
Abstract
Gene therapy remains one of the potential strategies to achieve a cure for HIV infection. One of the major limitations of anti-HIV gene therapy concerns recovering an adequate number of modified cells to generate an HIV-proof immune system. Our study addresses this issue by developing a methodology that can mark conditional vector-transformed cells for selection and subsequently target HIV-infected cells for elimination by treatment with ganciclovir (GCV). We used the herpes simplex virus thymidine kinase (TK) mutant SR39, which is highly potent at killing cells at low GCV concentrations. This gene was cloned into a conditional HIV vector, pNL-GFPRRESA, which expresses the gene of interest as well as green fluorescent protein (GFP) in the presence of HIV Tat protein. We show here that TK-SR39 was more potent that wild-type TK (TK-WT) at eliminating infected cells at lower concentrations of GCV. As the vector expresses GFP in the presence of Tat, transient expression of Tat either by Tat RNA transfection or transduction by a nonintegrating lentiviral (NIL) vector marked the cells with GFP for selection. In cells selected by this strategy, TK-SR39 was more potent at limiting virus replication than TK-WT. Finally, in Jurkat cells modified and selected by this approach, infection with CXCR4-tropic Lai virus could be suppressed by treatment with GCV. GCV treatment limited the number of HIV-infected cells, virus production, as well as virus-induced cytopathic effects in this model. We provide proof of principle that TK-SR39 in a conditional HIV vector can provide a safe and effective anti-HIV strategy.
Introduction
A
Multiple approaches have been taken for HIV gene therapy, reviewed by Herrera-Carrillo and Berkhout. 9 These strategies include elimination of CCR5 and/or CXCR4 by gene editing using zinc finger 6,10 and CRISPR, 11 gp41 fusion inhibitory peptide maC46, 12 dominant negative viral proteins, 13 –15 shRNA, 7,8 and chimeric CD4+ T cell receptor (TCR). 16 Although many of these approaches have shown preclinical promise, the growing consensus suggests that a combination gene therapy approach incorporating multiple targets would be required to achieve long-term HIV suppression without ART, and possibly a cure. 9,17 Hence, there is a need for new anti-HIV gene therapy strategies that use novel genes and approaches.
The use of conditional vectors combined with cytotoxic or suicidal gene therapy would be an alternative approach to eliminate HIV-infected cells in patients in vivo. This can be brought about by various genes known to be classical inducers of apoptosis, 18 thereby leading to the destruction of “cells of interest” by apoptosis. Several suicidal gene therapy approaches have been tried in the past for eliminating HIV-infected cells. 19 –24 However, the approach has not been widely accepted because of the major limitation of lack of specificity required to target only HIV-infected cells and the modest efficacy of the suicide gene. In this regard, a point mutant of the herpes simplex virus (HSV)-1 thymidine kinase gene, TK-SR39, 25 has been shown to have severalfold higher activity than its wild-type counterpart (TK-WT) and has been tested in cancer gene therapy settings. 26 –29 TK-WT and TK-SR39 are kinase enzymes that phosphorylate the prodrug ganciclovir (GCV), which subsequently is incorporated in cellular DNA and inhibits DNA polymerases. 30 As the cytotoxic activity of this strategy requires both GCV and the TK gene to be present, it provides an additional layer of safety to be developed for in vivo gene therapy settings.
Finally, an important consideration for a successful HSC-based gene therapy is the engraftment of sufficient numbers of modified stem cells in the patient, so that they are able to eventually regenerate an HIV-proof immune system. Previous clinical studies using genetically modified stem cells in vivo have suggested the presence of low or undetectable numbers of modified cells in the peripheral blood a few months after administration. 5,8,31 Thus, there is an urgent need for ways to select the genetically modified stem cells in vitro before administration to patients to ensure robust engraftment.
In this study, we describe the use of a conditional cytotoxic gene therapy vector, pNL-GFPRRESA-TK-SR39, capable of specifically eliminating HIV-infected cells. This was achieved by using a highly active TK gene variant, SR39, in a conditional Tat-dependent lentiviral vector to specifically eliminate HIV-infected cells. Selective killing of HIV-infected cells was achieved by a combination of HIV Tat protein-dependent TK gene expression and GCV, providing several layers of safety while maintaining efficacy. Moreover, as the conditional vector expresses green fluorescent protein (GFP) under the control of HIV Tat, transient expression of Tat by Tat RNA, or using a nonintegrating lentiviral vector (NIL), allowed for selection of the gene-modified cells. We successfully modified and selected both TZM-bl and Jurkat cells with our vector and suppressed HIV replication by treatment with GCV.
