
Abstract
Retroviral vector-mediated gene therapy is promising, but genotoxicity has limited its use in the clinic. Genotoxicity is highly dependent on the retroviral vector used, and foamy viral (FV) vectors appear relatively safe. However, internal promoters may still potentially activate nearby genes. We developed insulated FV vectors, using four previously described insulators: a version of the well-studied chicken hypersensitivity site 4 insulator (650cHS4), two synthetic CCCTC-binding factor (CTCF)-based insulators, and an insulator based on the CCAAT box-binding transcription factor/nuclear factor I (7xCTF/NF1). We directly compared these insulators for enhancer-blocking activity, effect on FV vector titer, and fidelity of transfer to both proviral long terminal repeats. The synthetic CTCF-based insulators had the strongest insulating activity, but reduced titers significantly. The 7xCTF/NF1 insulator did not reduce titers but had weak insulating activity. The 650cHS4-insulated FV vector was identified as the overall most promising vector. Uninsulated and 650cHS4-insulated FV vectors were both significantly less genotoxic than gammaretroviral vectors. Integration sites were evaluated in cord blood CD34+ cells and the 650cHS4-insulated FV vector had fewer hotspots compared with an uninsulated FV vector. These data suggest that insulated FV vectors are promising for hematopoietic stem cell gene therapy.
Introduction
R
Increasing the safety of retroviral vectors can be approached in several different ways, and the choice of vector parent virus is paramount. Retroviruses have specific integration site preferences that, in addition to removing the native viral enhancers, can be used to increase the safety of retroviral vectors. 20 Vectors developed from foamy virus (FV), the only member of the Spumaretrovirinae subfamily of retroviruses, have an integration profile that is potentially safer than other retroviral vectors. 20 –24 FV vectors have a reduced preference for integration within CpG islands known to be DNA regulatory regions and near promoters compared with GV vectors, and a reduced likelihood to integrate within genes relative to lentiviral (LV) vectors. 20,21 In addition to a more desirable integration profile, FV vectors also have a lower propensity to dysregulate nearby genes when directly compared with GV and LV vectors. 25 These increased safety features, in addition to a large therapeutic transgene-carrying capacity 17 and broad tissue tropism including human mobilized peripheral blood and cord blood-derived CD34+ cells, 23,26 suggest that FV vectors are a promising vector for safer HSC gene therapy.
FV vectors appear to be relatively safe, but the internal promoter-enhancers necessary for therapeutic gene expression could potentially activate nearby genes. A proven approach to reduce this source of genotoxicity is to add chromatin insulators to retroviral vectors. 18,27 –29 Chromatin insulators are DNA sequences that protect genes from inappropriate transcription, often by recruiting proteins necessary for chromatin remodeling and scaffold attachment. 30 –32 There are two types of insulators: barrier insulators and enhancer-blocking insulators. 30 Barrier insulators are sequences that allow for an abrupt change from closed to open chromatin and protect genes from being silenced. Enhancer-blocking insulators, hereafter referred to as insulators, prevent enhancer elements from acting on promoters when the insulator is between an enhancer and promoter. We hypothesized that adding insulators to FV vectors might improve their safety.
Here four previously described promising candidate insulators were evaluated for use in FV vectors. A 650-bp version of the well-studied chicken hypersensitivity site 4 (cHS4) originally described by Arumugam and colleagues 33 and three synthetic insulators described by Gaussin and colleagues 34 were tested. The 650-bp cHS4 (650cHS4) is a combination of the 5′ 250-bp core, which contains a CCCTC-binding factor (CTCF)-binding site, and the 3′ 400 bp of the 1.2-kbp cHS4. 35,36 This 650cHS4 insulator reduces the genotoxicity of GV and LV vectors. 18,33 Of the three synthetic insulators tested here, two have multiple binding sites for CTCF and the third has multiple binding sites for the CCAAT box-binding factor/nuclear factor 1 (CTF/NF1). The CTF/NF1 protein is believed to be involved in barrier insulation and its DNA-binding sequence seems to impart enhancer-blocking activity when presented in multiple repeats. 34,37 –39 Reduced genotoxicity of GV vectors has previously been demonstrated with all of these insulators. Using a series of assays, including a plasmid-based assay to evaluate insulator activity and a novel shuttle vector approach to assess the fidelity of insulator transfer from the vector plasmid to the integrated vector provirus, a cHS4-based insulated FV vector was found to be promising and evaluated for its relative genotoxicity.
Materials and Methods
Cell culture
All cell lines were maintained at 37°C and 5% CO2. HEK293T and HT1080 fibroblasts were cultured in HyClone high-glucose Dulbecco's modified Eagle's medium (DMEM) (SH30022.01; Thermo Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (FBS) (S11550; Atlanta Biologicals, Lawrenceville, GA) and penicillin–streptomycin (50 U/ml) (17-602E; Lonza, Walkersville, MD). WEHI-3 cells (TIB-68; American Type Culture Collection [ATCC], Manassas, VA) were maintained between 2 × 105 and 2 × 106 cells/ml in Iscove's modified Dulbecco's medium (IMDM) (12-722F; Lonza) supplemented with 10% FBS, 0.05 M 2-mercaptoethanol (Sigma-Aldrich, St. Louis, MO), and penicillin–streptomycin (50 U/ml). For the production of WEHI CM IL-3 tissue culture additive, 2 × 105 cells/ml were plated and cultured for 5 days. Medium was then collected, clarified by centrifugation at 400 × g, filtered through a 0.22-μm (pore size) filter (Millipore, Bedford, MA), and stored at –20°C. 32D cells (CRL-11346; ATCC) were maintained between 2 × 105 and 1 × 106 cells/ml in RPMI 1640 (SH30027.01; Thermo Scientific) supplemented with 10% FBS, 10% WEHI CM, and penicillin–streptomycin (50 U/ml). For interleukin (IL)-3 depletion experiments, 32D cells were grown in RPMI, 10% FBS, and penicillin–streptomycin (50 U/ml) after extensive washing with phosphate-buffered saline (PBS).
