
Abstract
Malignant gliomas (MGs) are highly vascularized, aggressive brain cancers carrying a dismal prognosis. Because of their high vascularity, anti-angiogenic therapy is a potential treatment option. Indeed, the anti-vascular endothelial growth factor (VEGF) antibody bevacizumab has demonstrated promising results in clinical trials. Similarly, adenovirus-medicated Herpes simplex virus thymidine kinase and ganciclovir (AdHSV-tk/GCV) suicide gene therapy has established itself in clinical trials as a potential novel therapeutic strategy for MGs. In this study, we demonstrate the feasibility of combining adenovirus-mediated soluble VEGF receptor-1 anti-angiogenic gene therapy with AdHSV-tk/GCV suicide gene therapy to treat experimental MGs. Our results reveal that, apart from inhibiting angiogenesis, other anti-tumor mechanisms, such as reduction of infiltration by tumor-associated macrophages/microglia, may contribute to the improved therapeutic benefit of combination therapy.
Introduction
M
Soluble VEGF receptor-1 (sVEGFR-1) (Kendall and Thomas, 1993) inhibits angiogenesis by sequestering VEGF because of its high affinity, and by dominant-negative heterodimerisation with transmembrane VEGRs (Kendall and Thomas, 1993; Kendall et al., 1996; Thomas, 1996; Barleon et al., 1997; Cebe-Suarez et al., 2006). Adenovirus-mediated sVEGFR-1 (AdsVEGFR-1) gene therapy has been successful in inhibiting angiogenesis in several preclinical cancer models in both primary and metastatic settings (Goldman et al., 1998; Kong et al., 1998; Sallinen et al., 2009). In most of these studies, the efficacy was evaluated as a monotherapy. A few studies have combined sVEGFR-1 with chemotherapies such as 5-FU (Zhang et al., 2005), cisplatin (Gao et al., 2007), and paclitaxel (Sopo et al., 2012), and oncolytic virotherapy. Combination with other anti-angiogenic agents (Graepler et al., 2005) has shown synergy with sVEGFR-1. Our group has previously demonstrated successful combination of soluble VEGF receptors 1, 2, and 3 (Sallinen et al., 2009; Sopo et al., 2012), and 1 and 3 (Sallinen et al., 2011) with soluble Tie-2 in an ovarian cancer model.
Combination with cytotoxic therapy could possibly overcome some of the inherent drawbacks of anti-angiogenic therapy, mentioned before. Adenovirus-mediated Herpes simplex virus type-1 thymidine kinase and ganciclovir (AdHSV-tk/GCV) suicide gene therapy is one of the well-characterized and successful treatments for MG in preclinical (Tyynela et al., 2002) and clinical trials (Sandmair et al., 2000; Immonen et al., 2004; Westphal et al., 2013). Previous work from our group demonstrated synergism between AdHSV-tk/GCV and anti-angiogenic endostatin gene therapy in a renal cell cancer model (Pulkkanen et al., 2002). AdHSV-tk/GCV and AdsVEGFR-1 gene therapies have not been combined to treat experimental cancer models before. The objective of our study was to evaluate the preclinical feasibility, efficacy, and safety of combining these two therapies to treat MG.
Herein, we demonstrate for the first time the feasibility of successfully combining AdHSV-tk/GCV therapy with AdsVEGFR-1 gene therapy to treat MG in an animal model. The study demonstrates the ability of this gene therapy combination to enhance the therapeutic outcome in an orthotopic, syngeneic rat MG model in an immunocompetent host. AdsVEGFR-1 therapy, in spite of not being able to significantly inhibit angiogenesis in these aggressive tumors, was able to reduce the recruitment of tumor-promoting, CD68+, tumor-associated macrophages/microglia (TAM) into the tumors.
Materials and Methods
Cell lines and cell culture
Adherent BT4C rat MG cells were grown at 37°C in the presence of 5% CO2 in Dulbecco's modified Eagle's medium containing L-glutamine and glucose, supplemented with 10% fetal bovine serum (HyClone).
Adenovirus vectors
All vectors were replication-deficient, first-generation, E1–E3-deleted, serotype 5 adenoviruses driven by a human cytomegalovirus promoter. Adenoviruses carrying cDNA for HSV-tk (GenBank V00470) (AdHSV-tk; Cerepro), manufactured in HEK-293 cells, purified by density gradient centrifugation, were supplied by FinVector Vision Therapies. The titer of the vector was 1.5×1012 vp/ml. Production of the adenovirus vector encoding cDNA for the first five immunoglobulin-like domains of human VEGFR-1 fused in-frame with the sequence encoding for Fc-portion of human IgG is described elsewhere (Pajusola et al., 1993; Takayama et al., 2000; Bhardwaj et al., 2005). The functional titer was 2.7×1012 vp/ml. Adenovirus vectors encoding LacZ (AdLacZ) had a functional titer of 1.3×1012 vp/ml.
Assessment of BT4C cell viability after AdsVEGFR-1 gene transfer
Subconfluent BT4C cells were transduced with AdsVEGFR-1 vector at multiplicity of infections (MOIs) 0, 0.1, 1, 5, 10, and 25 to assess the impact of viral transduction and the sequestration of VEGF by sVEGFR-1 on cell viability. Cell viability was measured 24, 48, 72, 96, 120, and 144 hr after transduction, using a 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay (Promega CellTiter 96 AQueous One Solution Cell Proliferation Assay kit) according to the manufacturer's instructions. Absorbance was detected at 490 nm using a spectrophotometer.
Effect of AdsVEGFR-1 gene transfer on AdHSV-tk/GCV-mediated cytotoxicity
To evaluate the impact of transduction by AdsVEGFR-1 on AdHSV-tk/GCV-mediated cytotoxicity, subconfluent BT4C cells were transduced with AdHSV-tk and AdLacZ or AdsVEGFR-1 vectors, alone or in combination at an MOI of 0, 5, or 10 on day 1. GCV was added at a concentration of 1 μg/ml for 24 hr on day 3, and the cell viability was measured 24, 48, 72, and 96 hr after, as described before.
