
Abstract
Neurotrophic factors (NTFs) hold great potential as therapeutic agents in the treatment of neurodegenerative conditions, including Parkinson's disease (PD), in which the progressive loss of dopamine neurons in the substantia nigra pars compacta causes severe motor symptoms. There is extensive evidence that in preclinical animal models of PD NTFs are both neuroprotective and neurorestorative. In particular, glial cell line-derived neurotrophic factor (GDNF), neurturin (NRTN), cerebral dopamine neurotrophic factor, and mesencephalic astrocyte-derived neurotrophic factor have shown great potential to restore dopamine neurocircuitry. Although some previous clinical trials have demonstrated limited efficacy of GDNF and NRTN, there are several concerns raised with these studies. Moreover, open-label studies with GDNF as well as a study with NRTN showed clinical improvement, particularly in patients with early-stage PD. Indeed, as previous clinical trials with NTFs were associated with several technical problems, there is a great need for further investigations. In this review we discuss the emerging and existing possibilities to use NTFs as neurorestorative agents and the ways to improve their efficacy, and compare gene therapy and recombinant protein therapy approaches for restoring the dopamine circuitry in PD.
Introduction: Concept of Restoring Dopamine Phenotype
P
Why dopamine neurons in the SNpc start to die is currently unknown. The selective vulnerability of SNpc dopamine neurons in familial forms of PD has been linked to mutations of specific genes (such as SNCA, Parkin/PARK2, UCHL1, PINK1, DJ1/PARK7, LRRK2, and several others; see updated list at the OMIM website 7 ) by genome-wide association studies (GWASs). 8,9 Moreover, various cellular and molecular mechanisms such as mitochondrial dysfunction, endoplasmic reticulum (ER) stress and the unfolded protein response (UPR), decreased support of neurotrophic factors (NTFs) in maintaining dopamine neurons, increased oxidative stress, neuroinflammation, as well as impairment of autophagy have been suggested to play a role in dopamine neurodegeneration. 10 –13 Because there is no single cause for the neurodegeneration of dopamine neurons it is likely that the selective vulnerability of SNpc neurons is caused by their physiological properties. It has been shown that the use of L-type Ca2+ channels and ATP-sensitive K+ channels for their pacemaking activity, which is specific for SNpc dopamine neurons, is a likely cause of increased cellular stress. 14 –18 Despite many years of intensive research, efficient treatments that could slow down or stop degeneration of dopamine neurons are currently not available for PD. Therefore, one of the top priorities in PD studies is the search for molecules with potential clinical application in protecting and repairing the functions of adult dopamine neurons in vivo. 19
The neurorestoration concept is based on both clinical and animal studies revealing that the SNpc dopamine neurons start to die in a retrograde manner. The “dying back” degeneration of nigrostriatal dopamine neurons starts from the caudate putamen (also known as the striatum) and progresses toward the cell bodies in the SNpc.
20
–22
This has been shown also in the MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) mouse model of PD, where systemic injections produce relatively more robust deficits in the striatum than in the midbrain.
23,24
A similar phenomenon has also been observed in genetic models of PD.
25
–28
Although most of the genetic animal models of PD based on GWASs
29
show none or only mild deficits in dopamine phenotypic markers, the study by Wade-Martins and colleagues demonstrated that transgenic mice carrying the human wild-type α-synuclein locus and expressing a high level of α-synuclein exhibit a parkinsonian phenotype of age-dependent motor impairments and loss of dopamine neurons.
30
Many of the genes identified in GWASs as risk factors for PD, such as Parkin, Pink-1, DJ-1, and LRRK2, are associated with mitochondrial function,
31
and a genetic deletion of the mitochondrial transcription factor A gene (Tfam, which is not mutated in patients with PD) in MitoPark mice causes clear motor and dopamine deficiency.
