
Abstract

Introduction
T
History
In 2014, the concept of using a modified virus to transfer genes in vivo is standard practice, widely used throughout the biomedical research community. In the late 1980s, however, when laboratories such as ours started thinking about delivering genes in vivo, the concept was new, and the challenge daunting.
We began strategizing how to effectively transfer genes in vivo as a follow-up to our development of a therapy for α1 antitrypsin (AAT) deficiency, an autosomal recessive disorder associated with a serum deficiency of AAT and development of emphysema (Crystal, 1990). After almost a decade of work, we had developed augmentation, using weekly intravenous infusions of human plasma-purified AAT to treat the deficiency state (Gadek et al., 1980; Wewers et al., 1987). After that therapy was approved by the FDA, we began thinking about strategies in which we could use gene therapy to produce endogenous AAT, circumventing the need for weekly infusions with the purified protein. At that time, the most effective gene transfer method into primary cells was with a Moloney murine leukemia-based retrovirus. We used a retrovirus construct from E. Gilboa of Memorial Sloan–Kettering Cancer Center to transfer the normal human AAT cDNA to mouse fibroblasts (Garver et al., 1987). Although we were able to generate glycosylated, physiologically “normal” human AAT by the fibroblasts, the amounts produced were far below that needed to treat AAT deficiency, thus obviating an ex vivo cell-based strategy to effectively treat the deficiency state. This observation led us to the concept that it would be a lot more efficient if we could use a virus to transfer the human AAT gene directly to the lung (the site of disease) or liver (the normal site of AAT production) in vivo.
Early In Vivo Studies
In 1989, I received a call from Paul Tolstoshev, a former postdoctoral fellow who was the scientific director of the Strasbourg-based biotech company, Transgene. He told me of a collaboration they had initiated with Michel Perricaudet, an adenovirus virologist at Institute Gustave-Roussy in France, to develop a replication-deficient adenovirus as a gene delivery strategy. As a pulmonary physician, I was well aware that the adenovirus was capable of infecting the lung epithelium and recognized that this might be an effective means to deliver genes directly to the lung. Michel graciously offered to train us in the use of adenoviruses, and I sent David Curiel (then a postdoctoral fellow in our laboratory) to Michel's laboratory to learn the technology so that we could transfer it to our laboratory. David did so, came back with the adenovirus components and the 293 cell line to produce the recombinant vectors, and taught Melissa Rosenfeld (another postdoctoral fellow; now M. Ashlock) how the adenovirus system worked. By deleting the E1 genes to prevent replication, and the E3 genes to make more room for the transgene, the common human serotype 5 adenovirus could be converted to a vector that had sufficient room for a promoter and transgene and was replication deficient (Fig. 1). We quickly established the system in our laboratory, and in one of those rare eureka moments in any scientist's career when you recognize that an observation in your laboratory may have significant implications, we observed that an E1−E3− adenovirus coding for β-galactosidase was strikingly effective in transferring genes in vivo (Fig. 2). This quickly led to a publication in Science, representing the first demonstration of efficient in vivo transfer of a gene to experimental animals with high levels of organ-specific expression. Soon afterward, we demonstrated that an adenovirus vector could be used to effectively transfer and express the normal human AAT cDNA to the liver, the natural site of AAT expression (Jaffe et al., 1992).
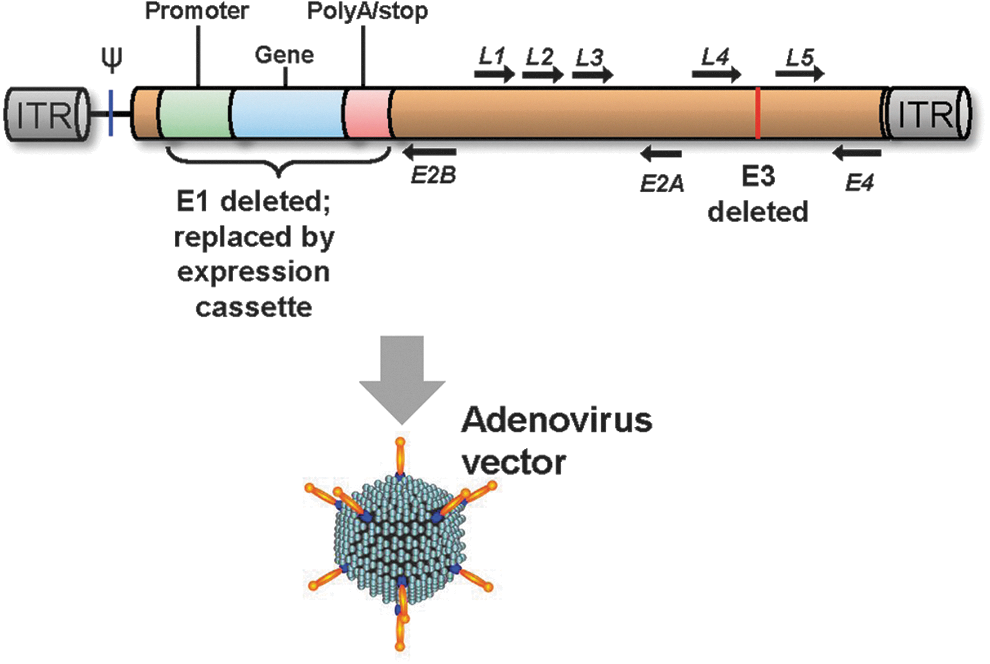
Schematic of a typical adenovirus gene transfer vector genome. In an adenovirus vector, the early (E) genes in the E1 region are deleted (to prevent replication) as is the E3 region (to make more room for the expression cassette). The inverted terminal repeats (ITR), packaging signal (ψ), and the late (L) genes remain in the vector. The deletions allow for an expression cassette of up to 7–8 kb. A typical expression cassette, including a promoter, the transgene, and stop/polyA sequences, is inserted into the deleted El region. The construct is typically packaged in 293 cells, a cell line that expresses the human adenovirus E1 region, thus providing the components necessary for replication. The vector enters cells through the fiber interacting with the coxsackie-adenovirus (CAR) receptor and secondary integrin (e.g., αVβ3–5) receptors.