ELISA for VEGF, sVEGFR-1, and binding of rat VEGF to human sVEGFR-1
In vitro expression of sVEGFR-1 and the effect of that on the production of VEGF by BT4C cells (5×105) transduced with AdHSV-tk and/or AdsVEGFR-1 at MOIs 0 and 25 were measured 1 week after transduction by ELISAs specific for rat VEGF (R&D Systems) and human sVEGFR-1 (R&D Systems) according to manufacturers' instructions. The conditioned media (CM) were collected 24 hr after serum starvation and stored at −70°C until ELISAs. Cells were harvested at the same time and stored at −70°C for RT-PCR. Simultaneous transduction by both vectors was used to assess the effect of double-vector transduction on the transgene expression and VEGF production by the cells. A custom-made ELISA was used to assess the binding of rat VEGF into human sVEGFR-1. Briefly, the 96-well ELISA plates were coated with 100 μl of rat VEGF164 (R&D Systems) at 1000 and 500 ng/ml concentrations in 0.1 M NaHCO3 coating buffer. Human VEGF165 (R&D Systems) was used as the control. After 2 hr of blocking, the wells were incubated for 1 hr at room temperature with 100 μl of human sVEGFR-1-Fc protein at 5 and 500 ng/ml concentrations. Bound human sVEGFR-1-Fc was detected by incubating for 1 hr with rabbit polyclonal anti-human IgG-Fc peroxidase antibody (A8792; Sigma-Aldrich) in 1/20,000 dilution, followed-by standard ELISA protocol. Optical density was measured at 450 nm with a correction wavelength at 540 nm using a spectrophotometer.
Quantification of HSV-tk and sVEGFR-1 mRNA by RT-PCR
In vitro expression of HSV-tk and sVEGFR-1 mRNA by BT4C cells, 1 week after transduction with AdHSV-tk or AdsVEGFR-1 alone and in combination at MOIs 0 and 25, was carried out by RT-PCR on cell samples collected earlier. Briefly, total RNA was extracted from the cells by RNeasy Mini Kit (Qiagen) according to manufacturers' protocol, followed by DNAse treatment (Thermo Scientific). RNA was reverse transcribed to cDNA by M-MuLV reverse transcriptase (RevertAid; Thermo Scientific). Relative transgene expression was measured by RT-PCR (StepOnePlus Real-Time PCR system; Applied BioSystems) using SYBR Green Master Mix gene expression assays (Applied BioSystems) with TK primers or sVEGFR1 primers and beta actin (ACTB) as the endogenous control for normalization. Relative mRNA expression compared with the control BT4C cells was quantified using comparative CT (ΔΔ CT) method.
Tube formation assay
The functionality of AdsVEGFR-1 was tested by a tubular formation assay using a V2a Kit (TCS CellWorks) according to manufacturer's instructions. CM from BT4C cells transduced with AdsVEGFR-1, AdHSV-tk, the combination and the control cells collected previously were diluted 1:10 with the optimized growth medium provided by the manufacturer. Digital images were obtained using an inverted phase microscope (Olympus BX41) under 10× magnification on day 15. Tube formations were quantified in a blinded manner using CellF imaging software (Olympus Soft Imaging Solutions).
Pharmaceutical compounds
Ganciclovir (Cymevene 500 mg; Roche) was diluted in 10 ml of water for injection to achieve 50 mg/ml stock solution for injections.
In vivo experimental protocols
All animal experiments were approved by the Animal Welfare and Ethics Committee of University of Eastern Finland. Immunocompetent, inbred, male BDIX rats (Charles Rivers Laboratories) weighing 175–200 g were inoculated with a total of 10,000 BT4C cells in 5 μl of Opti-Mem (Invitrogen) into the anesthetized (fentanyl citrate and midazolam) rat brain at a depth of 2.5 mm, above the right corpus callosum, over 2–3 min, using a 27G, 25 μl Hamilton syringe (Hamilton). Needle was left in place for 5 min and then slowly withdrawn to avoid backflow. On posttumor implantation (p.i.) days 14 and 15, intratumoral gene transfers were conducted using multiple-site vector injection technique. Depending on the study group (Table 1), 15 μl of AdHSV-tk and/or 10 μl of AdsVEGFR-1 was injected using a Hamilton syringe placed in the stereotactic device (David Kopf Instruments). Vector injections of 7.5 μl of AdHSV-tk and/or 5 μl of AdsVEGFR-1, per each location, were carried out at vertical depths of 2.0 and 2.5 mm from the bregma level, 10 min apart. Needle was left in place for 10 min at each depth for proper dissipation of the vector solution. For the group receiving both vectors, AdHSV-tk gene transfer was done first. AdHSV-tk-transduced groups received intraperitoneal GCV treatment starting day 5 after gene transfer for 14 days at a dosage of 50 mg/kg/day per rat, given in two divided doses. Serum samples collected after completing the treatment were analyzed for bilirubin, alkaline phosphatase (AFOS), alanine aminotransferase (ALT), and creatinine at Kuopio University Hospital laboratory.
No surviving animals.
When calculating the mean and median survivals, the data censored at the primary end point of 50 days were considered as deaths.
Days, days posttumor implantation; n, number of rats; n/a, not applicable; SEM, standard error of the mean.
Magnetic resonance imaging
The presence of intracranial tumors was verified on p.i. day 13 by magnetic resonance imaging (MRI). Follow-up MRIs were done on p.i. days 28 and 42. For MRI, anesthesia was induced with 5% isoflurane in a mixture of 70%:30% N2O:O2, and was maintained at 1.5% isoflurane. MRIs were done using 4.7 T small-animal MR scanner (Magnex Scientific) interfaced to Varian Unity Inova console. Noncontrast-enhanced, T2-weighted spin-echo sequence with an echo time of 0.08 sec and repetition time of 2.5 sec was used to generate 17 coronal images of 1 mm thickness with no gap between slices to cover the tumor area in the rat brain. Images were of 256×256 matrices with a field of view of 4.0×4.0 cm. Total tumor volume was calculated by delineating the tumor area in all the image slices, processed, and analyzed with Matlab version 7.1b (MathWorks, Inc.) using premade macros.
