
Abstract
The idea of treating disease in humans with genetic material was conceived over two decades ago and with that a promising journey involving development and efficacy studies in cells and animals of a large number of novel therapeutic reagents unfolded. In the footsteps of this process, successful gene therapy treatment of genetic conditions in humans has shown clear signs of efficacy. Notably, significant advancements using gene supplementation and silencing strategies have been made in the field of ocular gene therapy, thereby pinpointing ocular gene therapy as one of the compelling “actors” bringing gene therapy to the clinic. Most of all, this success has been facilitated because of (1) the fact that the eye is an effortlessly accessible, exceedingly compartmentalized, and immune-privileged organ offering a unique advantage as a gene therapy target, and (2) significant progress toward efficient, sustained transduction of cells within the retina having been achieved using nonintegrating vectors based on recombinant adeno-associated virus and nonintegrating lentivirus vectors. The results from in vivo experiments and trials suggest that treatment of inherited retinal dystrophies, ocular angiogenesis, and inflammation with gene therapy can be both safe and effective. Here, the progress of ocular gene therapy is examined with special emphasis on the potential use of RNAi- and protein-based antiangiogenic gene therapy to treat exudative age-related macular degeneration.
Introduction: Retinal Angiogenesis as a Target for Gene Therapy
B
Presently, no cure exists for avascular AMD. However, several therapies are available to treat exudative AMD, including preventative vitamin supplementation, photodynamic therapy (PDT), and antiangiogenic drugs. While therapy based on vitamin supplementation shows no or little effect, current strategies have focused on treating the neovascular lesion, the hallmark of exudative AMD, to slow the progression of vision loss or even, in some cases, to improve vision. Even though the effect of PDT in slowing the progress of AMD is often temporary, the treatment can be successful for selected patients. The most promising treatment strategy being increasingly used to limit the progression of exudative AMD is based on antiangiogenic therapy. The molecular basis of exudative AMD is not well understood, but several growth factors have been implicated, including vascular endothelial growth factor A (referred to as VEGF). By targeting the abnormally high levels of this specific proangiogenic growth factor, which has been shown to be a key regulator of pathological neovascularization in the retina and the choroid, anti-VEGF therapy has markedly changed the outcome of the treatment. Nevertheless, repeated monthly treatments by means of intravitreal injections of, for example, ranibizumab (trade name Lucentis), which is the drug of preference, are necessary and will be required for a lifetime because of the short half-life of the drug in the vitreous. The requirement for these repeated intravitreal eye injections by ophthalmologists is costly and even though the treatment in most cases is safe, there is an accumulated risk of infection, glaucoma, cataract, and trauma, suggesting that new sustained treatment strategies are required for exudative AMD.
Gene Transfer to Retinal Pigment Epithelium Cells
There are several characteristics of the eye that make it an appealing target organ for gene therapy. 4 First, the eye is relatively small compared with other organs and has a sophisticated compartmentalized anatomy, providing localized transfer of small volumes of gene transfer vectors in close vicinity to the target cells. Another important attribute favoring retinal gene therapy is that cell proliferation in the adult eye is limited. This allows the use of nonintegrating gene transfer vectors, such as integration-deficient lentivirus (IDLV) 5 and recombinant adeno-associated virus (AAV), 6 a nonpathogenic human parvovirus, for persistent gene expression. Moreover, as the eye is a relatively accessible organ the risk of complications related to operational procedures is reduced. Last, but not least, the compartmentalized architecture of the eye and the excluding blood–retina barrier reduce the extent of vector distribution outside the eye. This is important in order to limit the severity of possible immune responses following delivery of immunogenic gene transfer vectors.
Pathophysiology of AMD
Presently, the exact pathophysiology of AMD is not known, but alterations of the complicated interplay between PR cells and the retinal pigment epithelium (RPE), as well as changes in the arteriosclerotic vasculature, are believed to play a pivotal role. 7 Age-related macula degeneration affects the macula, which is the part of the eye that is responsible for sight in the center of the field of vision. The early stage of AMD is asymptomatic with normal visual acuity and is characterized by the formation of large and irregular drusen composed of multicomponent, heterogeneous aggregates, and alterations in pigmentation of the RPE cells. 8 However, late AMD causes loss of central vision and is characterized by either pigment epithelium detachment with or without choroidal neovascularization (CNV) or geographic atrophy in local areas of RPE or PR cells.
CNV is characterized by choroidal capillary growth through Bruch's membrane beneath the RPE cell layer. This vascular pathology is most classically seen in exudative or “wet” AMD. Research has shown that the RPE cells are able to control angiogenesis below the retina by constitutive synthesis and secretion of both pro- and antiangiogenic growth factors, including VEGF, 9,10 and pigment epithelium-derived factor (PEDF). 11 An increase in proangiogenic factors and/or a decrease in antiangiogenic factors within the retina microenvironment may thus be critical for the development of CNV leading to AMD. Gene transfer of RPE cells by means of viral transduction has therefore attracted substantial interest, and significant progress toward sustained RPE cell transduction has been achieved.