28
In both α-synuclein-overexpressing and in MitoPark mice, the deficiency of dopamine neuronal markers is more robust in the striatum than in the midbrain. Similarly, the clinical symptoms in patients with PD appear when striatal DOPA (
Is the way nigrostriatal dopamine neurons die something specific for this population of neurons or a general phenomenon? Studies of axotomized cholinergic neurons show that the dying neurons first lose their phenotype, but retain cytoarchitectural structure after their phenotypic markers are gone. 34,35 Similarly, in dying nigrostriatal dopamine neurons labeled with green fluorescent protein (GFP) the GFP fluorescence is still detected after the phenotypic markers are already lost. 36 This phenomenon of losing phenotype before actual neuronal death gives a possibility to apply neurorestoration therapy aiming to restore the dopamine phenotype while the neurons are still viable. In addition, there is certainly a population of dopamine neurons that have lost their synaptic contacts and are carrying degenerated axons. These neurons should also be possible targets for neurorestorative disease-modifying therapy. The longer PD progresses, the smaller the number of dopamine neurons to which phenotype can be restored. NTFs have other effects as well, such as the ability to induce sprouting of dopamine terminals, upregulate tyrosine hydroxylase and enhance dopamine synthesis, stimulate synaptic transmission, and so on, that all can take part in neurorestoration. However, it is likely that the more advanced the patient's disease is, the smaller the treatment effect will be.
Besides neurorestoration therapy with NTFs there are other therapeutic approaches for PD. The symptomatic treatment approaches have been successful and resulted in dopamine replacement drug treatments 37 –39 that are successfully alleviating the symptoms in patients with PD, but not preventing the disease progression. Another novel restorative approach is to differentiate dopamine neurons from patient-specific induced pluripotent stem cells and then transplant these cells into the striatum, and studies in rats demonstrate the feasibility of such therapy. 40 However, the striatal transplantation of grafts from embryonic midbrains into patients is a surgical procedure with a high risk of complications. Although transient clinical benefits were observed, the grafted neurons die over time. 41 –43 Also, several transcription factors have been found to be important for neuroblasts gaining dopaminergic fate during development. Expression of transcription factors, such as those encoded by Mash1 and Nurr1, could also be one of the treatment approaches to stimulate dopamine neurogenesis in adult neuronal stem cells. 44 Finally, small-molecule NTF mimetics and small molecules enhancing synthesis of endogenous NTFs are currently under development. 45,46 Thus, the current therapeutic strategies can be broadly divided into three categories: the treatment of PD symptoms with dopamine replacement drugs, neuron replacement therapies using grafts of embryonic midbrain cells or differentiated stem cells, and neurorestoration approaches using NTF or NTF mimetics. In contrast to symptomatic treatments, both neuron replacement and neurorestoration with NTFs can potentially offer a cure for PD. However, the NTF-based neurorestorative therapy may be more feasible, as in addition to creating novel neuronal connections it aims to preserve and enhance the existing nigrostriatal network in the adult brain.
