
Abstract
Adenovirus type 5 (Ad5) is a commonly used vector for gene therapy, but its efficacy is limited by high seroprevalence and off-target hepatic and splenic sequestration. In order to circumvent these limitations, the use of vectors derived from rare species adenoviruses is appealing. The opportunity to retarget rare species vectors to defined cell types through the incorporation of peptide ligands would be advantageous, particularly in targeting tumors and disseminated metastases. We used predictive structural modeling to assess the CD, DG, HI, and IJ loops of the Ad48 fiber knob and identify optimal incorporation locales for the 20-mer peptide, A20FMDV2 (A20). A20FMDV2 targets ανβ6 integrin, which is overexpressed in human carcinomas. Recombinant Ad48 fiber knob proteins Knob48, Knob48-CD-A20, Knob48-DG-A20, Knob48-HI-A20, and Knob48-IJ-A20 were engineered and purified after expression in Escherichia coli. We confirmed that Knob48, Knob48-CD-A20, and Knob48-IJ-A20 formed stable homotrimers. However, Knob48-DG-A20 and Knob-HI-A20 failed to form a trimer. All A20-modified knob proteins blocked the transduction of Ad5-EGFPA20 via ανβ6, demonstrating that the inserted A20 peptide was functional. In conclusion, we show that the CD and IJ loops of Ad48 represent suitable sites for targeting peptide incorporation. Interestingly, in vitro gene transfer mediated by the non-factor-X-binding Ad48 vector was not sensitive to immunoglobulins and complement when incubated in the presence of mouse serum, unlike Ad5. These data support the future generation of the corresponding Ad48 viral vectors, Ad48-CD-A20 and Ad48-IJ-A20, which may offer favorable characteristics for targeted delivery in vivo.
Introduction
T
Vector engineering has been facilitated by the resolution of the crystal structure of the Ad5 fiber knob domain (Xia et al., 1994). The fiber knob exists as a homotrimer, with the topology of an eight-stranded, antiparallel β-sandwich, with interspersing loop regions (Xia et al., 1994). These loop regions vary from 8-55aa residues and are generally designated the AB, CD, DG, GH, HI, and IJ loops (Xia et al., 1994; Santis et al., 1999). The insertion of targeting ligands within the flexible, surface-exposed HI loop is the most common approach to retargeting Ad5 (Coughlan et al., 2010). However, the CD and IJ loops of Ad5 also have been shown to successfully accommodate the RGD-4C peptide ligand and permit engagement with surface expressed αν-integrins (Lord et al., 2006). Selected ligands have included peptides that contain RGD motifs (Dmitriev et al., 1998; Belousova et al., 2002; Coughlan et al., 2009, 2012b) or various cell-selective peptides identified by phage display technology (Xia et al., 2000; Nicklin et al., 2001; Work et al., 2004; Denby et al., 2007).
It is thought that successful retargeting of Ad5 to disease-specific targets may help overcome poor delivery and help to reduce vector-related toxicity. We previously reported the successful retargeting of Ad5 to ανβ6 integrin (Coughlan et al., 2009, 2012b) through the HI-loop incorporation of an ανβ6-selective ligand (A20FMDV2), derived from foot and mouth disease virus (FMDV2) (DiCara et al., 2007, 2008). This integrin is overexpressed in numerous human carcinomas (Sipos et al., 2004; Thomas et al., 2006) and has been independently identified as a prognostic indicator (Ahmed et al., 2002; Bates et al., 2005; Hazelbag et al., 2007). The tissue expression of ανβ6 is limited to epithelial remodeling events, including carcinogenesis and wound healing, and is not normally expressed on adult epithelia (Breuss et al., 1995).