Plasmids and plasmid construction
Insulator elements have been previously described. 33,34 Spacers in synthetic insulators were derived from mouse utrophin cDNA (GenBank accession number: BC062163.1) as previously described. 34 All insulators were synthesized with flanking NsiI restriction sites. The FV vector plasmid FV-PGW was previously described 40 and carries an enhanced green fluorescent protein (EGFP) reporter transgene expressed from a human phosphoglycerate kinase (PGK) promoter. FV-SGW is an FV vector plasmid containing an EGFP reporter transgene expressed from a spleen focus-forming virus (SFFV) promoter. These FV vector plasmids contain an added NsiI restriction site at the U3 deletion site of the 3′ LTR to facilitate inserting insulators. The FV-SGWKO vectors were developed by cloning an SalI restriction site-flanked R6Kγ bacterial origin of replication and kanamycin resistance cassette into the SalI restriction site within FV-SGW. The enhancer-blocking test plasmid was created by synthesizing the cytomegalovirus (CMV) enhancer, multiple cloning site with NsiI restriction site, CMV minimal promoter expressing mCherry, and poly(A) signals into a pUC57 backbone. The PGK-driven EGFP expression cassette was derived from the vector FV-PGW. 40 The 1.2-kbp cHS4 insulator was amplified by PCR from chicken genomic DNA (69233; Novagen/EMD Millipore, Darmstadt, Germany), using primers 5′-ATATTCCCCCATCCTCACTGA-3′ and 5′-GAAGAAAAGAAGCAGGCTTTC-3′ and Phusion high-fidelity polymerase (Thermo Scientific). Insulators were cloned into the NsiI restriction site of all plasmids and vectors. The LTRs of FV-PGW and insulated FV constructs were cloned into the multiple cloning site of the enhancer-blocking test plasmid with ClaI and PvuII. All insulators were synthesized by Blue Heron Biotechnology (Bothell, WA) or GenScript (Piscataway, NJ).
Vector production and titer
Vectors were produced on HEK293T cells plated on poly-
Enhancer-blocking activity assay
HEK293T cells (4 × 105) were plated on poly-
Shuttle vector rescue for insulator fidelity
HT1080 fibroblasts were infected with the insulated FV-SGWKO vector, sorted for EGFP expression to greater than 95% transduction, and expanded to at least 1.5 × 106 cells. DNA was then extracted from the cells, digested with the restriction enzyme NdeI, ligated and transformed into electrocompetent TransforMax EC100D pir + Escherichia coli (ECP09500; Epicentre, Madison, WI), and plated on kanamycin-containing plates. Colonies were isolated and sequenced by Beckman Coulter Genomics Services (Danvers, MA), using primers 5′-TATGCCTCCCGCTATGCTCG-3′ and 5′-CCTGTGGAACACCTACATCTG-3′ for the 5′ and 3′ LTRs, respectively. Retrieved sequences were aligned to vector sequence data using the BioEdit alignment tool (Ibis Biosciences, Carlsbad, CA). Retrieved sequences that did not contain chromosome junctions were further processed to verify integration. DNA was extracted from selected colonies, digested with AvrII, and sequenced (Macrogen Korea, Seoul, South Korea) using primer 5′-TAAACCGACTTGATTCGAGAA-3′.
32D genotoxicity assay
32D cells (2 × 106 or 1 × 108) were plated in log phase at 5 × 105 cells/ml in 32D WEHI CM medium and transduced with CL-SGN, FVSGW, or FVSGW-650cHS4-R vector at a multiplicity of infection (MOI) of 2 or with LV-SFFVEGFP at an MOI of 0.4, resulting in 3–5% transduction as previously described. 18 CL-SGN and LV-SFFVEGFP transductions were carried out in the presence of protamine sulfate (4 μg/ml). Cells were cultured for 48 hr, to allow for transduction, and then enriched by flow sorting for EGFP-expressing cells (S3 cell sorter; Bio-Rad, Hercules, CA). Sorted cells were allowed to recover in 32D WEHI CM medium for 24 hr and then thoroughly washed with PBS and transferred to RPMI supplemented with FBS and penicillin–streptomycin. Forty-eight hours later, cells were counted and plated at a concentration of 5 × 105 cells/ml in MethoCult semisolid medium (04230; Stemcell Technologies, Vancouver, BC) containing RPMI. Colonies were scored after 4–5 weeks of incubation.
Transduction of human cord blood CD34+ cells
Human cord blood CD34+ progenitor cells (2C-101; Lonza) were stimulated and maintained between 5 × 105 and 1 × 106 cells/ml in IMDM with 10% FBS and a 100-ng/ml concentration each of recombinant human interleukin (rhIL)-3, rhIL-6, recombinant human stem cell factor (rhSCF), recombinant human thrombopoietin (rhTPO), rhFlt-3, and recombinant human granulocyte colony-stimulating factor (rhG-CSF) (ProSpec, Ness-Ziona, Israel). After 24 hr of culture, 1 × 106 cells were harvested and exposed to FVSGW or FVSGW-650cHS4-R on CH-296-coated suspension culture plates at an MOI of 10 for 16 hr. Cells were then transferred to standard tissue culture-treated plates maintained for up to 10 days. Transduced cells were sorted by EGFP at 5 days posttransduction. At 10 days, remaining cells were harvested and DNA was extracted with a Gentra Puregene tissue kit (Qiagen, Valencia, CA).