The first example of effective in vivo gene transfer using an adenovirus vector. Top: Examples from a notebook in 1991 from the Crystal laboratory, Pulmonary Branch, the National Heart, Lung, and Blood Institute, of a lung of a cotton rat that had received intratracheal E1−E3− adenovirus coding for β-galactosidase under control of an RSV promoter 7 days earlier. Shown is a control and with AdRSVβgal vector. There is extensive β-galactosidase expression throughout the lung. Bottom: The publication in Science was the first article describing effective in vivo gene transfer with a recombinant replication-deficient adenovirus (Rosenfeld et al., 1991).
Around the same time that we were developing the in vivo gene transfer methods to the lung using an E1−E3− serotype 5 adenovirus, the cystic fibrosis transmembrane conductance regulator (CFTR) gene was identified and characterized (Kerem et al., 1989; Riordan et al., 1989; Rommens et al., 1989). Recognizing that adenoviruses were trophic for the human airway epithelium, we realized that our in vivo gene transfer strategy would be ideal to transfer the human CFTR cDNA to the airway epithelium to treat cystic fibrosis. We quickly constructed an E1−E3− serotype 5 adenovirus gene transfer vector with an expression cassette that included the normal human CFTR cDNA under control of a constitutive promoter. This led to a publication in Cell documenting effective gene transfer and expression of CFTR to the airway epithelium in vivo (Fig. 3) (Rosenfeld et al., 1992).
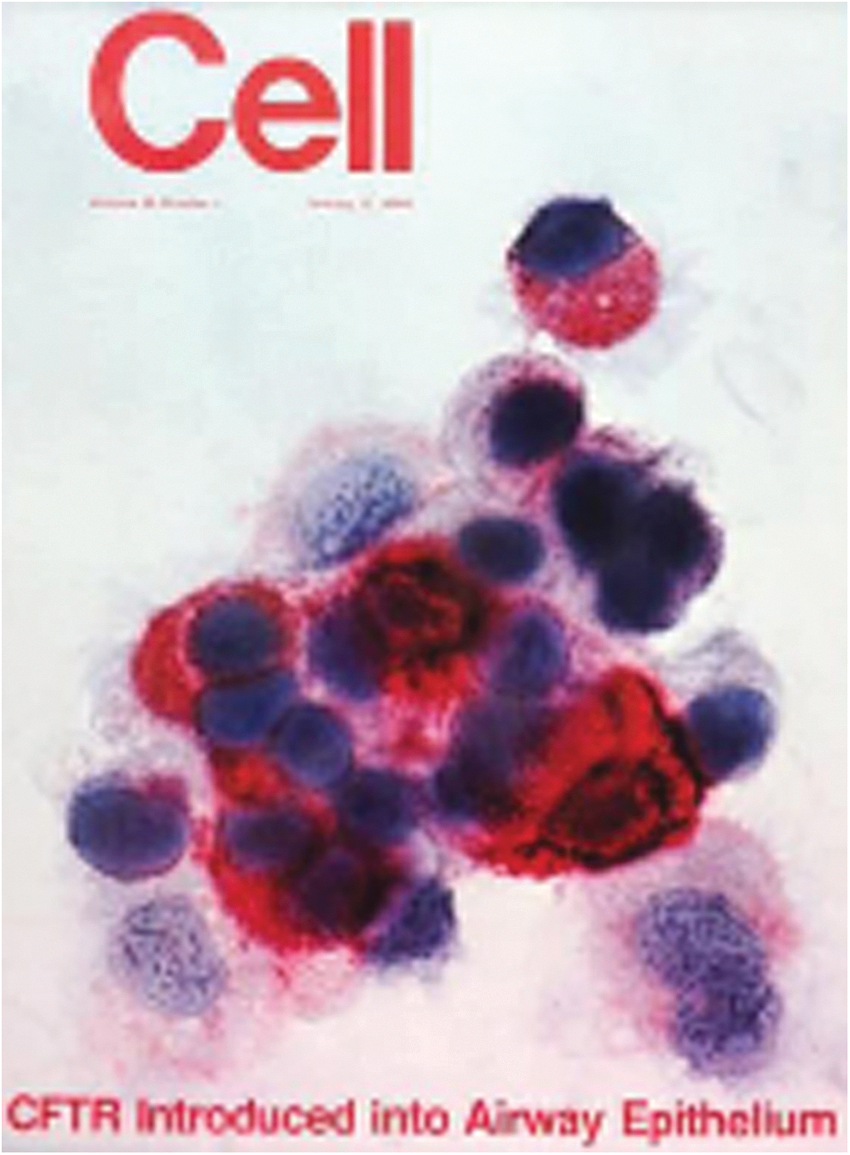
The publication in Cell demonstrating effective in vivo transfer of the human cystic fibrosis transmembrane conductance regulator (CFTR) cDNA to the epithelium of cotton rats. The expression of the human CFTR protein in the airway epithelium was detected by an antibody 2 weeks after intratracheal administration of an E1−E3− serotype adenovirus vector expressing the human CFTR cDNA (Rosenfeld et al., 1992).