However, Ad5-based vectors are limited by high seroprevalence in humans (Parker et al., 2009; Cheng et al., 2010; Barouch et al., 2011; Bradley et al., 2011a,b) and a natural tropism for hepatocytes after iv delivery (Kalyuzhniy et al., 2008; Vigant et al., 2008; Waddington et al., 2008) in multiple animal models (Alba et al., 2010, 2011). The hexon of Ad5 binds to coagulation factor X (FX) with high affinity (Kalyuzhniy et al., 2008; Waddington et al., 2008), and in vivo, liver gene transfer is facilitated by bridging of this Ad5:FX complex to heparan sulfate proteoglycans expressed on the surface of hepatocytes (Bradshaw et al., 2010; Duffy et al., 2011). This represents a major challenge to achieving high-efficiency extrahepatic delivery after iv delivery. Furthermore, it recently has been shown that FX binding to the Ad5 hexon facilitates evasion from neutralization by natural immunoglobulins and complement in serum from mice (Xu et al., 2013). Therefore, the innate sensitivity of Ad-based vectors to inactivation in serum, particularly those that do not bind FX, also should be considered when developing viruses for applications that required iv delivery. The undesirable attributes associated with Ad5 have prompted the investigation of diverse Ad species, with the aim of developing optimal platform vectors for gene therapy. Many species of D adenoviruses have low seroprevalence in humans (Abbink et al., 2007; Barouch et al., 2011) and do not bind to FX (Waddington et al., 2008). Therefore, these viruses represent an attractive alternative to Ad5 as they have the potential to evade not only liver gene transfer but also preexisting Ad5 immunity. We have recently shown that Ad48 has reduced accumulation in the liver and largely targets MARCO+marginal zone macrophages surrounding the white pulp in the spleen after iv delivery (Coughlan et al., 2012a). It previously has been demonstrated that adenoviral vectors with increased splenic uptake can successfully be retargeted to defined tissues (Alba et al., 2010). Therefore, efforts to retarget Ad48 and investigate its potential as a liver-detargeted platform vector for cancer gene therapy are warranted.
In contrast to the large number of studies that have assessed the suitability of Ad5 for selective retargeting, such approaches using rare serotype Ad vectors remain in their infancy, although fiber pseudotyping is commonly employed (Chiu et al., 2001; Nakayama et al., 2006; Parker et al., 2013). Attempts previously have been made to genetically modify the fiber of species B (Ad35) (Matsui et al., 2009), D (Ad19p fiber) (Denby et al., 2007), and F (Ad41 short fiber) serotypes (Hesse et al., 2007). The insertion of peptides within the EG, HI, and IJ loops of these fiber knobs was found to be compatible with fiber trimerization and resulted in successful modification of tropism (Denby et al., 2007; Hesse et al., 2007). In this study, we have assessed the feasibility of peptide incorporation into the CD, DG, HI, or IJ loops of the Ad48 fiber knob. The candidate peptide, A20FMDV2, was selected because of its tumor selectivity in human carcinomas, making it a clinically relevant targeting ligand for assessment. Modified Ad48 fiber knob proteins were produced as recombinant proteins, assessed for their structural integrity and the functionality of the inserted A20FMDV2 peptide.
