
Abstract
Inhibition of the proinflammatory transcription factor NF-κB has previously been shown to attenuate the inflammatory response in tissue after injury. However, the feasibility and efficacy of aerosolized adeno-associated viral (AAV) vector-delivered transgenes to inhibit the NF-κB pathway are less clear. Initial studies optimized the AAV vector for delivery of transgenes to the pulmonary epithelium. The effect of repeated nebulization on the integrity and transduction efficacy of the AAV vector was then examined. Subsequent in vivo studies examined the efficacy of aerosolized rAAV2/6 overexpressing the NF-κB inhibitor IκBα in a rodent endotoxin-induced lung injury model. Initial in vitro investigations indicated that rAAV2/6 was the most effective vector to transduce the lung epithelium, and maintained its integrity and transduction efficacy after repeated nebulization. In our in vivo studies, animals that received aerosolized rAAV2/6-IκBα demonstrated a significant increase in total IκBα levels in lung tissue relative to null vector-treated animals. Aerosolized rAAV2/6-IκBα attenuated endotoxin-induced bronchoalveolar lavage-detected neutrophilia, interleukin-6 and cytokine-induced neutrophil chemoattractant-1 levels, as well as total protein content, and decreased histologic indices of injury. These results demonstrate that aerosolized AAV vectors encoding human IκBα significantly attenuate endotoxin-mediated lung injury and may be a potential therapeutic candidate in the treatment of acute lung injury.
Introduction
A
Acquired inflammatory respiratory disorders such as chronic obstructive pulmonary disease (COPD), asthma, or ARDS, which occur because of various polygenic and exogenous influences, may be candidates for transient gene therapy approaches where short-term overexpression of protective genes or the suppression of damaging genes may attenuate inflammation. 13,14 In clinical ARDS, NF-κB pathway gene polymorphism activation can modify susceptibility to and severity of cases. 15,16 Cell activation initiated by stimuli such as gram-negative bacterial endotoxin activates various signaling pathways, which converge to phosphorylate and activate the IκB kinase complex proteins (IKK), which then phosphorylate and inactivate the IκB proteins, which dissociates from NF-κB. Activated NF-κB translocates to the nucleus and binds to specific cognate-binding sequences in the promoter or enhancer regions of various target genes to initiate transcription. 5 The evidence for potential therapeutic strategies emerging from inhibition of the NF-κB pathway is demonstrated by decreasing injury in nonseptic ALI models, including pulmonary ischemia–reperfusion, 17 endotoxemia, 9 and smoke-induced pulmonary inflammation. 18
Pulmonary aerosol deposition patterns may have significant implications for absorption profile and resulting pharmacokinetics, with deposition in the respiratory airways, typically resulting in greater absorption than that seen in the conducting airways. 19 –21 Direct liquid instillation to the pulmonary epithelium results in a heterogeneous pattern that deposits locally, predominantly in larger airways, which is of limited benefit when delivery to distal areas of the lung is required. Aerosolization offers the potential of a more diffuse, homogeneous pattern with deposition in the gas-exchanging smaller airways and alveolar regions. 22,23
Considering these issues, we wished to examine the feasibility and efficacy of aerosol delivery of transgenes to modulate the NF-κB pathway in the pulmonary epithelium. Initial studies optimized the adeno-associated viral (AAV) vector for delivery of transgenes to the pulmonary epithelium. We then examined the effect of repeated nebulization on the integrity and transduction efficacy of the AAV vector. Subsequent in vivo studies examined the efficacy of aerosolized rAAV2/6 overexpressing the NF-κB inhibitor, IκBα, in a rodent endotoxin-induced lung injury model. We hypothesized that (1) the AAV vector would efficiently transduce the pulmonary epithelium in vitro and in vivo, (2) aerosolization of AAV vector would not diminish its structural integrity or transduction efficacy, and (3) aerosolized AAV-IκBα would reduce the severity of endotoxin-induced lung injury in a rodent model.
Materials and Methods
rAAV2, rAAV2/5, and rAAV2/6 manufacture
The IκBα superrepressor, incorporating S32A and S36A mutations, is not susceptible to phosphorylation at these serines and therefore resistant to subsequent degradation. A plasmid carrying this mutated gene was a kind gift from R. Zwacka (National University of Ireland, Galway, Ireland).
Empty pTRUF and pTRUF-IκBα plasmids were packaged with the relevant AAV serotype vectors through dual transfection, with pDP helper-free REP/CAP plasmids (Plasmid Factory, Bielefeld, Germany), into HEK293T cells and isolated by subsequent iodixanol density gradient centrifugation. After DNase and proteinase K digestion, viral titer was quantified by real-time PCR (GeneAmp PCR 9600 system; PerkinElmer, Waltham, MA), using primers directed against the cytomegalovirus (CMV) promoter, as compared with a standard curve constructed with the pTRUF plasmid at defined copy number. Primers used were as follows: CMV promoter (sense, 5′-TGGAAATCCCCGT GAGTCAA-3′; antisense, 5′-CATGGTGATGCGGTTT TGG-3′).
Animal care
Specific-pathogen-free adult male Sprague Dawley rats (Harlan, Bicester, UK) weighing between 400 and 525 g were used in these experiments. All experimental work was approved by the Animal Care Research Ethics Committee of the National University of Ireland, Galway and conducted under license granted by the Department for Health, Ireland. On arrival, the rats were allowed to acclimatize for at least 1 week under standardized conditions of temperature (21–22°C), humidity (45–55%), and light (12 hr light:12 hr dark schedule). Animals had access to rat chow and tap water ad libitum.