Survival
Survival was calculated in days from tumor cell implantation to the sacrifice or death of an animal. Primary end point was to follow-up the rats in survival study for day 50 after tumor implantation. During the experiment, the animals were observed daily and body weight was measured regularly, and the animals were sacrificed with CO2 if the criteria for euthanasia defined by the Animal Welfare and Ethics Committee were met. Animals from histology groups were sacrificed approximately 10 days after completing treatment or at a corresponding point of tumor progression for histological analyses. Rats were transcardially perfused with 100 ml of 1% PBS. Samples from liver, spleen, kidney, and the whole brain were fixed in 4% paraformaldehyde at 4°C overnight and then changed into 15% sucrose solution having a pH of 7.4 at 4°C. Samples were processed and embedded in paraffin blocks, and were sectioned into 5-μm-thick slices.
Immunohistochemistry
Randomly selected paraffin-embedded tumor sections were stained with hematoxylin and eosin for general tumor morphology. Immunohistochemical staining was done using anti-rat primary antibodies for angiogenesis using anti-CD34 (AF4117; R&D Systems; 1/100), rat VEGF (ab1316; AbCam; 1/200), human sVEGFR-1 (ab9540; AbCam; 1/100), CD68 (MCA341R; AbDSerotec; 1/100), CD8 (MCA48R; AbDSerotec; 1/50), CD4 (MCA55R; AbDSerotec; 1/50), and CD25 (MCA273R; AbDSerotec; 1/50). Kidney, liver, and spleen samples were chosen as positive controls. Secondary antibodies were biotinylated horse anti-mouse IgG or goat anti-rabbit IgG (Vector Laboratories). Vectastain ABC-kit (Vector Laboratories) and DAB Plus (Zymed Laboratories) were used to form the color precipitate. Counter staining was done with hematoxylin (Merck KGaA). Staining for apoptosis was done using ApoTag apoptosis detection kit (Chemicon International) according to the manufacturer's instructions. The stained sections were examined under microscope and imaged using Olympus Bx41 inverted phase microscope (Olympus) under 20× magnification. Quantitative analysis of tumor angiogenesis, CD68, CD8, and apoptosis was done in a blinded manner using CellF image analysis software (Olympus Soft Imaging Solutions).
Statistical analyses
The data were analyzed with GraphPad Prism Version 5.01 statistical software (GraphPad Software, Inc.). Survival data were analyzed by Kaplan–Meier survival plots and the curves were compared using Mantel–Cox log-rank test with a Bonferroni-corrected p-value for significance set at 0.01. Column statistics were analyzed using one-way ANOVA or Kruskal–Wallis test and Bonferroni or Dunn's post-hoc test were used to compare groups.
Results
Viability of BT4C cells after AdHSV-tk and AdsVEGFR-1 transductions
Transduction with AdsVEGFR-1 did not affect the viability or the growth pattern of BT4C cells over a wide range of MOIs (0.1–25) up to 6 days after transduction (Fig. 1A). Maximum gene expression by adenovirus vectors is achieved 4–5 days after gene transfer. Hence, lack of growth inhibition even after 6 days indicates that the vector at these MOIs is not toxic to the cells and sequestration of VEGF by sVEGFR-1 does not affect the in vitro growth pattern of BT4C rat MG cells, in spite of VEGF being considered a growth factor in many cancers, including MG (Plate et al., 1992). Transduction of BT4C cells with AdLacZ or AdsVEGFR-1 at MOIs 5 or 10 had no significant impact on the cytotoxicity of AdHSV-tk/GCV over the measured period of cell viability (Fig. 1B and C). Combination of AdHSV-tk with AdLacZ was used as a control.
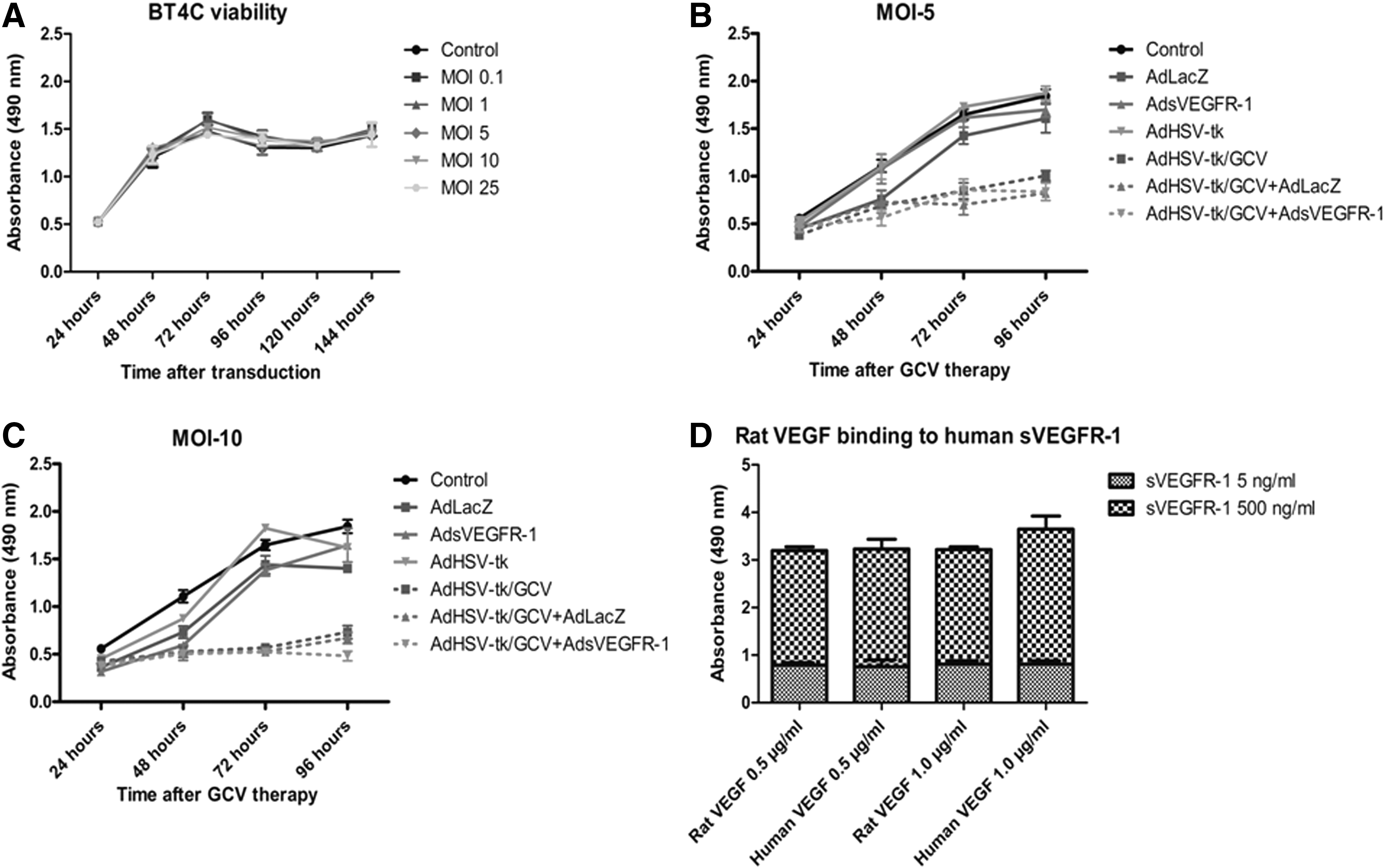
Viability of BT4C cells over time, measured by absorbance at 490 nm using an MTS assay.