Materials and Methods
Predictive modeling of recombinant knob proteins
Chimeric A20-modified fiber protein sequences were created using VectorNTI software and were subjected to structural modeling analysis using the automated mode of SWISS-Model (Guex and Peitsch, 1997; Schwede et al., 2003; Arnold et al., 2006). Predicted models for the fiber of Ad48 were based on the known structure of Ad37, selected automatically by the software. The propensity for post-RGD helix formation within the A20FMDV2 peptide was modeled using AGADIR software.
Construction of expression vectors
The pQE30 expression plasmid containing the wild-type Ad5 knob (Knob5) or Ad5 knob containing the A20FMDV2 peptide within the HI loop (Knob5-A20) previously has been described (Coughlan et al., 2009). Genomic DNA corresponding to aa168–373 of the Ad48 fiber knob domain (GenBank; EF153473.1) was synthesized by GeneArt (Invitrogen) and named pMA-Knob48. Engineered BamHI/HindIII restriction sites flanking the pMA-Knob48 sequence were incorporated to facilitate subcloning into pQE30. Smaller fragments of sequence that corresponded to the CD, DG, HI, and IJ loops containing the A20FMDV2 peptide were designed and synthesized so as to allow subsequent assembly of full chimeric Knob48 sequences by SOEing polymerase chain reaction (PCR; splicing by overlap extension) (Horton, 1995), a method that permits hybridization of overlapping complementary sequences within PCR fragments (see Fig. 1A for a schematic overview of primer design). The assembly of Knob-CD-A20 was performed as follows: Sequence containing A20FMDV2 within the CD loop was synthesized by GeneArt (pMA-CD-A20), beginning at the BamHI site to the PKPPT motif (aa168–297). The sequence corresponding to the left arm of the protein was amplified using BamHI-For 5′-TTTAAGGAT CCGGTGCCATTACAGTAGGAA-3′ and PKPPT-Rev 5′-
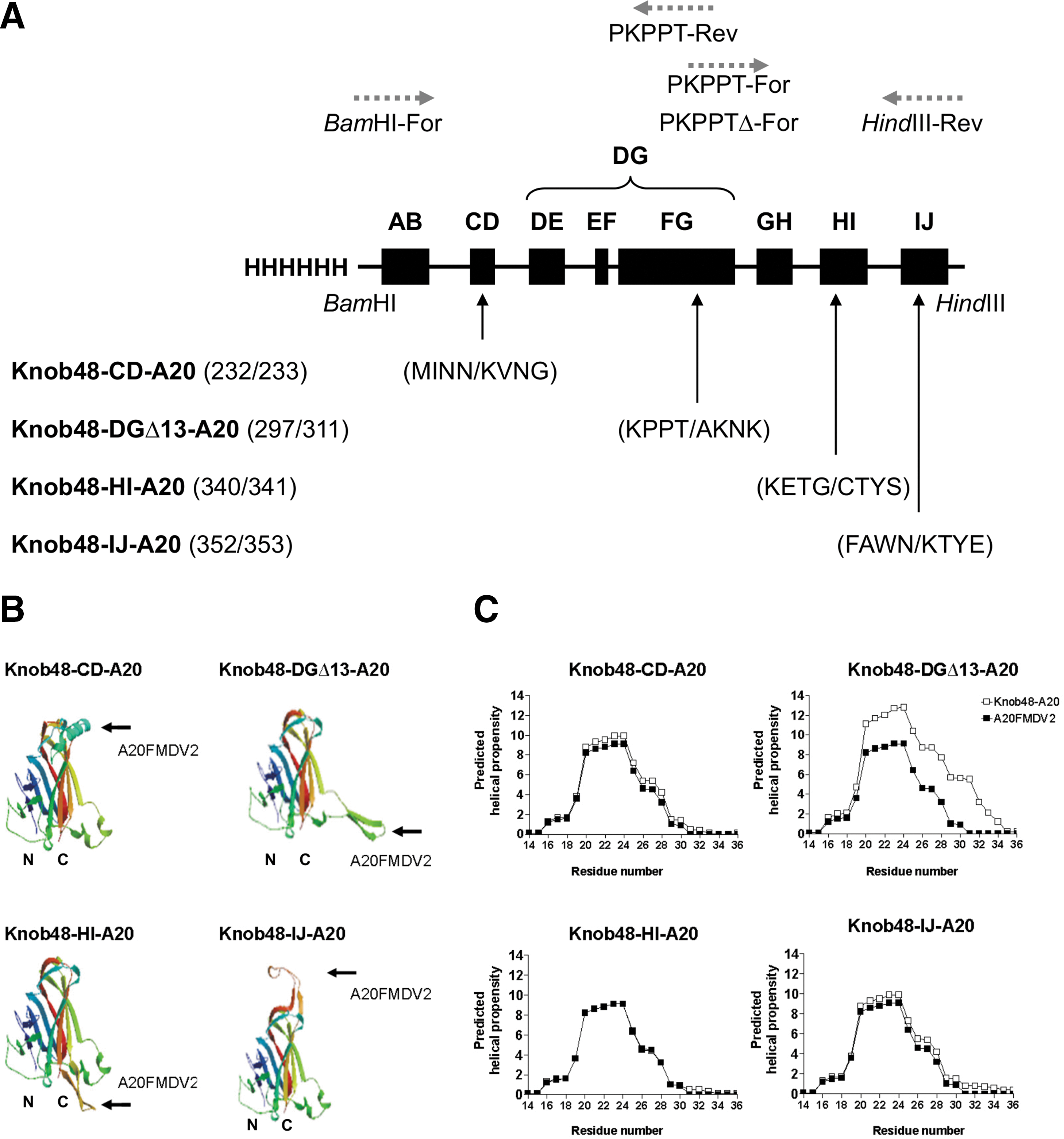
Construction and structural modeling of modified Ad48 fiber knob domains.
Ad48 fragments containing the A20FMDV2 peptide in the HI/IJ loops were ordered from GeneArt. The left arms of the chimeric Knob48-HI-A20 or Knob48-IJ-A20 were generated by performing PCR on unmodified pMA-Knob48 using BamHI-For and PKPPT-Rev, as before. The right arms, containing the A20FMDV sequence, were both amplified using PKPPT-For/HindIII-Rev, using either pMA-HI-A20 or pMA-IJ-A20 as template DNA. The resulting chimeric PCR products were then subjected to a final round of PCR using BamHI-For and HindIII-Rev, designed to amplify the entire fragment. Products were ligated into the pQE30 expression plasmid and subsequently transformed into SG13009 Escherichia coli for recombinant protein production as described (Coughlan et al., 2009).
