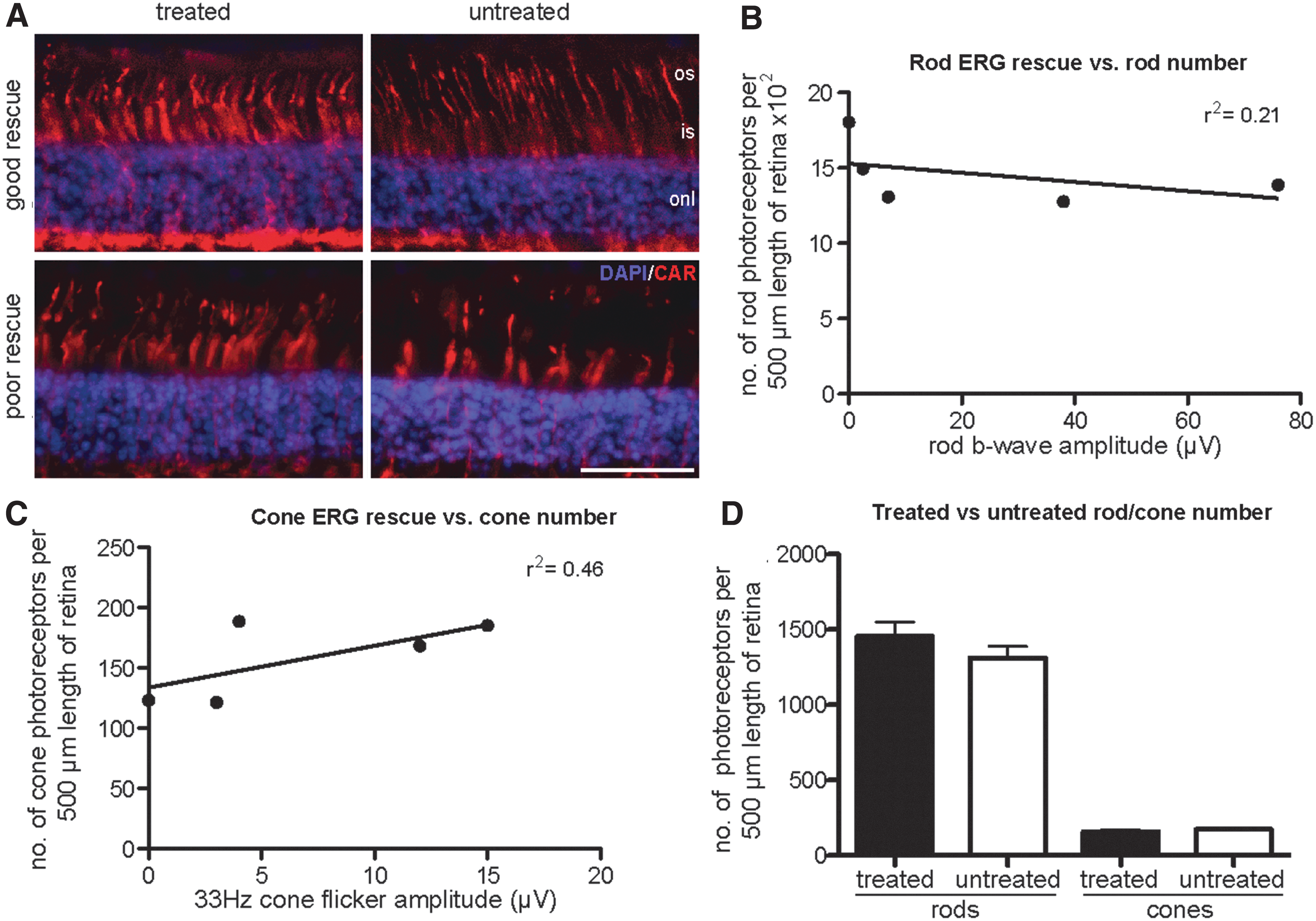
Quantification of rod and cone density in treated and untreated areas was performed using analysis of immunohistochemistry on frozen retinal sections from five treated eyes. Representative images labeled for cones (red: cone arrestin [CAR]) counterstained with the nuclear stain DAPI (blue) are shown
Discussion
In this study, we found that gene augmentation therapy using an rAAV2 vector expressing human RPE65 under control of the human RPE65 promoter improved visual function in 13 of 13 eyes of Rpe65-deficient dogs that were treated between 2 to 6 years old. Functional rescue was still possible in these dogs despite the progressive loss of photoreceptors and accumulation of lipid droplets in the retinal pigment epithelium that is known to develop with age in this model (Wrigstad et al., 1992; Wrigstad et al., 1994; Annear et al., 2011a; Mowat et al., 2013).