Aerosol administration to rats
Animals were anesthetized with intraperitoneal ketamine (Ketalar, 80 mg·kg–1; Pfizer Healthcare, Dublin, Ireland) and inhalational isoflurane (Abbott Laboratories, Dublin, Ireland) at a concentration of 2% in oxygen (fraction of inspired oxygen [F
Endotoxin/vehicle instillation
The experimental injury model was based on those previously reported, with several modifications. 25,26 Five days after vector/vehicle instillation, rats were reanesthetized and endotoxin (lipopolysaccharide [LPS]; 5 mg·kg−1) derived from Escherichia coli serotype 055:B5 (Fluka, Poole, UK) in 0.3 ml of normal saline or vehicle was instilled intratracheally.
Primary cell isolation and cell line culture
Animals were anesthetized with intraperitoneal ketamine (Ketalar, 80 mg·kg–1; Pfizer Healthcare) and xylazine (Xylapan, 8 mg·kg–1; Vétoquinol, Dublin, Ireland) and then killed by exsanguination. Primary type II epithelial cells isolated from Sprague Dawley rats (PT2SD) were cultured for a minimum of 24 hr before subsequent experimental use. 27 A549 cells (adenocarcinomic human alveolar basal epithelial cells; European Collection of Cell Cultures, Porton Down, UK), L2 bronchial epithelial cells (ATCC, Middlesex, UK), and PT2SD cells were used to assess transduction by AAV serotypes in vitro. All cells were grown to confluence on plastic plates or tissue culture flasks (Corning, New York, NY) at 37°C in a humidified incubator saturated with a gas mixture containing 5% CO2 in air.
Determination of β-galactosidase expression levels in the lung posttransduction
Expression levels of β-galactosidase in tissue were characterized with a standardized assay kit (Galacto-Star chemiluminescent reporter gene assay system; Applied Biosystems, Bedford, MA) according to the manufacturer's instructions.
RT-PCR of IκBα
Approximately 200 mg of recovered lung tissue was homogenized in 2 ml of TRIzol reagent (Sigma-Aldrich, Arklow, Ireland), using a TissueRuptor homogenizer (Qiagen, Germantown, MD). RNA was isolated through subsequent chloroform phase separation and isopropanol precipitation. One microgram of RNA was used to generate cDNA, using a reverse transcription kit (Promega, Madison, WI). Relative levels of transgene were quantified by real-time PCR (GeneAmp PCR 9600 system; PerkinElmer), using primers directed against human IκBα and normalized relative to a glyceraldehyde-3-phosphate dehydrogenase (GAPDH) housekeeper control. Primers used were as follows: human IκBα (sense, 5′-GTCAAGGAGCTGCAGGAGAT-3′; antisense, 5′-CCATGGTCAGTGCCTTTTCT-3′) and rat GAPDH (sense, 5′-CTGGCATTGCTCTCAATGACA-3′; antisense, 5′-TTACTCCTTGGAGGCCATGTA-3′). 8
Western blot for IκBα
Approximately 200 mg of recovered lung tissue was homogenized in 2 ml of phosphate-buffered saline (PBS) containing cOmplete, Mini, EDTA-free proteinase inhibitor cocktail (Roche Diagnostics, Burgess Hill, UK), using a TissueRuptor homogenizer (Qiagen). Insoluble material was precipitated at 12,000×g for 10 min in a minifuge, and protein content was assessed by bicinchoninic acid (BCA) assay. Fifty micrograms of protein was then applied to a 4–20% gradient polyacrylamide gel, using denaturing loading buffer and boiling, and electrophoresed. Protein was then transferred to nitrocellulose in a mini Trans-Blot cell (Bio-Rad, Mexico City, Mexico) and nonspecific sites were blocked with 5% (w/v) nonfat milk in PBS. Blots were probed with rabbit anti-IκBα antibody (Cell Signaling Technology, Danvers, MA) followed by an anti-rabbit IgG–horseradish peroxidase (HRP) conjugate antibody (Sigma-Aldrich). Blots were soaked in SuperSignal chemiluminescent substrate (Thermo Fisher Scientific, Waltham, MA) and imaged. Densitometry was performed with ImageJ software and band intensity was normalized by subsequent blotting with a β-actin–HRP antibody (Sigma-Aldrich). 28
Tissue sampling and assays
After 5 days, heparin (400 IU·kg–1; CP Pharmaceuticals, Wrexham, UK) was administered intravenously, and the animals were killed by exsanguination. Immediately postmortem, the heart–lung block was dissected from the thorax and bronchoalveolar lavage (BAL) collection and analysis was performed as previously described. 24 The concentrations of interleukin (IL)-6, and cytokine-induced neutrophil chemoattractant (CINC)-1 in BAL fluid, were determined in commercially available enzyme-linked immunosorbent assays (R&D Systems Europe, Abingdon, UK). The total protein infiltrate present in BAL fluid was determined with a BCA assay kit (Pierce, Thermo Fisher Scientific, Rockford, IL).
Histologic and stereologic analysis
The left lung was isolated and fixed for morphometric examination as previously described. 29 The extent of histologic lung damage was determined by blinded assessors using quantitative stereological techniques as previously described. 30 Briefly, the vertical axis of each left lung was identified and the lung was cut perpendicular to this axis into 4-mm-thick slices with a sharp blade beginning at a position chosen by random number within the first slice. These tissue slices were then embedded in paraffin and sections (10 μm) from each slice were mounted on slides and stained with hematoxylin and eosin. One field of view from each lung section was chosen, by means of the random selection of grid co-ordinates from a fixed point at the extreme lower left tip of the lung section. Each field was examined by light microscopy (model BX51; Olympus, Mason Technologies, Dublin, Ireland) at×10 magnification, and the images were digitally captured and stored in 8-bit (256 level) format and displayed with analySISD imaging software (version 1.20; Olympus Europa, Hamburg, Germany). A 100-point counting grid was then digitally superimposed over the image and a touch count was performed for each of the following: acinar tissue, nonacinar tissue, and airspace. Acinar volume fraction represented all alveolar tissue including epithelium, endothelium, connective tissue, and inflammatory cells. Nonacinar fraction meant bronchial or vascular space, and airspace described the gaseous component of the lung.