Recombinant protein expression
Recombinant protein expression and purification under native conditions was performed as described previously (Coughlan et al., 2009), with final yields improved when protein induction was performed at 20°C for 16 hr instead of 37°C for 4 hr. In order to assess protein solubility, aliquots of induced E. coli cultures were resuspended in lysis buffer [50 mM Tris pH8.0, 300 mM NaCl, 10 mM imidazole, 1% (v/v) NP40 nonidet substitute, 1 mg/ml lysozyme, 1 mM β-mercaptoethanol] and incubated on ice for 30 min. Suspensions were subjected to 6×10 sec rounds of sonication at 200–300 W with 10 sec pauses between. Samples were analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The ability of recombinant knob proteins to form homotrimers was verified by semidenaturing SDS-PAGE, comparing boiled and unboiled samples. The concentrations of purified recombinant knob proteins were determined by Bicinchoninic Acid Assay (Perbio Science) as directed by the manufacturer.
Competitive inhibition using recombinant knob proteins
In order to confirm that the A20FMDV2 peptide was functional and capable of binding to its target receptor, ανβ6 recombinant knob proteins were tested for their ability to block Ad5-EGFPA20-mediated transduction or detection of ανβ6 integrin by flow cytometry. Both assays have been described in detail elsewhere (Coughlan et al., 2009). EGFP or ανβ6 receptor expression was detected by flow cytometry using a FACS Canto II (Beckton Dickinson) and analysis using FACS DIVA software, acquiring 10,000 gated events. To calculate IC50 values, dose–response curves were generated with GraphPad Prism version 4.0 (GraphPad Software) after nonlinear regression analysis.
Neutralization of Ad5 and Ad48 by naïïve murine serum
To investigate if Ad48 is also subject to the same serum sensitivity previously described for Ad5 (Xu et al., 2013), we assessed viral transduction of SKOV3 human ovarian carcinoma cells under normal conditions as well as in the presence of murine sera (which contains FX) or in the presence of sera+X-bp (to deplete FX present in sera). SKOV3 cells (1×104 cells/well) were selected as they are a low CAR expressing cell line, but retain expression of integrins and heparin sulfate proteoglycans used by adenoviral vectors for cell entry (Bradshaw et al., 2012). Serum from C57Bl/6 mice was separated from fresh whole blood, diluted to 80–90% in RPMI-1640 supplemented with 2% globulin-free BSA (Sigma-Aldrich) and incubated with 1.0×109 vp Ad5/Ad48 in a final volume of 50 μl. X-bp was added to similar test samples at 80 μg/ml to deplete FX in the serum. Controls consisted of Ad5/Ad48 alone in serum-free RPMI-1640 supplemented with 2% globulin-free BSA (Sigma-Aldrich). Vectors were incubated with serum or medium for 30 min at 37°C and then placed on ice. Mixtures were diluted 200-fold in serum-free RPMI-1640 medium (50 μl into 10 ml). Cells were rinsed with serum-free medium and 100 μl of diluted vector (1000 vp/cell) was added to triplicate wells for 2 hr at 37°C. The inoculum was then replaced with medium containing 2% heat-inactivated FBS. After approximately 16 hr, cells were rinsed with PBS and harvested for determination of luciferase activity (Luciferase Assay System; Promega) and protein content using the Bicinchoninic Acid Assay (Perbio Science).
Results
Predictive modeling of recombinant protein structure
Ad5 and Ad48 fiber knob protein alignments were generated using the ClustalW multiple sequence alignment program (EMBL-EBI). We selected peptide insertion sites within the CD, HI, and IJ loops (Fig. 1A) as identified by Lord and colleagues as suitable for peptide incorporation in Ad5 (Lord et al., 2006). The A20FMDV2 sequence was inserted immediately after amino acids at position Asn232 in the CD loop (MINN/KVNG), after Thr297 in the DG loop (KPPT/AKNK) with the deletion of 13aa, after Gly340 in the HI loop (KETG/CTYS), and after Asn352 within the IJ loop (FAWN/KTYE). We used predictive structural modeling to visualize the fiber knob of Ad48 and modified Ad48 knob proteins featuring the A20FMDV2 peptide within the CD, DG, HI, and IJ loops (Fig. 1B). The crystal structure of the Ad48 knob has not yet been determined. Therefore, we used the automated mode of SWISS-Model, which is based on the known crystal structure of Ad37 (Burmeister et al., 2004) (PDB code: 1uxaA). Predictive models showed that the A20FMDV2 peptide appeared to be externally exposed, projecting away from the Ad48 fiber knob. Therefore, we hypothesized that the A20FMDV2 peptide, within these conformations, could be accessible for binding to its target receptor, the integrin ανβ6.