Quantitative real-time PCR for relative retroviral vector integration copy number
DNA extracted from 5-day in vitro-cultured transduced CD34+ cells was assessed for copy number by quantitative real-time PCR assay for the EGFP transgene carried by the retroviral vectors. Forty nanograms of DNA was amplified in triplicate with the EGFP primers 5′-TGAGCAAAGACCCCAACGAG-3′ and 5′-TCGTCCATGCCGAGAGTGAT-3′. As an internal control, the β-globin gene was amplified with primers 5′-ATATCCCCCAGTTTAGTAGTTGGA-3′ and 5′-CCACAAGTATCACTAAGCTCGC-3′. PCR-amplified DNA for standard amplification curves was generated with the previously described primers. Standard curves were run in triplicate with a 10-fold dilution series ranging from 100 to 107 copies. Reactions contained 10 μl of Applied Biosystems SYBR green master mix (Thermo Fisher Scientific, Grand Island, NY) and a 0.2 μM concentration of each primer in a total volume of 20 μl. Quantitative real-time PCR was performed on a CFX384 Touch (Bio-Rad) under the following thermal cycling conditions: 95°C for 10 min, 40 cycles of 95°C for 15 sec and 61°C for 1 min, and a final extension at 65°C for 5 min. Vector copy number per cell is expressed as the ratio of EGFP gene copies to β-globin copies.
Modified genomic sequencing-polymerase chain reaction and integration site analysis
Three micrograms of genomic DNA was analyzed by modified genomic sequencing-polymerase chain reaction (MGS-PCR) as previously described
45,46
and sequenced on an Illumina MiSeq (Genomic Sequencing and Analysis Facility, University of Texas-Austin, Austin, TX). Briefly, genomic DNA was sheared to an average fragment size of 1.5 kbp with a HydroShear (Digilab, Marlborough, MA) set to a speed code of 12. Fragment ends were repaired (terminator end repair kit; Lucigen, Middleton, WI) and a linker sequence was ligated to the fragments. Fragments then underwent two sets of 30-round exponential PCR. The first, exponential PCR was done with biotin-tagged FV LTR primer 5′-ACCGACTTGATTCGAGAACC-3′ and linker primer 5′-GACCCGGGAGATCTGAATTC-3′. After MACS (magnetic-activated cell-sorting) bead purification (Dynabeads M-280 streptavidin; Thermo Fisher Scientific) to enrich for biotin-tagged fragments, samples were amplified by PCR with FV LTR primer containing a sample identifier sequence (XXXXXXX) and an Illumina MiSeq platform adaptor sequence (in boldface) 5′-
Results
Development of insulated FV vectors
We chose four insulators, previously shown to be effective in GV or LV vectors, for our studies (Fig. 1a). The first insulator we used is derived from the well-studied chicken hypersensitivity site four (cHS4). The cHS4 insulator is a 1.2-kbp genomic region within the chicken β-globin locus that binds many proteins, including CTCF, and has been extensively used in retroviral vectors to reduce genotoxicity. Previous studies have shown that cHS4 reduces retroviral vector titers. 33,50 Thus, for our studies we used the 650-bp version originally described by Arumugam and colleagues, 33 which has similar activity to the full-length cHS4. We also evaluated synthetic insulators described by Gaussin and colleagues. 34 These insulators have repeated CTCF-binding elements in order to increase their efficacy. 6xCTCF and 12xCTCF contain repeats of either the CTCF-binding sites from cHS4 and the blocking element α/δ (BEAD) insulator from the human α/δ T cell locus (6xCTCF) or the consensus CTCF-binding site (12xCTCF). 51,52 The 7xCTF/NF1 insulator is not based on CTCF binding but has seven repeats of the binding site for CTF/NF1. This insulator was originally described as a barrier insulator but may have blocking activity as well when repeated in tandem. 34,37,38
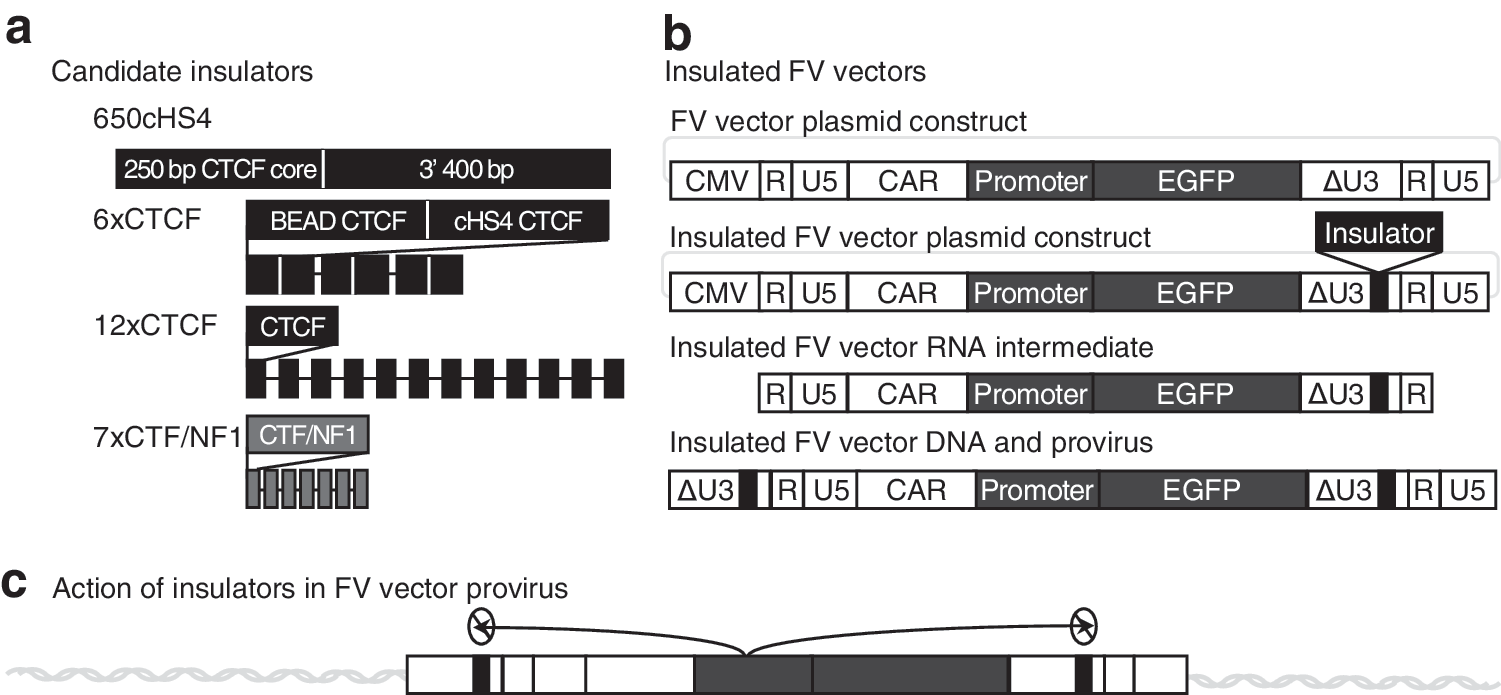
Insulated foamy viral (FV) vectors.