Viral vectors for RPE-specific gene therapy
The timeline and progression of AMD differs on a person-to-person basis, and the speed in which degeneration advances can be largely attributed to how a patient takes care of his/her eyes and overall health. However, development of AMD in humans may progress over a period of years. This implies that efficient clinical applicability of antiangiogenic AMD gene therapy in the long-term most likely requires sustained and effective transduction of RPE cells. In this context, viral vectors display several attractive features, including the possibility of changing the cellular pseudotyping as well as limiting transgene expression to the target cell by implementing tissue-specific promoters 12,13 and safety elements such as miRNA target sites. 14 Several studies have shown that subretinal delivery of viral vectors, including AAV 15,16 and lentivirus vectors (LV), 17 results in both efficient and sustained transduction of RPE cells (Fig. 1). Even though the diffusion of the viral vectors from the vitreous to the outer retinal cells is limited by physical barriers, another, less invasive injection route based on intravitreal injection is now gaining ground (Fig. 1) (see below). Researchers have also investigated other approaches besides virus-mediated delivery to transfer therapeutic genes to retinal cells. Even though improvements in non-viral-mediated delivery, involving transfection and electroporation of plasmid DNA 18 as well as DNA mini-circles, 19,20 have been obtained, presently the most promising delivery systems for efficient and persistent ocular gene therapy are based on AAV- and LV-based vector technology.
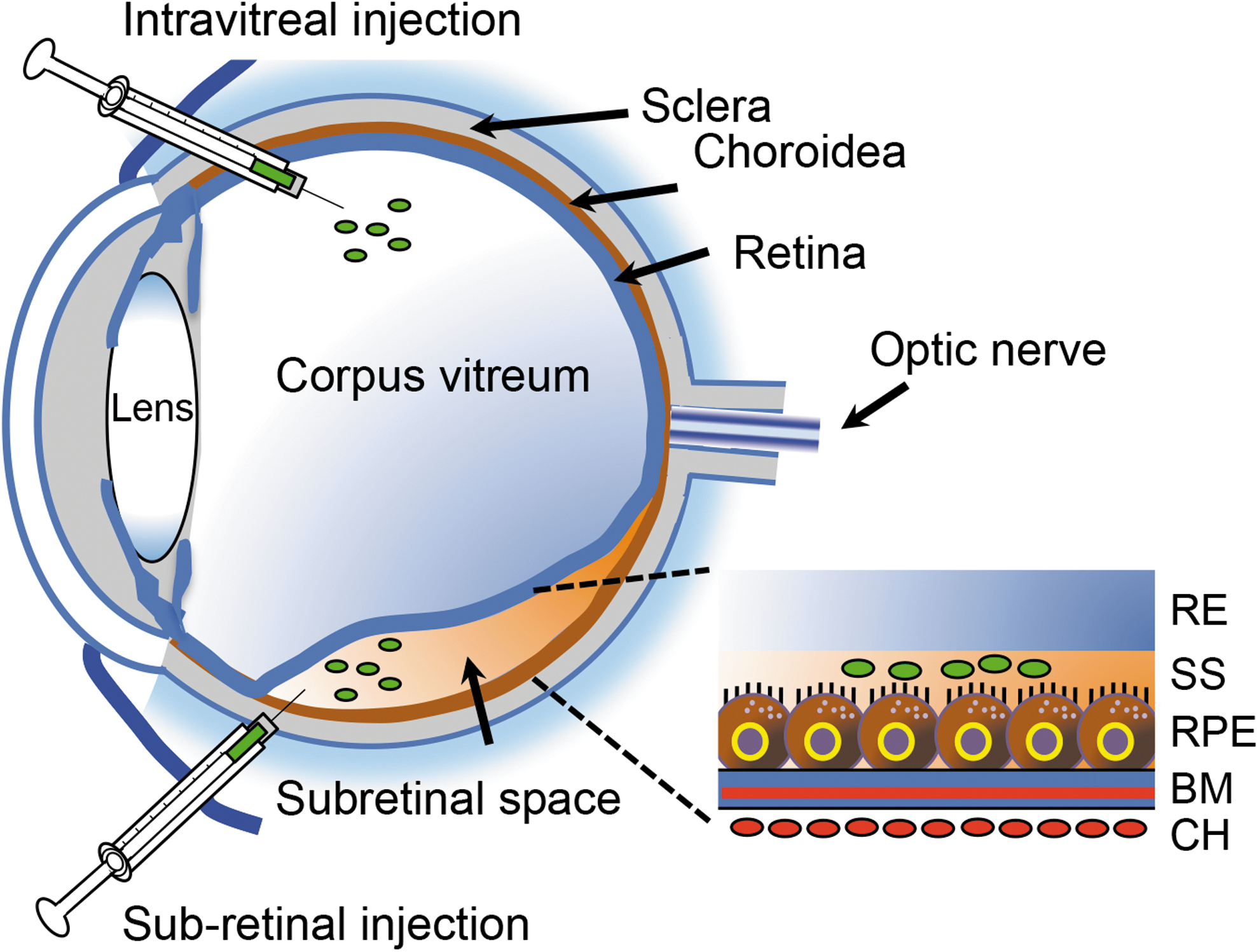
Injection routes of viral vectors used in ocular gene therapy for inhibition of CNV. Depiction of subretinal and intravitreal injection routes for the administration of viral vectors (green) used in ocular gene therapy. The route of administration and the manipulation of the capsid can be exploited to target cells of interest and obtain cell-specific gene expression. In the case of subretinal administration, a small volume of vector particles is injected in the subretinal space separating the RPE and PR cells. During this procedure, an injection-induced detachment of the retina, often referred to as “bleb formation,” is generated, thereby enabling effective administration of viral vector particles to cells in the vicinity of the injection site. RPE cells are typically targeted with high efficacy, but transduction of other cells of the outer retina is also observed. Following reabsorption of viral particles, the retina folds back. The intravitreal injection route is less invasive compared with its subretinal counterpart. Even though intravitreal injection provides transduction to cells of the outer retina, highest efficacy is obtained by subretinal administration. BM, Bruch's membrane; CH, choroid; CNV, choroidal neovascularization; PR, photoreceptor; RE, retina; RPE, retina pigment epithelium; SS, subretinal space.