It previously has been demonstrated that the specificity of the A20FMDV2 peptide for ανβ6 is dependent on the formation of a critical post-RGD, C-terminal α-helical loop and that residues flanking the peptide can affect this conformation (DiCara et al., 2007). Previous studies have used AGADIR, a freely available algorithm that predicts the helical behavior of monomeric peptides (Muñoz and Serrano, 1997), to theoretically predict if A20FMDV2 forms this post-RGD helix (DiCara et al., 2007). Therefore, we also used this approach to model if the A20FMDV2 peptide could assume its critical helical conformation within the CD, DG, HI, or IJ loops of modified Knob48 proteins (Fig. 1C). The AGADIR analysis included the A20FMDV2 peptide sequence within each loop, along with 10 adjacent amino acid residues corresponding to the flanking adenoviral sequence at each site (40aa in total). Each modified Knob48 sequence (filled square ▪) was compared with the native A20FMDV2 peptide sequence (open square □). The RGD motif begins at residue number 17–19 followed by the post-RGD sequence, LQVL. Predicted post-RGD percentage helix formation was as follows: 2.22% for Knob48-CD-A20, 3.43% for Knob48-DGΔ13-A20, 2.03% for Knob48-HI-A20, 2.38% for Knob48-IJ-A20, and 2.70% for the A20FMDV2 peptide alone. The algorithm prediction was favorable, with all configurations retaining similar helical propensity to the A20FMDV2 peptide alone. However, it is worth considering that the AGADIR software is predictive and does not confirm the ability to form post-RGD helices. Therefore, we decided to construct expression vectors for each of the A20-modified knob proteins in order to produce recombinant protein.
Production and elution of recombinant knob proteins
Denatured, boiled samples corresponding to each elution fraction were analyzed by SDS-PAGE. We successfully expressed and purified A20-modified recombinant Knob48 proteins and detected protein at the expected size of ∼25 kDa (Fig. 2A–D). Uninduced controls were also analyzed and did not contain a band of this size (data not shown). After SDS-PAGE analysis, elution fractions with high purity were pooled and dialyzed against PBS. With the exception of Knob48-HI-A20 (Fig. 2C), all recombinant proteins were produced to high yields (>0.5 mg/ml). Incorrect structural folding can result in protein proteolysis or can alter the solubility of the protein, affecting its yield. Therefore, in an attempt to explain the low yield obtained with Knob48-HI-A20, we tested its solubility (Fig. 2E). Aliquots were taken from induced E. coli cultures (16 hr) and cells pelleted by centrifugation. Cells were lysed in lysis buffer, lysates centrifuged, and crude