
Abstract
Adeno-associated virus (AAV) is an ideal choice for gene delivery; however, its further development has been limited owing to its low transduction efficiency. DNA-damaging agents can improve AAV-mediated transgene expression. Hyperthermia, as one of the oldest documented tumor treatment modalities, can cause DNA damage as well. However, combined treatment consisting of hyperthermia and AAV-mediated gene therapy has not been reported yet. In this work we investigated whether therapy consisting of AAV-mediated pigment epithelium-derived factor (PEDF) delivery combined with hyperthermia has synergistic antitumor effect on established solid tumors. We produced the recombinant AAV encoding PEDF (rAAV-PEDF). The therapeutic effect of rAAV-PEDF plus hyperthermia was evaluated in a subcutaneous fibrosarcoma mouse model, and the possible mechanism of antitumor effect was investigated. We found that rAAV-PEDF could infect a murine fibrosarcoma cell line (Meth-A) and express PEDF protein with bioactivity in vitro. In addition, in vivo experiments suggested that the combination of rAAV-PEDF with hyperthermia could significantly suppress tumor growth and prolong survival time of treated mice. Immunofluorescence studies indicated that the combination therapy could inhibit angiogenesis and induce apoptosis in tumor tissues. An immunohistochemistry assay of tumor tissue showed that PEDF expression in the combined treatment group was significantly higher than in the rAAV-PEDF group, which implied that hyperthermia could improve the expression of PEDF protein in vivo. No significant differences were observed in each group by hematoxylin–eosin staining of major organs, serum chemistry test, and complete blood assay. These results indicate that the combination of rAAV-PEDF with hyperthermia has synergistic therapeutic effects on established solid tumors, with no side effects. In addition, hyperthermia could improve AAV-mediated transgene expression, which suggests that hyperthermia could serve as a promising adjunctive therapy for AAV-mediated gene therapy.
Introduction
A
Angiogenesis is essential for tumor growth and metastasis; therefore, inhibition of tumor vascularization is considered a feasible approach for cancer therapy (Folkman, 1971; Weis and Cheresh, 2011). Because the universality and efficacy of this approach have been demonstrated on the basis of both experimental and clinical data, antiangiogenic therapy has evolved into an integral component of current standard anticancer treatment (Abdollahi and Folkman, 2010). With the development of antiangiogenic therapy, more antiangiogenic agents have been found, such as pigment epithelium-derived factor (PEDF), angiostatin, endostatin, TNP-470, thalidomide, and so on (Ma et al., 2002). PEDF, a 50-kDa secreted glycoprotein that belongs to the serine protease inhibitor superfamily, is widely expressed in most tissues of the body. It was first identified as a product of pigmented cells of the neural retina (Tombran-Tink and Barnstable, 2003a). In addition to its neurotrophic and neuroprotective functions, PEDF has been shown to be a more promising angiogenesis inhibitor than any other known endogenous angiogenic inhibitors, including endostatin, angiostatin, and thrombospondin-1 (Tombran-Tink and Barnstable, 2003b; Wu et al., 2012). PEDF specifically inhibits only new blood vessel growth and spares the preexisting vasculature, and has no direct effects on normal blood vessels in the body; this makes it an appealing candidate as an inhibitor of tumor angiogenesis (Ek et al., 2006).
The success of gene therapy is not only highly dependent on the selected gene but also on efficient and safe gene delivery vectors (Verma and Somia, 1997; Li and Huang, 2000). An ideal gene therapy vector should be nontoxic, nonimmunogenic, easy to produce, and efficient in protecting and delivering the therapeutic gene to target cells (Chung et al., 2011). Adeno-associated virus (AAV), as an ideal gene vector, has many of the desirable features of other viral vectors, such as retroviruses and adenovirus, but without their potential drawbacks for application in gene therapy (Samulski et al., 1991). AAV is a simple, nonpathogenic, and replication-defective single-stranded DNA virus. It can infect a variety of cell types, and its DNA can integrate preferentially into human chromosome 19 (Samulski et al., 1991). Recombinant AAV (rAAV) has the ability to infect both dividing and nondividing cells in vitro and in vivo. Moreover, it can maintain efficient and long-term gene transduction in vivo without cytotoxicity and cellular immune responses, the hallmark features of AAV-mediated gene transfer (Xiao et al., 1996; Kay et al., 2001).
Although AAV has many advantages, its low transgene expression efficiency, which is due mainly to limited second-strand synthesis in vivo, is still an obstacle to its application (Ferrari et al., 1996; Fisher et al., 1996; Jiang et al., 2011). Previous studies have proven that some DNA-damaging agents, such as cisplatin, doxorubicin, tritiated thymidine, ultraviolet light, and γ irradiation, could significantly improve the transduction efficiency of AAV (Alexander et al., 1994; Jiang et al., 2011). Hyperthermia, as an adjuvant therapy, usually combined with conventional therapy such as chemotherapy, radiotherapy, and surgical resection, can cause DNA damage as well (Eppink et al., 2012). However, combined treatment consisting of hyperthermia and AAV-mediated gene therapy has not been reported yet, and whether they have synergistic effects on antitumor therapy remains unclear. Therefore, in this study, we constructed an rAAV encoding PEDF (rAAV-PEDF) and evaluated the therapeutic effects of rAAV-PEDF combined with hyperthermia in a mouse fibrosarcoma model. In addition, in vitro correlation between DNA damage of tumor cells and PEDF expression was investigated.
Materials and Methods
Cells and animals
Meth A is a transplantable 3-methylcholanthrene-induced sarcoma of BALB/c origin passaged as an ascetic tumor. AAV-293 cells were purchased (240073; Stratagene/Agilent Technologies, Santa Clara, CA) and used to package AAV particles. AAV-293 cells were grown in Dulbecco's modified Eagle's medium (GIBCO DMEM; Life Technologies, Carlsbad, CA) with 10% fetal bovine serum (GIBCO FBS; Life Technologies), and Meth-A cells were cultured in GIBCO RPMI 1640 (Life Technologies) supplemented with 10% FBS. According to a standard procedure (Jaffe et al., 1973), primary human umbilical vein endothelial cells (HUVECs) were isolated from human umbilical cord veins donated by a puerpera, and then maintained in complete EGM-2 medium (Lonza Group, Basel, Switzerland). HUVECs were used for experiments after passages 3–5.
Female BALB/c mice, age 6 to 8 weeks, were purchased from the Laboratory Animal Centre of Sichuan University (Chengdu, China) and kept under specific pathogen-free conditions. After being allowed to acclimate for 1 week, the mice were used for experiments. All animal procedures were performed in accordance with the protocol approved by the Institutional Animal Care and Treatment Committee of Sichuan University.
Production of rAAV-PEDF
Production of rAAV-PEDF (serotype 2) was carried out according to previous work (Wu et al., 2012). In brief, human PEDF cDNA was first amplified from the pBLAST49-PEDF vector (purchased from InvivoGen [San Diego, CA] and stored in our laboratory) by PCR, and then inserted into the pAAV-MCS expression vector (commercial AAV helper-free system; Stratagene/Agilent Technologies). According to the instruction manual, three plasmids, namely pAAV-PEDF plasmid, pAAV-RC plasmid, and pHelper plasmid, were cotransfected into AAV-293 cells to produce rAAV-PEDF particles. The control vector rAAV-null was produced in a similar manner. Purification of all rAAV vectors were done as described previously (Blouin et al., 2004; Grieger et al., 2006). The appearance of rAAV particles was observed by transmission electron microscopy (TEM) (H-600; Hitachi, Tokyo, Japan). rAAV particle purity was evaluated by electrophoresis in 10% sodium dodecyl sulfate (SDS)–polyacrylamide. Particle titers were measured by dot-blotting assay. All particles were stored at 4°C in phosphate-buffered saline (PBS) before use.