Binding of rat VEGF into human sVEGFR-1
The modified ELISA binding assay reiterated the binding of ratVEGF164 to the human sVEGFR-1-Fc protein (Fig. 1D). There were no significant differences in the binding of the sVEGFR-1-Fc protein in high (500 ng/ml) or low (5 ng/ml) concentrations into rat and human VEGF at 1000 and 500 ng/ml VEGF concentrations (Fig. 1D), respectively, confirming almost equal binding potency.
Effect of combination gene transfers on VEGF, sVEGFR-1, and HSV-tk levels
As expected, BT4C rat MG cells produced VEGF in high concentrations (>30 ng/ml). Transduction by either vector alone or in combination did not have any major impact on VEGF secretion by these cells (Fig. 2A). Transduction with AdsVEGFR-1 and the vector combination produced sVEGFR-1 at almost equal concentrations of >700 pg/ml (Fig. 2B). The transgene construct used in the study was from the human sVEGFR-1, and the ELISA technique was specific for human sVEGFR-1. Accordingly, no human sVEGFR-1 was detected from the control and AdHSV-tk-transduced cells. RT-PCR results revealed 7.9- and 6.1-fold increases compared with the control BT4C cells in the relative HSV-tk mRNA levels in the AdHSV-tk- and vector combination-transduced cells, respectively, and >5-fold increase in the sVEGFR-1 mRNA level in both AdsVEGFR-1 and combination groups (Supplementary Fig. S1; Supplementary Data are available online at
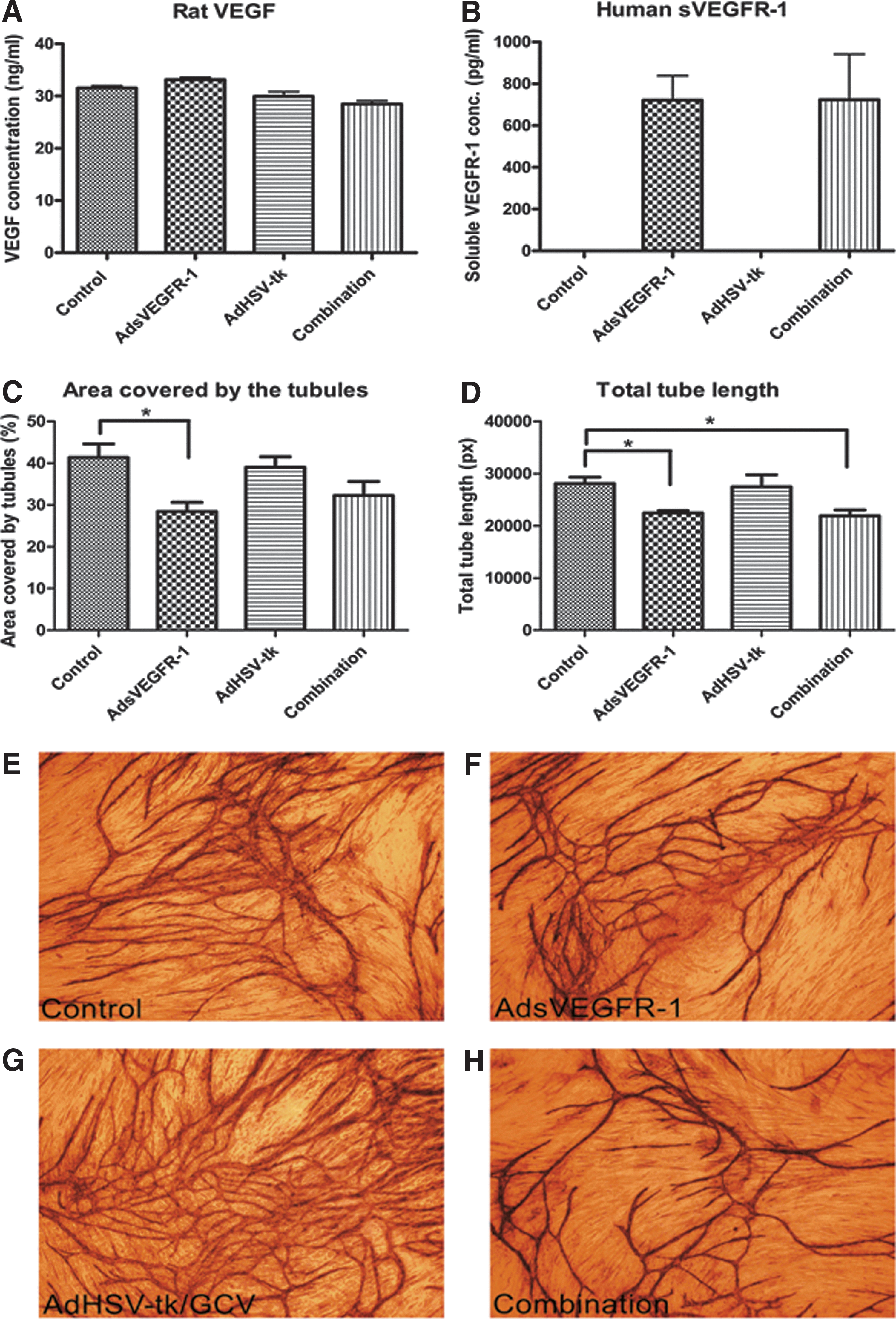
Secretion of
The functionality of AdsVEGFR-1
Analysis of tube formation revealed that the sVEGFR-1 produced and secreted into the CM by the transduced cells is functional. The percentage of area covered by the tubes and the total tube lengths were less in AdsVEGFR-1 and the combination groups, where the CM contained sVEGFR-1 as compared with the control and AdHSV-tk groups (Fig. 2C and D). Of the parameters, the area covered by the tubes in the AdsVEGFR-1 group and the total tube length in AdsVEGFR-1 and the combination groups were significantly (p<0.05) less compared with the respective control groups. Representative images from each group are presented (Fig. 2E–H). These results indicate that sVEGFR-1 is functional and capable of suppressing VEGF-mediated angiogenesis when transduced either alone or in combination with AdHSV-tk into BT4C cells.