Similar to previously developed insulated retroviral vectors, 53,54 the insulators were positioned in the U3 region of the 3′ LTR of replication-incompetent FV vectors (Fig. 1b). 17,40 Enhancer elements function bidirectionally but insulators can prevent the enhancer activation of promoters only when between the enhancer and promoter. 30 To best prevent enhancer-mediated dysregulation of nearby genes, the therapeutic transgene cassette needs to be flanked by insulators. Placing the insulator in the U3 region of the 3′ LTR uses the retroviral/vector genome replication strategy in order to accomplish this flanking. During vector production the vector genome is transcribed from a plasmid and then reverse transcribed to make integration-competent replicate vector genomes. During reverse transcription the 3′-LTR U3 region is copied to the 5′ LTR. Addition of the insulator to the U3 region of the 3′ LTR results in vector genomes and subsequent integrated vector proviruses with insulators in both the 5′ and 3′ LTRs, effectively blocking any activity from the internal promoter on the upstream or downstream flanking host genome (Fig. 1c). To reduce the potential negative impact of insulator sequences on titer, the insulators were placed at the site of a 582-bp deletion made during previous vector development. 17 All developed vectors contain an EGFP gene cassette controlled by a PGK promoter or SFFV promoter in the location where a therapeutic transgene would be located.
Enhancer-blocking activity assay
To assess the insulating activity of the candidate insulators, we designed a dual-fluorescence enhancer-blocking test plasmid that features a separated CMV enhancer and minimal promoter (CMVe/CMVmin) controlling mCherry expression and a control EGFP expression cassette (Fig. 2a). Insulators were placed between the CMVe and CMVmin and the ratio of mCherry to EGFP expression was compared (Supplementary Fig. S1; supplementary data are available online at

Enhancer-blocking activity of candidate insulators.
Enhancer-blocking activity was found for all insulators with a broad range of activity (Fig. 2b). The 7xCTF/NF1 insulator was the least potent insulator and the 12xCTCF insulator was the most potent. Interestingly, there was a significant difference between the activities of the forward- and reverse-oriented 650cHS4 insulators, which was not observed with the synthetic insulators. To further investigate how insulators would function within the FV vector, the entire LTR from insulated FV vectors was then cloned between the CMVe and CMVmin (Fig. 2c). Inserting insulators into the FV vector LTR sequence significantly decreased the activity of the 6xCTCF insulator from 30 to 47%. For other insulators, insulator function stayed the same or was improved. Interestingly, the forward-oriented 650cHS4 insulator had observably stronger enhancer-blocking activity when tested in the presence of the FV LTR (45%) than in the absence of the LTR (80%) (Fig. 2b and c).
Effect of insulators on FV vector titer
LTR-insulated FV vectors containing an EGFP expression cassette were made in HEK293T cells and titered on HT1080 human fibroblasts (Fig. 3). For these experiments EGFP acted as a marker for transduction, which could be visualized by fluorescence microscopy and quantitated by flow cytometry. The 7xCTF/NF1 insulator had a minimal effect on FV vector titer. The two CTCF-based insulators with repeated CTCF elements, 12xCTCF and 6xCTCF, dramatically reduced titers. 6xCTCF was not evaluated further because of the low titers. Neither the size of the insulator, as seen with LV vectors, 50 nor the strength of the insulator had a consistent effect on titer. Interestingly, the titer of FV vectors with insulators in the forward orientation were consistently lower than FV vectors with insulators in the reverse orientation. However, the reverse-oriented 650cHS4 insulator could still be produced at a clinically relevant titer, less than 5-fold lower than the titer of an uninsulated FV vector, and more than 107 transducing units/ml after vector concentration. The choice of promoter to drive EGFP did not affect the changes in titer (data not shown).