Prior to this study, the results of gene augmentation therapy had been reported in 10 eyes of Rpe65-deficient dogs older than 2 years of age. These reports included one eye of a dog injected at 2.5 years old that failed to show ERG or vision-testing rescue, although, subsequently it was shown by immunohistochemistry to have appropriate RPE65 transgene product in the RPE, suggesting that failure was not due to technical injection issues or failure of RPE transduction by the viral vector (Rolling et al., 2006; Le Meur et al., 2007). This could indicate that this dog was close to the upper age limit for treatment success. However, the results of our current study suggest that this is not the case with all eyes treated showing improvement in vision at dim-light levels using an objective vision-testing measure. Additionally, 12 of the 13 eyes treated had an improvement in both rod and cone function as assessed by ERG; with lowering of dark-adapted response thresholds, the appearance at the lower flash intensities of waveforms with a shape typical of rod-mediated responses increases in waveform amplitudes and recordable cone flicker responses (prior to therapy, cone flicker responses were not recordable from these eyes). There was a significant correlation between the degree of rod and cone rescue.
Over the range of ages (2 to 5.9 years) tested in this study, we did not find a correlation of ERG outcomes (b-wave amplitude at 0.0 log cdS/m2 and cone flicker amplitudes) with age. This suggests that within this range of ages sufficient rescuable photoreceptors remain to provide functional rescue detectable by the assessment methods that can be used in dogs. The only significant positive correlations with ERG outcomes were a correlation between improved cone flicker amplitudes and with inclusion of the area centralis in the injected area and also with a larger treated area. Inclusion of the area centralis was also positively correlated with improved vision-testing performance. The link between ERG and vision-testing outcomes and inclusion of the area centralis in the treated area may be because the central retina in the normal canine has a greater number of photoreceptors per unit area than the peripheral retina (Mowat et al., 2008), and in Rpe65-deficient dogs there is relative preservation of the outer nuclear layer in the central and superior retina with a more severe degeneration peripherally (Wrigstad et al., 1994; Cideciyan et al., 2013; Mowat et al., 2013). It is also possible that although the ERG stimulus used a Ganzfeld bowl and the pupils were fully dilated, the degree of light stimulation of the central retina might still be greater than that of the peripheral retina. The lack of correlation of outcome with age was unexpected as our previous studies showed that the older dogs will have lower numbers of surviving photoreceptors at the time of treatment (Mowat et al., 2013). However, one should consider here the relatively low numbers of dogs used and a possible masking effect of other variables such as dose, volume injected, size of the subretinal injection, and precise region of the retina treated; it is difficult to control the direction of spread of the subretinal injection bleb and therefore difficult to standardize among animals.
Histological analysis of five of the treated eyes was performed to measure rod-and-cone photoreceptor numbers in treated and untreated areas. Although there was some variation in rod-and-cone numbers between the eyes examined, this did not correlate with ERG rescue. This would suggest that loss of photoreceptors was not a limiting factor for success of gene therapy in the age range of Rpe65-deficient dogs investigated in this study. Additional factors to consider that might influence the degree of rescue and could be the basis of future gene therapy studies include the health of both the remaining photoreceptors and the RPE. The effect of therapy on preservation of photoreceptors could not be evaluated on the five eyes examined histologically because of the relatively short time that had elapsed following treatment. Other animals in this study are being maintained for longer-term assessment of rescue. The longer-term assessment of photoreceptor preservation in dogs treated at an older age and the assessment of adjunctive therapy such as anti-apoptotic agents and growth factors aimed to preserve photoreceptors is particularly relevant in view of the recent report by Cideciyan et al. indicating in both human and dog that gene therapy treatment after the retina has entered a phase of photoreceptor degeneration can improve function but does not halt further progression of retinal degeneration (Cideciyan et al., 2013).