Materials and Methods
Cells, transfection, and reagents
293T cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS). Transfections were performed with Fermentas TurboFect reagent (Thermo Fisher Scientific, Waltham, MA). TZM-bl cells were obtained from J. Kappes through the NIH AIDS Reagent Program (Germantown, MD) and cultured in DMEM supplemented with 10% FBS. TZM-bl infections were performed in the presence of DEAE-dextran (20–40 μg/ml; Sigma-Aldrich, St. Louis, MO). Jurkat and Jurkat-Tat cells were kindly provided by the NIH AIDS Reagent Program and maintained in RPMI medium supplemented with 10% FBS. For Jurkat-Tat cells, medium was supplemented with Gibco G418 (800 μg/ml; Thermo Fisher Scientific). These CD4+ T cells stably express HIV-1 Tat and are readily infected with HIV. CD34+ stem cells (STEMCELL Technologies, Vancouver, BC, Canada) were cultured in StemSpan serum-free expansion medium (SFEM) supplemented with CC110 cocktail (STEMCELL Technologies). The anti-herpes simplex virus drug ganciclovir was from Sigma-Aldrich, and integrase inhibitor 118-D-24 was kindly provided by the NIH AIDS Reagent Program.
DNA constructs, cloning, and mRNA production
The TK-WT and TK-SR39 genes (kind gift from M.E. Black, Washington State University, Pullman, WA) were amplified with specific primers and cloned into pcDNA3.1, using an Invitrogen pcDNA3.1 directional TOPO expression kit (Thermo Fisher Scientific). The conditional vector pGFPRRESA was kindly provided by Dr. Y. Wu (George Mason University, Fairfax, VA), and the TK-WT and TK-SR39 genes were cloned into the vector followed by authentication by sequencing and Western blotting. The pEV280-Tat101 construct was kindly provided by the NIH AIDS Reagent Program and cloned into an Invitrogen ViraPower lentiviral expression system (Thermo Fisher Scientific) for packaging into viral particles. The nonintegrating lentiviral packaging vector LENTI-Smart NIL was from Invivogen (San Diego, CA). Tat mRNA was prepared with an Ambion mMESSAGE mMACHINE T7 ULTRA transcription kit (Thermo Fisher Scientific). The full-length HIV proviral clones pNL-Lai, 32 pNL(AD8), 33 Bal, 34 JRSCF, 35 pNL-YU2, 36 and 89.637 have been previously described. GFP-expressing control lentiviral vector (pTY-EFeGFP) was kindly provided by the NIH AIDS Reagent Program and packaged as described subsequently.
Viral stock preparation and concentration
For preparation of TK viral stocks, 293T cells were transfected with the packaging DNA construct (TK vectors), helper DNA (pHP-dl-N/A), and vesicular stomatitis virus G glycoprotein (VSV-G) plasmid. For preparation of Tat-NIL viral stocks, cells were transfected with Tat DNA cloned into the lentiviral LENTI-Smart NIL packaging vector and VSV-G DNA. Viral stocks were harvested 48 hr posttransfection and either used directly for infection or after concentration, using Vivaspin columns (Sartorius, Goettingen, Germany), or ultracentrifugation at 20,000 × g for 90 min at 4°C.
Titration of viral stocks and transductions
SR39 lentiviral particles were titrated in Jurkat-Tat cells whereas Tat-NIL particles were titrated in sorted TZM-SR39/Jurkat SR39 cells. Briefly, 5 × 105 cells were infected with 2-fold dilutions of viral stocks starting with a maximum of 100 μl in the presence of Polybrene at 10 μg/ml. Cells were transferred to a 24-well plate and spinoculated at 800 × g, 35 min, 32°C in a final volume of 200 μl. Cells were washed with medium 3–5 hr postspinoculation and GFP expression was determined 72 hr postinfection. For transduction, cells were infected with different viral stocks either in the presence of DEAE-dextran (20–40 μg/ml; Sigma-Aldrich) for adherent cells or Polybrene (10 μg/ml; Sigma) for suspension cells in a total volume of 200 μl (100 μl of virus +100 μl of medium with Polybrene/DEAE-dextran). Suspension cells were spinoculated at 800 × g for 35 min at 32°C. After infection of Jurkat cells with Tat-NIL particles, cultures were treated with the integrase inhibitor 118-D-24 to further minimize the possibility of lentiviral integration.
Western blotting
To determine expression of TK protein in cells transduced with lentiviral vectors or transfected with plasmid DNA, the cells were lysed in radioimmunoprecipitation assay (RIPA) buffer (Sigma-Aldrich) 48 hr posttransfection/transduction. Lysates were run on Invitrogen 4–12% NuPAGE Bis-Tris gels (Thermo Fisher Scientific) and transferred onto Invitrogen polyvinylidene difluoride (PVDF) membranes (Thermo Fisher Scientific). Blots were blocked with 5% nonfat milk in Tris-buffered saline–Tween 20 (TBS-T) for 1 hr, followed by incubation with primary anti-TK antibody (diluted 1:1000; Santa Cruz Biotechnology, Santa Cruz, CA) overnight at 4°C. After three washes in TBS-T the blots were incubated with horseradish peroxidase-conjugated antibody (diluted 1:10,000; Sigma-Aldrich) for 1 hr. The blots were developed by enhanced chemiluminescence substrate, using the Pierce SuperSignal west femto maximum sensitivity substrate (Thermo Fisher Scientific), and images were acquired with a Syngene (Frederick, MD) gel imaging system.