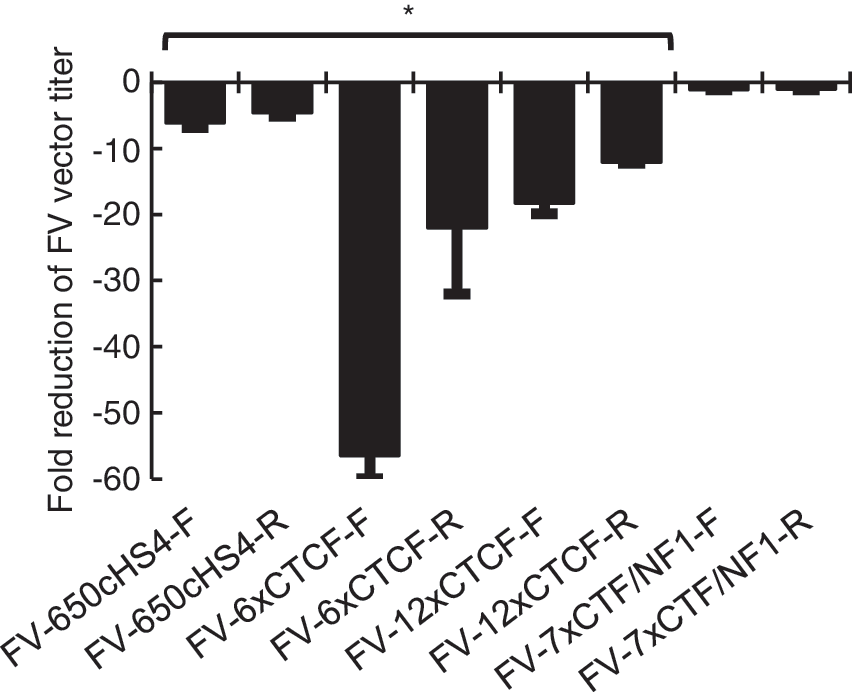
Titers of insulated FV vectors. HT1080 fibroblasts were transduced with FV vector preparations with the indicated insulators, and titers were determined by flow cytometry for EGFP expression. Columns represent the fold reduction in titer as compared with an uninsulated FV vector control. *p < 0.05 compared with control.
Fidelity of transfer of insulators to proviral LTRs
For insulators to be effective, they must be retained as the 3′ LTR of the vector plasmid is copied to the 5′ and 3′ LTRs of the integrated vector provirus during vector production, reverse transcription, and integration. Previous studies have identified problems with the retention of repeated elements within retroviruses as well as retaining a tandem repeat of the 250-bp cHS4 core. 55 –57 To evaluate the fidelity of transfer of insulators during vector production and transduction a shuttle vector rescue (SVR) approach was used to isolate integrated vector proviruses and sequence the LTRs (Fig. 4a and b). 20 The SVR FV vector contains an R6Kγ bacterial origin of replication and a kanamycin resistance cassette between the transgene expression cassette and the FV vector LTR. After integration with the SVR FV vector, the host cell genome will contain integrated vector proviruses with bacterial origins of replication and kanamycin resistance cassettes. The DNA from the transduced cells can now be extracted and digested with a restriction enzyme that cuts the genomic DNA but does not cut the integrated provirus. Of the digested DNA, only the fragments containing integrated proviruses will have kanamycin resistance cassettes. These fragments can then be ligated and transformed into bacteria for isolation on kanamycin-containing plates. In this way, the complete provirus can be captured as a plasmid, and both LTRs can be sequenced for direct comparison. SVR thus allowed us to rapidly evaluate the fidelity of transfer of insulators from vector plasmids into both vector provirus LTRs. Both 650cHS4 and 7xCTF/NF1 were retained at high frequency regardless of orientation (Fig. 4c and Supplementary Table S1). The 12xCTCF insulator was not efficiently retained, with a dramatic decrease in fidelity when the insulator was in the forward orientation. For all captured proviruses where loss of an insulator occurred, the deletions were large (at least 150 bp) and only one to three CTCF-binding sites remained when the insulator was in the forward orientation. With few exceptions, the provirus 5′ and 3′ LTRs were identical, suggesting the errors occurred during reverse transcription. The insulator with the lowest fidelity tested in the fidelity assay, 12xCTCF in forward orientation, resulted in the lowest titer FV vector.