GDNF and CDNF/MANF Families of Neurotrophic Factors: Effects of Recombinant Proteins
In this review we focus on two NTF families: (1) the glial cell line-derived neurotrophic factor (GDNF) family, including its family member neurturin (NRTN). GDNF and NRTN are the two most studied NTFs in animal models of PD, and they have also been tested in clinical trials 47 ; and (2) the CDNF/MANF family, consisting of cerebral dopamine neurotrophic factor (CDNF) 48 and its homolog mesencephalic astrocyte-derived neurotrophic factor (MANF). 49,50
Classically, NTFs are small secretory proteins regulating neuronal number, neuronal survival, neurite branching, synaptic plasticity, and neuronal phenotype by binding and activating their cognate receptors. The GDNF family is distantly related to the transforming growth factor-β superfamily. 51 GDNF family ligands (GFLs) signal as homodimers through transmembrane receptor tyrosine kinase (RET) by first binding to specific GDNF family receptor α (GFRα). In addition, alternative GFL receptors have been identified such as neural cell adhesion molecule (NCAM) and syndecan-3. 52,53 GDNF was first purified from rat glioma cell line culture medium, and recombinant protein added to the cell culture medium was found to promote survival of embryonic dopamine neurons, 54 whereas NRTN was originally identified through its ability to promote the survival of cultured sympathetic neurons. 55 Thereafter, in animal models of PD, intrastriatally and intranigrally administered recombinant GDNF protein has been shown both to protect and restore the nigrostriatal dopamine system from toxin-induced neurodegeneration. 48,56 –59 Similarly, recombinant NRTN protein has shown neuroprotective and neurorestorative efficacy in animal models of PD. 60,61
CDNF and MANF comprise a new family of proteins that have also been shown to protect dopamine neurons from degeneration in animal models of PD, but more importantly they restore dopamine circuitry after MPTP or 6-hydroxydopamine (6-OHDA) treatment when injected into the striatum as recombinant proteins. 23,48,59 CDNF and MANF have drastically different in vitro neuroprotective mechanisms compared with GDNF and NRTN (Table 1). Overexpressed MANF protects cultured mouse sympathetic neurons from apoptosis; however, MANF and CDNF proteins added to the culture medium do not elicit any protection, 48,62 suggesting that sympathetic neurons lack plasma membrane receptors for MANF and CDNF. In contrast to GDNF, radioactively labeled MANF and CDNF did not bind to or were not taken up by cultured sympathetic neurons 62 (and M. Saarma, personal communication).
Comparison of properties of glial cell line-derived neurotrophic factor, neurturin, cerebral dopamine neurotrophic factor, and mesencephalic astrocyte-derived neurotrophic factor
aa, amino acid; CDNF, cerebral dopamine neurotrophic factor; GDNF, glial cell line-derived neurotrophic factor; MANF, mesencephalic astrocyte-derived neurotrophic factor; NRTN, neurturin; pI, isoelectric point.
MANF and CDNF are ER-resident proteins and secreted on ER stress in a calcium-dependent manner. 48,63,64 The loss of MANF leads to activation of the UPR and ER stress response pathways in knockout mice. 65 CDNF and MANF inhibit cell death, regulate ER stress and the UPR, and can protect neurons from ER stress 66 ; however, the precise molecular mechanism of their action is not yet clearly defined, although it has been suggested that MANF interacts with KDEL receptors 64 and GRP78 (glucose-regulated protein 78 kDa). 63
Although both GDNF- and CDNF/MANF family proteins have been shown to have effects in animal models of PD there are also limitations in their therapeutic use. In general, recombinant proteins have been successful drugs for several diseases. 67 However, there are several challenges when using proteins as therapeutic drugs for PD or other diseases of the brain. Proteins do not pass the blood–brain barrier (BBB) and therefore should be injected directly into the brain parenchyma. The production of recombinant proteins can be expensive and thus the costs of treatment for a single patient can be high. The administration of proteins into the brain requires stereotaxic surgery and delivery techniques are not trivial. The diffusion of GDNF, and particularly NRTN, in the brain is restricted because of their binding to heparin sulfate proteoglycans in the extracellular matrix (Table 1). 52,68,69 Also, the solubility of recombinant proteins can be problematic, and that is indeed the case with wild-type NRTN (Table 1). 70 The production method can also be important for the activity and solubility of the protein. For example, GDNF produced in Escherichia coli is nonglycosylated whereas GDNF produced in mammalian cells is glycosylated and more stable. 71 The advantages of protein delivery are that protein receptor-mediated signaling is limited by its half-life, and the administration techniques via pumps can be intermittent and easily terminated. One of the suggested problems in the failed phase 2 GDNF clinical trial was the limited diffusion of GDNF from the injection catheters into the brain parenchyma. 72,73 This could at least partly explain the controversy concerning why a phase 2 clinical trial was unsuccessful whereas the small open-label studies with a different delivery catheter system were successful. 74 –76