Effect of combination gene therapy on rat MG tumor volume
There was no significant difference in the mean tumor volumes among the groups before starting the treatments at day 13 (Fig. 3A and Table 1). The MRI at 28 days p.i. revealed that AdsVEGFR-1 gene therapy had no effect on controlling tumor growth in this aggressive MG model. AdHSV-tk/GCV and the combination groups had significantly lower (p<0.001) mean tumor volumes compared with the control and AdsVEGFR-1-only groups, but not compared with each other, suggesting that AdHSV-tk/GCV is solely responsible for the effect. MRI 42 days p.i. could not be compared with the control or AdsVEGFR-1 groups because of the lack of surviving animals. Surprisingly, the mean tumor volume of the AdHSV-tk/GCV group was smaller compared with the group that received the treatment combination, even though the difference was not statistically significant. Representative MRI pictures are given in Fig. 3B.

Mean tumor volumes measured by MRI on 13, 28, and 42 days posttumor implantation
Effect of combination gene therapy on rat MG survival
AdsVEGFR-1 alone was not able to improve the survival of tumor-bearing rats (Fig. 3C). Compared with the control group (37 days), the median survival was reduced by 13.5% in this group (32 days). AdHSV-tk/GCV increased the median survival (41 days) by ∼11% and 28% compared with the control and AdsVEGFR-1 groups, respectively. In combination, AdHSV-tk/GCV and AdsVEGFR-1 were able to increase the median survival (47 days) by 27% and ∼47% compared with these two groups, respectively. Moreover, compared with the AdHSV-tk/GCV group, this was a ∼15% increase (Table 1). Survival in the combination group was significantly better (p<0.01) compared with both control [hazard ratio 0.24 (0.08–0.7)] and AdsVEGFR-1 groups [hazard ratio 0.20 (0.06–0.68)] (Fig. 3E and G). These results indicate 76% and 80% risk reductions, respectively. However, in spite of having a better trend toward survival, the difference did not reach significance compared with the AdHSV-tk/GCV group (Fig. 3H). No other comparison reached Bonferroni-corrected level of significance (p<0.01) (Fig. 3D and F). Furthermore, at the primary end point of 50 days p.i., the combination group had 50% of the animals still alive as opposed to 16% in the AdHSV-tk/GCV group, further emphasizing that the combination may be better than AdHSV-tk/GCV alone in enhancing long-term survival in this model. Neither the control group nor the AdsVEGFR-1 group had any surviving animals at this time point (Table 1).
Immunohistochemistry
As expected, human sVEGFR-1 expression was limited to AdsVEGFR-1 and the combination gene therapy groups (sVEGFR-1: Fig. 4B and D). VEGF levels in the BT4C tumors seem to be diminished in those two groups (VEGF: Fig. 4B and D), providing in vivo confirmation for the functionality of AdsEVGFR-1. However, the CD34 staining for angiogenesis did not show significant difference between the study groups (Fig. 4, CD34). The quantification of angiogenesis confirmed that there were no differences in the mean capillary density (Fig. 5A) between groups.