The First Human Studies
With the successful demonstration of effective adenovirus-directed expression of the normal human CFTR cDNA in the airway epithelium of experimental animals, our laboratory in the intramural program at the National Heart, Lung, and Blood Institute, and laboratories led by Jim Wilson (initially at Michigan and then at University of Pennsylvania) and Mike Welsh at Iowa and his colleagues at Genzyme began to seriously consider using adenovirus vectors in vivo to treat the pulmonary manifestations of cystic fibrosis in humans. In a historic (for the gene therapy field) meeting at the NIH DNA Recombinant Advisory Committee meeting on December 4, 1992, with over 200 scientists, media, venture capitalists, and representatives from pharma in the audience, all three groups had protocols approved. One of the concerns discussed was the theoretical risk for in vivo recombination of the E1−E3− adenovirus with remnants of endogenous viral sequences, generating a recombinant lethal virus that would infect the community. As a precaution, we constructed two negative pressure rooms in the NIH Clinical Center to contain any recombinant viruses that might be generated. We were lucky enough to win the race to be the first to use a recombinant virus for in vivo gene therapy in a human. On April 16, 1993, a 23-year-old man with cystic fibrosis homozygous for the ΔF508 mutation of the CFTR gene was the first human to undergo in vivo gene therapy with administration of an E1−E3− recombinant adenovirus vector coding for the normal human CFTR cDNA to the nasal epithelium. One day later, using a fiberoptic bronchoscope, we administered 2×108 plaque-forming units of the same vector to the airway epithelium, an event that got worldwide media attention (Fig. 4; Supplementary Fig. S1; Supplementary Data are available online at
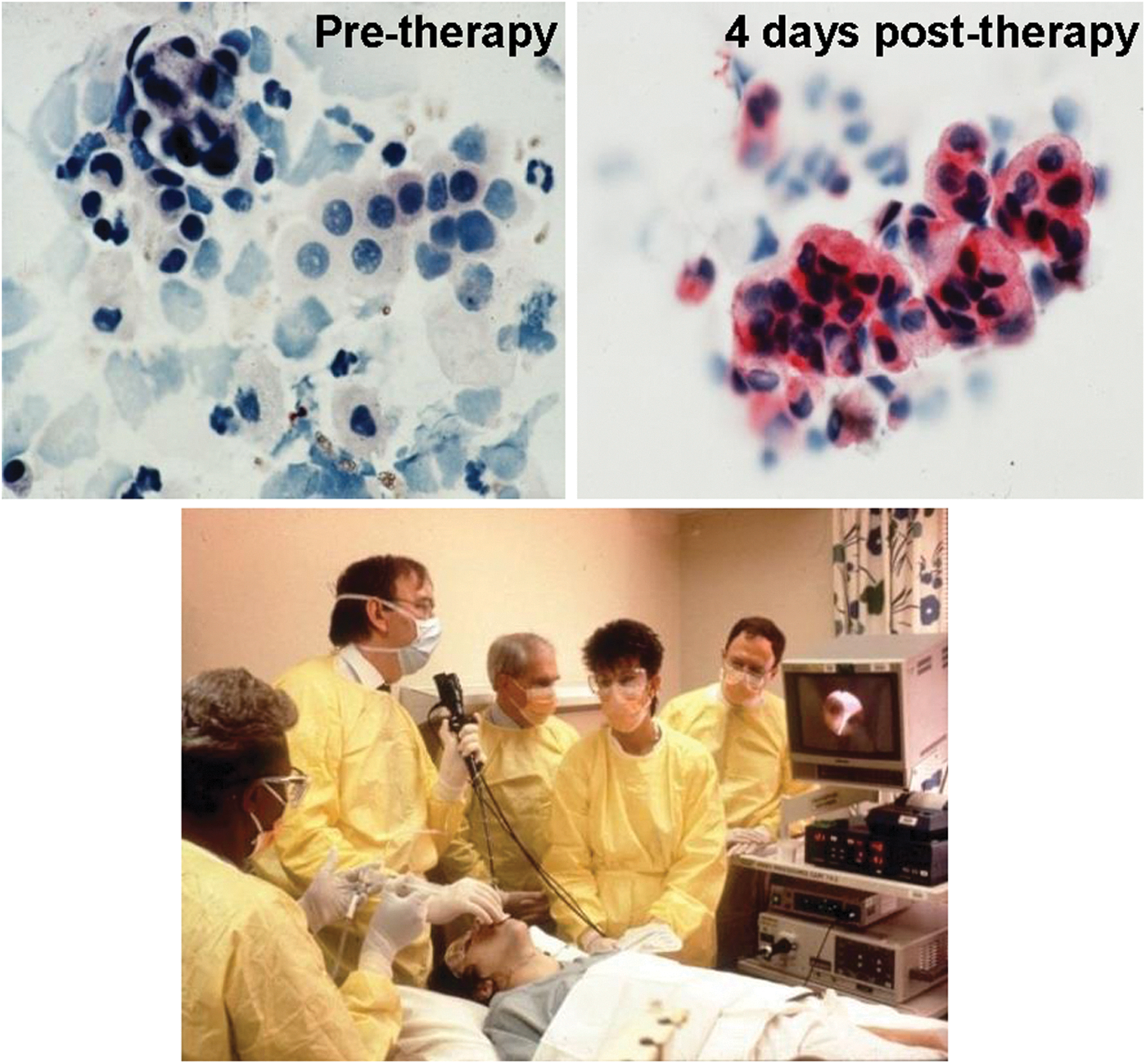
The first human gene therapy with a recombinant virus. On April 16, 1993, at the Clinical Center, NIH, a 23-year-old man with cystic fibrosis received an adenovirus coding for the normal human CFTR cDNA to the nasal epithelium. On the next day, the patient underwent fiberoptic bronchoscopy and 2×108 plaque-forming units of the vector was