Viral Vectors
Viral vectors, especially recombinant helper-free adeno-associated viruses (AAVs), provide a safe, fast, and robust way to efficiently transduce various neuronal populations. 77,78 Importantly, AAVs have already been used as efficient vectors for gene therapy in patients with PD without serious adverse effects. 79,80 For neurodegenerative diseases AAV is at the forefront of therapeutic approaches for gene delivery. 81
Depending on the gene of interest, several features of the viral vector need to be considered. Whether the protein mediates its action on dopamine neurons in an extrinsic or intrinsic manner can influence which serotype or promoter to choose. Our experience is that nigrostriatal dopamine neurons are among the most difficult to target both anatomically and technically. To target dopamine neurons in the SNpc one also needs to keep in mind that these neurons are relatively more sensitive to stress caused by accumulation of misfolded and aggregated proteins. Thus, a promoter driving excessively high expression of a transgene can lead to protein accumulation, activation of UPR pathways, ER stress, and neurodegeneration. To selectively target dopamine neurons, careful choice of viral serotype and promoter is of great importance. The previous 81 and current clinical trials (Table 2) have used AAV2 as a vector for gene therapy, whereas studies indicate that AAV5 is more effective in transducing mouse and primate neurons 82,83 ; thus, choosing nonoptimal vector might have contributed to the failure of some initial clinical trials. Immunity to wild-type AAV is another issue complicating gene therapy development. First, it should be noted that the immune response to AAV injected into the human brain has been poorly studied, and it could be different from the response to AAV injected into the blood. For example, the brain does not reject cell grafts in a similar manner to other organs in the body, 84 and indeed, no inflammatory or immune reactions either to AAV capsid or to the transgene were observed in the brains of monkeys and rats after intracranial injections of AAV2. 79 Another important issue is that AAV has more than 100 known serotypes that can be used to avoid immune responses to repeated AAV injections. 81 Most humans naturally develop anti-AAV immunity during the first years of life, and the development of immunity has also been shown to depend on the time of AAV administration: The immune response is greater if the patient has been given AAV once already. In addition, neutralizing antibodies against AAV5 are less prevalent compared with AAV2, 85 and thus using AAV5 may elicit a lower immune response. However, AAV capsids are highly conserved and therefore the neutralizing anti-AAV antibodies cross-react with a wide range of AAV serotypes. 86 Several strategies have been tested to overcome immunity to AAV, such as using empty viral capsids to adsorb anti-AAV antibodies, injecting higher AAV doses, or using balloon catheters followed by saline flushing. 86 Careful consideration is appropriate when deciding whether viral vector is to be injected into the substantia nigra or striatum: Studies involving GDNF gene therapy approaches in the rat 6-OHDA model of PD have demonstrated that striatal delivery results in a better neuroprotective and neurorestorative effect than by delivery above the SNpc. 87 –89 In addition to nigral delivery being less effective, stereotaxic injections above the SNpc in humans are also more challenging. When targeting the deeper brain regions the risk for damaging the vasculature is greater. However, in the latest AAV2-NRTN clinical trial AAV was administered both to the striatum and substantia nigra. 79 This was done mainly because NRTN is known to bind tightly to heparin sulfate proteoglycans, which restricts its diffusion in the brain parenchyma.
Current clinical trials of neurotrophic factors using proteins or viral vectors for Parkinson's disease
AAV, adeno-associated virus; GDNF, glial cell line-derived neurotrophic factor; r-metHuGDNF, recombinant methionyl human GDNF.