AAV vectors
Delivery of AAV to the subretinal space results in sustained transduction of RPE cells, 21 implying that vectors based on AAV may provide long-term treatment of angiogenesis defects in RPE cells. Since the discovery of the pronounced applicability of AAV as a gene therapy vector, numerous recombinant AAV (rAAV) vectors have been developed. The most important landmark is the possibility of packaging viral genomes of one serotype (typically AAV2) into the capsid of another serotype. This maneuver improves gene expression kinetics and the cellular tropism. 22,23 Thus, it has been shown that AAV2 serotypes 1, 2, 4, 5, 7, 8, and 9 transduce RPE cells following subretinal delivery. 15,24 –26 At least three other important findings have emerged in the field of rAAVs: the development of (1) self-complementary (sc)AAV, (2) tyrosine variants, and (3) variants enabling transduction of RPE/retinal cells following intravitreal injection. In the former case, increased transduction efficacy is obtained because of immediate transcription of double-stranded DNA without the requirement for de novo DNA synthesis or base pairing between single-stranded vector genomes. 27 On the downside, it should, however, be mentioned that the packing capacity of scAAV vectors (∼2.4 kb) is only a half of single-stranded AAV vectors (∼4.7 kb). However, effective delivery of large genes to the retina by dual AAV vectors has recently been demonstrated. 28 In the case of the tyrosine variants, enhanced intracellular trafficking is obtained—and hence increased transduction efficacy—by the mutation of surface-exposed tyrosine residues on the capsid, thereby rendering the internalized virus particles less sensitive to proteasome-mediated degradation because of the lack of ubiquitination. 29 Regarding the latter case, progress toward diffusion of the viral vectors from the vitreous, through the inner limiting membrane, to the outer retinal cells, and hence efficient retinal transduction after delivery to the intravitreal space, has been achieved using novel capsid-mutated AAV vectors, including tyrosine variants. 30 Thus, surface-exposed tyrosine (Y) and threonine (T) residues on the capsids of AAV2, AAV5, and AAV8 were altered to phenylalanine (F) and valine (V), respectively. 31 Notably, the capsid mutant AAV2 (quad Y-F) was shown to transduce cells of both the inner and outer retina, including PR and RPE cells, following intravitreal injections in mice and canines, respectively. 30,32
Novel primate-derived hybrid recombinant AAV serotypes providing shuffled AAV capsid domains are also gaining more ground in clinical applications in the case of vector retreatment. 33 Recently, by using in vivo–directed evolution, a novel AAV vector (AAV-7m8-CAG-GFP) was shown to effectively deliver genetic material to the outer retina from the vitreous. 34 Notably, high-magnification imaging revealed the transduction of retinal ganglion cells and Müller cells, but also of amacrine cells, bipolar cells, rods, cones, and RPE, implying that AAV-7m8-CAG-GFP is capable of transducing cells far away from the injection site (Fig. 3). With further refinement, capsid-mutant AAV2 vectors show promise for retinal gene delivery through an intravitreal approach.
LV vectors
Vectors based on LV, which is a retrovirus, have been used widely in ex vivo gene therapy. Notably, there is a growing interest in using LV vectors for ocular gene therapy. This is mainly because of the LV vectors' capability to transduce nondividing cells. Further to this, the packaging capacity of LV vectors is approximately 10 kb, which is almost twice as high as that of single-stranded AAV vectors. The LV vectors integrate into the host genome with a potentially safer integration profile than other retroviruses. Hitherto, self-inactivating LV vectors derived from the human immunodeficiency virus (HIV) are the best studied. However, vectors based on non-human-derived types of lentiviruses, including equine infectious anemia virus (EIAV) and the simian immunodeficiency virus (SIV), are also being used because of their better safety profiles. In this context, SIV vectors pseudotyped with Sendai virus envelope proteins have shown effective but also swift gene transfer in the retina after subretinal exposure to the vector for only a few minutes in a mouse model. 35 To further improve safety, the risk of insertional mutagenesis can be reduced by using IDLV vectors carrying a mutation in the integrase-coding region. As a result, the integrase activity is eliminated and the viral genomes remain as episomal transcriptional templates. Even though comparable vector titers and transduction percentages, such as integration-proficient LV (IPLV) vectors, have been noted, the expression level from IDLVs is reduced by 50–90% compared with IPLVs. 36 –39 Regardless of surface proteins, subretinal injection of LV vectors primarily transduces RPE cells. 17 This is in contrast to the AAV vectors. However, in some eyes, using certain vectors (EIAV.CMV.eGFP), the transduction of other retinal cell types, including PR inner and outer segments, is evident using ubiquitous promoters driving reporter gene expression. 17 Further to this, traumatic delivery, by intentional creation of an incision into the retina during subretinal vector injection, enables gene delivery to retinal neuronal and glial cells, demonstrating the capacity of LV vectors to transduce neurosensory retinal tissue. 17 This finding may thus support the role of physical barriers in influencing vector tropism. Another important difference between the two vector systems is the fact that the LV vectors typically show inefficient targeting of outer retinal cells following injection in the intravitreal space, irrespective of species and pseudotype, because of the size of the LV vector. 17
Molecular Strategies for Antiangiogenic Therapy