Flow cytometry and cell sorting
For determination of GFP+ cells after transduction with various lentiviral stocks, cells were harvested and run on a 10-color Beckman Coulter (Brea, CA) Gallios flow cytometer. At least 20,000 events for each sample were acquired. Data were analyzed with FlowJo software (FlowJo, Ashland, OR). For the sorting of GFP+ cells, TZM-bl or Jurkat cells were transduced with the TK vectors followed by either RNA transfection (TZM-bl) or Tat-NIL transduction (Jurkat). Forty-eight hours posttransduction, GFP+ cells were sorted with the FACSAria II system (BD Biosciences, San Jose, CA). Propidium iodide (PI) staining (BD Pharmingen, San Diego, CA) was used to differentiate live from dead cell populations in accordance with the manufacturer's protocol.
Cytotoxicity assay
Cells seeded in 96-well plates were induced to express the TK gene either by transfection or transduction and treated with various concentrations of GCV. Cell viability was determined 48 hr later, using the CellTiter-Glo luminescent cell viability assay (Promega, Madison, WI), which determines the number of viable cells in culture based on quantitation of ATP as an indicator of metabolically active cells. Plates were read for luciferase activity, using a FLUOstar Omega plate reader (BMG Labtech, Offenburg, Germany) and data were analyzed with the Omega Data analysis software.
Virus replication assay
For multiple-round replication studies, parental Jurkat, Jurkat-TK-WT, or Jurkat-SR39 sorted cells were infected with ∼5 × 105 reverse transcriptase (RT) cpm of NL-Lai virus 32 for 2–3 hr at 37°C. The cells were treated with various concentrations of GCV immediately postinfection. The cultures were split 1:3 every second or third day postinfection, and culture supernatants were harvested for determination of RT activity and infectious virus. RT activity was determined as described previously. 14 For infectious virus determination, the TZM-bl indicator cell line was infected with 2 μl of harvested culture supernatants. Luciferase activity was determined 24 hr postinfection, using the britelite plus reporter gene assay system (PerkinElmer, Waltham, MA), and plates were read on a luciferase plate reader (BMG Labtech).
Data analysis
Most assays were conducted in triplicate and data represent means ± SD of triplicate observations. Data were analyzed and statistical analysis was performed with Excel software (Microsoft, Redmond, WA). Curve fits and IC50 (median inhibitory concentration) calculations were performed with SigmaPlot software (Systat Software, San Jose, CA). Flow cytometry data were analyzed with FlowJo software (FloJo). For fluorescence microscopy, images were acquired with a Nikon Ti microscope (Nikon Instruments, Melville, NY) and analyzed with NIS-Elements AR software.
Results
TK-SR39 construct shows significantly higher cell killing compared with its wild-type counterpart
The kinase activity of the HSV TK gene has been used for several scientific applications such as selection of hybridomas, 38 identification of dividing cells, 39 positron emission tomography (PET) scanning for active tumors, 40 and clinical chemistry, 41,42 including suicide gene therapy. 43 –45 HSV-1 TK has been successfully used for gene therapy in a wide variety of animal models and tried in clinical trials for human cancers. 46 –55 Cancer cells expressing the TK gene can be rendered susceptible to the prodrug GCV by inhibiting DNA synthesis. Although GCV is the drug of choice for several viral infections, its use may be limited in part because of toxicity. To overcome this phenomenon, Kokoris and Black 25 described the use of TK mutant SR39, which shows significantly higher activity compared with the wild-type enzyme as a result of enhanced phosphorylation, allowing the use of GCV at much lower concentrations. To test the efficacy of TK-WK and TK-SR39 for cell killing in our system, we cloned the above-described genes into the mammalian expression vector pcDNA3.1. The TK-SR39 gene exhibited significantly higher cell killing (∼180-fold) at lower GCV concentrations (IC50, 0.1447 μM) when compared with the wild-type gene (IC50, 26.05 μM) (Fig. 1A) in transfected HeLa cells. Both vectors expressed the TK protein to similar levels as determined by Western blot analyses (Fig. 1B), which demonstrated two bands corresponding to the two different isoforms of the TK protein. 56 Thus, the TK-SR39 gene exhibits better cell killing compared with its wild-type counterpart at lower GCV concentrations.

The thymidine kinase mutant SR39 (TK-SR39) construct shows significantly higher cell killing compared with its wild-type counterpart (TK-WT).