cell lysates containing soluble protein separated from the pellet containing insoluble protein. Boiled and unboiled fractions for each were analyzed by SDS-PAGE. An uninduced negative control and positive control (purified Knob48) were run in parallel to estimate the correct MW band (data not shown). We detected a clear band of ∼25 kDa within the insoluble material that corresponded with the expected size of the Knob48-HI-A20 protein (Fig. 2E). This band was not as strong in the soluble protein fraction, indicating that the low yields observed with Knob48-HI-A20 were because of poor solubility of this particular recombinant protein.
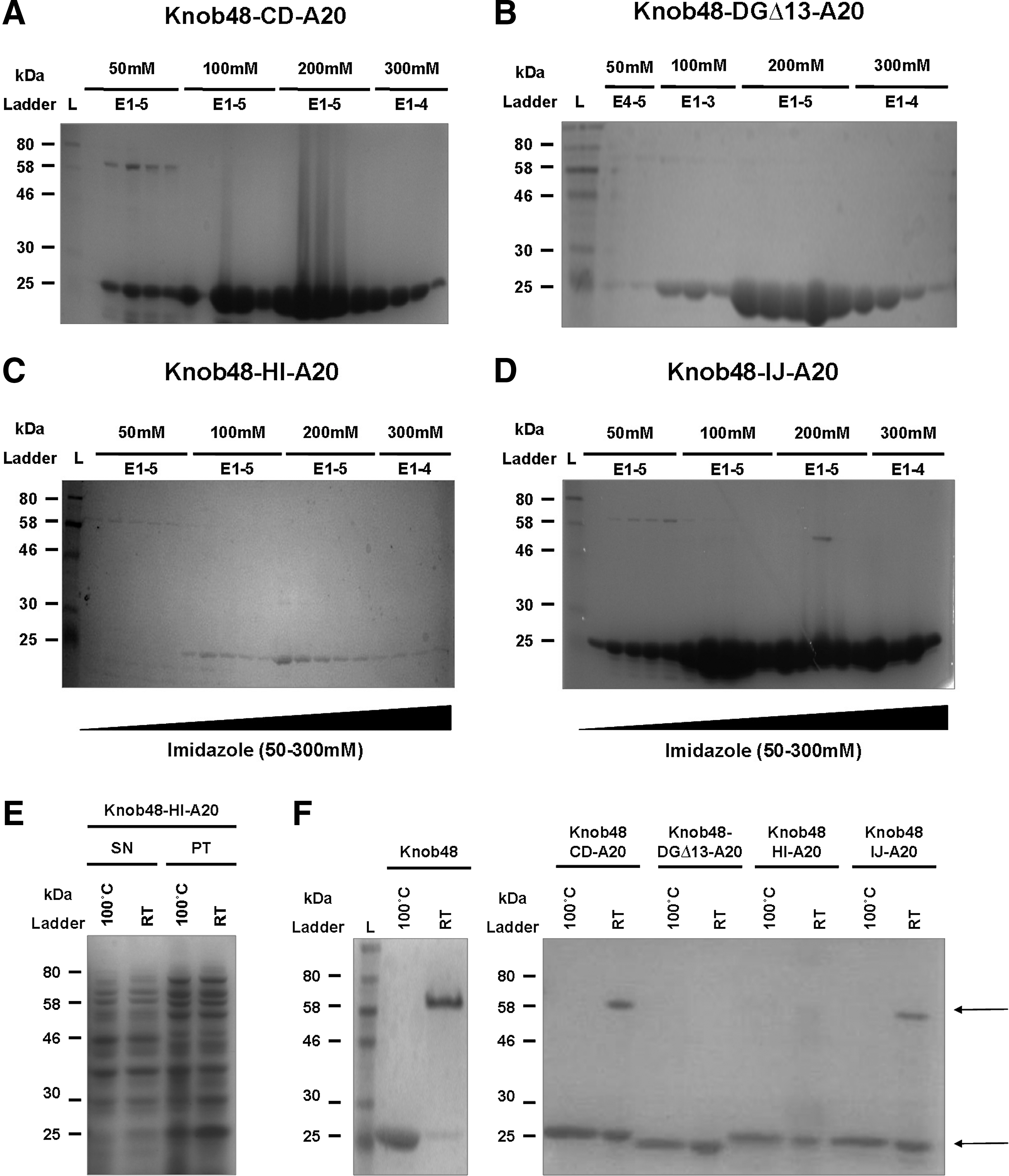
Recombinant protein production and characterization.
Characterization of modified knob proteins
Trimerization of the adenoviral fiber is critical for adequate viral assembly. Therefore, we tested the trimerization of purified Knob48 and all A20-modified Knob48 proteins, comparing 10 μg boiled and unboiled (room temperature, RT) fractions by SDS-PAGE (Fig. 2F). Under semidenaturing conditions (RT), the unmodified Ad48 fiber knob, Knob48, successfully formed a trimer with an observed size of ∼58 kDa. The fully denatured, boiled sample produced a band at ∼25 kDa in agreement with the predicted size of the Knob48 monomer. Both Knob48-CD-A20 and Knob48-IJ-A20 formed intact trimers, detected at the correct size. Bands corresponding to the monomeric size were also observed, because of partial denaturation of the proteins by SDS in the loading buffer. We did not detect bands for Knob48-DGΔ13-A20 or Knob-HI-A20 at the expected trimer size, suggesting that the insertion of the A20FMDV2 peptide had compromised the structural integrity of the fiber knob.
The successful multimerization of modified fiber knob proteins after insertion of targeting peptides does not guarantee the retention of peptide functionality. In order to confirm that the A20FMDV2 peptide was functional and capable of binding to its target receptor, the integrin ανβ6, we performed a competition assay using each recombinant knob protein to inhibit the transduction of Ad5-EGFPA20 (Fig. 3A). We previously have shown that Ad5-EGFPA20 transduces BT-20 cells (low CAR/high ανβ6) with high efficiency in a ανβ6-dependent manner (Coughlan et al., 2009). Positive controls, Knob5-A20 and anti-ανβ6 antibody 53A.2, both blocked the transduction of Ad5-EGFPA20 (p≤0.0001), whereas Knob5 had no effect (p=0.31). Importantly, all A20-modified Knob48 proteins, but not Knob48, inhibited Ad5-EGFPA20-mediated transduction to a similar extent as Knob5-A20, indicating that the inserted peptide was functional.

Characterization of modified knob proteins.