The discovery of VEGF as being the major stimulating factor of subretinal neovascularization, as well as clinical findings providing a transient effect of antibody-based anti-VEGF therapy, has stimulated researchers from the gene therapy society to develop effective and sustained genetically based anti-VEGF therapeutics, overcoming the repeated monthly injection. Two major antiangiogenic strategies based on the delivery of viral vectors encoding therapeutic proteins or noncoding RNAi effector molecules for gene therapy of exudative AMD have therefore been pursued (Fig. 2). In the case of the protein-mediated approach, recent research has focused on proteins exerting antiangiogenic effects either by targeting the VEGF protein directly or by inhibiting the effects of VEGF. Prominent examples of such proteins include PEDF, soluble Flt1 (sFlt1), endostatin, and angiostatin (Fig. 2, right side). The protein PEDF is found to be one of the most potent endogenous inhibitors of angiogenesis, 40 and overexpression of this secreted glycoprotein ameliorates the phenotype in several models of ocular vascularization. 41 –43 sFlt-1 is an alternative spliced form of the FLT1 gene that retains the ligand-binding domain, but lacks the intracellular kinase and membrane-spanning domains of the full-length membrane-bound Flt-1 (mbFlt-1) protein. 44 As an endogenous inhibitor of VEGF-mediated angiogenesis, sFlt1 is believed to exert its antiangiogenic effect either by direct binding and subsequent sequestration of VEGF, or by the formation of inactive sFlt1-mbFlt1 heterodimers. 45 Endostatin and angiostatin are secreted endogenous inhibitors of angiogenesis and both have been reported to have an effect on new blood vessel formation. 46,47

The principle of RNAi- and protein-based antiangiogenic gene therapy. Delivery of viral vectors encoding small RNAi effectors or proteins inhibiting synthesis, function, or effect of VEGF. miRNA biogenesis and examples of RNAi-mediated anti-VEGF gene therapy are shown on the left side of the figure. The concept of protein-mediated antiangiogenic gene is depicted on the right side. Following transcription from either endogenous or vector-derived genes by RNA polymerase II or polymerase III, the pri-miRNA transcript enters a processor complex constituted by Drosha and DGCR8. After cleavage (depicted with a “C” shown in yellow), the resulting pre-miRNAs as well as vector-derived shRNAs (such as sh953 and miR(5,7,9) 56 ) may then bind to the exportin-5 RanGTP complex, enabling them to be exported to the cytoplasm. Then pre-miRNAs, shRNAs, and artificial siRNAs (like bevasiranib, 80 AGN211745, 82 and PF-04523655 84 ) associate with Dicer and TRBP, an RNA-binding protein. This initiates the second cleavage event leading to the generation of miRNA/miRNA duplex with an approximated length of 22 nt. In the optimal situation, the guide strand of the duplex or ectopic siRNAs is then loaded into RNA-induced silencing complex (RISC), a multiprotein complex responsible for targeted degradation of mRNA. The catalytic activity of the RISC is facilitated by, for example, Ago2, which via the important PIWI domain cleaves messenger RNA if the guide strand has perfect complementary base paring. (Right side) Antiangiogenic gene therapy based on delivery of viral vectors encoding proteins for either (1) direct interaction and subsequent inhibition of VEGF, or (2) interaction with proteins affecting the function of VEGF. Examples of proteins belonging to these two classes are sFlt173 and PEDF, 69 respectively. (Adapted from Borel et al. 55 )
As an alternative strategy to protein-based inhibition of VEGF, several studies have investigated the possibility of using RNAi-based strategies 48 –53 involving noncoding RNA hairpin molecules (RNAi effectors), including shRNAs and artificial miRNAs that target the mRNAs for proangiogenic genes (Fig. 2, left side). RNAi is a biological process in which small RNA molecules incorporated in an enzymatic complex inhibit gene expression by guided cleavage or translational repression of complementary (target) mRNA (reviewed in ref. 54 ). As illustrated in Fig. 2 the origin of double-stranded RNA molecules can be either endogenous, primary miRNAs transcribed from the genome, or, as in the case of gene therapy purposes, exogenous artificial primary miRNAs or shRNA expressed from viral or plasmid templates, or synthetic double-stranded RNA molecules delivered to the cell by nonviral approaches. Depending on the design, the DNA-based or RNA-based templates are processed by the RNAi pathway machinery comprising four central elements: (1) Drosha RNAse III endonuclease (processing of pri-miRNA to pre-miRNA); (2) exportin-5 (export receptor providing active transport of pre-miRNA from the nucleus to the cytoplasm); (3) Dicer (processing of pre-miRNA to siRNA duplexes) comprising guide (complementary to the target mRNA) and passage strands resulting in the production of approximately 22-nt-long miRNA/miRNA duplex molecules; and (4) RNA-induced silencing complex (RISC) (preferential incorporation of guide strand and subsequent identification of and binding to the target mRNA strand, followed by Ago2-mediated “slicer” activity), whereas the passenger strand is usually degraded.
To further harness RNAi effectors for clinical purposes and at the same time reduce toxicity, cell and animal studies have been performed involving (1) generation of rules for the screening of “elite” shRNAs/siRNAs by using algorithms that take into account secondary structural features of the target RNAs and the thermodynamic stability of the siRNAs' ends 55 ; (2) enhanced processing by the utilization of endogenous miRNA clusters 56 ; (3) reduction of the “off-targeting” effect by limiting the incorporation of passenger strands into RISC by Dicer-independent processing of hairpin RNAs, 57 by following rational design rules for selecting artificial RNAi effectors whose seeds have a low incidence in 3′UTRs, which is the predominant location of off-target events, 58 or by circumventing Dicer slippage 59 ; (4) reduction of toxicity by utilizing tissue-specific promoters 60 and miRNA-based safety elements for the detargeting of vector gene expression, 14,61 as well as employing low concentrations of multiple siRNAs directed against the same target 56,62 ; (5) the reduction of potential toxicity by designing Ago2-specific siRNAs/shRNAs to avoid competition with endogenous miRNAs 63 ; and (6) improvement of efficacy through simultaneous modulation of potentially functionally related genes by introducing fully complementary tough-decoy (TuD) molecules targeting a single cellular miRNA. 64,65
Preclinical studies of RNAi effectors involving expression of anti-VEGF shRNAs or miRNAs have shown significant downregulation of proangiogenic proteins (Fig. 3). 56,66 –68 Recently, the utilization of shRNAs for scAAV8-mediated anti-VEGF therapy has been described, showing potent reduction of VEGF and reduced CNV in a laser-induced mouse model. 53 Thus, from these and many other studies it has become clear that RNAi has expanded the field of viral gene therapy from gene augmentation to gene knockdown.