Development and characterization of conditional, cytotoxic anti-HIV gene therapy vector
The use of “conditional” cytotoxic gene therapy vectors for targeting HIV is optimal because (1) it is safe in that it causes minimal damage to uninfected cells by selectively targeting infected cells; (2) it minimizes the generation of resistant viral mutants, as the approach kills virus-infected cells rather than targeting a viral or cellular protein; (3) it can be packaged into virus-like particles (VLPs), and hence it offers ease of delivery and adaptability to modify a large number of cells; and (4) it has the potential to eliminate viral reservoirs, as the progeny cells arising from modified HSCs resist future infection. To achieve this objective, we cloned the TK-WT and TK-SR39 conditionally cytotoxic genes into the previously described conditional vector pNL-GFPRRESA, 20 which expresses the gene of interest under the control of HIV Tat. This vector was anticipated to have three unique properties: (1) It should express the TK gene only in the presence of HIV Tat; (2) on Tat expression or HIV infection the cells should also express GFP; and (3) when treated with GCV, the TK-expressing, HIV-infected cells should be eliminated (see Fig. 11). Hence, keeping in mind the above-described facts, expression of TK-WT and TK-SR39 genes by the conditional lentiviral vector (Fig. 2A) was tested by transient transfection in HeLa cells in the presence or absence of viral protein Tat. As seen in Fig. 2B, the pNL-GFPRRESA vector expressed TK-WT and TK-SR39 proteins only in the presence of HIV-1 Tat. Moreover, only cells transfected with Tat DNA expressed GFP when compared with control pcDNA-transfected cells (Fig. 2C). Having confirmed the Tat dependence of our conditional vector, we next asked whether this vector was capable of inhibiting HIV production by specific cytotoxicity in the presence of GCV. As shown in Fig. 2D, in cotransfection experiments with HIV pNL4-3, the TK-SR39 construct was highly effective at inhibiting HIV production compared with TK-WT. Both constructs inhibited infectious virus production only in the presence of GCV, and addition of GCV in the absence of conditional cytotoxic vector (compare control and control + GCV) had minimal effect on HIV inhibition. These data demonstrate that the pNL-GFPRRESA-TK vectors express the TK-WT or TK-SR39 gene in the presence of viral protein Tat and potently inhibit HIV when treated with GCV.
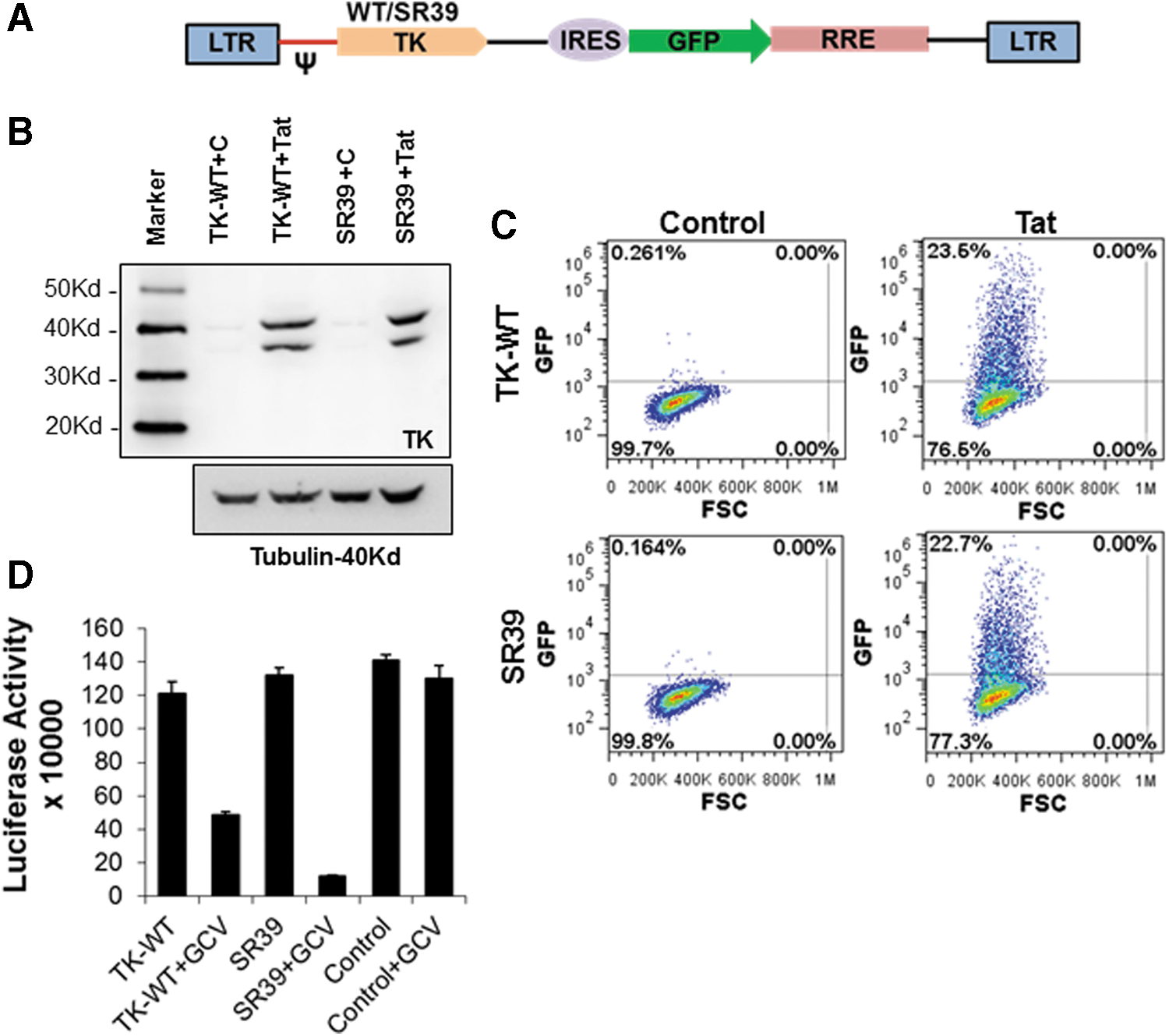
TK-WT and TK-SR39 genes cloned in the conditional vector pGFPRRESA express the TK gene and green fluorescent protein (GFP) in the presence of HIV Tat and inhibit HIV production in the presence of GCV.