As Knob48-CD-A20 and Knob48-IJ-A20 were the only modified fiber knobs that trimerized and would therefore be suitable for engineering a retargeted Ad48 vector, we determined their IC50 values, to compare this value to the previously described IC50 for Knob5-A20 (Coughlan et al., 2009). BT-20 cells were preincubated with increasing concentrations of Knob48 (−control), Knob5-A20 (+control), Knob48-CD-A20, or Knob48-IJ-A20 (Fig. 3B), in order to block the detection of ανβ6 integrin using the anti-ανβ6 antibody, 53A.2. Knob48 did not inhibit the detection of ανβ6 at any concentration tested, whereas Knob5-A20, Knob48-CD-A20, and Knob48-IJ-A20 inhibited detection of ανβ6 with IC50 values of 0.04, 0.51, and 0.68 μg/105 cells, demonstrating that the A20FMDV2 peptide insertion was functional in blocking its target receptor, ανβ6.
Neutralization of vectors by naïve murine serum
It recently has been shown that FX binding to the Ad5 hexon appears to have an auxiliary role in shielding the virus from immune attack (Xu et al., 2013), in addition to the previously described effect in mediating hepatocyte transduction via engagement with heparin sulfate proteoglycans (Kalyuzhniy et al., 2008; Waddington et al., 2008; Alba et al., 2009, 2010; Bradshaw et al., 2010). However, it is not clear whether adenovirus serotypes that do not bind FX are sensitive to similar effects from serum as are observed when FX binding to Ad5 is prevented. Therefore, we compared the sensitivity of Ad5 and Ad48 to neutralization by murine sera using an in vitro assay (Fig. 4). The transduction of Ad5 in low CAR ovarian carcinoma cell line, SKOV3, was enhanced ∼1.8-fold in the presence of murine sera. In the presence of X-bp, which binds to the Gla domain of FX with high affinity, blocking binding to Ad5 hexon (Waddington et al., 2008), transduction was significantly decreased, consistent with the findings of a previous study (Xu et al., 2013). Importantly, in the presence of murine sera with or without X-bp, transduction of SKOV-3 cells mediated by Ad48, a known non-FX-binding serotype (Waddington et al., 2008; Coughlan et al., 2012a), was not affected, suggesting that Ad48 is not sensitive to the same mechanism that mediates immune attack on Ad5 vectors that are devoid of FX binding. These data further support the suitability of Ad48 for systemic delivery strategies.

Neutralization of Ad5 and Ad48 by naïve murine serum. Ad5 or Ad48 (1×109 vp) were incubated with RPMI-1640+2% BSA, 80–90% murine serum in RPMI-1640+2% BSA, or 80–90% murine serum in RPMI-1640+2% BSA+X-bp, for 30 min at 37°C. Virus suspensions were diluted 200-fold in serum-free RPMI-1640 and 100 μl added to low CAR-expressing SKOV3 cells, seeded in triplicate at 1×104 cells/well (MOI 1000 vp/cell), for 2 hr at 37°C before virus was washed off. Luciferase transgene expression was quantified ∼16 hr posttransduction and relative light units (RLU) normalized to mg total protein. Graph shows transduction as a percentage of control (Ad5/Ad48 transduction with serum free media alone). **p<0.01.
Discussion
In this preliminary study we have identified functional peptide insertion sites within the fiber knob domain of Ad48. We have demonstrated that the CD and IJ loops of the Ad48 fiber tolerate the insertion of the 20aa tumor-selective peptide, A20FMDV2. Incorporation of the peptide at these sites had no detrimental effect on the trimerization of the fiber protein, a process that is critical in the viral assembly process. Although trimerization of recombinant fiber proteins cannot guarantee the successful rescue of modified Ad48 viral particles (Xia et al., 2000), these data suggest that such modifications could be compatible with the assembly of a fiber-modified Ad48 virus in the future. In support of this, the candidate A20FMDV2 peptide has already been incorporated into the fiber of Ad5 and virion integrity retained (Coughlan et al., 2009, 2012b). Importantly, we confirmed that the inserted A20FMDV2 peptide was capable of recognizing its target receptor, the integrin ανβ6, and that A20-modified recombinant Knob48 proteins could competitively inhibit ανβ6 in a dose-dependent manner. The IC50 values for Knob48-CD-A20 and Knob48-IJ-A20 were ∼10-fold higher than the A20-modified construct derived from Ad5, Knob5-A20. It is unclear how this may impact upon recognition of the target receptor ανβ6 in vivo. However, it may be possible to improve upon this in future constructs by combining peptide insertions within both the CD and IJ loops.