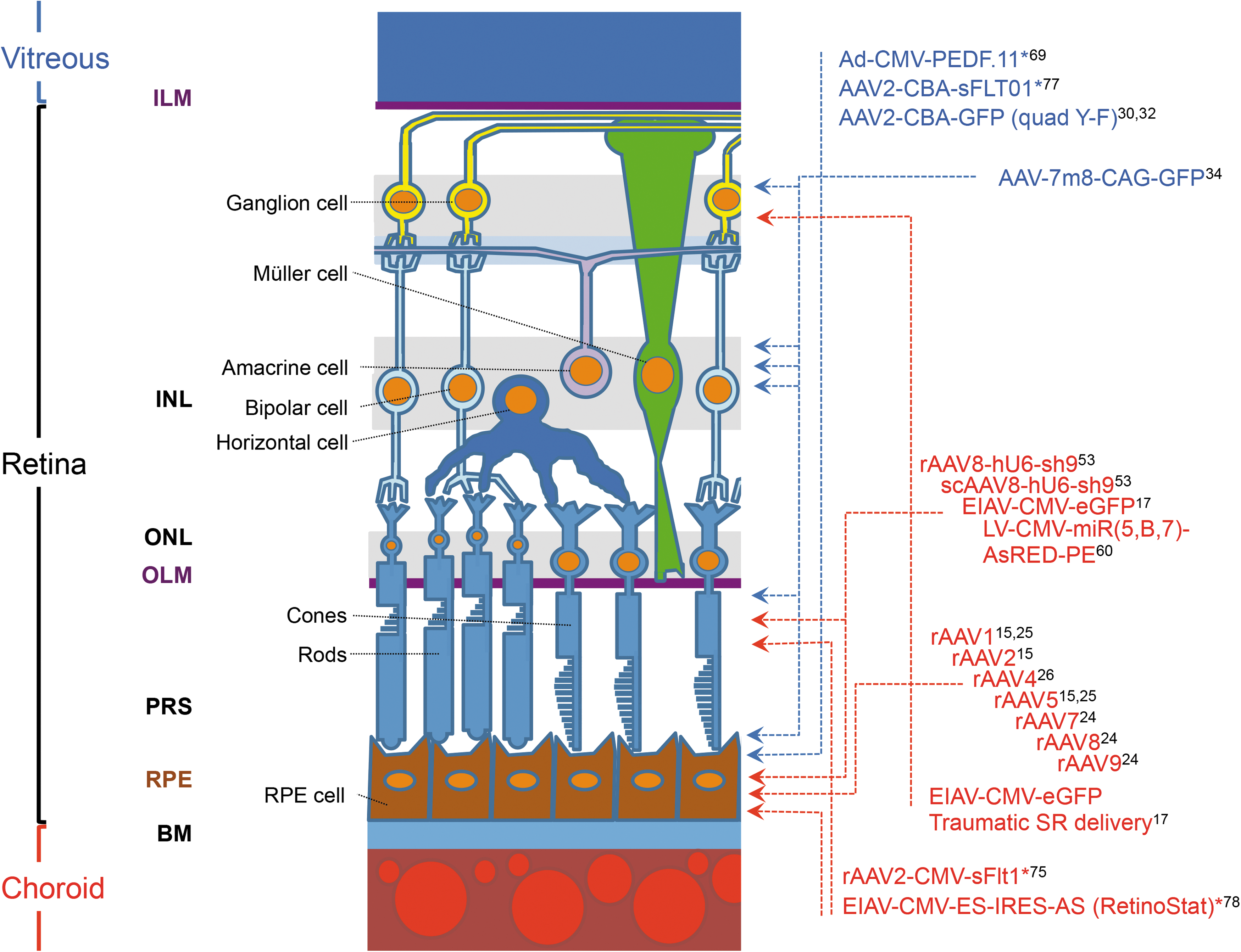
Transduction profile of AAV and LV vectors after subretinal or intravitreal injection. Schematic showing the cells in the retina compartment that are targeted by subretinal or intravitreal administration of representative AAV- and LV-based vectors. The injection route of the utilized vectors is depicted by red or blue names representing subretinal or intravitreal injection, respectively. Vectors used in clinical trials are marked by asterisk. In general, AAV- and LV-based vectors typically result in sustained gene expression restricted to the RPE monolayer after subretinal administration. Transduction of PR cells is also evident using subretinal delivery of AAVs. By using certain vectors (EIAV.CMV.eGFP) or by the creation of an incision in the retina at the time of subretinal injection, transduction of other retinal cell types—including PR cells, retinal neuronal cells, and glial cells–is possible. Interestingly, transduced RPE cells are evident after intravitreal delivery of modified rAAV capsid variants, including quad Y-F and 7m8. AAV, adeno-associated virus; BM, Bruch's membrane; ILM, inner limiting membrane; INL, inner nuclear layer; LV, lentivirus vectors; OLM, outer limiting membrane; ONL, outer nuclear layer; PRS, photoreceptor segment; rAAV, recombinant AAV; RPE, retinal pigment epithelium.