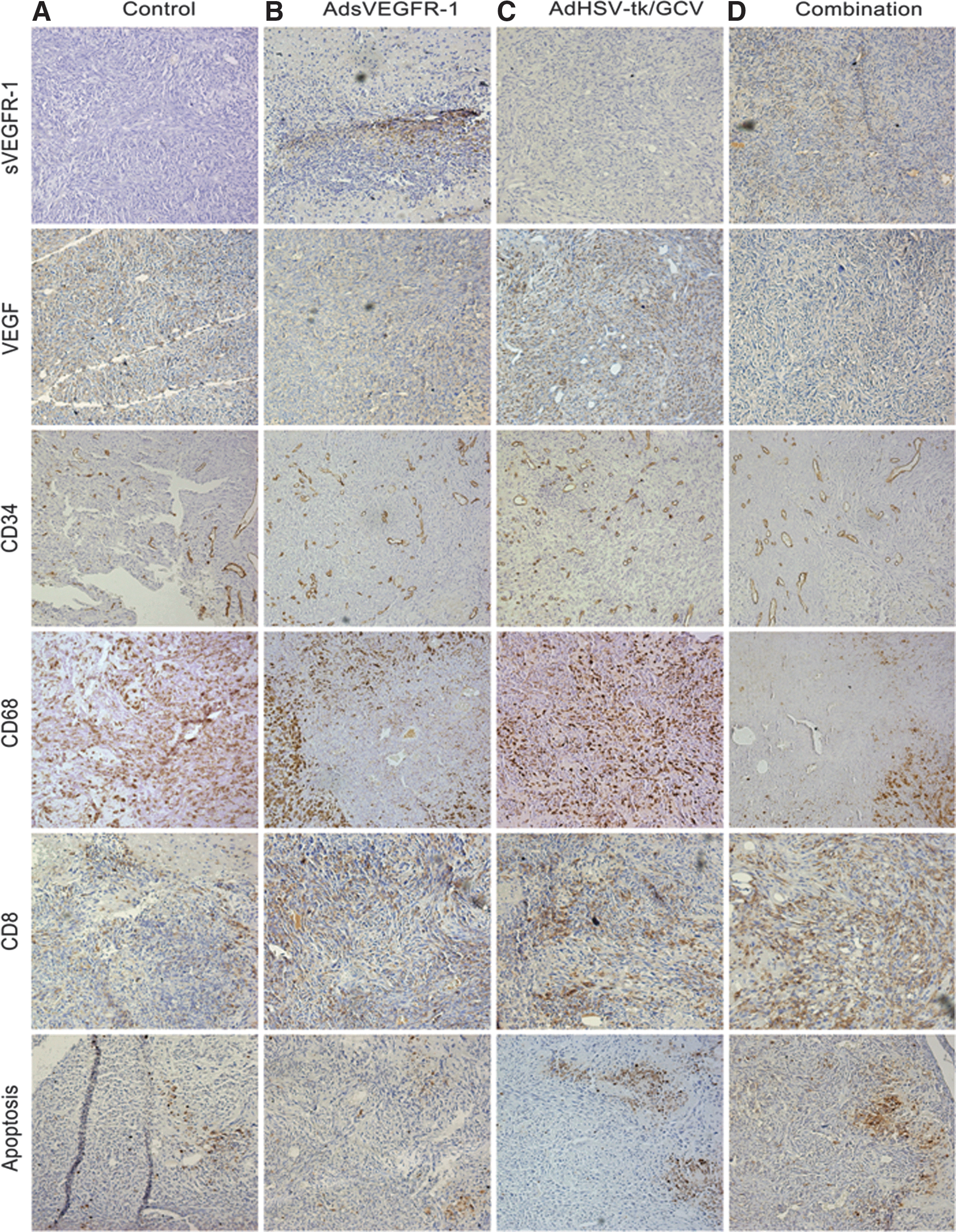
Immunohistochemical staining for human sVEGFR-1, rat VEGF, angiogenesis by CD34, tumor infiltrating macrophages/microglia by CD68, tumor infiltrating cytotoxic T-lymphocytes by CD8, and apoptosis by ApoTag staining from the control
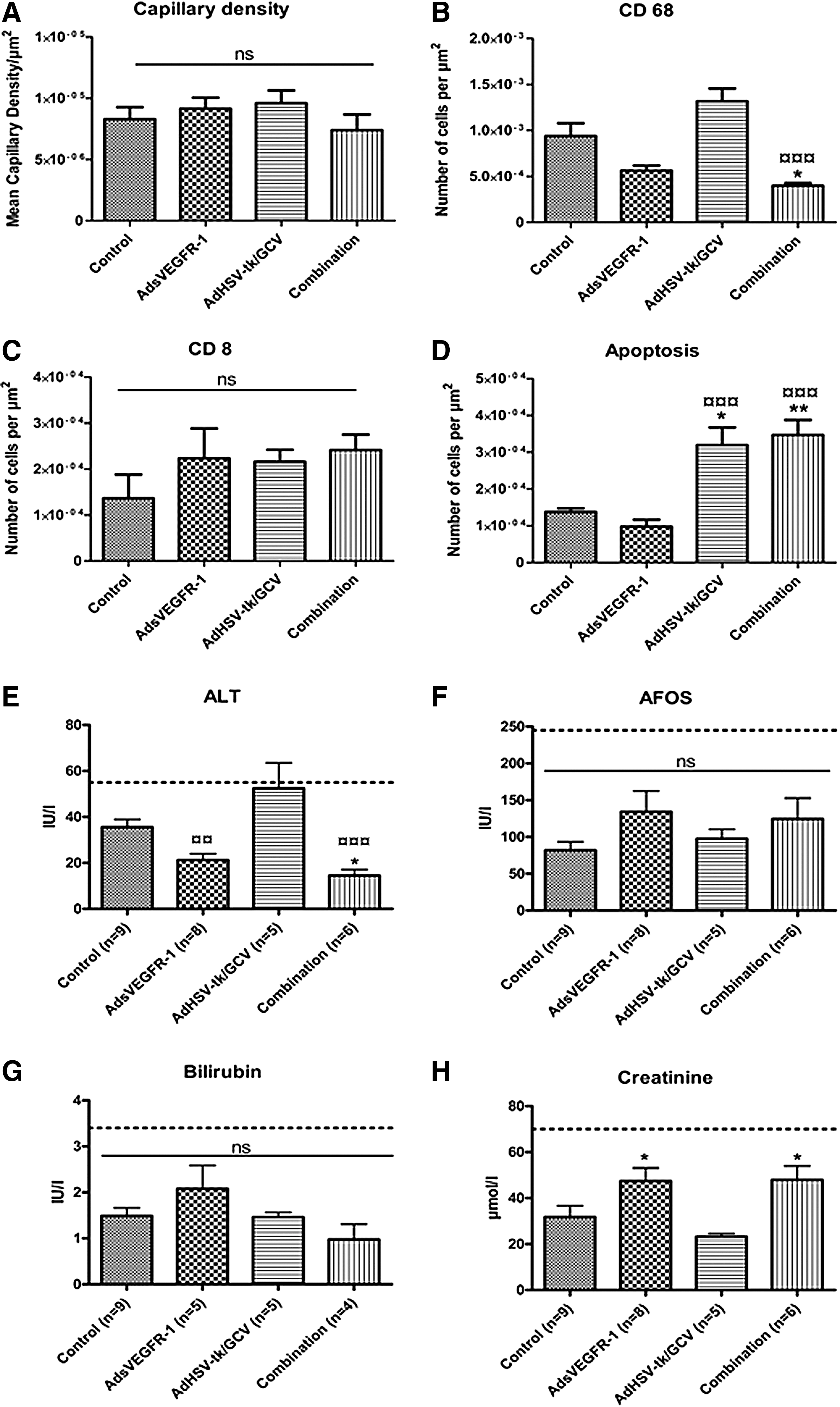
Since viral vectors were used to deliver the transgenes into the tumors, infiltration of inflammatory cells was studied using immunostainings for CD68-, CD8-, CD4-, and CD25-expressing cells. No expression of CD4 or CD25 was observed (data not shown). On the other hand, tumors were grossly infiltrated by CD68+ cells (CD68: Fig. 4A–D). On close inspection, it was evident that a specific area around the gene transfer needle-track in AdsVEGFR-1 and combination groups was devoid of CD68+ cells (CD68: Fig. 4B and D) as opposed to diffuse, uniform TAM infiltration in the control and AdHSV-tk/GCV groups (CD68: Fig. 4A and D). CD68+ cell density was less in the AdsVEGFR-1 and the combination groups compared with the control and AdHSV-tk/GCV groups, and this difference was significant in the combination group compared with the control (p<0.05) and AdHSV-tk/GCV (p<0.001) groups (Fig. 5B). No other comparisons reached statistical significance. All groups had some CD8+ T-lymphocyte infiltration (Fig. 4, CD8) and the differences in the CD8+ cell densities among the groups were not significant (Fig. 5C). Apoptotic cells were observed in all groups (Fig. 4, apoptosis), but were significantly higher in AdHSV-tk/GCV and the combination groups (Fig. 5D) as expected.