Assessment of pulmonary epithelial gene transfer and expression
Transduced rat lung epithelia were characterized with monoclonal pan-cytokeratin antibody [C-11] (phycoerythrin [PE]) (Abcam, Cambridge, UK). Appropriate PE-conjugated immunoglobulin isotype controls were also used (Abcam). Green fluorescent protein (GFP) generated by the instilled AAV-delivered GFP IκBα plasmid construct was measured in the fluorescein isothiocyanate (FITC) channel. A single-cell suspension was obtained by collagenase digestion of rat lung homogenate. For staining, cells were first washed with fluorescence-activated cell-sorting (FACS) buffer (PBS containing 2% fetal calf serum and 0.01% NaN3 [Sigma-Aldrich]) and then stained on ice for 30 min in 1:20 antibody FACS buffer solution. Unbound antibody was removed by washing twice with FACS buffer and cells were resuspended in FACS buffer for analysis with a FACSCanto flow cytometer (BD Biosciences, Oxford, UK).
Compensation parameters were established on the cytometer, using appropriately stained single cells. Unlabeled beads (CaliBRITE unlabeled beads; BD Biosciences) were added to FACS samples for detection of absolute number of cells, and dead cells were identified with propidium iodide (PI) staining solution. Data were analyzed with FlowJo software (Tree Star, Ashland, OR) and results are presented as a percentage of cell population.
Statistical analyses
All data were checked for normality before statistical analysis. Data distribution was assessed using Kolmogorov–Smirnov tests. Data were analyzed using an unpaired Student t test and are shown as means±standard deviation. To compare differences between groups, one-way analysis of variance (ANOVA) coupled with the Bonferroni multiple comparison test was used. Lung histology was analyzed by two-way ANOVA, with group as the first factor and histologic classification (tissue, airspace, extraacinar) as the second factor. Asterisks indicate level of significance (*p<0.05; **p<0.01; ***p<0.0001). All statistics were performed with GraphPad Prism 5.0 (GraphPad Software, La Jolla, CA)
Results
Determination of optimal AAV vector serotype for transduction of pulmonary epithelium: In vitro studies
The transduction efficiencies of rAAV2, rAAV2/5, and rAAV2/6 expressing LacZ were compared in three different pulmonary epithelial cell lines: A549 cells, L2 bronchial epithelial cells, and PT2SD cells. The transduction efficiency of the three rAAV serotypes is shown (Fig. 1A) (n=10 for each group). Across all cell types tested, rAAV2/6-LacZ resulted in the highest transduction efficiency (p<0.0001). The least efficient was rAAV2/5-LacZ, which consistently had the lowest β-galactosidase activity levels across the cell types.

AAV vector aerosolization preserves structural integrity and transduction efficacy: In vitro studies
The process of aerosol generation during both ultrasonic and jet nebulization is known to create heat and shear forces, respectively, thus potentially affecting the formulation being nebulized. Using an alternative aerosol generator technology, an actively vibrating mesh nebulizer (Aeroneb® Lab; Aerogen, Galway, Ireland), the effect of two rounds of nebulization on the titer of the rAAV serotypes under investigation was determined by real-time PCR. The effect of nebulization on 1×1012 drp in 0.3 ml of sterile 0.9% saline is shown in Fig. 2A. After collection of nebulized vector, each sample was immediately subjected to a second round of nebulization, mimicking our protocol for intratracheal aerosol administration. On characterization, the titer of rAAV2-LacZ decreased by 17.66±1.44% (p<0.0001), the titer of rAAV2/5-LacZ decreased by 5.67±1.46% (p<0.01), and a significant titer reduction was also recorded for rAAV2/6-LacZ, 7.66±1.44% (p<0.0001) (n=5 for each group).