In vitro expression of PEDF
Briefly, Meth-A cells were infected with rAAV-null or rAAV-PEDF at a multiplicity of infection (MOI) of 105 infectious particles per cell in serum-free medium, and with normal saline (NS) as noninfection control. Four hours later, the medium was replaced with complete medium for further culture. After 72 hr, conditioned media and cells were collected. The conditioned media were concentrated by filtration (5 kDa; EMD Millipore, Billerica, MA), and then PEDF levels were determined by ELISA. A commercial PEDF ELISA kit (Chemicon, Temecula, CA) was used according to the manufacturer's instructions.
The cells were lysed, and protein concentrations were determined with a modified Lowry protein assay kit (Pierce Biotechnology/Thermo Scientific, Rockford, IL) and equalized before loading. The samples were loaded onto 10% SDS–polyacrylamide gels and transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA) for Western blotting analysis. A mouse anti-human PEDF monoclonal antibody (R&D Systems, Minneapolis, MN) was used as the primary antibody, and horseradish peroxidase (HRP)-conjugated secondary antibody (Abcam, Cambridge, UK) was used as the secondary antibody. Protein bands were detected with an enhanced chemiluminescence (ECL) detection system (Pierce Biotechnology/Thermo Scientific).
In vitro proliferation assay
To determine the effect of PEDF derived from rAAV-PEDF-infected Meth-A cells on HUVECs, a proliferation assay was conducted as previously described (Wu et al., 2012). Briefly, HUVECs (1×104 per well) were plated into 96-well plates and incubated for 48 hr. The conditioned media from Meth-A cells infected with rAAV-PEDF, rAAV-null, or NS were collected, 2-fold serially diluted (from 1:2 to 1:64), and then added into each well in triplicate. After further incubation for 48 hr, the proliferation of HUVECs was measured by the methyl thiazolyl tetrazolium (MTT) method.
In vitro wound-healing assay
HUVECs (5×104 per well) were plated into 24-well plates and allowed to grow to 80% confluence. The cells were then wounded with pipette tips and washed gently twice with PBS. Conditioned media from the three groups mentioned previously (rAAV-PEDF, rAAV-null, and NS) were collected and diluted with fresh EGM-2 medium (1:32), and then these media (1 ml/well) were added to the wounded cell monolayers. After incubation for 18 hr, images were taken with a digital camera (Olympus, Tokyo, Japan). Migrated HUVECs were counted manually in three representative fields in each well, and the percentage of inhibition was expressed relative to the NS group (control) as 100%.
In vitro Transwell invasion assay
Transwell plates (24-well, 8.0 μm, polyethylene terephthalate [PET]; Corning Life Sciences, Lowell, MA) were coated with 25% Matrigel (BD Biosciences, San Jose, CA). After Matrigel polymerization, each bottom chamber was filled with 500 μl of complete EGM-2 medium and each upper chamber was filled with 90 μl of HUVEC suspension (2×104 cells per well). Ten microliters of conditioned media from one of the three groups mentioned previously (rAAV-PEDF, rAAV-null, or NS) was added into the upper chamber, respectively. The cells were allowed to migrate for 24 hr, nonmigrated cells were removed with cotton swabs, and migrated cells were fixed 100% methanol and stained with 0.05% crystal violet. Images were taken with a digital camera attached to an inverted microscope (Olympus), and the migrated HUVECs were counted manually in three representative fields in each well. The percentage of migrated cells was expressed relative to the NS well, which was set at 100%.
In vitro capillary-like tube formation assay
The tube formation assay was conducted as previously described, with some modifications (Pang et al., 2009; Wu et al., 2012). Briefly, prechilled Matrigel (BD Biosciences) was pipetted into prechilled 24-well plates on ice (50 μl/well). After Matrigel was polymerized at 37°C, HUVECs suspended in EBM-2 medium were seeded onto the Matrigel (3×104 cells per well). Conditioned media from the three groups mentioned previously (rAAV-PEDF, rAAV-null, or NS) were then pipetted into the wells, respectively. After incubation for 6 hr, the cells were photographed with an inverted microscope (Olympus). Three independent experiments were performed.
In vivo synergistic antitumor effects
Viable Meth-A cells were harvested and then injected subcutaneously into the right hind limb of each mouse (each mouse, 5×105 cells). After 4 days, when the tumors were palpable, the mice were randomly divided into the following 5 groups: NS, rAAV-null, rAAV-PEDF, hyperthermia (Hyper), and rAAV-PEDF+Hyper groups (12 mice per group, of which 6 mice were used to observe tumor growth and the other 6 mice were used to observe survival time).
Gene therapy was performed when tumor volume reached about 5×5 mm. rAAV-null, rAAV-PEDF, and rAAV-PEDF+Hyper groups received intratumoral injections of rAAV-null or rAAV-PEDF (2.5×1010 vector genomes [VG]/50 μl/mouse), whereas the NS group was given 50 μl of NS. On days 3 and 10 after gene therapy, hyperthermia was conducted in the Hyper group and the rAAV-PEDF+Hyper group. The tumor-bearing legs of mice in the Hyper and rAAV-PEDF+Hyper groups were heated by immersion in a water bath at 42±0.5°C for 1 hr, whereas those in other groups were immersed in a water bath at 37±0.5°C for 1 hr (Song, 1984).
In the tumor growth inhibition assay (six mice per group), tumor volume and body weight were monitored every 3 days. Tumor volume was calculated according to the following formula: tumor volume (mm3)=0.5×length×width2. On day 25 after cell inoculation, when the mean tumor volume reached nearly 4000 mm3, the mice were killed. Serum samples, tumor tissues, and major organs were collected for further study. To further investigate antitumor activity in the rAAV-PEDF+Hyper group, survival times of the mice were observed (six mice per group). When the mice became moribund, they were killed by cervical dislocation.
Immunofluorescence analysis
Microvessel density (MVD) and apoptosis in tumor tissues were detected by immunofluorescence analysis, and the staining process was carried out according to previously published methods (Wu et al., 2012). Briefly, frozen sections of tumors were fixed in prechilled acetone. Intratumoral microvessels were stained with a rat anti-mouse CD31 polyclonal antibody (diluted 1:50; BD Biosciences) and a rhodamine-conjugated second antibody (Abcam). For quantitative assessment of apoptosis, terminal deoxynucleotidyltransferase-mediated nick-end labeling (TUNEL) was done according to the protocols of an in situ cell death detection kit (DeadEnd fluorometric TUNEL system; Promega, Madison, WI) (Wu et al., 2012). In the tissue sections, at least four equal-sized fields were randomly chosen and analyzed (field magnification, ×200) by fluorescence microscopy (Leica Microsystems, Wetzlar, Germany). MVD was calculated by counting the number of microvessels in each field, and the apoptotic index was determined as the ratio of apoptotic cell number to the total cell number in each field. Data are shown as the mean apoptotic index±SD for each group.