The safety of gene therapy combination
All measured serum parameters in all groups were within normal limits for rats. Liver function tests, AFOS and bilirubin, did not show significant differences between the groups (Fig. 5F and G). However, ALT level in the AdHSV-tk/GCV group was reaching the upper limit of normality and was significantly higher compared with the AdsVEGFR-1 and the combination groups. The difference in the ALT levels between the control and the combination groups was also statistically significant (Fig. 5E). Transient elevation of ALT levels has been reported with AdHSV-tk/GCV suicide gene therapy before (Immonen et al., 2004). The renal function test, serum creatinine, was within the normal range in all groups, but the differences between AdsVEGFR-1 and the combination groups compared with the AdHSV-tk/GCV group were statistically significant (Fig. 5H).
Discussion
MGs are highly vascularized tumors with increased VEGF expression, which makes them, theoretically, highly amenable to anti-angiogenic therapy. With the approval of bevacizumab, anti-angiogenic agents have entered the MG therapeutic arena. Soluble VEGFR-1 has been studied extensively as an anti-angiogenic gene therapy against a variety of cancers, including MG (Goldman et al., 1998; Ohlfest et al., 2005; Harding et al., 2006), and it has some theoretical advantages over bevacizumab. Being a parenteral therapy, bevacizumab has the disadvantage of evoking systemic adverse effects, while tumor-directed AdsVEGFR-1 therapy is less likely to carry this risk. The inherent cytostatic nature of anti-angiogenic therapies necessitates the combination with cytotoxic therapies, such as chemotherapy, for the therapeutic success. Several ongoing clinical trials are evaluating the efficacy of bevacizumab in combination with chemotherapy, in the adjuvant setting, for the treatment of newly diagnosed MGs (Gilbert et al., 2013; Herrlinger et al., 2013). Moreover, combination of different therapies may help to overcome the development of treatment resistance, which is a major problem in cancer care. Thus far, AdHSV-tk/GCV and AdsVEGFR-1 gene therapies have not been combined for the treatment of MG or any other cancer. AdHSV-tk/GCV has been successfully combined with endostatin gene therapy for the treatment of renal cell cancer (Pulkkanen et al., 2002), while sVEGFR-1 has been combined with oncolytic viruses (Zhang et al., 2005; Guse et al., 2010), other sVEGFRs (Sallinen et al., 2009), soluble Tie2 (Sallinen et al., 2011), and chemotherapy (Sopo et al., 2012). All previous studies with sVEGFR-1 gene therapy for MG were conducted on nude mouse models. Therefore, this is the first study to test the efficacy of sVEGFR-1 in an immunecompetent orthotopic rat MG model using adenovirus vectors to deliver sVEGFR-1 gene therapy.
Similar to human MGs, BT4C rat MGs did express high amounts of VEGF (>30 ng/ml). However, it is not clear to what extent VEGF acts as an autocrine/paracrine growth factor for MG cells. The lack of growth inhibition even up to 6 days after transduction by AdsVEGFR-1 indicates that the vector at these MOIs was not toxic to the cells and the degree to which VEGF was inhibited by sVEGFR-1 did not affect the growth pattern of BT4C rat MG cells. We did not differentiate which of the VEGF isoforms are being secreted by BT4C cells. However, VEGF165 is typically produced by cancers, and in human MGs, isoforms 121 and 165 have been reported (Berkman et al., 1993). VEGFR-1 can bind all isoforms of VEGF, and our modified ELISA confirmed the binding of rat VEGF164 to human sVEGFR-1. In MGs, VEGF expression far exceeds that of sVEGFR-1 (Lamszus et al., 2003). Even though MGs produce high amounts of endogenous VEGF and sVEGFR-1, the VEGF:sVEGFR-1 ratio is increased, tipping the balance toward an angiogenic phenotype (Plate et al., 1994; Lamszus et al., 2003). Increasing sVEGFR-1 production within the tumors by gene therapy may help to shift this balance in favour of anti-angiogenesis, even though VEGF levels are not reduced remarkably. It must also be emphasized at this point that apart from sequestering VEGF, sVEGFR-1 can also act through dominant negative heterodimerisation with VEGFR-2 (Kendall et al., 1996; Thomas, 1996; Barleon et al., 1997; Cebe-Suarez et al., 2006). In vitro transduction of BT4C cells with AdsVEGFR-1 or AdHSV-tk alone and in combination led to the production of human sVEGFR-1 protein, and sVEGFR-1 and HSV-tk mRNA at almost equal concentrations of >700 pg/ml, and ∼5- and ∼6–8-fold increase, respectively. Simultaneous transduction by two different vectors did not have a significant impact on the production or the functionality of these transgene constructs. This in vitro data supports the simultaneous gene transfer methodology applied in the in vivo experiment to be effective in expressing the transgene constructs without interfering with the production of each other.