Similarly, the choice of promoter driving NTF expression is important, and AAV5 vectors using the human synapsin1 (hSYN) promoter for neuron-specific expression 90 have been shown to most efficiently target GFP expression to neurons in the rat striatum. 91 Our own results in transducing primary mouse embryonic midbrain cultures with lentiviral vector using three different promoters (CMV, elongation factor [EF]-1α, or hSYN) clearly show that the hSYN promoter is the most specific in targeting embryonic dopamine neurons in primary mouse midbrain cultures (Fig. 1a–c). Lentiviral vector with the CMV promoter targeted only a few dopamine neurons. We observed more GFP-positive dopamine neurons with EF-1α than with CMV. However, with the EF-1α promoter the levels of GFP expression in targeted neurons were clearly lower compared with hSYN. We obtained similar results from in vivo experiments with the same lentiviral preparations after stereotaxically injecting them just above the substantia nigra of adult rats. All three vectors showed similar distribution, but with hSYN and, to lesser extent, with EF-1α vector GFP immunostaining can also be seen in the substantia nigra pars reticulata fibers (Fig. 1d–f), indicating expression of GFP in dopamine neurons. In contrast, CMV-driven GFP expression was not observed in the substantia nigra pars reticulata neurites. Accordingly, the colocalization of GFP and tyrosine hydroxylase (TH) immunostaining was clearly highest with the vector using the hSYN promoter (Fig. 1g–i). Although we cannot exclude that CMV promoter-driven GFP expression caused the increased death of dopamine neurons, we find that this option is rather unlikely. Because transduction with the CMV-GFP lentiviral vector showed hardly any TH-positive neurons in our experiments we conclude that CMV is not an optimal promoter for targeting dopamine neurons, at least in the mouse and rat. Also, there can be differences between species and laboratory animal strains. Nonoptimal selection of the CMV promoter to drive rat intranigral CDNF and MANF expression using lentiviral vectors 92 may explain why there was no neuroprotective effect of CDNF or MANF when used alone, in contrast to our studies with recombinant proteins in rats and mice. 21,48,59 Interestingly, CMV promoter-driven AAV vectors for intrastriatal CDNF expression were efficient in a rat 6-OHDA model of PD, 93,94 demonstrating the importance of both the site of injection and promoter for the therapeutic outcome.
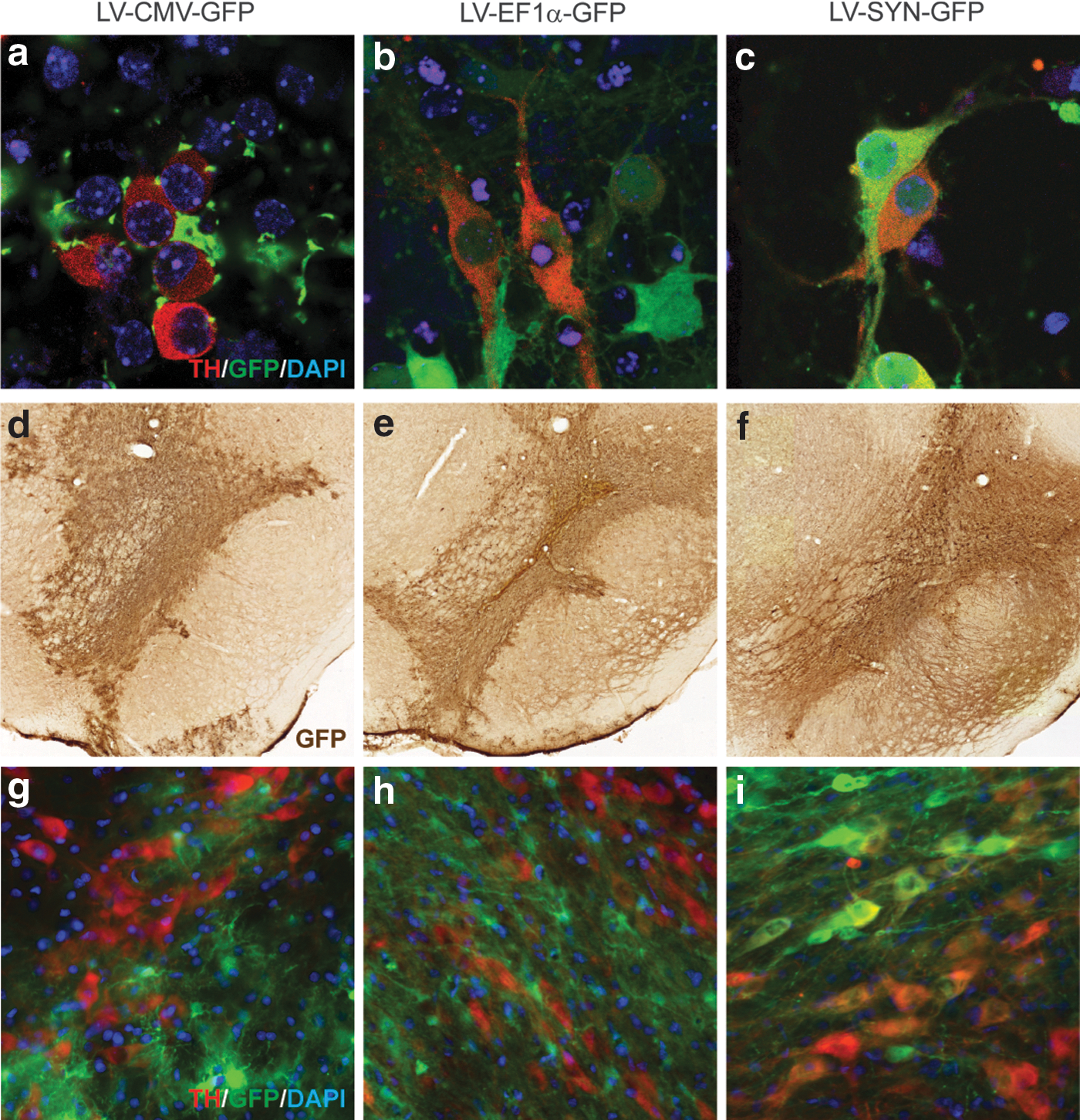
Expression of green fluorescent protein (GFP) and its colocalization with tyrosine hydroxylase (TH) immunostaining in cultured embryonic dopamine neurons
Continuous expression of NTFs with gene therapy vectors has caused significant side effects. 95,96 Intranigral injections of GDNF actually increased the loss of striatal neurons in rats after ischemic stroke, 95 which may be related to stimulatory effect of GDNF on dopamine neurons. Continuous expression of GDNF may lead to increased dopamine release and dopamine-induced toxicity in the striatum. 