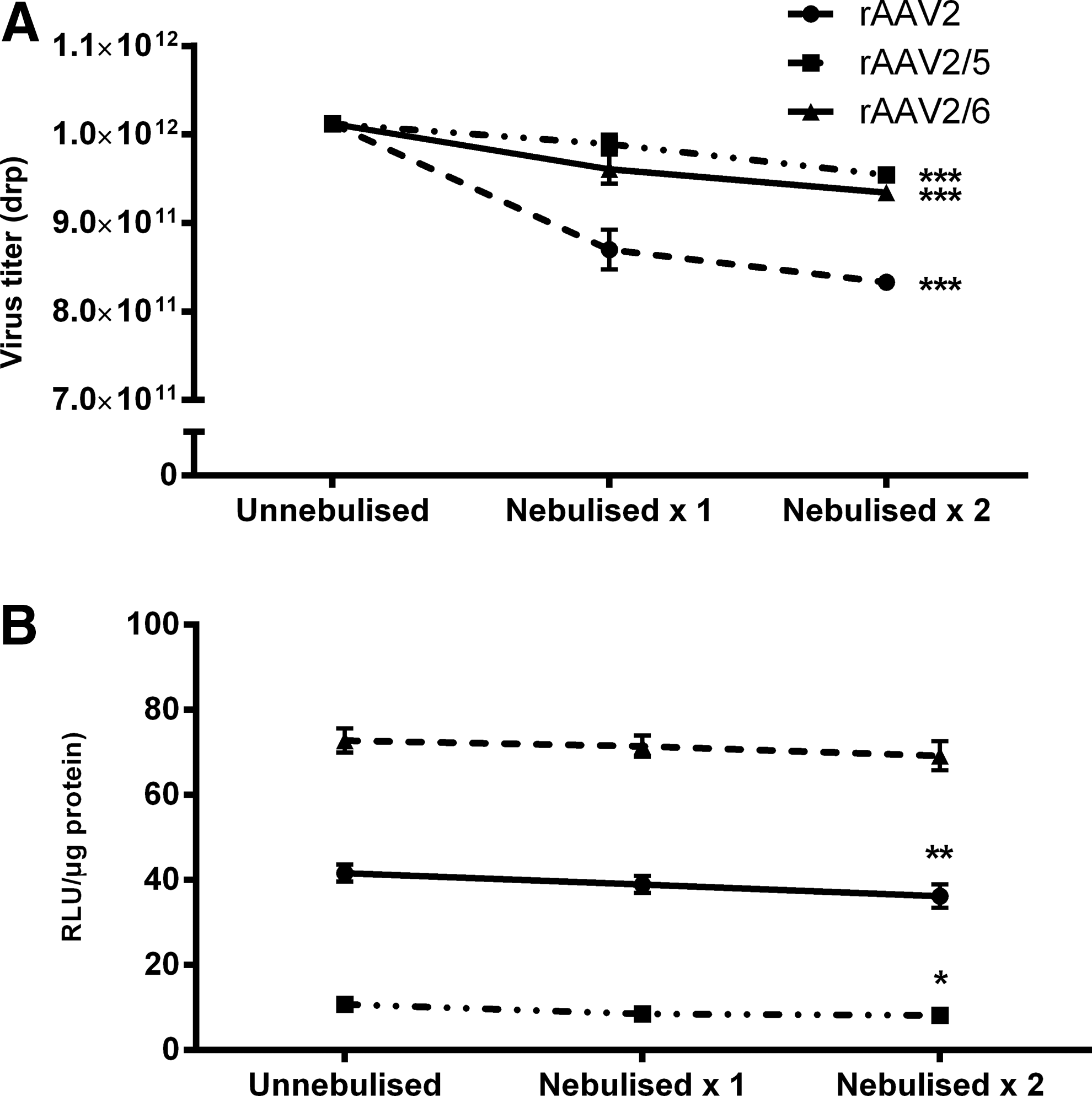
The effect of two rounds of nebulization on the in vitro transduction efficiency of the rAAV vectors is shown in Fig. 2B. Transduction efficiency was quantified by measuring the activity levels of β-galactosidase in A549 cells, after coincubation. A significant decrease in the β-galactosidase activity levels (5.47 relative light units [RLU]/μg protein) was noted for rAAV2-LacZ (p<0.01). The β-galactosidase activity levels also decreased significantly when rAAV2/5-LacZ was nebulized twice (2.58 RLU/μg protein; p<0.05) (relative light units standardized per microgram of protein). However, no reduction in transduction efficiency was seen after two rounds of nebulization of rAAV2/6-LacZ, for which a 3.59-RLU/μg protein decrease in β-galactosidase activity was recorded (n=5 for each group).
In vivo studies
Transduction efficiency of AAV serotypes
Each rAAV serotype being evaluated was delivered by aerosolization at a dose of 1×1012 drp in 0.3 ml of sterile 0.9% saline (Fig. 1B). Five days postadministration, the transduction efficiency of rAAV2/6-LacZ (4.02 RLU/μg protein) was significantly higher than that of both rAAV2-LacZ (2.17 RLU/μg; p<0.0001) and rAAV2/5-LacZ (0.66 RLU/μg; p<0.0001) (n=5 animals for each group). These results are in broad agreement with the performance recorded in the isolated Sprague Dawley primary type II epithelial cells tested in vitro.
Aerosol-delivered AAV-IκBα inhibits endotoxin-induced lung injury
rAAV2/6 vector encoding the IκBα or null gene was administered to rats via aerosolization 5 days before insult, in order to allow time for appreciable expression of the transgene. LPS or vehicle was administered intratracheally on day 5, and animals were killed 24 hr later.
IκBα was delivered and expressed in the lung
Across all treatment groups, exogenous human IκBα gene expression was analyzed by quantitative RT-PCR after LPS or vehicle administration (Fig. 3A). In the IκBα gene+LPS injury treatment group there was a 127.82±3.21-fold increase in IκBα levels compared with the sham group. This increase can be attributed to exogenous human IκBα in rat lung after rAAV2/6-mediated delivery. Three pooled samples of two animals each were analyzed in triplicate.

Homogenized lung tissue was examined for total IκBα protein levels by Western blotting followed by densitometry (Fig. 3B). As expected, LPS injury and null gene+LPS injury groups exhibited reduced levels of IκBα as part of an inflammatory NF-κB pathway response. This degradation was significantly ameliorated in the IκBα gene+LPS injury treatment group (p<0.05 vs. null gene+LPS injury group) and, indeed, protein levels were increased beyond that seen in the sham controls (p<0.05).
Analysis by FACS was performed on rat lung cotransduced with IκBα and GFP at a titer of 1×1010 drp in 0.3 ml of saline at 96 hr in order to assess the efficiency of pulmonary epithelial cell transduction by the rAAV2/6 vector employed in the current study. A PE-labeled epithelial specific antibody (pan-cytokeratin) was used to identify epithelial cells from the lung cell suspension (Fig. 4A–F), and FITC excitation (Fig. 4G) was used to calculate the percentage of cells that were transduced with GFP. Of the epithelial cells stained by the cytokeratin–PE antibody in lungs that received rAAV 2/6-IκBα, 21% were positive for GFP (Fig. 4F), whereas overall 4.3% of events were positive for GFP.