delivered through a catheter to the bronchial epithelium. The airways can be seen on the monitor. In the photo, left to right: staff nurse delivering the vector via a syringe, G. McElvaney, R. Crystal, staff nurse, and J. Hay. Successful gene transfer to the airway epithelium was demonstrated by antihuman CFTR antibody detection of CFTR before (left) and 4 days after (right) vector administration. The CFTR protein is stained red (Rosenfeld et al., 1992).
Limits Secondary to Antiadenovirus Immunity
After our initial Science article, our laboratory, along with many others, carried out a number of studies showing that adenovirus vectors were effective at transferring genes to most organs in vivo (for a list of our adenovirus vector studies from 1991 to present, see the Supplementary Material). In our initial enthusiasm, however, we and others did not recognize an important biologic fact that the human adenovirus is highly immunogenic, and that this immunogenicity would limit the time of expression, despite how highly efficient gene transfer is by adenovirus (Yang et al., 1994; Worgall et al., 1997; Hackett and Crystal, 2008; Hartman et al., 2008; Nemerow, 2009; Zaiss et al., 2009; Shayakhmetov, 2010; Thaci et al., 2011). This was demonstrated by many laboratories in experimental animals (Yang et al., 1996; Suzuki et al., 1998; Hartman et al., 2008), and strikingly demonstrated to us in our subsequent human study of repetitive airway epithelial gene therapy for cystic fibrosis (Harvey et al., 1999). This study demonstrated that we could achieve therapeutic levels of CFTR mRNA in the airway epithelium of patients with cystic fibrosis with the first administration, but this would quickly wane, and subsequent administrations were limited by immunity against the vector, with reduced yield of expression with each repeat of administration (Fig. 5). This combined innate, humoral, and cellular immunity evoked by the adenovirus vectors was eventually recognized to be responsible for the short, 1–2-week expression of adenovirus vectors administered in vivo to immunocompetent experimental animals and humans and was responsible for dose-limiting adverse events (Yang et al., 1994, 1996; Jooss and Chirmule, 2003).