Histology and immunohistochemistry analysis
To further observe histological changes, all collected tissues including tumors, heart, liver, spleen, lungs, and kidneys were fixed in 4% paraformaldehyde for 48 hr and embedded in paraffin. The tissues were then sectioned and stained with hematoxylin and eosin (H&E) according to standard methods (Fischer et al., 2008).
To detect PEDF protein expression in tumor specimens, frozen sections of tumors were sectioned (thickness, 3–5 μm) and fixed in prechilled acetone for 30 min. After being washed with PBS, the tissues were incubated with primary antibodies (diluted 1:50, a mouse anti-human PEDF monoclonal antibody; R&D Systems) at 4°C overnight. In subsequent steps, the assay was performed with a Histostain-Plus kit (SP-9002; ZSGB-BIO, Beijing, China) according to the manufacturer's protocol.
ELISA
According to previously published methods (Wu et al., 2012), a commercial PEDF ELISA kit (Chemicon, Temecula, CA) was used to detect PEDF expression level in serum. Briefly, all collected serum samples were prediluted and then added to wells of a precoated ELISA plate. After a series of reactions, absorbance at 450 nm was measured with an ELISA reader (680; Bio-Rad).
Evaluation of possible adverse effects
In addition to histopathological observation of major organs (including heart, liver, spleen, lungs, and kidneys), blood samples were collected for complete blood count testing with an automatic blood analyzer (Abbott, Abbott Park, IL). In addition, serum samples were collected for liver and kidney function analysis with an automatic biochemical analyzer (AU7020; Hitachi) on day 25 after tumor inoculation.
Determination of DNA damage and PEDF expression
To determine the relation between DNA damage and AAV-mediated PEDF expression in vitro, Meth-A cells were divided into the following five groups: NS, rAAV-null, Hyper, rAAV-PEDF, and rAAV-PEDF+Hyper groups. In the rAAV-PEDF+Hyper group, Meth-A cells were infected with rAAV-PEDF at an MOI of 105 infectious particles per cell in serum-free medium for 4 hr, and then the medium was replaced with complete medium. After 24 hr, the cells were incubated at 42°C for 2 hr, and then cultured at 37°C for another 48 hr. Conditioned media were collected and concentrated by superfiltration (5 kDa; EMD Millipore), and PEDF levels were determined by ELISA. In the rAAV-PEDF group, cells were infected with rAAV-PEDF and treated as described previously without hyperthermia. In the Hyper group, cells were treated as described for the rAAV-PEDF+Hyper group but without the infection process. In the rAAV-null group, cells were infected with rAAV-null and treated without hyperthermia. In the NS group, cells were infected with NS as control, and treated without hyperthermia.
DNA damage in each group was determined by comet assay immediately after hyperthermia was done (Speit and Rothfuss, 2012). Briefly, frosted slides were coated with 1.5% normal melting point agarose (in PBS) and air dried. A cell suspension (20 μl, ∼12,000 cells) was mixed with 100 μl of low melting point agarose (0.5% in PBS) and added to the slides. After being dried, slides were lysed (pH 10) at 4°C for about 2 hr. Slides were then processed by alkali denaturation for 1 hr and electrophoresis (1 V/cm; pH>13) for 30 min at 4°C. One hundred randomly selected cells in each slide were stained with propidium iodide (PI) and analyzed. DNA migration was determined by measuring the percentage of DNA in the tail, using image analysis (Comet Assay IV; Perceptive Instruments, Haverhill, UK).
Statistical analysis
Statistical analysis was carried out with SPSS 15.0 software (IBM/SPSS, Chicago, IL). Comparisons of tumor volume and tumor weight were performed by one-way analysis of variance (ANOVA). Survival curves were generated by the Kaplan–Meier method and statistical significance was determined by Mann–Whitney U testing. A p value less than 0.05 on a two-tailed test was considered statistically significant.
Results
Production and identification of rAAV-PEDF
To produce rAAV-PEDF, PEDF cDNA obtained from the pBLAST49-PEDF vector was first cloned into the pAAV-MCS vector, which contains two inverted terminal repeats (ITRs) (Fig. 1A). The recombinant expression plasmid was then cotransfected into AAV-293 cells with the pAAV-RC plasmid and pHelper plasmid to produce rAAV-PEDF particles. Figure 1B shows a schematic overview of the production of recombinant AAV particles, using the AAV helper-free system. To visualize the morphological characteristics of rAAV-PEDF particles, an aliquot of purified particles was observed by TEM after negative staining. As presented in Fig. 1C, these particles were nonenveloped and icosahedral in shape; a large proportion of the AAV particles appeared to be full, with a few intermediates and empty particles rarely seen (Fig. 1C). The titers of rAAV-PEDF and rAAV-null measured by dot-blotting assay were 4×1012 and 1×1012 VG/ml, respectively. These results indicated that the rAAV-PEDF particles produced were of high concentration and purity.
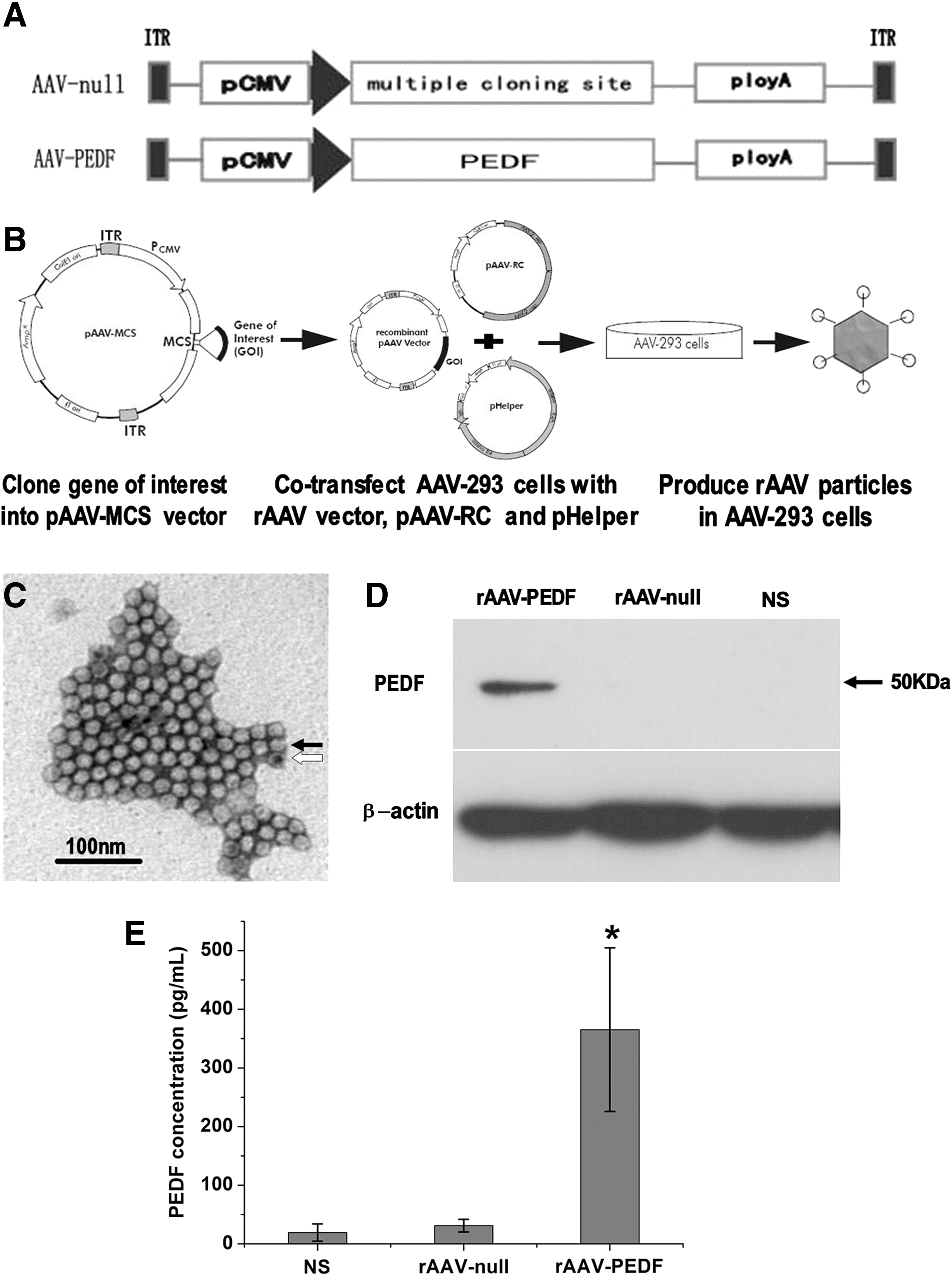
Production scheme and characterization of rAAV-PEDF.