pNL-GFPRRESA-TK-WT and pNL-GFPRRESA-TK-SR39 can be readily packaged into lentiviral particles and Tat DNA or RNA can be used to sort lentiviral vector-transduced cells
As demonstrated previously, in transfection experiments, the pGFPRRESA lentiviral vector expressed our proteins of interest (TK-WT or TK-SR39) only in the presence of HIV-1 Tat, which simultaneously resulted in GFP expression. Expression of GFP can be used to select cells that have been successfully modified with the gene therapy construct TK-SR39. For safety considerations, transient expression of Tat by Tat RNA rather than Tat DNA would be better suited for in vivo gene therapy purposes. We hence generated in vitro-transcribed HIV-1 Tat RNA and tested the efficacy of Tat expression in TZM-bl indicator cells that express luciferase in the presence of HIV Tat. As shown in Fig. 3A, TZM-bl cells transfected with various concentrations of Tat DNA or RNA showed robust luciferase expression, validating the effectiveness of Tat expression by RNA transfection. We next packaged our TK-WT and TK-SR39 genes into lentiviral particles and confirmed the Tat dependency of the pGFPRRESA vector. TZM-bl cells were transduced with pGFPRRESA-TK-WT or pGFPRRESA-TK-SR39 packaged virions followed by transfection with Tat RNA. The transduced and transfected TZM-bl cells were then assayed for GFP expression. As shown in Fig. 3B, Tat RNA transfection resulted in GFP-positive cells that were sorted to generate vector-transformed cell lines. This demonstrates that transient Tat expression by Tat RNA after transduction with our conditional lentiviral vector (TK-WT or TK-SR39) can be applied for cell enrichment/selection after genetic modification.

Tat RNA can be used to sort pGFPRRESA-TK-WT or pGFPRRESA-TK-SR39 lentiviral vector-transduced cells.
Sorted cells maintain dormant integrated lentiviral vector in a stable form
One of the primary requisites for a successful gene therapy vector is the ability of the integrated lentiviral construct to be stably maintained long-term in modified cells. We hence asked whether cells modified with our gene therapy vector pGFPRRESA-TK-SR39 maintained the TK-SR39 gene in a latent nonexpressed form over a prolonged period of time. For this purpose, the GFP-positive cells sorted previously were expanded in cell culture and monitored for GFP expression. As shown in Fig. 4A, sorted TK-WT- or TK-SR39-transduced cells lost GFP expression over time (Control). However, when retransfected with Tat DNA (data not shown) or Tat RNA, or when transduced with VSV-pseudotyped NL-Luc particles, they reexpressed GFP as determined by flow cytometry (Fig. 4A) and the TK gene as determined by Western blotting (Fig. 4B), confirming the maintenance of TK-WT and TK-SR39 genes in a stable but dormant form. To determine whether this dormant integration of the TK gene in the conditional lentiviral vector is stable, the above-sorted cells were maintained in culture for approximately 8 months (240 days) and then infected with VSV-pseudotyped NL-Luc viral particles to provide Tat protein. As shown in Fig. 4C, the TK-transduced cells maintained the TK-WT or TK-SR39 gene long-term in a stable form in vitro as evident by GFP expression. These data suggest that this conditional vector is ideal as a gene therapy vector as it not only allows for sorting of genetically modified cells but also maintains the TK gene in a dormant/latent but stable form.