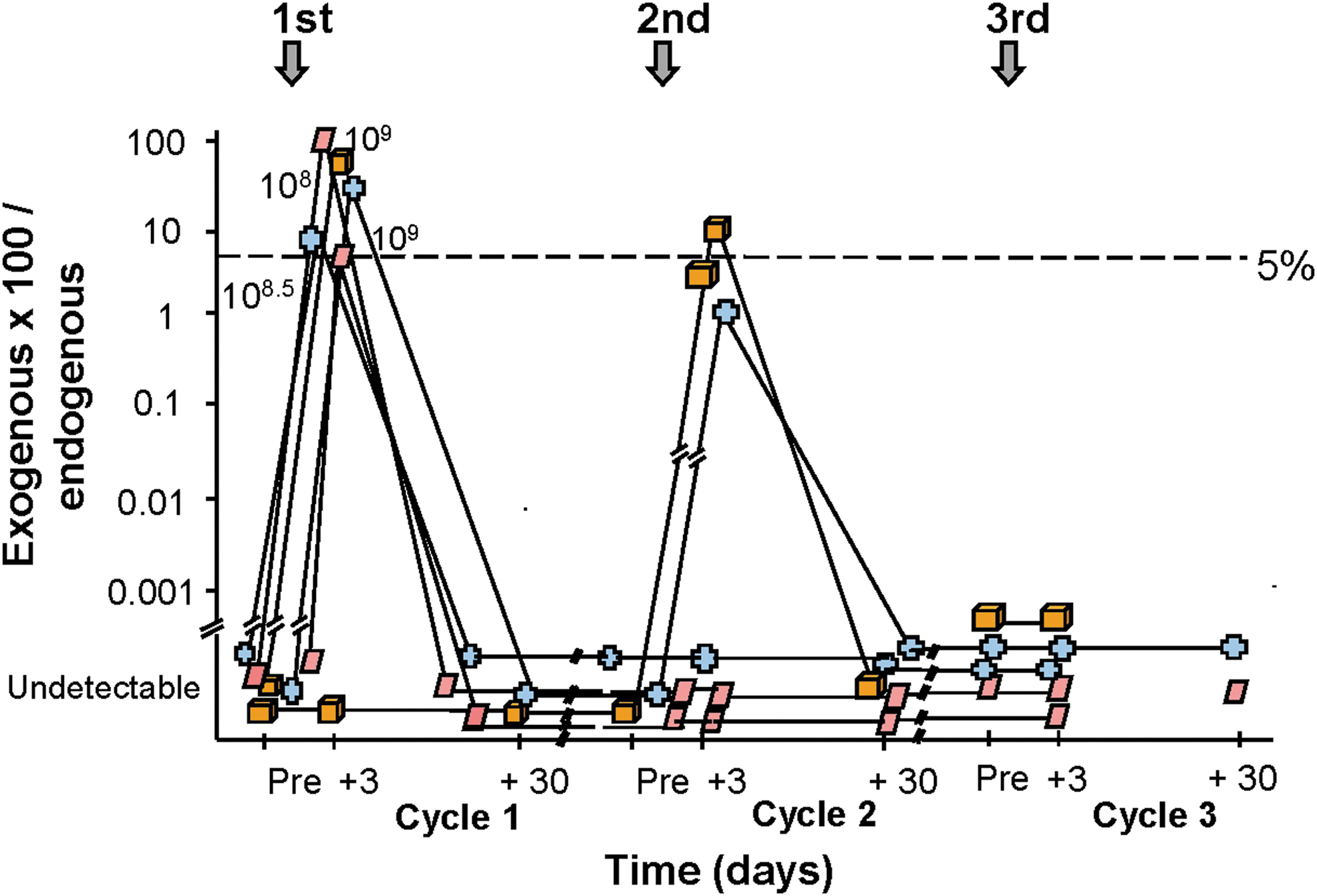
Quantitative assessment of the airway epithelium for the percentage of exogenous CFTR mRNA derived from the adenovirus vector compared with endogenous CFTR mRNA (individual's own CFTR gene expression) as a function of dose and time (baseline, days 3 and 30) after endobronchial spray of the first administration (cycle 1), second administration (cycle 2), and third administration (cycle 3) of the vector. The dashed line represents the target 5% level of exogenous vector-derived CFTR mRNA, that is, the level above which there should be sufficient levels of normal CFTR mRNA to correct the defect. Each symbol represents a different individual. Note that correction was achieved 3 days after the first administration (vector-derived mRNA levels all above the 5% level needed for correction), but this wanes by 30 days. Repeat administration (cycle 2) barely achieved this level, and the third administration resulted in no vector-derived CFTR mRNA expression. Adapted from Harvey et al. (1999).
Circumventing Antiadenovirus Immunity
The initial approach by us and others to limit the immunity evoked by adenovirus vectors was to either remove the responsible adenovirus genes from the vectors (such as the E2 and E4 genes and others) or add immune-suppressing genes to be expressed by the adenovirus vector (Gao et al., 1996; Raper et al., 1998; Weitzman, 2005; Bangari and Mittal, 2006; Dharmapuri et al., 2009; Seregin and Amalfitano, 2009). However, despite the elegant design of the molecular strategies to achieve this, these attempts to limit antivector immunity proved fruitless: the adenovirus is too immunogenic in humans, and the immune system too subtle in its ability to recognize adenovirus epitopes (Hackett et al., 2000; Weitzman, 2005; Yang et al., 2007; Dharmapuri et al., 2009; Sack and Herzog, 2009; Thacker et al., 2009; Ahi et al., 2011; Thaci et al., 2011). The only glimmer of hope in creating a “stealth” adenovirus vector has been the development of helper-dependent, so-called “gutless” adenovirus vectors, where all of the adenovirus genes are removed, with the necessary genes to create the vector provided by the producer cells (Kochanek et al., 2001; Kushwah et al., 2007; Segura et al., 2008; Dharmapuri et al., 2009; Thacker et al., 2009; Brunetti-Pierri and Ng, 2011; Vetrini and Ng, 2011). While the experimental animal studies with these vectors are encouraging, the effectiveness of “gutless” adenovirus vectors has not as yet been demonstrated in human trials.