In addition, Fig. 1D shows the PEDF expression in Meth-A cells treated with rAAV-PEDF, rAAV-null, or NS. As shown in Fig. 1D, only one band of 50 kDa, which coincided with the PEDF protein, was observed in rAAV-PEDF-infected cells, and no PEDF protein was detected in the control groups. Furthermore, Fig. 1E shows characterization of the PEDF secreted from rAAV-PEDF-transfected Meth-A cells. Conditioned media from rAAV-null-infected Meth-A cells and NS-treated Meth-A cells were used as a control. The PEDF concentration in the rAAV-PEDF group (365.3±139.4 pg/ml) was significantly higher than in the rAAV-null group (31.0±10.8 pg/ml; p<0.001) or the NS group (19.3±14.7 pg/ml; p<0.001). These results suggested that rAAV-PEDF particles could infect Meth-A cells, transfer the PEDF gene into cells, and produce secretory PEDF protein.
Biological activity of PEDF produced by rAAV-PEDF in vitro
It is well known that the potential of PEDF as a tumor inhibitor stems from its potential antiangiogenic activity (Ek et al., 2006). Endothelial cell migration and invasion are essential for angiogenesis and also play critical roles in tumor growth. To investigate whether PEDF protein produced from rAAV-PEDF also possesses antiangiogenic activity, the conditioned media from Meth-A cells infected with rAAV-PEDF or rAAV-null were collected, and the conditioned medium from untreated cells was used as a control. We then investigated the effects of those conditioned media on HUVECs in vitro by means of the wound-healing assay and Transwell invasion assay, respectively. As shown in Fig. 2A, B, and D, the migration and invasion of HUVECs were inhibited by secretory PEDF protein from Meth-A cells infected with rAAV-PEDF, but no significant difference was observed between the rAAV-null and NS groups.
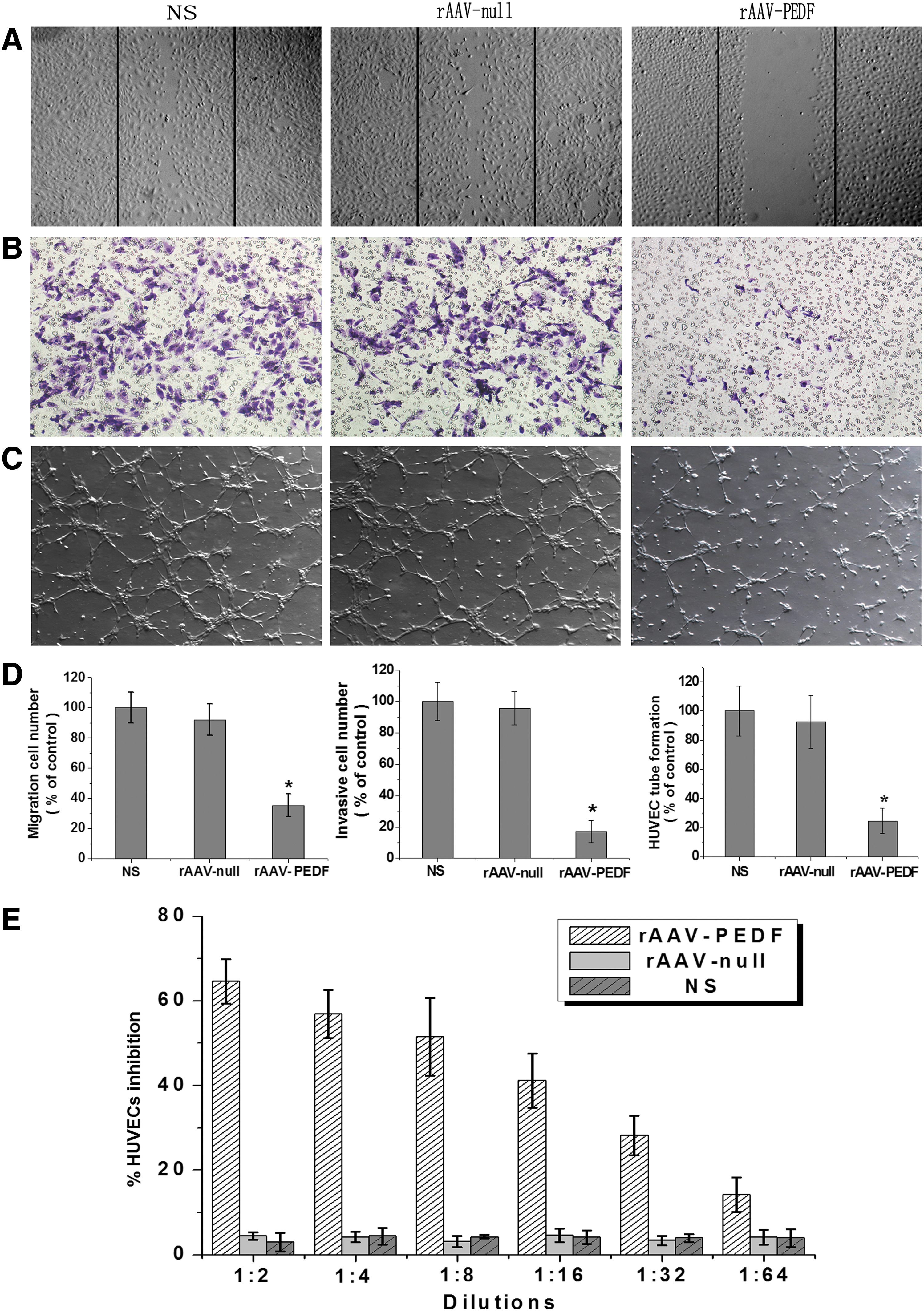
Biological activity of PEDF produced by rAAV-PEDF in vitro. To investigate the biological activity of PEDF produced by rAAV-PEDF, conditioned media from Meth-A cells infected with rAAV-PEDF or rAAV-null were collected after 72 hr. The conditioned medium from untreated cells was used as a control. The effects of those conditioned media on HUVECs were then researched.