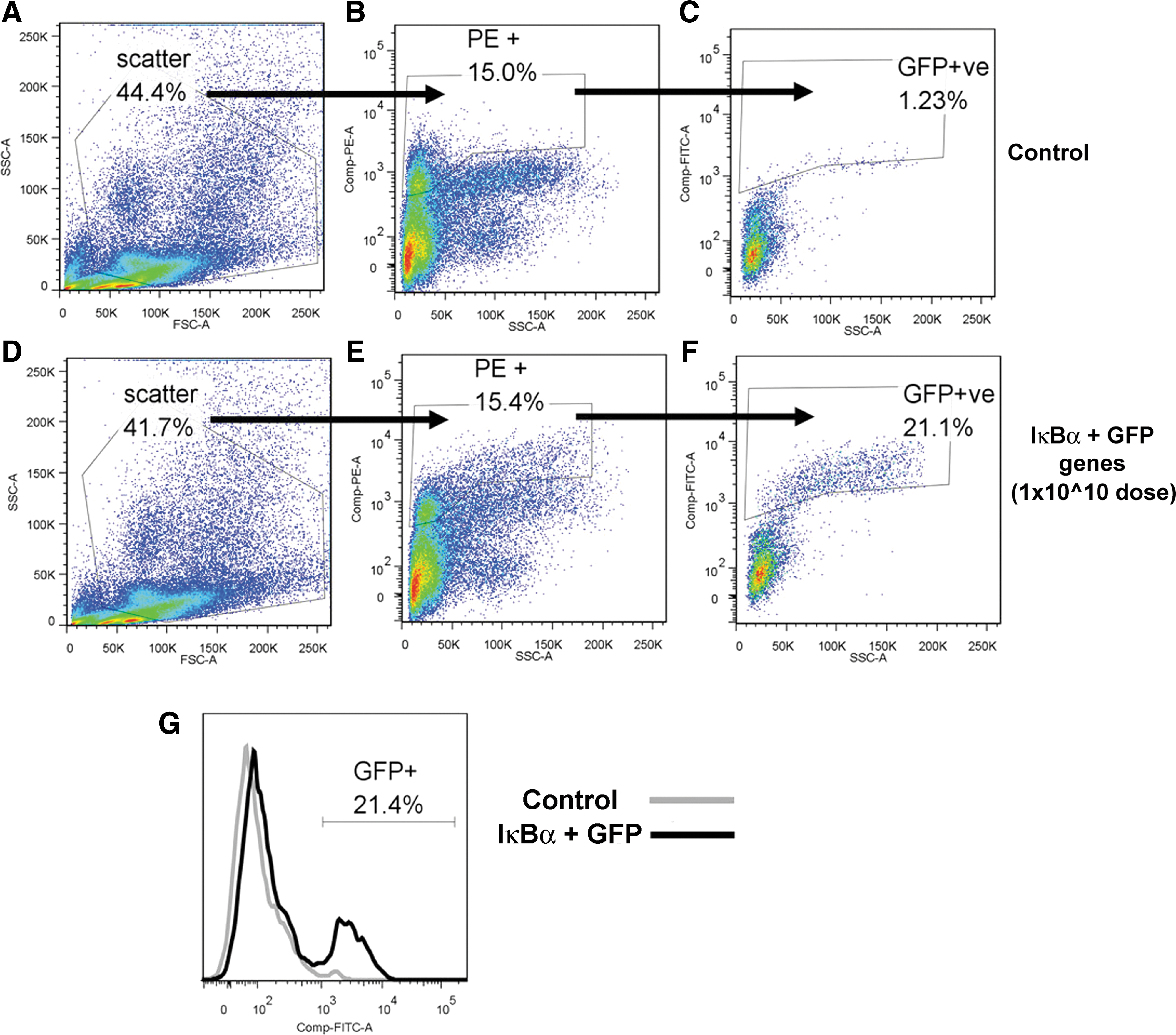
FACS analysis of rat pulmonary epithelial cells transduced with AAV expressing IκBα-GFP construct.
IκBα overexpression reduced lung inflammation
Endotoxin produced a significant inflammatory response compared with vehicle. IκBα overexpression attenuated—but did not eliminate—the pulmonary inflammatory response to endotoxin, attenuating the increase in alveolar concentrations of the key cytokines IL-6 and CINC-1 (Fig. 5A and B) and mitigating the increase in total alveolar inflammatory cell infiltration (Fig. 5C) and alveolar neutrophil infiltration (Fig. 5D). The total protein infiltrate present in BAL fluid from injured lungs also showed a significant improvement compared with injured animals (Fig. 6A).