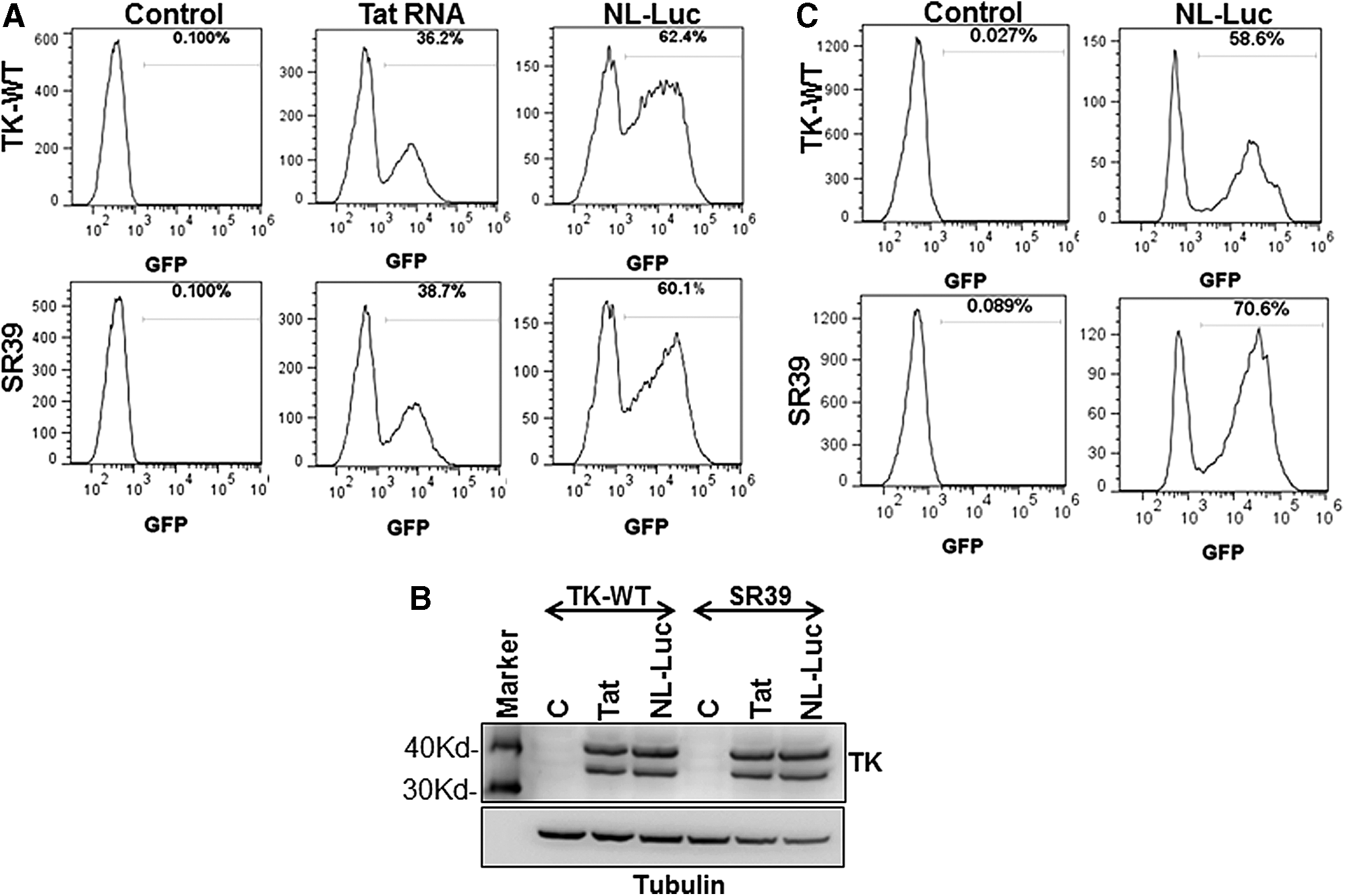
Sorted TZM-bl cells maintain dormant integrated lentiviral vector in a stable form. GFP-positive cells transduced with TK-WT or TK-SR39 lentiviral vector and sorted as described in Fig. 3 were expanded in cell culture for approximately 3–4 weeks.
Lentivirally expressed TK-SR39 construct shows significantly higher cell killing and HIV inhibition in the presence of lower GCV concentrations
We next tested the potency of the TK-SR39 gene in the pGFPRRESA lentiviral vector both in terms of cytotoxicity and HIV inhibition and compared it with TK-WT in the presence of various concentrations of GCV. It has been shown that the catalytically active site mutant of the TK gene, TK-SR39, is highly receptive to low concentrations of GCV as a result of enhanced phosphorylation. 25 To this end, TZM cells transduced with the pGFPRRESA-TK-WT or pGFPRRESA-TK-SR39 vector and sorted as described previously were either transfected with Tat DNA or infected with pNL-Luc/VSVG-pseudotyped virus to provide Tat in trans followed by treatment with various concentration of GCV. Cell viability was determined 48 hr later and, consistent with the findings in Fig. 1, the TK-SR39 construct exhibited significantly higher cell killing in the presence of lower GCV concentrations when compared with TK-WT (Fig. 5). As expected, the effect was more pronounced with NL-Luc infection (Fig. 5C) versus Tat DNA transfection (Fig. 5B), which could be due either to a higher percentage of cells being transduced versus transfected or to the presence of Rev protein in the former, which may further enhance expression from the vector. Interestingly, no significant loss in viability was evident in the absence of Tat protein (Fig. 5A).

Lentivirally expressed TK-SR39 construct shows significantly higher cell killing in the presence of lower GCV concentrations. Sorted TZM cells transduced with the TK-WT or TK-SR39 vector were either transfected with
We next looked at HIV inhibition in pGFPRRESA-TK-WT- or pGFPRRESA-TK-SR39-transduced cells in the presence of various GCV concentrations. As demonstrated in Fig. 6, multiple HIV strains were inhibited in both TK-WT- and TK-SR39-transduced cells treated with GCV. Interestingly, in most instances, the TK-SR39-transduced cells were more effective at HIV inhibition at lower concentrations of GCV when compared with the TK-WT cell line. The inhibition was similar for several of the R5 and X4 HIV isolates, suggesting that this approach was universally effective at inhibiting diverse HIV-1 strains.

HIV inhibition in cells lentivirally transduced with TK-WT or TK-SR39. Parental TZM cells or sorted TZM cells transduced with the TK-WT or TK-SR39 vector were infected with various HIV isolates. Four to 5 hr postinfection, culture medium was replaced with RPMI containing the indicated concentrations of GCV. Culture supernatants were harvested on day 3 (AD8, Bal, Lai, YU-2, 89.6) or day 6 (JRCSF) postinfection and used to infect TZM cells that were analyzed for luciferase activity 24 hr postinfection. Data represent means and SD of triplicate observations. *Significant difference (p < 0.05) in HIV production in TK-SR39-transduced cells compared with TK-WT.