Previous attempts to retarget rare serotype Ads have identified the HI loop as a suitable site for peptide incorporation. Hesse and colleagues highlighted the suitability of the EG, HI, and IJ loops within the short fiber of species F adenovirus, Ad41, for the insertion of an integrin targeting Arg-Gly-Asn (RGD) motif (Hesse et al., 2007). The FG and HI loops of species B adenovirus, Ad35, also have been shown to tolerate the insertion of RGD-containing peptides (Matsui et al., 2009). Both retargeting approaches resulted in improved αν-integrin-mediated transduction efficiency in vitro. Importantly, efficient in vivo retargeting has been achieved after the insertion of renal targeting peptides into the HI loop of an Ad19p fiber pseudotyped vector (Denby et al., 2007). As the HI loop has been identified as a functional site for peptide incorporation in many rare species Ads, we were surprised to find that insertion of A20FMDV2 within the HI of the Ad48 fiber knob domain was incompatible with trimerization. It is possible that we were unable to detect trimerization because of the formation of large insoluble aggregates of Knob48-HI-A20 when this recombinant protein is produced in E. coli. However, it is possible that this fiber modification would be capable of trimerization when in the context of an intact virion assembling in mammalian cells. Furthermore, it is possible that the insertion of alternative targeting peptides, with varying sequence or length, within the DG or HI loops of the Ad48 fiber may permit trimerization.
The RGD motif is frequently used to retarget Ad vectors to αν-integrins. However, αν-integrins are widely expressed on normal tissue and are not restricted to malignant tissue. Conversely, the A20FMDV2 peptide selectively targets the integrin ανβ6 (DiCara et al., 2007, 2008), a prognostic indicator that is overexpressed in numerous human carcinomas but is largely absent in corresponding healthy tissue. We previously have demonstrated improved targeting to ανβ6+tumor xenografts in vivo after the incorporation of the A20FMDV2 peptide into the HI loop of Ad5 (Coughlan et al., 2009, 2012b). Furthermore, this vector had an improved safety profile, with reduced hepatotoxicity after iv delivery. It will be interesting to determine whether A20FMDV2 targeting peptide within a non-FX-binding virus Ad48 capsid could further improve upon these desirable in vivo characteristics.
In contrast to Ad5, the basic biology of Ad48 is not well determined, highlighted by the fact that the native receptors for many species D Ads have not conclusively been determined, although CAR (Abbink et al., 2007), CD46 (Abbink et al., 2007), and sialic acids (Burmeister et al., 2004) have all been implicated. We recently characterized the in vivo biodistribution profile of Ad48 and concluded that this vector possesses many attributes that would make it an attractive alternative platform vector to Ad5 (Coughlan et al., 2012a). These are namely its low seroprevalence in humans (Barouch et al., 2011), its failure to interact with FX and transduce the liver (Waddington et al., 2008), and its predictable cytokine profile, a reflection of its high-level accumulation in the spleen (Coughlan et al., 2012a). High-efficiency retargeting to αvβ6 integrin could potentially limit this splenic accumulation by redirecting to tumor xenografts and subsequently reduce cytokine responses to Ad48-based vectors.
The development of retargeted Ads based on rare species viruses is limited by the fact that vector systems for many rare species adenoviral vectors, including Ad48, are not as widely available to the research community as Ad5. In recent years, substantial efforts have resulted in the generation of FX-binding-ablated Ad5-based vectors for systemic delivery approaches (Alba et al., 2009, 2010). It may therefore be feasible to pseudotype hexon-modified Ad5 vectors with our retargeted Ad48 fibers or fiber knob domains. However, as recent work by Xu and colleagues has demonstrated that FX is exploited by Ad5-based vectors to shield the virus from attack by natural antibodies and complement (Xu et al., 2013), the development of non-FX-binding Ad vectors that are not sensitive to this immune inactivation may be preferable. We have shown in this study that Ad48 vectors are not susceptible to this serum neutralization and may therefore represent optimal platform vectors for future retargeting strategies, particularly when systemic delivery is the route of administration.
To our knowledge, this is the first attempt to assess the potential for engineering the fiber of Ad48 for retargeting strategies. These findings demonstrate the feasibility of modifying the fiber knob domain of Ad48 and provide valuable information for targeting peptide strategies within these sites. Therefore, these results facilitate opportunities to develop therapeutic Ad48 vectors with improved and optimized biological characteristics in the future.
Footnotes
Acknowledgments
H.U.-K. was funded by a Society for General Microbiology Vacation Studentship Award (VS10/73) and B.P.D. by a Nuffield Foundation Research Bursary (URB/39438), awarded to L.C. This work was also funded by the BBSRC (BB/G016844/1, awarded to A.H.B). A.H.B. is supported by the British Heart Foundation Chair of Translational Cardiovascular Sciences (CH/11/2/28733). We would like to thank Nicola Britton and Gregor Aitchison for their invaluable technical assistance. We also thank Dr. John F. Marshall, Barts Cancer Institute, for kindly providing the function blocking anti-ανβ6 antibody, 53A.2.
Author Disclosure Statement
The authors declare that they have no conflict of interest.