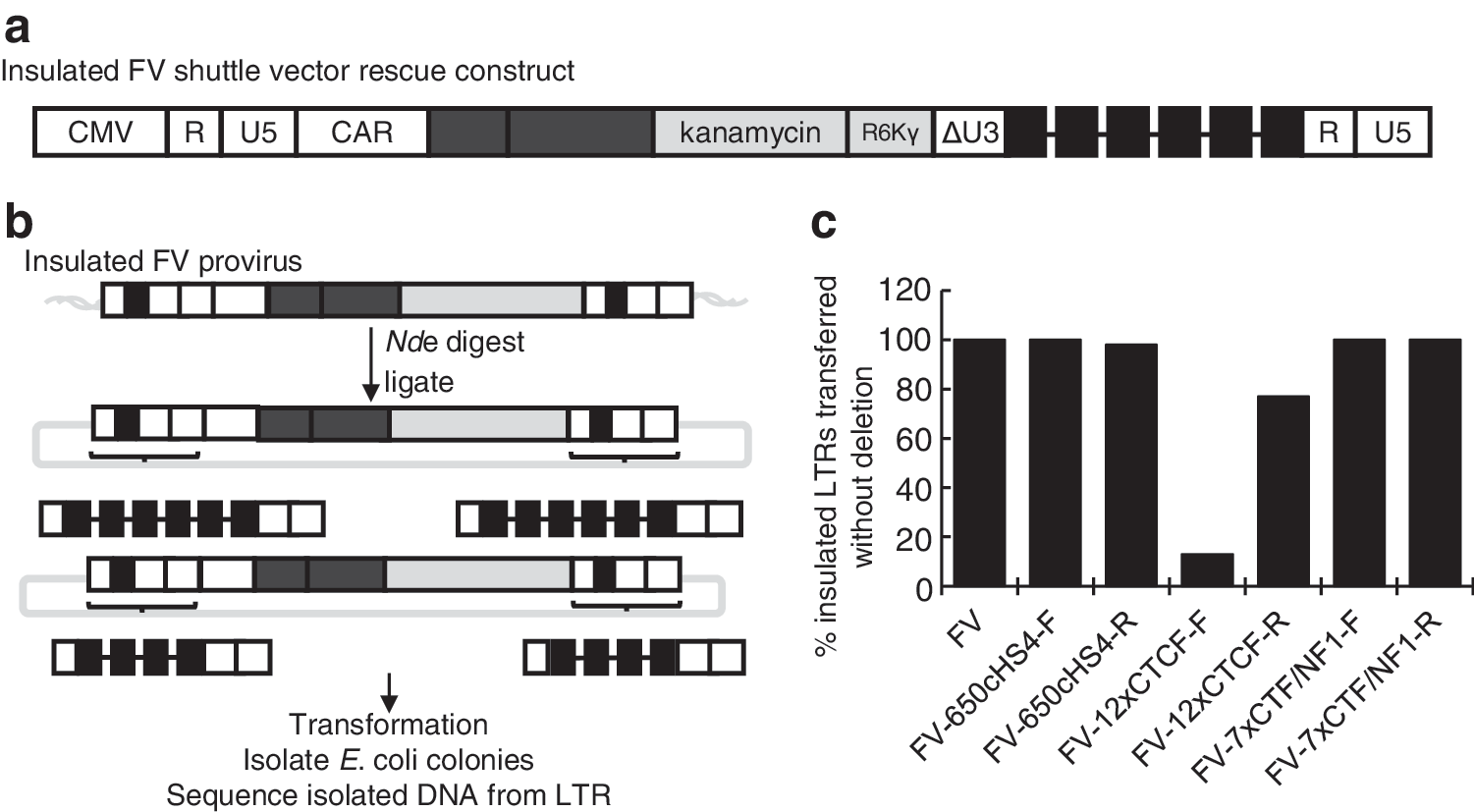
Shuttle vector rescue for analysis of insulator fidelity. A shuttle vector rescue strategy was used to isolate whole integrated vector proviruses and evaluate the presence of insulators from both the 5′ and 3′ LTRs.
Evaluating the safety of insulated FV vectors in vitro
The FV vectors containing the 650cHS4 in reverse orientation were determined to be the most promising insulated vectors for further studies because of a clinically relevant titer, significant enhancer-blocking activity, and efficient retention of insulators in both LTRs within integrated vector proviruses. Therefore we began assessing the genotoxicity of these vectors in vitro, using a previously described assay using 32D myeloid cells. 18 32D cells are an IL-3-dependent cell line where genotoxicity is measured by the frequency at which cells become IL-3 independent. IL-3 independence can be achieved by activation of autocrine IL-3 production or by activation of numerous oncogenes that negate the need for IL-3. 58 As described by Li and colleagues, 18 the 32D in vitro assay is 10-fold more sensitive to genotoxic events than assays using normal mouse bone marrow, and genotoxicity can be detected from GV with as few as 1 × 106 cells at 3% transduction efficiency.
Previous studies have suggested a reduced potential for FV vector genotoxicity compared with GV vectors. In anticipation of this, we used vectors with SFFV-controlled EGFP gene cassettes and increased the number of total cells in the assay. Although not a promoter currently used in retroviral gene therapy clinical trials, the SFFV promoter represents a worse-case scenario to fully stress the action of the insulators. For FV vectors we expanded the procedure to 1 × 108 cells at 5% transduction efficiency in order to detect possible genotoxicity. The 32D cell assay shows that FVSGW and FVSGW-650cHS4-R are both significantly less genotoxic than the control GV vector (CL-SGN) (Fig. 5a). Because LV vectors are becoming more popular for gene therapy studies because of their reduced genotoxicity compared with GV vectors, 28,59 1 × 108 32D cells were also transduced with an SFFV-containing LV vector (LV-SFFVEGFP). Consistent with previous findings, LV-SFFVEGFP is also significantly less genotoxic than CL-SGN with no detectable difference in the frequency of 32D colony formation compared with the FV vectors. Therefore, while CL-SGN consistently causes conversion of 32D cells to IL-3 independence, this conversion is not consistently detectable for FV or insulated FV and suggests that these vectors are both relatively safe and at least as safe as LV vectors.