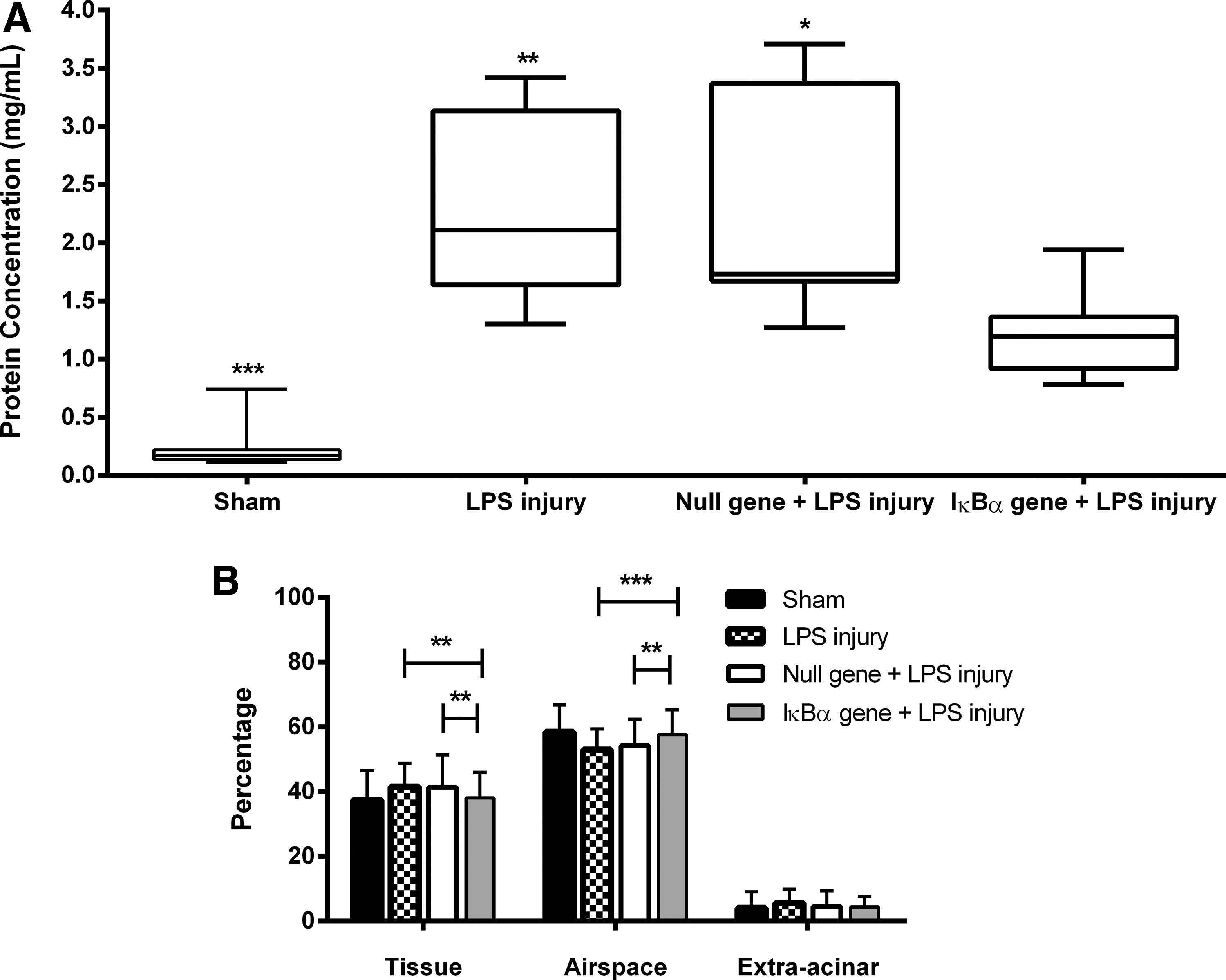
Quantitative stereologic analysis demonstrated that overexpression of IκBα significantly attenuated injury with respect to tissue density and airspace in the IκBα gene+LPS injury treatment group. IκBα overexpression maintained significant alveolar airspace fraction (LPS injury, p<0.0001; IκBα gene+LPS injury, p<0.01) and a significantly reduced alveolar tissue fraction compared with other LPS-treated groups (LPS injury, p<0.01; IκBα gene+LPS injury, p<0.01) (Fig. 6B). This demonstrates that in the case of NF-κB inhibition mediated by the IκBα gene, alteration of the alveolar wall as a result of edema and the accumulation of inflammatory infiltrates occurred to a lesser degree (Fig. 7).
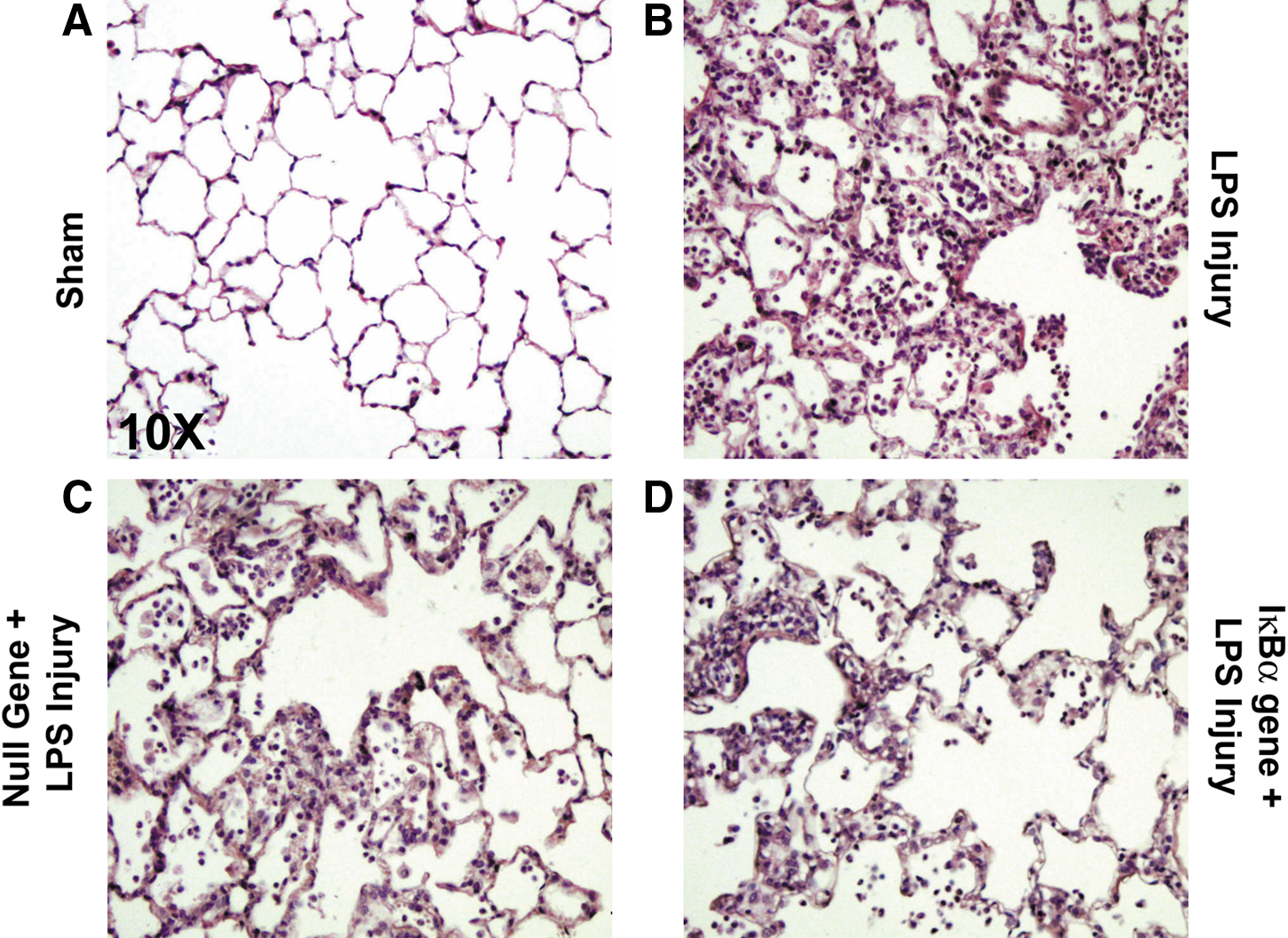
Photomicrographs (original magnification, ×10) of representative sections of hematoxylin–eosin (H&E)-stained lung tissue from each of the treatment protocols demonstrating the histologic level of ALI.
Discussion
Key findings
rAAV-mediated overexpression of the NF-κB inhibitory protein, IκBα, both in vitro and in vivo, attenuates the severity of the inflammatory response in the setting of endotoxin-mediated ALI, thus providing proof that (1) optimized aerosol delivery of the rAAV vector and the subsequent localized expression of a suitable transgene in the lung is capable of having a therapeutic effect on the markers of acute lung injury in a preclinical setting, and that (2) the effect of overexpression of a superrepressor form of IκBα can sufficiently reduce the activity of NF-κB to indirectly regulate a variety of proinflammatory cytokines. Insights into the nuclear activity and biological function of IκBα are essential for targeting the NF-κB pathway for future therapeutic approaches to inflammation and lung injury.
In vitro and in vivo serotype optimization studies
Preliminary in vitro investigations using the immortalized A549 (human) and L2 (rat) cell lines and primary isolated type II cells (PT2SD) confirmed the abilities of the tested AAV serotypes to transduce lung epithelial cells. PT2SD cells demonstrated a high correlation of transduction, as did the immortalized cell lines (Fig. 1A), with the rAAV2/6 serotype being similarly effective when targeting the Sprague Dawley rat pulmonary epithelium (Fig. 1B).