Antiangiogenic Ocular Gene Therapy
Successful antiangiogenic gene therapy in animal models has led to a clinical trial that was the first trial to use antiangiogenic therapy for neovascular eye diseases, including exudative AMD. 69 The trial was based on intravitreal injection of an adenovirus (Ad) and provided proof of concept for the delivery of genes encoding antianiogenic proteins as a gene therapy approach for the treatment of exudative AMD (see Fig. 3). 69 Twenty-eight patients with advanced neovascular AMD were recruited in a phase I clinical trial and given a single intravitreal injection of up to 109.5 particle units of Ad-CMV-PEDF.11. In contrast to previous studies, which suggested inflammation as a typical side effect in Ad vector-based gene therapy, the study showed that the vector was well tolerated with no serious adverse advents. Notably, the neovascular lesions receiving the highest vector doses showed less progression in size compared with the control group. Despite the fact that no follow-up studies were pursued, probably either because of the fear of introducing cytotoxic T lymphocyte-mediated immunological reactions or because the novel trial was unlikely to meet its primary end point, this important study has shown that expression of antiangiogenic proteins following viral delivery may prove invaluable for driving the future development of molecular therapeutics, overcoming the repeated monthly injection, and has set the direction of further clinical trials.
Recently, two other clinical trials have been initiated to evaluate the effect of the antiangiogenic protein sFlt1, a soluble VEGF antagonist. Instead of using Ad-mediated transfer, sFlt1 will be delivered by rAAV2 vectors (Fig. 3). A number of studies in animal models have documented that overexpression of sFlt1 in the eye was favorable in inhibiting CNV.
70
–75
This, together with the fact that sFlt1 is the soluble form of endogenously expressed VEGF receptor mbFlt-1, and hence should not be recognized as a foreign antigen when expressed from the AAV vector, has paved the way for an Australian phase I/II study (
In parallel, an American clinical research study will look at the safety and tolerability of a single administration of similar AAV2-based vector injected directly into the eye of 34 patients with neovascular AMD (clinicaltrials.gov, NCT01024998). This trial is mainly based on a thorough study in nonhuman primates showing that the AAV2-CBA-sFLT01 vector (Fig. 3), encoding the VEGF-binding domain of sFlt1 fused to a human IgG heavy-chain Fc domain under the control of a chicken β-actin promoter, 76 is well tolerated in general and capable of long-term expression. 77 However, in comparison to subretinal delivery, biodistribution analysis in nonhuman primates and rats shows traces of the vector in tissues other than ocular compartments. 77 Between 2 × 108 and 2 × 1010 AAV2-CBA-sFLT01 vectors, encoding the VEGF-binding domain of sFlt1 fused to a human IgG heavy-chain Fc domain under the control of a chicken β-actin promoter, 76 are intravitreally injected to explore the potential clinical applicability of this simplified administration route. The expected completion date is July 2018.
RetinoStat is a third example of protein-based inhibition of VEGF, and preclinical safety studies have delivered support for the initiation of the first LV-based clinical trial for treatment of exudative AMD (clinicaltrials.gov NCT01301443) (Fig. 3). The RetinoStat vector is based on the nonreplicating, nonhuman recombinant LV vector EIAV and is nonpathogenic in humans. 78 RetinoStat delivers two antiangiogenic genes, endostatin and angiostatin, under control of the CMV promoter (EIAV-CMV-ES-IRES-AS). 78 RetinoStat was engineered to express both angiostatic proteins because of the reported synergistic effects of angiostatin and endostatin on neovascularization. 47,79 RetinoStat is pseudotyped with the vesicular stomatitis virus G (VSV-G) protein, which facilitates entry into target RPE cells. Safety studies in rabbits and nonhuman primates showed that the vector was localized in the ocular compartment following subretinal injection, and found that the vector did not introduce structural changes in the eye or synthesis of antibodies to any of the components of RetinoStat. 78 Based on these encouraging findings, 18 patients have been enrolled into an American phase I dose-escalation study, and the estimated completion date is May 2015.
RNAi-Based Anti-VEGF Ocular Gene Therapy
As an alternative strategy to protein-based inhibition of VEGF, three clinical trials have investigated the safety, tolerability, and efficacy of RNAi-based molecules. RNAi has become a ubiquitous biological tool, and through different steps the mechanism is being harnessed for therapeutic purposes. 54 The ocular clinical trials involved the intravitreal injection of siRNA molecules, which have been chemically modified in order to increase their stability in corpus vitreum. Bevasiranib (also known as Cand5), a 21 nt RNA duplex, is designed to target the mRNA of VEGF. Phase I and II clinical trials showed promising results in terms of safety, tolerability, and efficacy (clinicaltrials.gov, NCT00722384 and NCT00259753, 80 ). Surprisingly, a phase III trial (clinicaltrials.gov, NCT00499590) involving injections of bevasiranib once every 8 or 12 weeks was terminated, because of insufficient efficacy of the siRNA monotherapy. Then followed a new phase III trial (clinicaltrials.gov, NCT00557791) designed to investigating the efficacy of the combinatory treatment of bevasiranib and Lucentis. Even though this trial was withdrawn before enrolment, because of the discovery that bevasiranib did not occur through the specific action of the siRNA targeting VEGF, but rather as a generic, sequence-independent property of the noninternalized siRNAs by the activation of the cell surface Toll-like receptor (TLR)-3, 81 the preclinical and clinical trials clearly support the safety and efficacy of bevasiranib.