Angiogenesis is a complicated process including various kinds of cells, but tube formation of endothelial cells is one of the key steps (Patan, 2004). On the basis of this point, we carried out a two-dimensional Matrigel assay to assess the potential effects of conditioned media on the capillary-like structure formation of HUVECs. We found that the percentage of tubular formation in the rAAV-PEDF group was much less than in the NS and rAAV-null groups (Fig. 2C and D). In addition, HUVEC proliferation in vitro was measured by the MTT method. According to Fig. 2E, conditioned medium from cells infected with rAAV-PEDF could inhibit HUVEC proliferation. Moreover, the inhibitory rate decreased with increased dilution. These results revealed that PEDF protein produced by rAAV-PEDF was highly bioactive and could block angiogenesis in vitro by inhibiting the migration, invasion, tubular formation, and proliferation of endothelial cells.
Inhibition of tumor growth in vivo
We subsequently evaluated the therapeutic effects of rAAV-PEDF and hyperthermia on solid tumor in vivo. Figure 3A shows representative photographs of tumors on day 25 after Meth-A cell inoculation. It reveals that single rAAV-PEDF or hyperthermia treatment resulted in a remarkable reduction of tumor growth compared with the control group, and that combination therapy displayed superior antitumor efficacy. Figure 3B shows that compared with the mean tumor volume of the NS group (3697.0±247.05 mm3), no obvious difference was observed in the rAAV-null group (3603.02±314.08 mm3). Tumor volumes in the Hyper group (1661.42±194.04 mm3), rAAV-PEDF group (1088.35±293.82 mm3), and rAAV-PEDF+Hyper group (347.27±254.97 mm3) were smaller than in the NS group (p<0.001). Meanwhile, compared with the Hyper group and rAAV-PEDF group, the rAAV-PEDF+Hyper group showed significant inhibitory effects (p<0.001), indicating that combination therapy was superior to monotherapy.

Tumor growth inhibition and survival advantage in mice. Meth-A cells were injected subcutaneously into the right hind limb of each mouse (5×105 cells for each mouse). After 4 days, when tumors were palpable, the mice were randomly distributed into the following 5 groups: NS (solid squares), rAAV-null (solid circles), Hyper (solid triangles), rAAV-PEDF (solid inverted triangles), and rAAV-PEDF+Hyper groups (solid rhombus) (12 mice per group, of which 6 mice were used to observe tumor growth and the other 6 mice were used to observe survival time). When tumor volume reached about 5×5 mm, gene therapy was performed by intratumorally injecting rAAV-null or rAAV-PEDF (2.5×1010 VG/50 μl/mouse); the NS group was given 50 μl of NS. On days 3 and 10 after gene therapy, hyperthermia was conducted. Tumor growth and body weight were monitored every 3 days. On day 25 after cell inoculation, the mean tumor volume reached nearly 4000 mm3, mice became moribund, and were killed.
Besides, as shown in Fig. 3C, the median survival time of the combination therapy (>100 days) was significantly longer than that in NS group (40 days), rAAV-null group (44 days), Hyper group (81 days), or rAAV-PEDF group (62 days), respectively. These results indicated that compared with NS group, either single hyper therapy or single gene therapy could extend survival time, but combination therapy had a better therapeutic effect. Notably, on day 15 after Meth-A cell inoculation, the weight of mice in NS and rAAV-null groups increased aggressively, and that of the mice received rAAV-PEDF therapy increased slightly. However, the body weight of mice in hyperthermia or combination group decreased compared with NS group (Fig. 3D). This appeared partly due to the fact that single rAAV-PEDF treatment could inhibit tumor growth and led to partial tumor regression. On the other hand, water bath could make mice uncomfortable and led to food intake reduction, and finally, the body weight of mice declined. Fortunately, mice restored and their weight began to increase slowly about 5–6 days after hyperthermia (Fig. 3D).
Inhibition of tumor angiogenesis
To analyze the antitumor mechanism, MVD in tumor tissues was measured by staining tumor sections with CD31 antibody. As shown in Fig. 4A, MVD of tumor tissues from rAAV-PEDF-treated mice (22.67±4.67, p<0.001; Fig. 4A, panel d) or hyperthermia-treated mice (37.67±4.63, p<0.001; Fig. 4 A, panel c) was clearly less than that of the NS group (56.67±5.72; Fig. 4A, panel a) or the rAAV-null group (58.0±8.53; Fig. 4A, panel b). Furthermore, MVD in the combination treatment group was the least of all (14.33±3.39, p<0.001; Fig. 4A, panel e). These results suggest that the inhibition of angiogenesis in tumors was responsible for tumor growth delay. In addition, combination therapy with rAAV-PEDF and hyperthermia had better therapeutic benefits for antiangiogenesis.