An alternative approach to providing longer-term gene expression is based on the development of adenovirus vectors of different serotypes. While the first adenovirus studies were carried out with human serotype 5, and to a lesser extent human serotype 2, subsequent studies have led to identification of multiple adenovirus serotypes, both human and nonhuman (Dobbelstein, 2003; Bangari et al., 2005; Stone and Lieber, 2006). This led to the concept of “seroswitch,” that is, to circumvent the antiadenovirus immunity elicited by the initial administration of an adenovirus gene transfer vector by administering an adenovirus vector comprised of a different serotype carrying the same gene, a strategy that is effective in experimental animals (Mastrangeli et al., 1996; Mack et al., 1997). However, while seroswitch will provide another 1–2 weeks of expression, antiadenovirus immunity against the second serotype will obviate longer-term expression.
Capitalizing on Antiadenovirus Immunity
While the immunogenicity of the adenovirus as a gene transfer vector limits the length of time of expression to a few weeks, this can be leveraged for specific therapeutic applications. First, short-term expression is ideal for clinical applications in which the goal is to build new biologic structures.
A good example is the use of adenovirus to express angiogenic genes such as vascular endothelial growth factor (VEGF) in the myocardium to generate new coronary vasculature. In this application, the antivector immunity limits expression of the VEGF to 1–2 weeks, which is ideal for initiating angiogenesis and short enough to prevent excess blood vessels and hemangioma formation (Mack et al., 1998; Patel et al., 1999). Together with Todd Rosengart (then a Weill Cornell cardiothoracic surgeon and now Chairman of Surgery at Baylor), we carried out the first human studies with direct administration to the heart of individuals with diffuse coronary artery disease of an adenovirus vector coding for VEGF 121, one of the two isoforms of VEGF (Fig. 6) (Rosengart et al., 1999a,b, 2013). In a new-generation strategy for cardiac angiogenesis, we are initiating a double-blinded, placebo-controlled trial for cardiac angiogenesis with a serotype 5 adenovirus coding for all three isoforms of VEGF, a construct that is 10- to 100-fold more potent than any single isoform (Rosengart et al., 1999a, 