Tat delivered by a nonintegrating lentiviral packaging vector can be used to sort TK-SR39-transduced cells
In the previous experiments it is clear that Tat DNA or RNA can be used to sort TK-WT- or TK-SR39-transduced cells, using Tat-dependent transient GFP expression. However, introduction of Tat DNA/RNA by various transfection methods as well as electroporation has the limitation of efficiency and associated cell toxicity, especially in lymphocytic cell lines. To address this issue, we cloned the Tat gene into a lentiviral packaging vector and packaged it into viral particles, using a nonintegrating lentiviral (NIL) helper construct. The integrase-deficient helper construct allows for delivery of the gene into cells by transduction, thus overcoming the toxicity and efficiency issues while retaining the advantage of transient GFP expression similar to Tat RNA transfection. To characterize the Tat-NIL particles, sorted TZM-TK-SR39 cells were infected with Tat-NIL-packaged lentiviral particles and monitored for GFP expression over a period of 15 days. As shown in Fig. 7A, cells infected with Tat-NIL lentiviral particles showed robust GFP expression on day 1 that gradually fell, with minimal GFP expression on day 12, which was further reduced by day 15. As control, a GFP vector packaged using an integrating helper construct (pHP-dl-N/A) maintained high GFP expression for more than 15 days. This suggests that expression of Tat by nonintegrating lentiviral particles could overcome some of the limitations associated with RNA transfection and provide an ample window of opportunity for sorting the GFP-positive cells.

The Tat-NIL (nonintegrating lentiviral) vector can be used to effectively sort TK-WT- and TK-SR39-transduced cells.
Having determined the safety and efficacy of the Tat-NIL vector, we next proceeded to modify a CD4+ lymphocytic cell line (Jurkat) with TK-WT or TK-SR39 lentiviral particles followed by infection with Tat-NIL lentiviral particles. Cells were then analyzed for GFP expression on day 3 (Fig. 7B) followed by sorting of GFP+ cells. The sorted cells were expanded in cell culture with a concomitant loss in GFP expression over time (Fig. 7C, control). Interestingly, once the cells were infected with NL-Luc to provide the Tat protein, the cells reexpressed GFP (Fig. 7C). These data demonstrate that Jurkat T cells could be stably transformed with pGFPRRESA-TK-WT or pGFPRRESA-TK-SR39 vectors that express GFP in the presence of viral protein Tat. To determine whether the sorted Jurkat cells were capable of expressing the TK protein in the presence of Tat, Jurkat-TK-WT or Jurkat-SR39 cells were infected with VSV-pseudotyped NL-Luc viral particles followed by Western blotting for the TK gene. As expected, TK gene expression was seen for both Jurkat-TK-WT and Jurkat-SR39 cells only after infection with NL-Luc (Fig. 7D). Moreover, after treatment with GCV a clear reduction in cell viability was evident for the Jurkat-SR39 cells (Fig. 7E), demonstrating the greater effectiveness of the TK-SR39 gene to induce cell killing compared with TK-WT at lower GCV concentrations. Thereafter, we also tested the feasibility of our approach in CD34+ stem cells. As shown in Fig. 7F, using this method we were able to modify and mark CD34+ stem cells after transduction with TK-SR39 followed by the Tat-NIL vector. Overall, these data demonstrate the effectiveness of the TK-SR39 gene and Tat-NIL vector to stably transduce T cells followed by their ability to be sorted by GFP expression. The data also demonstrate the effective conditional functioning of the TK gene only in the presence of HIV Tat protein and GCV.
TK-SR39-transduced cultures specifically deplete HIV-infected cells in the presence of GCV
We next asked whether our pGFPRRESA-TK-SR39 vector would prove efficacious in targeted elimination of HIV-infected cells in T cell lines. To this end, Jurkat-TK-WT or Jurkat-SR39 cells were infected with VSV-pseudotyped NL-Luc viral particles followed by treatment with GCV. On days 5 and 7, cell viability was determined by propidium iodide (PI) staining and infection by GFP expression, using flow cytometry. NL-Luc-infected cells undergoing apoptosis were determined as GFP+PI+ whereas infected and live cells were GFP+PI– populations. As shown in Fig. 8, in TK-SR39-transduced cells treated with GCV, there was a significant increase in GFP+PI+ cells when compared with cells cultured in the absence of GCV, suggesting that HIV-infected cells were specifically undergoing cell death (Fig. 8B). Interestingly, this increase was accompanied by a concomitant decrease in GFP+PI– cells (24.1 vs. 38.1% in 0 μM GCV and 6.2 vs. 70.4% in 20 μM GCV) (Fig. 8A). A similar trend was observed with TK-WT-transduced cells but to a lesser degree (26 vs. 35.3% in 0 μM GCV and 19.3 vs. 48.1% in 20 μM GCV). These findings suggest that the pGFPRRESA-TK-SR39 conditional vector is capable of specifically eliminating HIV-infected cells in the presence of GCV.