The effect of nebulization on the integrity of rAAV serotypes has not been adequately addressed previously. Although a significant reduction in titer for all three rAAV serotypes tested was observed, the transduction efficiency of the AAV2/6 vector before and after nebulization was unchanged, even after two cycles of aerosolization, suggesting the vector was not damaged by the nebulization process (Fig. 2A and B). These results are in agreement with a previously published characterization of the effect of nebulization on rAAV2 and rAAV2/5 using vibrating mesh nebulizer technology. 31 If no damage to the vector was seen in vitro, it would be expected that delivered AAV2/6 would retain its activity in vivo.
Aerosol characteristics and behavior dictate that in a small-bore diameter system, such as that used presently, droplet size is a factor of vital importance in the system's delivery performance. We have previously demonstrated that the experimental system used in the present study delivers a high percentage of the original dose to the lungs of a mechanically ventilated rat. This is achieved through the combined use of a low flow rate and low residual volume breath-actuated vibrating mesh nebulizer, generating a small droplet size appropriate for alveolar deposition. 32
Demonstration that nebulized AAV-IκBα reduces LPS injury
The presence of rAAV2/6-mediated exogenous human IκBα was confirmed by quantitative PCR. A 127.82±3.21-fold increase (p<0.0001) was noted in IκBα levels 24 hr after LPS administration (Fig. 3A). As a result of the activation of NF-κB by LPS, the amount of unbound IκBα at the local site is known to decrease as it is degraded by the 26S proteasome. This change in IκBα protein levels was confirmed by Western blot (Fig. 3B).
The efficacy of pulmonary epithelial cell transduction by the rAAV2/6 vector employed in this study was measured by flow cytometry 96 hr after instillation to calculate the percentage of lung epithelial cells that were transduced with GFP and IκBα. Of the epithelial cells stained with the cytokeratin–PE antibody and that received rAAV 2/6-IκBα, 21% were positive for GFP (Fig. 4F), whereas for the lung as a whole, 4.3% of events were positive for GFP, indicating targeted delivery to the pulmonary epithelium. Previous studies have shown the efficacy of rAAV2/6 serotype-delivered IκBα in lung injury models such as E. coli-induced injury 6 and ventilator-induced injury, 33 with both studies demonstrating attenuation of indices of injury and inflammation. The results of the present study further indicate the potential for significant levels of super-repressor transduction and hence the improved indices of injury seen here.