2013; Crystal et al., 2012). Another good example of capitalizing on antivector immunity to limit gene expression was our demonstration that intradermal administration of an adenovirus coding for sonic hedgehog would provide a burst of sonic hedgehog expression to resting hair follicles, with the resulting induction of hair growth (Fig. 7) (Sato et al., 1999).
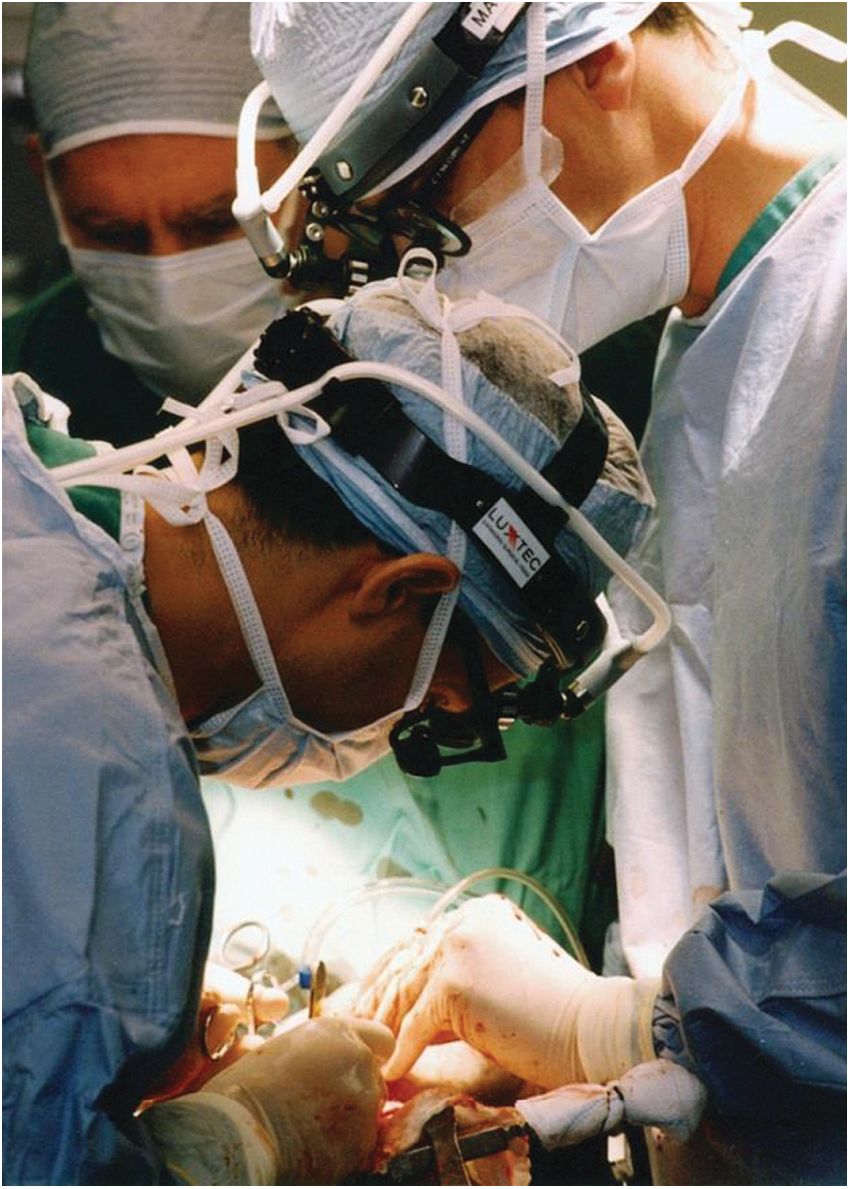
The first human trial of inducing cardiac angiogenesis with an adenovirus gene transfer vector. In 1997, we initiated a clinical trial to treat individuals with diffuse coronary artery disease with direct cardiac administration of an adenovirus coding for vascular endothelial growth factor (VEGF) 121 (Rosengart et al., 1999a,b, 2013). Clockwise, left to right: T. Rosengart, R. Crystal, and K. Krieger.
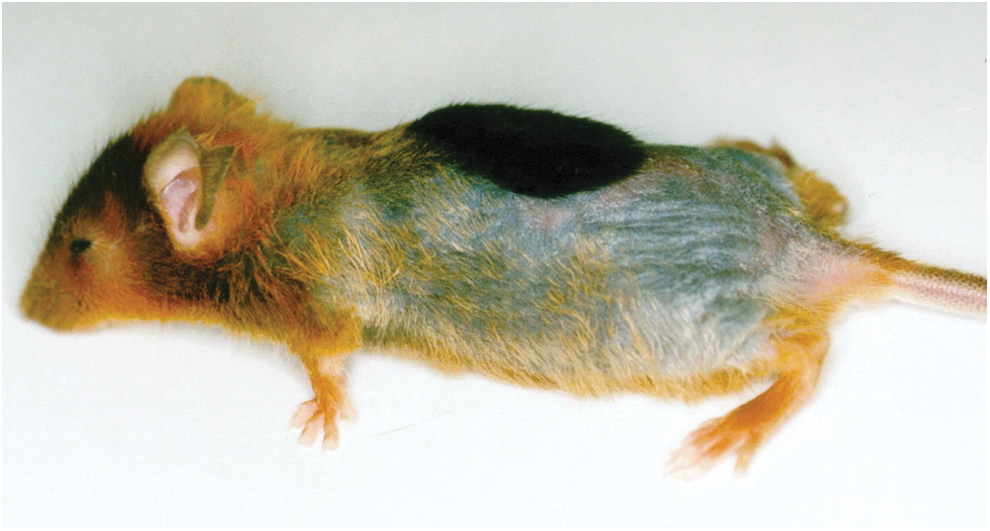
Induction of hair growth in a C57Bl/6 mouse 2 weeks after administration of an adenovirus vector coding for sonic hedgehog. To visualize hair growth, the hair of the mouse was bleached with blond hair dye to provide contrast for assessing new growth of the natural black hair of the mouse. The tuft of black hair is apparent (Sato et al., 1999).
Second, short-term expression is also ideal for applications where the goal is to destroy cells, as is the goal of anticancer strategies. This approach has been used to deliver directly to tumors cytotoxic genes, suicide genes that convert prodrugs to cytotoxic drugs, and immune-related genes to attract and/or activate local antitumor immunity (Wu et al., 2005; Tagawa et al., 2008; Matthews et al., 2009; Fukazawa et al., 2010; Aguilar et al., 2011; Aurisicchio and Ciliberto, 2012; Duarte et al., 2012; Deisseroth et al., 2013). Elegant molecular engineering strategies have been designed to target such vectors specifically to tumors (Kanerva et al., 2002; Hedley et al., 2006; Khare et al., 2011; Beatty and Curiel, 2012; Hangalapura et al., 2012; Kaufmann and Nettelbeck, 2012). Finally, with the recognition that antiadenovirus immunity is vigorous, many groups have made it even more so by generating replication-competent adenoviruses as therapeutic strategies to treat cancer, that is, to enlist the antiadenovirus immunity in killing cancer cells (Short and Curiel, 2009; Eager and Nemunaitis, 2011; Alemany, 2012; Cerullo et al., 2012; Russell et al., 2012; Choi and Yun, 2013).