95 Similarly, prolonged expression of GDNF in dopamine neurons decreased striatal TH expression, induced aberrant neuronal sprouting, and failed to improve motor performance in a rat 6-OHDA model of PD. 96 Also, the limitation of targeting NTFs into the brain via constitutively expressing AAVs is that in many cases the optimal therapeutic approach would be achieved via controlled and noncontinuous expression. There are currently two prominent approaches for controlled transgene expression. The first uses an autoregulatory Tet-On AAV vector containing a bidirectional tetracycline-responsive promoter driving the expression of reverse tetracycline transactivator and the transgene of interest. 97 The basal transgene expression in this system in the noninduced state is low, whereas after activation by doxycycline the expression levels are comparable or higher than obtained with the CMV promoter. 97,98 A similar system using mifepristone, an approved small-molecule drug, has been described 99 ; optimizing the AAV serotype and timing of GDNF transgene expression resulted in efficient neuroprotection in a rat 6-OHDA PD model. 100 The other approach is to use a dihydrofolate reductase-derived destabilizing domain (DD) fused to the transgene that leads to rapid degradation of the entire fusion protein; the DD is stabilized by the small-molecule ligand trimethoprim, which can penetrate the blood–brain barrier, allowing for controlled transgene expression. 101 This system has been used to drive reversible regulated GDNF expression in the rat brain. 102
The secretory pathway, secretion stimuli, splicing of pre-pro forms, and signal sequence can all have an effect on therapeutic potency. There are two conserved GDNF splice variants, [pre-(α)pro-GDNF] and [pre-(β)pro-GDNF], lacking 78 bp in the pro-domain-encoding region, expressed both in the CNS and in many nonneuronal tissues, that produce identical mature GDNF. 103 –105 These splice variants are secreted both as pro-GDNF as well as mature GDNF, but whereas (α)pro-GDNF is secreted constitutively, the secretion of (β)pro-GDNF depends on neuronal activity. 105 MANF and CDNF have canonical ER retention C-terminal sequences. The ER retention of MANF is mediated by the interaction of a C-terminal RTDL sequence with a KDEL ER protein retention receptor (KDELR). 64 CDNF has a similar C-terminal KTEL sequence. CDNF and MANF are secreted from cells in response to ER stress inducers such as thapsigargin and tunicamycin. 63,64,106,107
Current and Past Clinical Trials
Several clinical trials using NTFs to treat patients with PD have been performed, 11,79,81 and three are currently ongoing (Table 2). Despite the limited efficacy of previous trials they did provide important results and we can learn to improve current therapies. 79 As mentioned previously, for viral vector-based therapy, the careful choice of viral vector type, delivery method, promoter, virus dose, injection site, and possibility to control and fine-tune the transgene expression appears to be of great importance for a successful outcome. All these aspects need to be considered, based on the biological properties of the NTF used. The promoter needs to be selected on the basis of target expression level and target neuronal population. Also, the virus dose needs to be estimated on the basis of the desired optimal target expression level. For proteins with poor diffusion