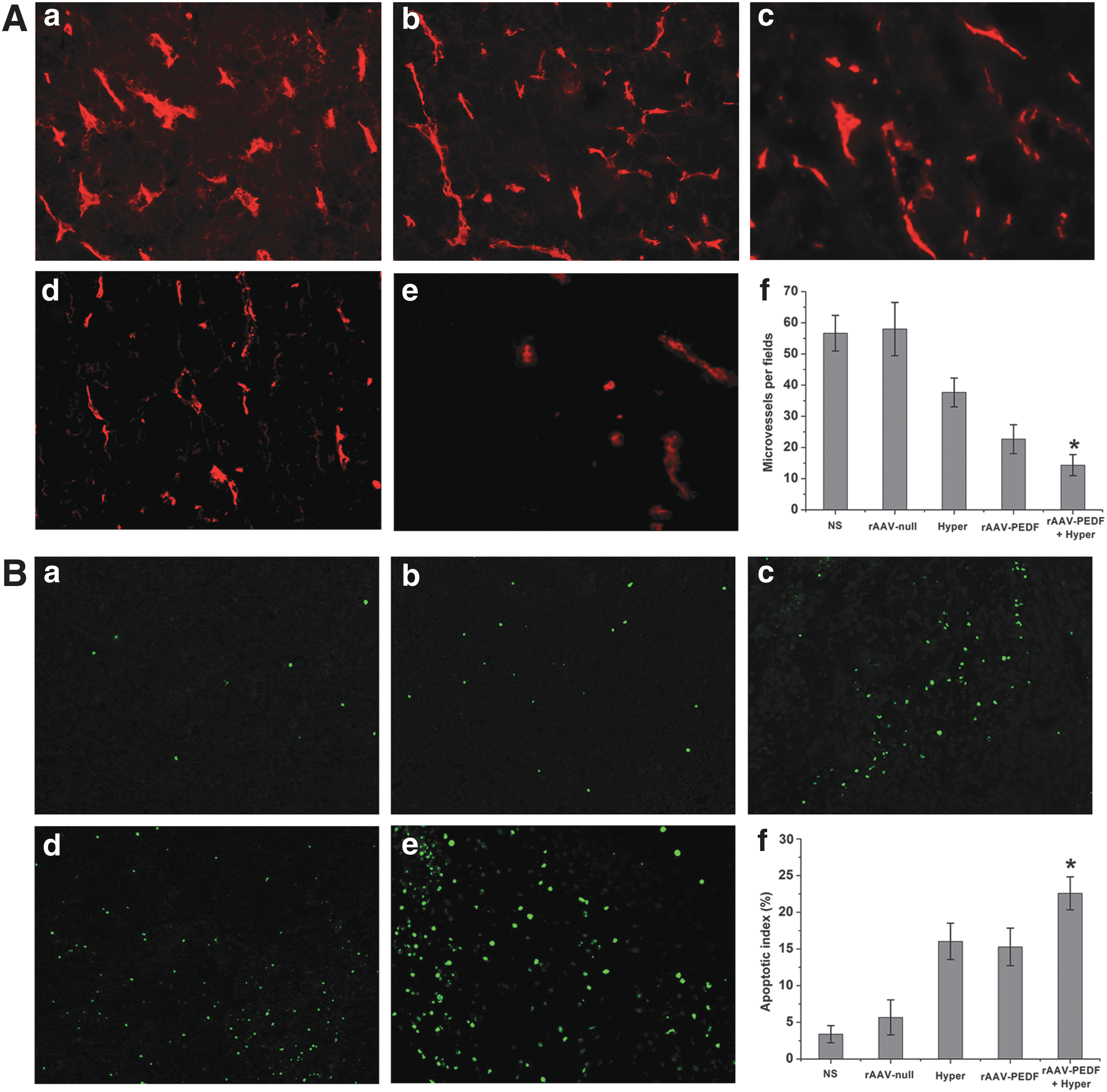
Immunofluorescence staining of tumor tissues (original magnification, ×200). On day 25 after cell inoculation, mice became moribund and were killed, and microvessel density (MVD) and apoptosis in tumor tissues were detected by immunofluorescence analysis. MVD was calculated by counting the number of microvessels in each field. The apoptotic index was determined as the ratio of apoptotic cell number to total cell number in each field, and data are shown as the mean apoptotic index±SD for each group.
Induction of apoptosis in tumor
The effect on tumor cell apoptosis was detected by immunofluorescence TUNEL staining assay. According to Fig. 4B, the apoptosis rate for the rAAV-PEDF+Hyper group (22.58±2.25%; Fig. 4B, panel e) was significantly higher than for the NS group (3.38±1.72%, p<0.001; Fig. 4B, panel a), rAAV-null group (5.67±2.39%, p<0.001; Fig. 4B, panel b), Hyper group (16.03±2.47%, p<0.001; Fig. 4B, panel c), or rAAV-PEDF group (15.28±2.56%, p<0.001; Fig. 4B, panel d). In addition, compared with the NS group, the apoptosis rates of the Hyper group and rAAV-PEDF group were higher (p<0.001), but there was no significant difference between them (p>0.05).
Histologic analysis of tumor
To detect histological changes in tumors, tumor tissues were obtained and processed for H&E staining. Figure 5 showed representative images of tumors from mice on day 25 after cell inoculation. Compared with control groups (Fig. 5A and B), apparently elevated cluster-like leukocyte infiltration was found in the Hyper group (Fig. 5C), rAAV-PEDF group (Fig. 5D), and rAAV-PEDF+Hyper group (Fig. 5E); of the latter three groups, the combination therapy group showed the most cluster-like leukocyte infiltration. In addition, compared with the NS group, significant necrosis was found in the hyperthermia-treated group (Fig. 5C), but the extent of tumor necrosis in the combination therapy group (Fig. 5E) was even greater. This phenomenon suggested that hyperthermia alone could lead to the necrosis of tumor, but combination therapy was more effective.
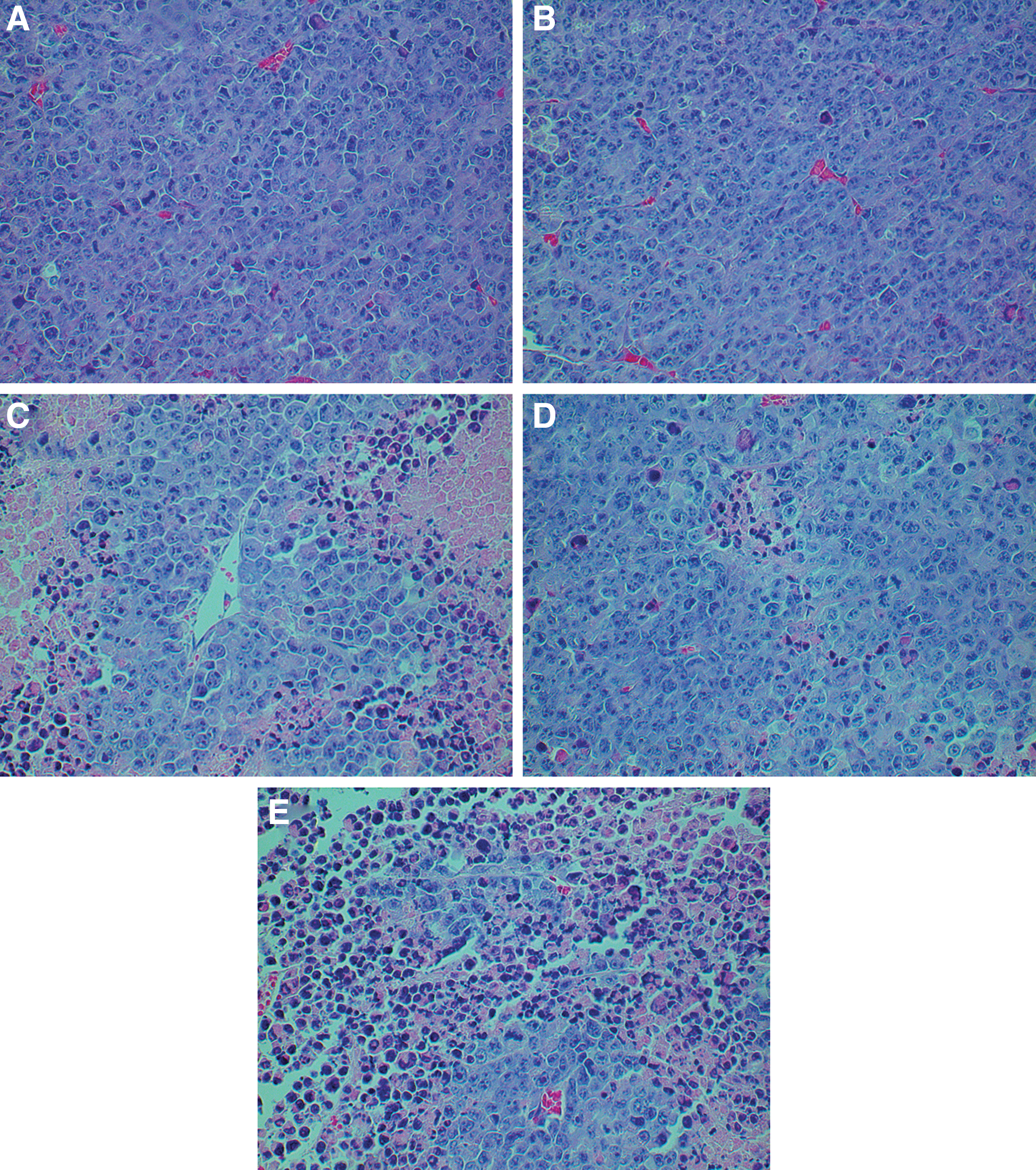
Hematoxylin–eosin (H&E) staining of tumor tissues (original magnification, ×400). To further observe histological changes, all tumor tissues collected on day 25 after cell inoculation were fixed in 4% paraformaldehyde for 48 hr and embedded in paraffin. The tissues were then sectioned and stained with H&E.