Third, we and others have leveraged the potent immunity against the adenovirus to create vaccines against the transgene (Krause and Worgall, 2011) or by inserting the antigen in the capsid (Lasaro and Ertl, 2009; Thacker et al., 2009; Matthews, 2011). We have also used this approach to develop vaccines against bioterrorism agents, such as anthrax and plague (Tan et al., 2003; Boyer et al., 2005; Hashimoto et al., 2005; Kasuya et al., 2005; Boyer and Crystal, 2006; Chiuchiolo et al., 2006; De et al., 2008; Sofer-Podesta et al., 2009; Boyer et al., 2010; Van Blarcom et al., 2010). In a variation of this approach, we have used adenovirus vectors to code for a monoclonal antibody against a pathogen, taking advantage of the rapid and robust adenovirus vector-mediated gene expression (8–12 hr) to generate immunity against a pathogen in a pandemic (Kasuya et al., 2005; Chiuchiolo et al., 2006). In the most recent iteration of this concept, we have developed vaccines against addictive drugs (cocaine or nicotine) by covalently attaching an analog of the addictive molecule to the adenovirus capsid proteins, with effective generation of humoral immunity against the addictive molecule (Hicks et al., 2011; Koob et al., 2011; Wee et al., 2012; De et al., 2013).
Adenovirus Vectors in 2014
With thousands of articles published regarding adenovirus vectors, and hundreds of human trials with adenovirus vectors, what have we learned and what is the role of adenovirus vectors in the gene therapist's armamentarium?
First, there is no question that the adenovirus is the most effective means of delivering genes in vivo. Most human cells express the coxsackie-adenovirus primary adenovirus receptor as well as the secondary integrin receptors (Sharma et al., 2009; Smith et al., 2010; Chen and Lee, 2013; Wolfrum and Greber, 2013), and thus are easily infected with adenovirus vectors, with consequent high of the transgene expression. However, because of the immunity evoked against the adenovirus capsid and low-level expression of adenovirus genes, the effective expression for adenovirus vectors in vivo in humans peaks at 1 week and is limited to about 2 weeks. This makes adenovirus vectors ideal for when the desired expression is short-term, such as for building new biologic structures. Adenovirus vectors can also be used in therapies for cancer, where inducing immunity against the cancer or directly killing the cancer cell is the goal. Finally, the combined immunity against the adenovirus together with the short time of expression is ideal for using the adenovirus as a platform for developing vaccines.
Second, despite all of the efforts to circumvent antivector immunity, we have learned that mother nature is smarter than gene therapy adenovirologists, and we have not been able to effectively circumvent the antiadenovirus immunity by engineering stealth adenovirus vectors. While gutless adenovirus vectors may be the answer, it is likely that the potent adenovirus capsid immunogens will outsmart the most clever vectorology.
Third, despite the concern over safety of using adenovirus vectors in relation to secondary innate and adaptive immune responses to the vectors, there is now extensive experience with adenovirus vectors in many different clinical applications, and the dose and routes that are safe are now well established (Crystal et al., 2002; Harvey et al., 2002; Journal of Gene Medicine, 2013). Worldwide, adenovirus vectors are the most common vector used in clinical trials: to date, there have been 476 human gene therapy studies that have been initiated using adenovirus vectors, representing 23.5% of all gene therapy trials (Journal of Gene Medicine, 2013).
Fourth, adenovirus vectors have been a fruitful laboratory for developing strategies to test whether modification of viral capsids can help to enhance therapeutic properties of the virus, such as organ (or cancer)-specific homing. These design strategies will continue, and will be helpful in limiting adverse events while allowing higher doses.
Finally, the conventional way to manufacture adenovirus vectors is in cell lines expressing the adenovirus E1 genes with subsequent purification of the produced vectors. These production strategies will undoubtedly improve in both efficiency and purity of intact, functional vectors.
In summary, in the 23 years since our initial demonstration of the effectiveness of in vivo gene transfer using recombinant replication-deficient adenovirus vectors, together with adeno-associated viruses and lenti/retrovirus vectors, the adenovirus now represents one of the three major viral vector categories in the gene therapy tool kit. Through studies with adenovirus vectors, we have come to understand the importance of immunity, both preexisting and that evoked by the vector, in the efficacy and safety of viral vectors. Adenoviruses have been important tools to explore issues of tropism and regulatable expression and, importantly, opened the door to effective clinical gene therapy. For high-level, short-term expression or for purposefully evoking immunity, the adenovirus vectors stand alone as the choice for the gene therapist. Through some serendipity, luck at being at the right place at the right time, and collaboration with many talented postdoctoral fellows and collaborators throughout the world, I was able to play a part in the development of the adenovirus vector system and translation of its use to the clinic.
Footnotes
Acknowledgments
I would like to thank my many postdoctoral fellows and collaborators at NIH, Weill Cornell, and throughout the world who contributed to the early development of the adenovirus as a gene transfer vector and the translation of the adenovirus technology to gene therapy to humans.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.