The finding that AdsVEGFR-1 was not capable of controlling the tumor growth rate in vivo is contrary to what is expected from an anti-angiogenic therapy. Nevertheless, we were surprised to see that animals in the combination group lived significantly longer compared with the AdHSV-tk/GCV group, which had a smaller mean tumor volume on p.i. day 42. Interestingly, similar results have been reported previously with the use of anti-angiogenic therapies (Jahnke et al., 2009), highlighting the possibility of indirect mechanisms such as reduction in peritumoral edema contributing to the therapeutic efficacy. In intracranial tumor models, animals often have to be sacrificed because of poor physical well-being, which is at least partly because of elevated intracranial pressure, obstruction of the ventricular system, peritumoral edema, and pressure effects. Clinical studies have also demonstrated a reduction in the peritumoral edema in patients after anti-angiogenic therapy that may have contributed to their general well-being, in spite of not having a significant impact on the progression-free survival or overall survival (Batchelor et al., 2007). One reason for the lack of efficacy of anti-angiogenic therapy in this study could be the late onset of therapy (gene transfer on days 14 and 15 p.i. and possible gene expression by day 19) as opposed to early onset of anti-angiogenic therapy in most other studies where therapeutic efficacy was established (Gerber and Ferrara, 2005). The reason for this delay was that gene transfers were done only after MRI verification of tumor existence, which could only be performed after 12 or 13 days p.i.
Even though angiogenesis was not inhibited, sVEGFR-1 gene therapy enhanced the efficacy of suicide gene therapy, especially in terms of long-term survival. Examination of histology samples revealed the expression of human sVEGFR-1 by the tumors, thereby excluding the possibility of lack of gene expression as a reason for failure to inhibit angiogenesis. As a single therapy, AdsVEGFR-1 did not confer any therapeutic benefit, keeping in line with the concept that anti-angiogenic therapy should be combined with a cytotoxic therapy for optimal benefit (Ibrahim et al., 2012). Possible mechanisms for the better therapeutic outcome in combination group could be the increased GCV delivery into the tumor because of vascular normalization by anti-angiogenic therapy and the reduced peritumoral edema and intracranial pressure, improving the general well-being of the animals. Alternatively, the depletion of CD68+ cells within MGs is another anti-tumor mechanism. CD68+ cells in MG can be as high as 30% (Watters et al., 2005), and similar results have been reported in BT4C rat MGs (Wirth et al., 2012). CD68 has been described by some to be a marker of both M1- and M2-phenotype macrophages (Ohri et al., 2009), while others propose that as a marker of M2 phenotype (Murdoch et al., 2008). Recent evaluations of human MG samples suggest that CD68 is expressed by both microglia and tumor cells, with a higher CD68 positivity associated with an increased malignancy grade and poor survival (Strojnik et al., 2009). Multiple protumor mechanisms have been attributed to TAMs (Watters et al., 2005), and their tumor-promoting role is well established in many cancers, including MG (Gabrusiewicz et al., 2011; Coniglio et al., 2012; Kennedy et al., 2013; Sielska et al., 2013). Moreover, several studies have demonstrated therapeutic benefit by inhibiting TAMs in MG (Gabrusiewicz et al., 2011; Cieslewicz et al., 2013; Pyonteck et al., 2013). In our study, we observed a depletion of CD68+ cells in the area around the gene transfer needle track in the AdsVEGFR-1 and combination treatment groups, as opposed to diffuse uniform TAM infiltration in the control and AdHSV-tk/GCV groups. Interestingly, similar results have been reported with anti-VEGF therapies in other cancers (Roland et al., 2009a,b). VEGF is a known chemoattractant for TAMs because of their expression of VEGFR-1 and/or VEGFR-2 (Muramatsu et al., 2010). Hence, it is apparent that the sequestration of VEGF by sVEGFR-1 has inhibited the CD68+ protumorigenic TAM infiltration into tumor, contributing to the anti-tumor response. No significant increase in the amount of CD8+ cells was observed in the tumors of virus-treated animals compared with untreated control animals, indicating that CD8+ cells did not contribute to the improved survival.
In the present study, we were able to successfully demonstrate the feasibility of combining two gene therapy strategies for the treatment of MG and to examine the putative anti-tumor mechanisms. The underlying mechanism of therapeutic efficacy needs further evaluation, as well as optimization of the treatment protocol. The overall results of the in vivo experiment indicate that the combination of AdHSV-tk/GCV and AdsVEGFR-1 is feasible in an orthotopic, syngeneic rat MG model with a better therapeutic outcome compared with the individual therapies. Most importantly, the present findings suggest that AdsVEGFR-1 in combination with cytotoxic therapies may enhance the long-term survival. Furthermore, our results demonstrate that AdsVEGFR-1 gene therapy is safe when delivered locally into the tumors, even though previous studies have reported adverse effects, such as liver and renal toxicities, with systemic administration (Mahasreshti et al., 2001, 2003). A major limitation for combination gene therapy, especially to the brain, is the limited vector volume that can be injected into the tumors. The development of more potent vectors and vectors expressing multiple transgene constructs could further improve therapeutic efficacy of gene medicines.
In conclusion, our study confirms, for the first time, the feasibility, safety, and efficacy of simultaneous combination of AdHSV-tk/GCV with AdsVEGFR-1 for the treatment of cancers such as MG. Apart from reducing angiogenesis, inhibition of infiltration by TAMs could be another important anti-tumor mechanism of AdsVEGFR-1 therapy that contributes to the therapeutic benefit.
Footnotes
Acknowledgments
The authors thank Joonas Malinen, Anneli Miettinen, Sari Järveläinen, Tiina Koponen, Svetlana Laitinen, and Jaana Siponen for their skilful technical assistance. Our gratitude is extended to Drs. Nihay Laham-Karam, Tiina Nieminen, Pyry Toivanen, Minna Kaikkonen, Johanna Närväinen, Timo Liimatainen, and Teemu Laitinen of A. I. Virtanen Institute for their generous contributions. This study was supported by the University of Eastern Finland, Kuopio University Hospital, and European Research Council. The authors thank FinVector Vision Therapies for providing the AdHSV-tk vector for the study.
Author Disclosure Statement
No competing financial interests exist for any of the authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.