AGN211745 (21-nt RNA duplex), targeting Flt1 mRNA, 82 was successfully used in a phase I clinical trial (clinicaltrials.gov, NCT00395057). However, a phase II trial was, as in the case of bevasiranib, terminated because of unspecific CNV suppression mediated by TLR-3 activation rather than a specific effect of AGN211745. To inhibit the TLR-3 activation, oligonucleotides of the PF-04523655 (clinicaltrials.gov, NCT00713518) siRNA designed to target the hypoxia-inducible RTP801 gene were methylated. 83 The preliminary data from a phase II clinical trial showed that dual-therapy together with Lucentis revealed higher efficacy compared with both Lucentis and PF-04523655 monotherapy. 84
In these trials, naked siRNA was delivered by intravitreal injections with the drawback of multiple injections, just as with conventional treatments. Another concern regarding siRNA-based therapy was the nonspecific suppression of CNV and retinal toxicity, 81,85 underscoring the importance of introducing modifications for abrogating TLR-3 activation or reducing the length of siRNAs below 21 nt, or using efficient intracellular delivery systems. Such changes should be used in order to enhance the specificity of the siRNAs and at the same time avoid retinal degeneration. Thus, to achieve synergy in a genetic intervention approach, a recent study explored the means of combining RNAi-based silencing of VEGF with delivery of the therapeutic protein PEDF. 60 Following the engineering of multigenic LV vectors, the potential of combining multiple miRNAs with protein expression for combined antiangiogenic activity was demonstrated using an RNApolII-driven expression cassette. This configuration allows effective production of intron-embedded miRNAs that are released upon transduction of target cells. The LV vector showed efficient knockdown of VEGF in vitro and provided in vivo evidence for localized transgene expression following the incorporation of the RPE-specific vitelliform macular dystrophy 2 promoter into the vector for elimination of expression in ganglion cells and thus the transport of gene products to the brain (Fig. 3). These results may thus have potential implications for the future development of gene delivery vehicles for combination therapy of neovascular diseases, including exudative AMD.
The identification of cellular miRNA sponges 86 and miRNA target sequences 87 regulating the endogenous level of miRNAs, and subsequently the expression of genes downregulated by miRNAs, may pave the way for introducing new approaches for targeted expression and reduction of angiogenic gene products. For instance, expression of miRNA-10 has been shown to reduce the level of sFlt1. 88 This allows VEGF to bind to the otherwise less preferable mFlt1, suggesting expression of ectopic miRNA-10 target sequence molecules as a novel tool to reduce CNV. For efficient elimination of a specific miRNA, or for simultaneous targeting of several different miRNAs, the target sequences could be expressed either as clustered tough decoys 89 –91 or as circular miRNA sponges. 86 These examples of using shRNA and miRNA target sequences expressed from a viral vector may also contribute to the development of effective anti-VEGF therapy.
Concluding Remarks and Future Considerations
During the last two decades, research findings in the field of viral ocular gene therapy have been a significant propellant in moving genetically based therapy from bench to bedside. 17,92,93 In this context, viral vector-based gene delivery has proved its worth as an extremely versatile platform, and vector improvements for viral gene therapy aim at designing vectors that can safely provide targeted delivery to a high number of cells with a low amount of viral vectors. As it stands, viral vectors based on LV and AAV are the most promising delivery systems for efficient and persistent ocular gene therapy. However, it is important to stress that the present viral vector collection is neither complete nor perfect, but the introduction of scAAV and optimization of dual AAV vectors as well as novel capsid-mutated AAV vectors clearly demonstrates what vector manipulation can accomplish in terms of enhancing safety, efficacy, intracellular trafficking, and diffusion, while at the same time reducing the therapeutic viral load. However, this advancement has also presented new challenges that need to be addressed in order to further increase the applicability of antiangiogenic gene therapy, including long-time efficacy, safe delivery of viral vectors to the outer retina, development of vectors providing effective delivery to the retina following intravitreal injection, and engineering of vectors enabling combination therapy.
Viral antiangiogenic gene therapy based on protein or shRNA/miRNA expression for treatment of exudative AMD shows efficacy both in model systems and in patients. Interestingly, clinical studies have suggested dual therapy based on siRNA and Lucentis. Similarly, preclinical studies have indicated potential implications for the future development of gene delivery vehicles for a tissue-specific combination therapy of neovascular diseases utilizing LV vectors as a platform for delivery of RNAi effectors and antiangiogenic proteins encoded from an RNApolII-driven expression cassette. 60 Implementation of safety elements based on miRNA target sites is another strategy to inhibit transgene expression in nontarget cells. 14,94 Recent studies have also implicated targeting or overexpression of specific miRNAs as important options to prevent pathological retinal angiogenesis. Overexpression of miR-126 mimics repressed VEGF expression in the RPE cells, 95 whereas targeting of miR-30 inhibits neovascularization by enhancing CCL2-dependent macrophage recruitment, suggesting that anti-miR30a-based therapy may be applicable for treating human neovascular eye diseases by targeting non-VEGF-driven pathways. 96
Targeted genome editing by LV protein transduction should also be considered as a possibility of reducing the VEGF level in RPE cells. CRISPR/Cas systems for genome editing have revolutionized biological research over the past 3 years, and their ability to make targeted changes in DNA sequences in living cells is now being applied to clinical medicine. 97,98 In parallel, it has been shown that LV protein transduction effectively facilitates codelivery of site-directed nucleases, like zinc-finger and TAL-effector nucleases and donor RNA. 99 Thus, as a novel approach to reduce the VEGF level in RPE cells copackaging of CRISPR/Cas proteins with guide strand in LV particles might lead to targeted and safe elimination of one of the VEGF alleles.