properties, mutants with better diffusion properties would be needed. In addition, many NTFs are potent molecules and therefore it is likely that regulated expression systems would be optimal. For therapeutic strategies based on protein delivery it is also important to develop reliable and robust methods to assess the biological activity of recombinant proteins before their injection into patients. Current clinical trials using intrastriatal infusions of AAV2-GDNF will employ an improved delivery method, carefully tested in a number of preclinical studies in primates. 108,109 The ongoing trials of recombinant GDNF protein also use an intermittent convection-enhanced delivery protocol that is expected to result in more homogeneous distribution of GDNF over larger areas of the striatum to prevent protein accumulation and toxicity observed in previous studies. 110 A phase 1/2 clinical trial for human recombinant CDNF protein is planned to start in 2015, using a novel intermittent convection-enhanced delivery method.
Future Directions
As discussed previously, to find a neuroprotective strategy for dopamine neurons in PD is not an easy task. To successfully develop treatment to stop the progression of the disease, several aspects should be taken into consideration. The physical properties of NTFs play a crucial role in their pharmacokinetics, which in turn influence their therapeutic effect. For example, the diffusion properties and secretion of NTFs can be limiting factors in the human brain. Furthermore, it is important to develop better diagnostic tools to detect the earliest stages of PD, enabling proper selection of subjects who can then be treated at an earlier disease stage. Patient selection criteria can be improved by modern brain imaging techniques.
For the viral vector-mediated gene expression approach it is important to use proper serotypes, promoters, and controlled transgene expression. For the protein-based approach, one should carefully select proper concentrations, delivery site, and injection method (continuous vs. intermittent). Use of proteins expressed and purified from mammalian cells should be seriously considered. More careful design of therapeutic vectors and optimization of protein delivery conditions will lead to the development of new clinical trials. Moreover, combination therapies with molecules having different mechanisms of action and similar neuroprotective effects could lead to better clinical outcomes.
Finally, we should continue our efforts to search for new molecules with neurotrophic potential and compounds that regulate levels of endogenous NTFs, including small-molecule compounds and relatively novel therapeutic candidates such as microRNAs or long noncoding RNAs, and improve delivery methods to selectively target dopamine neurons in the adult brain.
Thus, although NTF treatments have faced some drawbacks in the first clinical trials, the strategy to restore the dopamine phenotype with proteins possessing neurotrophic properties is a valid and fully viable therapeutic approach for PD and needs further investigation.
Footnotes
Acknowledgment
The authors thank Katrina Albert for critical reading of the manuscript.
Author Disclosure
The authors have no conflicts of interest to disclose.