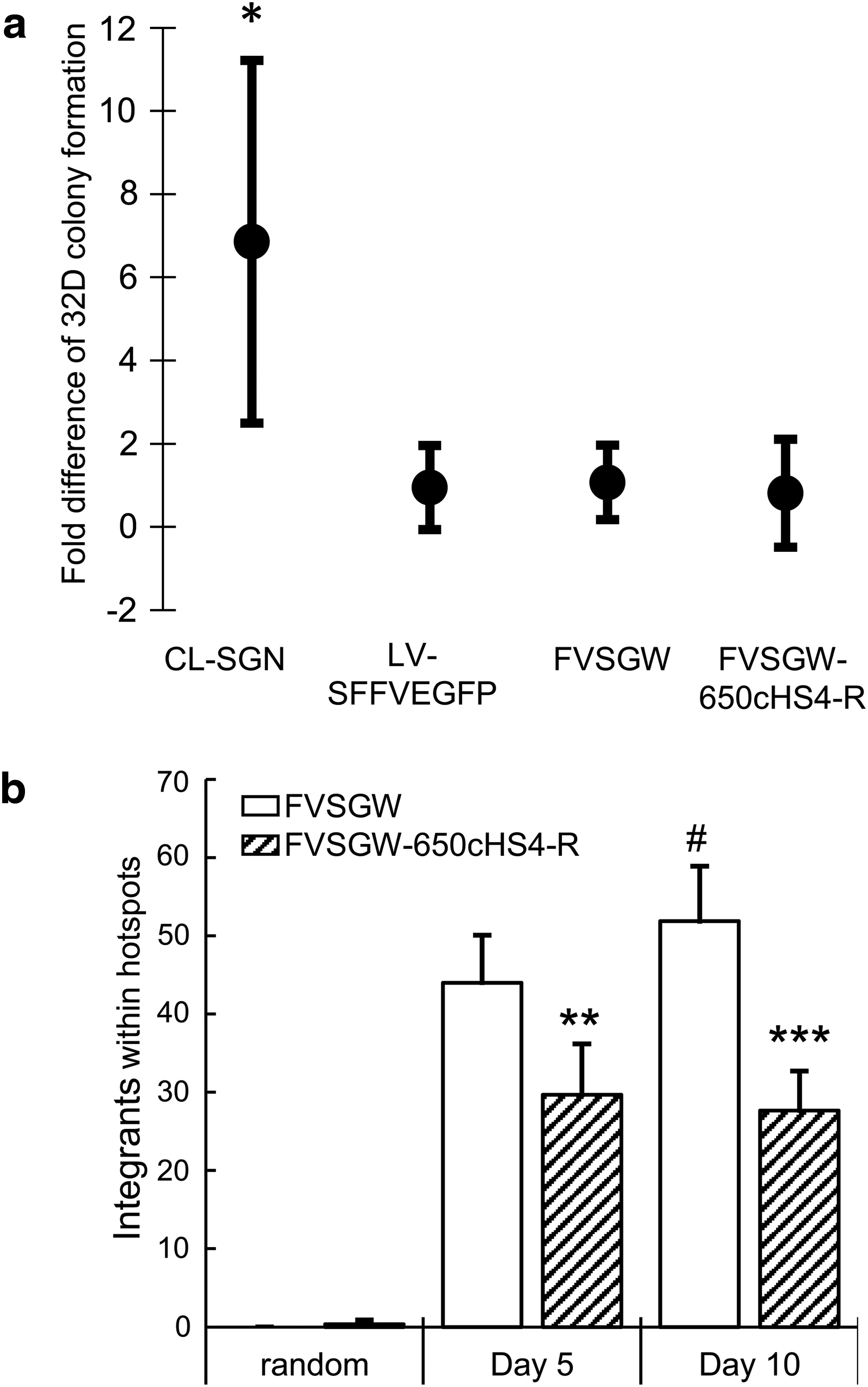
In vitro assessment of genotoxicity.
Reduced genotoxicity of insulated vectors in human cord blood CD34+ cells
Ultimately, insulated FV vectors used for HSC gene therapy will need to be safe in patient cells that are used in the clinic. We thus explored potential signs of genotoxicity in transduced and in vitro-cultured human CD34+ cord blood cells by performing RIS analysis of MGS-PCR-captured integration sites. Our goal was to determine whether the 650cHS4 insulator would affect the integration profile as well as evaluate integrations near proto-oncogenes and hotspots that may have clinical relevance and warn of potential genotoxic side effects in a clinically relevant target cell. Again, FV vectors containing a highly genotoxic SFFV promoter were used to maximize potential differences between the control uninsulated vector and the insulated vector. Cord blood CD34+ cells were transduced at an MOI of 10, resulting in 12.7 and 13.2% EGFP-expressing cells, respectively, for insulated and noninsulated FV vectors (Supplementary Fig. S2a). Before DNA extraction for RIS retrieval, samples were enriched to about 80% EGFP-expressing cells and equal transduction efficiency was verified by qPCR (Supplementary Fig. S2b and c). MGS-PCR of extracted DNA resulted in the capture of 4764 unique integrations from insulated FV vector-transduced samples and 9765 unique integrations from uninsulated FV vector-transduced samples (Supplementary Table S2). The integration profiles of FV and FVSGW-650cHS4-R were assessed after 5 days of in vitro culture (Supplementary Fig. S3). Addition of the cHS4 insulator did not significantly change the distribution of integration sites in proximity to transcription start sites (TSS) or DNase hypersensitivity sites, nor did it change the frequency of integrations within genes on day 5 after vector exposure (Supplementary Table S2). We also assessed integration sites for proximity to known proto-oncogenes and extended the culture time to 10 days to evaluate any potential skewing during in vitro culture. From the 14,529 integrations sites retrieved from day 5 and the additional 5976 sites and 13,120 sites retrieved from insulated and uninsulated FV vector-transduced samples, respectively, we were able to determine that there was no significant difference in the number of FV or FV-650cHS4-R vector integrations observed near proto-oncogenes (Supplementary Table S2).
We then assessed the frequency of integration hotspots. A hotspot is defined as an area of the genome where clusters of integrations are observed more frequently than expected by chance. On the basis of previous evaluations of GV and LV vector hotspots and common insertion sites we defined three integrations in a span of 50 kbp or less as a hotspot. 60,61 To avoid bias from integration site sample size differences, the MGS-PCR data sets were divided into a minimum of three nonoverlapping subsets of 1588 unique integration sites. Integrations within hotspots were then identified from these subsets. Fewer integration hotspots were observed from FVSGW-650cHS4-R-insulated FV vector-transduced CD34+ cells than from the uninsulated FV vector-transduced cells (Fig. 5b). We also observed a statistically significant increase in the number of hotspots captured from day 5 to day 10 in uninsulated FV vector-transduced samples that was not seen in the insulated counterpart. The integration hotspots were also less likely to be near known oncogene TSS, although these differences were not statistically significant (Supplementary Fig. S4). These data show that the 650cHS4 insulator reduced the frequency of integration hotspots when used in otherwise identical FV vectors.
Discussion
Retroviral vectors have enormous potential for HSC gene therapy, but genotoxicity continues to be a major concern for their clinical use. Here we describe the development of an insulated FV vector and show that in human cord blood CD34+ cells, fewer integrants occurred within hotspots compared with an uninsulated FV vector. We also describe a new rapid plasmid-based assay for insulator function and a shuttle vector assay to rapidly assess the fidelity of insulator transfer to the integrated vector provirus.