As evidenced by assays of the proinflammatory cytokines IL-6 and CINC-1, the increased concentration of IκBα protein was capable of binding unbound NF-κB that had been released as a result of the proinflammatory signal and so attenuated the proinflammatory cascade regulated by NF-κB (Fig. 5A and B). Proinflammatory IL-6 levels were seen to be reduced significantly in the prophylactic IκBα gene+LPS injury treatment group in comparison with the control injury groups (p<0.0001) (Fig. 5A). Previous reports of NF-κB inhibition have shown a reduction in IL-6 expression levels. 34 Although the reduction in IL-6 levels noted in the present study is quantitatively small, studies in murine sepsis and clinical ARDS have linked attenuation of IL-6 to improved survival. 35,36 The reduction in IL-6 levels correlated with that seen in CINC-1 levels (Fig. 5B).
Concomitant with the reduction in CINC-1 levels, a significantly lowered absolute number of neutrophils was recorded in the rAAV2/6-IκBα treatment group in comparison with both control injury groups (LPS injury, p<0.0001; IκBα gene+LPS injury, p<0.01) (Fig. 5D). CINC-1 acts as a functional chemoattractant for neutrophils in inflammation, and therefore reduced CINC-1 levels may provide some explanation for this reduced neutrophil count. This is consistent with previous observations. 37 The total BAL cell count was also significantly lower in the rAAV2/6-IκBα treatment group compared with control injury groups (p<0.01) (Fig. 5C). Significantly less protein was present in the BAL of rAAV2/6-IκBα-treated animals compared with both injury control groups (LPS injury, p<0.01; IκBα gene+LPS injury, p<0.05) (Fig. 6A), suggesting that overexpression of superrepressor IκBα is protective, ameliorating disruption caused to the alveolar epithelium and vascular endothelium.
Quantitative stereologic analysis demonstrated significantly less histologic injury with respect to tissue and airspace in the IκBα/LPS treatment group; and a significantly greater alveolar airspace fraction (LPS injury, p<0.001; IκBα gene+LPS injury, p<0.01) and a significantly reduced alveolar tissue fraction were recorded when compared with the LPS injury groups (LPS injury, p<0.01; IκBα gene+LPS injury, p<0.01) (Fig. 6B). The greater loss of alveolar architecture and thickening of alveolar septae in LPS-injured lungs results in impaired gas exchange, compared with the protection afforded by IκBα treatment. The closer-to-normal histology present in the IκBα gene treatment group should allow for improved gaseous exchange in the lungs and, thus, attenuate lung injury-associated hypoxemia.
Despite the reduction in proinflammatory signals seen across all experiments, as well as reduced histologic injury, the indices of proinflammatory signals for the rAAV2/6-IκBα/LPS treatment group were significantly higher than those measured in uninjured animals. LPS is a potent activator of proinflammatory signals and thus it is extremely difficult to completely abrogate its proinflammatory effect. Despite the considerable research to date in LPS-induced ALI, there are no reports of complete reversal of LPS-induced injury. While IκBα inhibits only the canonical pathway of NF-κB, a number of other mechanisms may be responsible for cytokine production.
Limitations
This study involved administration of vector 5 days before the initiation of lung injury. In the clinical setting, lung injury is generally well established at presentation. It is not clear what effect IκBα overexpression might have if introduced after lung injury or if the inflammatory process already underway would hinder IκBα transduction. However, there are certain situations, such as prior major intraabdominal or cardiovascular procedures, in which an insult, such as the development of endotoxemia, is a relatively frequent occurrence. 38 Intratracheal delivery of transgene after the development of ALI/ARDS may also be difficult, particularly in a context of atelectasis and edema, reducing the therapeutic usefulness of intrapulmonary delivered therapies. Although gene transfer in this environment has been shown to be successful, 39 –41 it is likely to be associated with reduced gene transfer efficiency. Also, the time course of ARDS is significantly longer than the 24 hr assessed here. Differences between treatment groups that were not apparent at 24 hr may be more obvious after a number of days. Further experiments with a longer treatment period might be useful in giving insights into the likely clinical impact of pulmonary IκBα therapy for ARDS. The optimal timeframe for gene expression may also be longer than 5 days, However, a study of this nature would require a different model of injury than LPS. We are unable to furnish any data relating to any potential influence of LPS on transgene expression in the current model.
As type II pneumocytes represent approximately 14% of the total alveolar population and cover approximately 5% of the total lung surface area, 42 there are limitations to their use in vitro. Finally, because of major anatomic and physiological differences between rats and humans, aerosol-based therapeutic strategies would need to be evaluated in large animal studies before translation to clinical applications.
In conclusion, we have demonstrated the effectiveness of aerosol delivery of rAAV2/6-IκBα, which inhibits the proinflammatory effects of NF-κB in the setting of endotoxin-induced ALI. There is no evidence that nebulization resulted in any degradation of viral activity. This innovative therapeutic approach provides the proof of concept for a single prophylactic delivered intervention and could serve as the basis for a potential protective strategy, based on aerosol-based vector delivery, for patients against the establishment of ARDS.
Footnotes
Acknowledgments
This work was funded by a Centre for Science Engineering and Technology (CSET) grant (03/CE2/B312) from Science Foundation Ireland and by the European Research Council (ERC-2007-StG 207777).
Author Disclosure Statement
B.D.H, J.D., D.O'T., and J.G.L. declared no competing financial interest. R.MacL. is Senior Scientist at Aerogen Ltd., and T.O'B. is a founder, director, and equity holder in Orbsen Therapeutics.