Increasing PEDF expression in vivo by rAAV-PEDF combination with hyperthermia
To explore whether the PEDF expression level was altered in mice, we detected the expression of PEDF in tumor tissue by immunohistochemistry and in serum by ELISA. Figure 6A shows representative images of tumor tissues, and only in the rAAV-PEDF group (Fig. 6A, panel d) and rAAV-PEDF+Hyper group (Fig. 6A, panel e) was PEDF protein expression detected. The level of PEDF expression was significantly higher in the combination treatment group (Fig. 6A, panel e) than in the rAAV-PEDF group (Fig. 6A, panel d). In addition, the PEDF concentration in serum from rAAV-PEDF-treated mice (13.12±3.14 ng/ml; p<0.001) (Fig. 6B) was about 4-fold more compared with the NS group (3.58±1.24 ng/ml) (Fig. 6B). A nearly 2-fold increase in serum PEDF was seen in the combination treatment group (20.65±3.51 ng/ml; p<0.001) (Fig. 6B) compared with the rAAV-PEDF single treatment group (13.12±3.14 ng/ml) (Fig. 6B). These results revealed that hyperthermia could increase the level of PEDF expression and further improve the effect of antiangiogenic gene therapy.
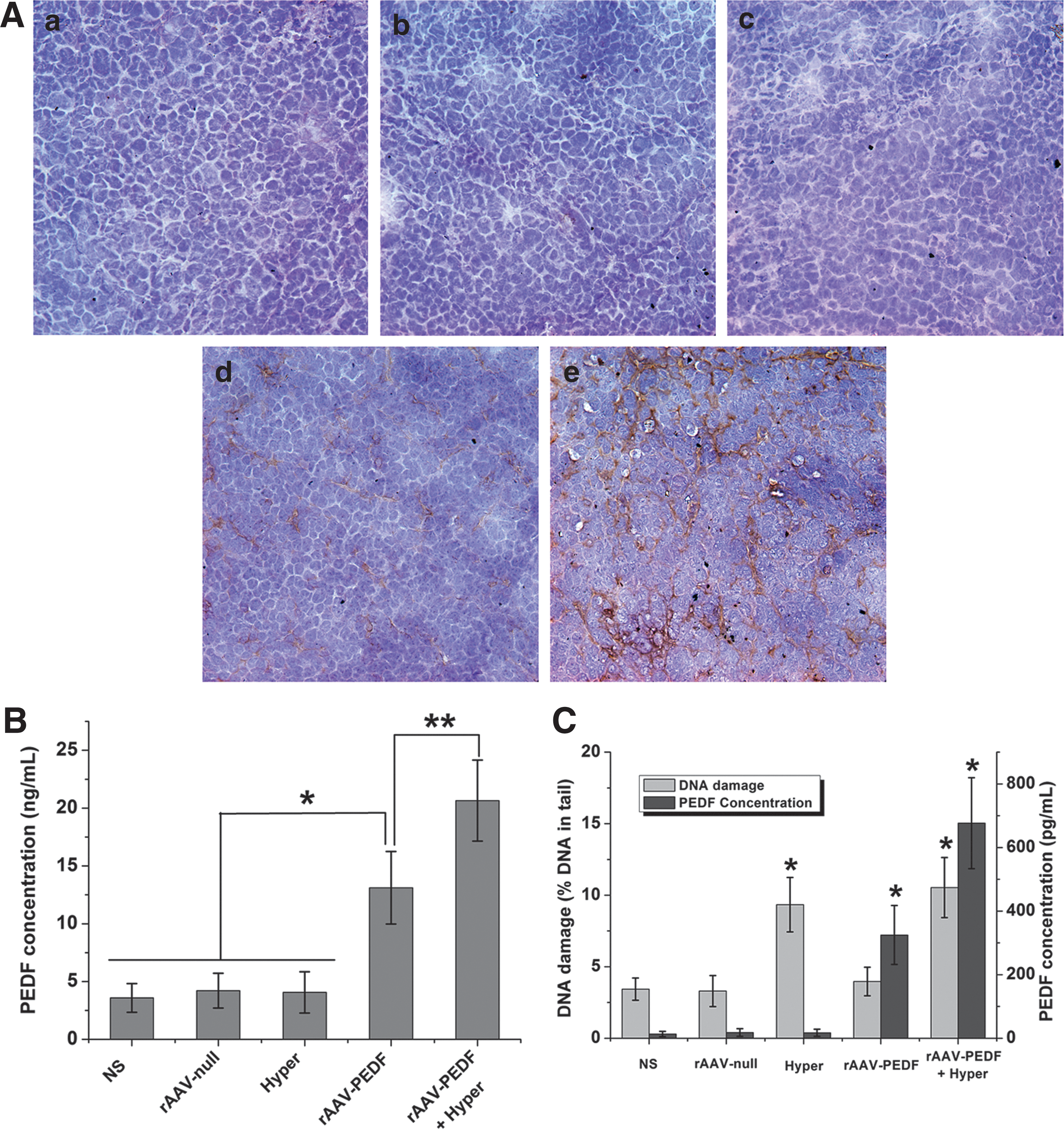
PEDF expression level in tumor tissue and serum, and relation between DNA damage and AAV-mediated PEDF expression in vitro. To explore whether the PEDF expression level was altered in mice, on day 25 after cell inoculation we detected the expression of PEDF in tumor tissue by immunohistochemistry and that in serum by ELISA.
Observation of potential toxicity
We further investigated the systemic toxicity or potential side effects of the treatment. All mice were observed each day, including their general condition, body weight, mortality, and so on. We found that the food intake and activity of mice were decreased slightly during the first few days (usually 3–5 days) after hyperthermia treatment; other than this, no other obvious signs of side effects were observed (data not shown). In addition, H&E-stained sections of major organs were observed by microscopy, and no significant histopathological lesions were found (Supplementary Fig. S1; Supplementary data are available online at
Relation between DNA damage and PEDF expression in vitro
To investigate the correlation between DNA damage and AAV-mediated PEDF expression in vitro, a comet assay and ELISA were conducted. As shown in Fig. 6C and Supplementary Fig. S4, compared with the NS, rAAV-null, and rAAV-PEDF groups, DNA damage in the Hyper group and rAAV-PEDF+Hyper group increased by nearly 2-fold after incubation at 42°C. At the same time, the AAV-mediated PEDF expression level in the rAAV-PEDF+Hyper group was 676.6±142.9 pg/ml, which was higher than that in the rAAV-PEDF group (324.9±92.8 pg/ml; p<0.001), Hyper group (17.3±11.3 pg/ml; p<0.001), rAAV-null group (18.4±12.0 pg/ml; p<0.001), or NS group (13.4±8.4 pg/ml; p<0.001). These results suggested that AAV-mediated PEDF expression was associated with DNA damage, and that DNA-damaging agents such as hyperthermia could increase the AAV-mediated gene expression level.