An insulated FV vector should have potent insulating activity and must be produced at high-enough titer for efficient HSC transduction. It should also maintain insulator fidelity during transfer of the insulator from the 3′ vector plasmid LTR to both LTRs of the integrated vector provirus in order to flank the transgene cassette. We found that the previously described 12xCTCF and 6xCTCF insulators were potent in a plasmid-based assay; however, they greatly reduced FV vector titers below what can be used in a clinical setting. Also, when the fidelity of transfer of the 12xCTCF insulator was evaluated, 33% (reverse orientation) to 87% (forward orientation) of integrated proviruses had deletions of the repeated CTCF elements. Taken together, these data suggest that repeated elements of CTCF mediate potent insulating activity, but future designs need to reduce recombination during vector production. This may be possible by creating repeated CTCF elements that are not based on a consensus CTCF element, but instead use different CTCF elements with mismatches to reduce the potential for recombination. Evaluation of CTCF-binding sites in the human genome has established hundreds of CTCF-binding domains, so designing repeated CTCF elements with potent insulating activity that do not reduce vector titers may be possible. 62 –65
Only the FV vectors with 650cHS4 in the reverse orientation fulfilled the criteria of significant insulating activity, high titer, and high fidelity of insulator transfer from vector plasmid to vector provirus. By using vectors with a strong SFFV internal promoter, we showed in the established and sensitive 32D assay that the FV vectors were significantly less genotoxic than a GV vector (CL-SGN) and similar in genotoxicity to an LV vector (LV-SFFVEGFP) (Fig. 5a). Neither the LV vectors nor the FV vectors produced consistently reproducible transformation of 32D cells to factor independence. As such, we were unable to establish a difference in genotoxicity between FVSGW and FVSGW-650cHS4-R with this assay. However, when integration sites in cord blood CD34+ cells were compared, a potential difference in safety was observed between the uninsulated and insulated FV vectors. The global distribution of captured integration sites from both vector-transduced samples were similar, as expected, with no significant difference in distribution of integrations near gene TSS, integrations within genes, or near or in known proto-oncogenes (Fig. 5b and Supplementary Table S2). We also did not identify clonal dominance within any of the samples, consistent with FV vector-transduced samples being highly polyclonal with and without an insulator. 41,66 Despite these similarities, a significant reduction in the number of observed integration sites within hotspots was observed when FV vectors were insulated. The effect is seen as early as 5 days after vector exposure and increases over time as indicated by a significant increase in the number of observed integrations within hotspots in uninsulated FV vector-transduced samples (Fig. 5b). Furthermore, we did not observe a change in the number of integrations within hotspots of insulated FV vector-transduced samples over time. Therefore our data show the insulator is potentially reducing the effects of the strong promoter on the surrounding host genome, thus reducing skewing from the initial polyclonal distribution of the transduced samples and inhibiting the proliferation of cells containing integration within a hotspot.
We speculate that the initial distribution of integration sites with respect to hotspots is not significantly different for the uninsulated and insulated FV vectors. This is because the insulator element is not expected to alter the integration profile. However, as early as day 5, the uninsulated vector is potentially promoting a growth advantage of hotspots (Fig. 5b). By day 10 this difference is significantly greater, which is consistent with 5 days being sufficient time for skewing of hotspots. In this study we chose to use a highly genotoxic promoter, SFFV, in order to maximize the potential to observe a reduction of skewing by an insulator element. In the setting of gene therapy a less genotoxic promoter would be preferred, but it must still provide sufficient transgene expression to correct the disease phenotype. 67,68 Also, genotoxicity has still been observed in some vectors with housekeeping promoters such as PGK, which may still benefit from the use of insulators in a clinical setting. 39 Additional studies to evaluate the effect of the FV-650cHS4-R-insulated FV vector on skewing in repopulating cells are warranted, but our data in human cord blood CD34+ cells suggest that insulated FV vectors may further improve FV safety.
Evaluating insulators in both forward and reverse orientation brought to light some interesting characteristics that should be taken under consideration when developing insulated vectors. We found that insulators can have variable activity depending on their orientation and that this can be changed in the context of the vector LTR. We also found that placing an insulator in the FV vector LTR can increase or decrease the activity of an insulator (Fig. 2b and c). The direct validation and comparison of insulator activity within the context of a retroviral LTR as we have shown here has previously been understudied, and is critical to assess insulators for use in retroviral vectors. Our enhancer-blocking activity assay allowed rapid assessment of insulators in either orientation and allows rapid assessment of activity within a LTR. As new CTCF-based and also non-CTCF-based insulator sequences continue to be identified, the plasmid-based enhancer activation assay we describe here should allow high-throughput analysis of candidate insulators to efficiently identify novel promising insulators.
In summary, our findings provide additional evidence that FV vectors are relatively safe for gene therapy. We developed a high-titer 650cHS4-insulated FV vector that transfers the insulator element with high fidelity to both proviral LTRs. We also showed that this insulated FV vector may be safer than an uninsulated FV vector, based on the frequency of integrants in hotspots in cord blood CD34+ cells. Previous studies with GV and LV vectors strongly suggest that insulators add to the safety of vectors 18,33,34,39 and this also seems to be the case for FV vectors. Our data strongly support further preclinical evaluation of insulated FV vectors for HSC gene therapy.
Footnotes
Acknowledgments
This research was supported by NIH grants AI097100 and AI102672 (G.D.T.). The authors thank Victor Bii for sharing the gammaretroviral vector CL-SGN before publication.
Author Disclosure
The authors have no competing financial interests in relation to the work described.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.