One of the older dogs in the current study (04-245-OS aged 5.2 years at the time of injection) is of particular interest because there was no detectable rescue of ERG function, although dim-light vision was improved. Clearly it requires fewer functional photoreceptors to make a detectable difference in visual function compared with the number of summed photoreceptor responses required to make a difference in recordable full-field flash ERG. Similar findings are seen in dogs with other retinal degenerative conditions that have useful vision, and yet, an ERG response is not recordable (Petersen-Jones, personal observations). There is a similar finding in the RPE65-LCA patients treated by gene augmentation therapy where even in those patients with the greatest improvement in visual function the full-field flash ERG was not improved (Bainbridge et al., 2008; Cideciyan et al., 2008, 2009 , 2013; Hauswirth et al., 2008; Maguire et al., 2008; Banin et al., 2010; Jacobson et al., 2012). In the current study, the eye with the lack of ERG rescue had one of the larger subretinal injection blebs, a factor which overall showed a positive correlation with cone ERG rescue. Furthermore, the animal was treated using the same batch of vector as the other eyes, and the eye did not develop funduscopic lesions such as tapetal hyperreflectivity in the injection area that would indicate marked retinal thinning; and indeed, histology revealed that there was RPE65 expression and good preservation of photoreceptors in this eye (as assessed by counting rod and cone photoreceptor nuclei per unit length of retina Figure 5A, B & C). This would suggest that there may be factors in the retina of Rpe65-deficient animals other than the numbers of surviving photoreceptors that influence outcome. Additional studies of older dogs at an age in which treatment is less successful would be valuable in the quest to identify factors that may limit treatment success.
The treatment procedure itself is not without potential deleterious effects and indeed ophthalmoscopically detectable fundus changes developed in the region of the subretinal injection bleb in some animals. These included patchy tapetal hyper-reflectivity, which is a clinical indication of retinal thinning, and development of pigment spots that possibly indicated changes in the RPE. Patchy retinal thinning was recorded in a previous gene augmentation therapy trial in Rpe65-deficient dogs (Le Meur et al., 2007). The current generation of viral vectors used in retinal gene therapy require delivery by subretinal injection to transduce the retinal pigment epithelium. Creating a retinal detachment has deleterious effects on the retina, with studies indicating that there is significant induction of cytokines associated with apoptotic pathways within 72 hr of retinal detachment (Nakazawa et al., 2006; Mandal et al., 2011). It is not clear whether the retinal changes that developed in the injected area are the result of the physical detachment of the retina, a result of the viral vector and expression of the transgene, or a combination of both. Patchy retinal degeneration has also been recorded in a reporter gene study in normal dogs and developed only in the eyes receiving the highest titers; the timing of the degeneration suggested that this was an immune reaction to either the high levels of the expressed transgene (green fluorescent protein) or the high concentration of rAAV particles (Beltran et al., 2010).
There are substantial differences between the pathology in patients and in dogs and between the results of gene therapy in these species that make a direct comparison complex. One major difference is the rod photoreceptor degeneration in the patients, which is not replicated to the same extent in dogs. A second difference is the fact that rescue of ERG activity, the primary outcome measure in the animal studies, has not been observed in any of the subjects in any of the current clinical trials. Nevertheless, as shown in this study in dogs, the clinical trial studies do not demonstrate a substantially worse treatment effect in young adult subjects compared with children. This suggests that rescue of RPE function and retinoid cycling may be feasible to a similar extent in young and older subjects, and that any limitations on the ability to treat even older subjects successfully are the likely to be the result of photoreceptor degeneration. The current study shows that the use of older Rpe65-deficient dogs in gene augmentation therapy will be valuable in investigating the factors, other than the number of surviving photoreceptors, that limit the success of such therapy. An important feature that makes the canine Rpe65-deficient model particularly relevant compared to mouse models is the presence in the dog of an area centralis and visual streak. These retinal regions have greater photoreceptor densities compared with the peripheral retina (Mowat et al., 2008). This more closely reflects the photoreceptor distribution in humans than murine models that do not have regions of higher photoreceptor density. The use of older Rpe65-deficient dogs in gene augmentation therapy will not only allow the investigation of factors that limit functional rescue but also enable studies of protocols designed to improve the preservation of photoreceptors when the retina is already partially degenerate at the time of treatment.
We conclude that the use of Rpe65-deficient dogs older than 6 years of age for testing gene augmentation therapy may more closely reflect the challenges that are faced in treating human subjects. Although important for proof-of-concept and other preclinical studies, the use of young Rpe65-deficient dogs does not accurately reflect the clinical situation in which subjects are being treated when there are potentially fewer responsive photoreceptor cells remaining.
Footnotes
Acknowledgments
This work was supported by the British Retinitis Pigmentosa Society, The Midwest Eye Banks and Transplantation Center Research Program, Michigan State University College of Veterinary Medicine Purebred Dog Endowment Fund, and Myers-Dunlap Endowment for canine health. J.W.B. is a Wellcome Trust Advanced Fellow. R.R.A. and J.W.B. are investigators at The NIHR Centre for Ophthalmology at UCl and Moorfields Eye Hospital. We acknowledge Dr. Cheri Johnson for assistance with our dog colony breeding programs.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.