Discussion
Tumor growth and dissemination are angiogenesis dependent, and solid tumors cannot grow beyond 2–3 mm in diameter without vascularization (Hanahan and Folkman, 1996). On the basis of this point, antiangiogenic gene therapy is considered one of the most promising and feasible antitumor strategies. Such an approach acts by cutting off the blood supply to tumors, not by acting directly on tumor cells (Ma et al., 2002). PEDF is one of the most appealing angiogenesis inhibitors and has been extensively studied, because it specifically inhibits only new blood vessel growth while having no effect on normal blood vessels in the body. In addition, the nonpathogenic human parvovirus AAV has gained attention for gene therapy in many phase 1/2 clinical trials in human (Moss et al., 2007; Wright, 2011). Our previous study indicated that rAAV-PEDF could suppress tumor growth and metastasis in a colorectal peritoneal carcinomatosis (CRPC) mouse model by a single intraperitoneal administration (Wu et al., 2012). Although rAAV-PEDF showed an inhibitory effect on tumor growth and metastasis in the CRPC model, the therapeutic effects of AAV-mediated gene therapy need to be improved (Ferrari et al., 1996; Fisher et al., 1996; Jiang et al., 2011).
A combination of antiangiogenic factors with other antiangiogenic factors or cytotoxic therapies can be more effective than single antiangiogenic therapy and show better efficacy (Abdollahi et al., 2003; Li et al., 2005). Previous studies reported that on exposure to some DNA-damaging agents, cells might provide sites for AAV integration, which could lead to enhanced efficiency of AAV transduction (Alexander et al., 1994; Jiang et al., 2011). Hyperthermia showed direct tumor cell-killing ability, which has been recognized in association with DNA damage as well (Roca et al., 2003; Chen et al., 2008; Bettaieb et al., 2013). However, whether hyperthermia is able to enhance AAV transduction in solid tumor remains unclear. Therefore, to improve the anticancer effect of AAV-mediated gene therapy, we conceived a novel strategy that combined rAAV-PEDF gene therapy with hyperthermia; this combination not only acted to cut the blood supply to tumors, but also directly killed tumor cells. In addition, hyperthermia may improve the transduction efficiency of AAV and enhance the therapeutic effect of AAV-mediated gene therapy. To our knowledge, our study is the first to demonstrate the synergistic antitumor effects of AAV-mediated PEDF with hyperthermia on solid tumor. Our results (Fig. 6C) showed that DNA damage of Meth-A cells increased with hyperthermia, and AAV-mediated PEDF expression was enhanced with the increase in DNA damage. This phenomenon revealed that the synergistic effect of hyperthermia and rAAV-PEDF gene therapy may contribute to enhanced transduction efficiency of AAV by hyperthermia-induced DNA damage. After hyperthermia, DNA damage of tumor cells increased, which improved the transduction efficiency of AAV. As a result, PEDF concentration was increased at the tumor site, and angiogenesis was inhibited dramatically. Tumor cells with DNA damage induced by hyperthermia might be more susceptible to PEDF-mediated antiangiogenic therapy, because they cannot repair the DNA damage, which eventually leads to cell death. In addition, there may other possible mechanisms underlying the synergy between rAAV-PEDF gene therapy and hyperthermia, and we will investigate other possible mechanisms in our future work.
Several observations have been made in this study concerning the antitumor effects and mechanism of action of AAV-mediated PEDF combined with hyperthermia on solid tumor. The combination of AAV-mediated PEDF with hyperthermia showed an improved therapeutic effect on the subcutaneous Meth-A mouse model. The combined effects of antiangiogenesis, tumor cell killing, and improved transduction efficiency of rAAV-PEDF by hyperthermia are probably responsible for the enhanced antitumor activity. These suggestions are supported by the findings in this study. Compared with the NS group, although single therapy with rAAV-PEDF or hyperthermia could suppress solid tumor growth, the combination therapy of rAAV-PEDF and hyperthermia exhibited stronger antitumor activity and led to a longer life span for tumor-bearing mice (Fig. 3). In addition, compared with single therapy, there were more apparent apoptotic cells in combination-treated tumor tissues (Fig. 4B), and the combination treatment did dramatically decrease microvessel density and increase treatment response (Figs. 3 and 4A). To investigate whether hyperthermia is able to enhance AAV transduction in vivo, we detected the expression of PEDF in tumor tissue and serum by immunohistochemistry and ELISA, respectively. There was higher expression of PEDF in mice treated with a combination of rAAV-PEDF therapy and hyperthermia than in those receiving single rAAV-PEDF therapy (Fig. 6). In addition, the ELISA results indicated that on day 25 after tumor cell inoculation, the PEDF level in the rAAV-PEDF+Hyper group was significant higher than in the other groups. These results provided further evidence that hyperthermia could enhance the expression of AAV-mediated target gene in vivo, which may in part result from hyperthermia improving the therapeutic effect of rAAV-PEDF monotherapy. In addition, AAV is a safe gene delivery vector, and this notion was further supported by the fact that AAV vector can offer target gene expression without severe toxic effects (Supplementary Figs. S1–S3).
The results of our research indicated that (1) rAAV-PEDF could infect Meth-A cells and express PEDF protein with biological activity (Figs. 1 and 2); (2) the combination of rAAV-PEDF and hyperthermia exhibited significant synergistic effects in a mouse Meth-A fibrosarcoma model (Figs. 3 –5); (3) hyperthermia could improve the PEDF expression level from rAAV-PEDF in vivo (Fig. 6); and (4) both AAV-mediated gene therapy and combination therapy were safe, with no obvious toxicity observed (Supplementary Figs. S1–S3). Above all, the combination therapy of rAAV-PEDF gene therapy and hyperthermia may have potential applications in future clinical cancer therapy. In addition, other therapy that could cause DNA damage (e.g., chemotherapy by cisplatin or doxorubicin, etc.) could be combined with AAV-mediated gene therapy to achieve synergistic effects in cancer therapy, which may serve as a new therapeutic strategy in the clinic.
Conclusions
In summary, our studies demonstrated new potential for the AAV vector in the application of an antiangiogenic strategy to treat established solid tumors. In addition, hyperthermia could improve AAV-mediated transgene expression in vitro, and the combination administration of hyperthermia and rAAV-PEDF had a significantly synergistic effect on solid tumor, suggesting that hyperthermia could serve as a promising adjunctive therapy for AAV-mediated gene therapy. Additional research is warranted to further study the underlying molecular mechanisms of hyperthermia-induced enhancement of AAV-mediated gene expression.
Footnotes
Acknowledgments
The authors thank Dr. Xiaobing Wu (State Key Laboratory for Molecular Virology and Genetic Engineering, Beijing, China) for help with the production of rAAV particles. This work was financially supported by National Natural Science Foundation of China (NSFC81301961) and China Postdoctoral Science Foundation (2013M531969).
Author Disclosure Statement
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.