In light of the immense advancements in the high-throughput genotyping and the analysis of the human genome, new targets for mono or combinatory antiangiogenic gene therapy will appear. Recently, an association study provided new evidence to support ABCG1 as a susceptibility gene for exudative AMD. 100 Similar, in-depth fine-mapping of 34 loci associated with AMD has led to the identification of 14 novel disease-associated haplotypes, 101 and a whole exome sequencing study has identified several variants in novel candidate genes that may contribute to AMD. 102 Notably, the latter analysis further establishes the extracellular matrix pathway as an important component of AMD pathogenesis.
Oxidative stress and inflammation are interrelated biological events and both have been identified to play significant roles in the pathogenesis of AMD. 103 Recent studies have therefore focused on to investigate anti-inflammation effects. For instance, studies involving downregulation of the gene encoding the p22phox enzyme, which produces reactive oxygen species, show anti-inflammatory effect in ARPE-19 cells by regulating MAPK and NF-κB pathways. 104 These studies support the notions that p22phox plays an important role in inflammation accompanied by oxidative stress and imply that p22phox may provide a novel therapeutic target for oxidative induced inflammation.
An important issue in respect to using RNAi effectors is toxicity and how to obtain high specificity and a low off-target effect. It seems that toxicity primarily depends on the stem–loop design and the dose. 54 Hitherto, the design of “elite” shRNAs/siRNAs has attracted much attention and promising preclinical results have been obtained, for example, by Dicer-independent processing of hairpin RNAs. 57 Incorporation of RNA aptamers into, for example, the loop of shRNAs or artificial miRNAs is another interesting approach that might have an impact on future RNAi-based therapy. Recent research has shown that RNA aptamers can control the expression of RNAi effectors 105,106 as well as modify the miRNA pathway. 107 RNA aptamers may thus be a valuable tool for restricting shRNA expression and modulation of miRNA levels. However, further research in this field is needed in order to obtain clinical-relevant data. In addition, as geographic atrophy is an important factor in AMD, especially in the dry form, which cannot be treated, additional approaches besides antineovascular-based therapy are required for treatment of AMD. Novel studies show that interleukin 33/ST2 signaling regulates inflammatory response in choroidal stroma, suggesting that possible novel targets may include components of the IL-33/ST2 axis. 108 In addition, another recent study has demonstrated that intrevitreal injection of a P2XR7 antagonist reduces subretinal monuclear phagocyte toxicity and suggest that P2XR7 antagonist treatment might help at preventing PR cell loss in dry AMD. 109
Host immune response is of utmost importance because an inflammatory response may reduce the efficacy of the therapy as well as introduce irreversible damage in the tissue. However, in the context of ocular gene therapy, several studies have shown that immune responses in the eye are far fewer compared with what has been found after systemic injection of AAVs. 110 –113 Notably, a recent clinical study found the subretinal injection method to be safe 1 year after readministration. 114 Contrary to this finding, the production of neutralizing antibodies is stimulated following intravitreal injection in animal models. 115 However, preliminary safety and tolerability results of intravitreal administration of an rAAV2 vector encoding human wild-type mitochondrial NADH dehydrogenase 4 gene in patients with Leber hereditary optic neuropathy overall showed that safety and tolerability of a single intravitreal injection was good in all cohorts. 116 These studies allow for dose escalation necessary to identify the highest tolerated dose that will be used in upcoming studies (clinicaltrials.gov, NCT02064569) of clinical efficacy of the vector, and may thus pave the way of using the intravitreal injection route in treatment of other ocular diseases. Another important challenge is to reduce the risk of genotoxicity, and different approaches using more efficient vectors to avoid multiple integrations in a single cell, or expression cassettes that avoid long-distance enhancer interactions or incorporate insulator sequences, as well as optimized IDLVs, have been suggested.
As another approach to treat exudative AMD, targeted, transposon-based ex vivo gene therapy is currently being investigated. 117 Genetically modified RPE and iris pigment epithelium (IPE) cells to produce elevated levels of PEDF will be transplanted to the subretinal space of patients with exudative AMD to inhibit CNV by augmented PEDF secretion. An advantage of this method is that it is nonviral. However, even though RPE and IPE cells have shown potential following transplantation, the robustness and efficacy of this approach still remains unclear. To circumvent intravitreal and subretinal injections, a recent study has investigated the effects of the multikinase inhibitor regorafenib following noninvasive administration on ocular neovascularization. 118 Notably, regorafenib shows efficacy in two different rodent models of ocular neovascularization and a phase II trial is currently recruiting participant (NCT02222207). As an eye drop formulation would be noninvasive and self-administered by patients, this approach seems highly attractive. Alternatively, a recent study has shown that a single ocular administration of a biodegradable microsphere delivery system releasing a highly potent anti-VEGF domain antibody-based molecule from a intravitreal depot facilitates effective ocular drug levels and protection against laser CNV in nonhuman primates. 119
In conclusion, these highly encouraging data prove that viral-mediated antiangiogenic ocular gene therapy as such is beyond the proof-of-concept threshold and will most likely, in combination with the promising findings from non-viral-based experiments and the growing support by substantial investments by the pharmaceutical industry, play an important role in the development of efficacious long-term antiangiogenic gene therapy treatment of ocular diseases, including AMD and proliferative diabetic retinopathy.
Footnotes
Acknowledgments
The author would like to thank Aarhus University, Denmark, for funding the research. In addition, this work was supported by Gene Therapy Initiative Aarhus (GTI-Aarhus) funded by the Lundbeck Foundation (Grant No. R126-2012-12456) and The Danish Eye Foundation.
Author Disclosure
No competing financial interests exist.