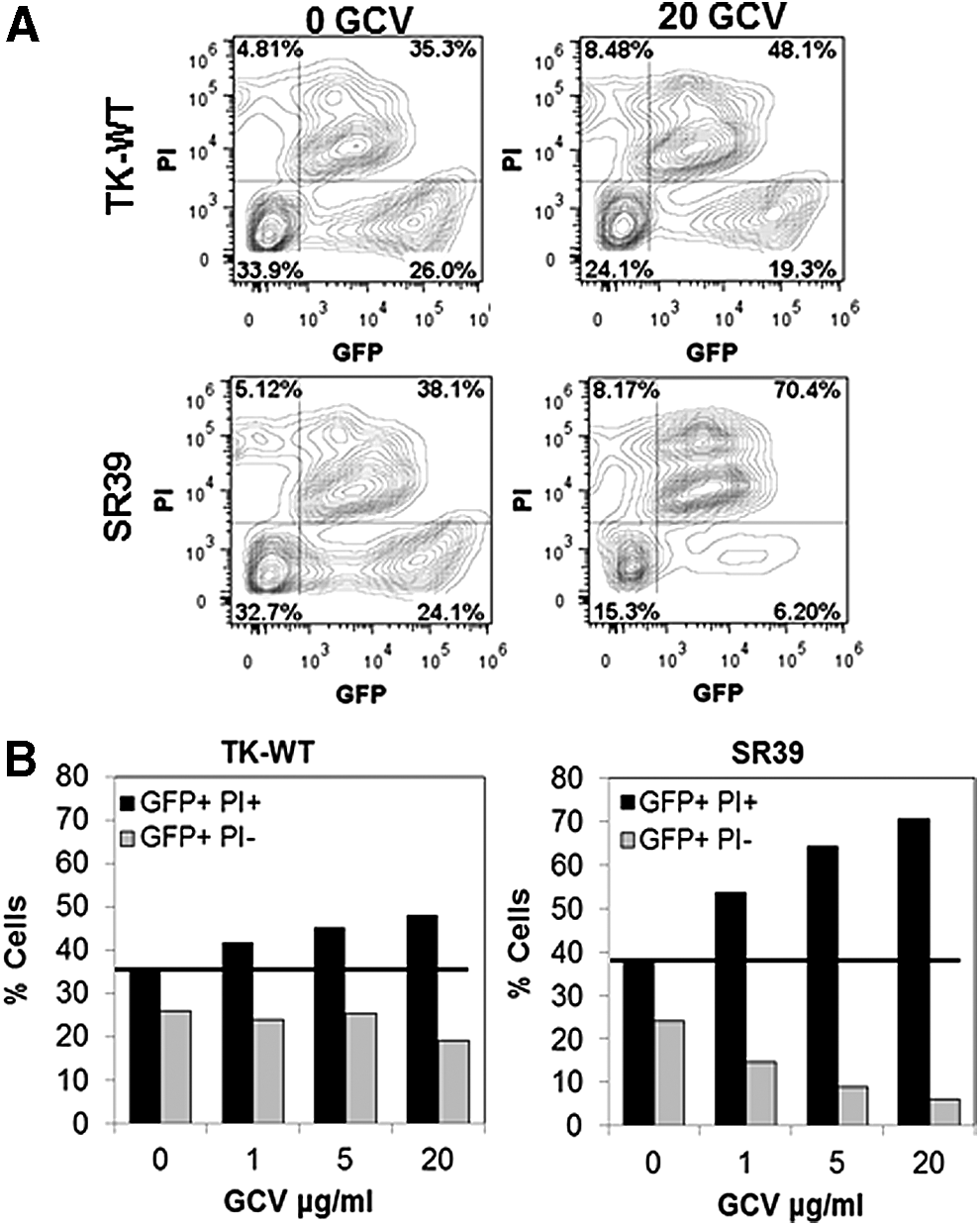
TK-SR39-transduced cultures specifically deplete HIV-infected cells in the presence of GCV. Sorted TK-WT- or TK-SR39-transduced Jurkat cells were infected with VSV pseudotyped NL-LUC followed by culture in the presence of various concentrations of GCV. On days 5 and 7, cells were stained with propidium iodide (PI) and analyzed by flow cytometry.
TK-SR39-transduced cells inhibit HIV replication and associated cytopathic effects in the presence of GCV
We next asked whether pGFPRRESA-TK-SR39-transduced Jurkat T cells in the presence of GCV were capable of limiting multiple-round HIV replication in cultures in vitro. For this, Jurkat-SR39 cells were infected with wild-type Lai virus (∼5 × 105 RT cpm) and monitored for RT activity and infectious virus in culture supernatants along with changes in cell viability and GFP expression. As shown in Fig. 9A and B, parental Jurkat cells infected with wild-type HIV showed robust virus replication at all GCV concentrations. Surprisingly, in this highly acute system of virus replication, TK-SR39 cells treated with GCV showed a significant, dose-dependent reduction in HIV replication as evidenced by a decrease in infectious virus (Fig. 9A) and RT activity (Fig. 9B) in culture supernatants. The inhibition of HIV replication by TK-WT was moderate and better than that of parental Jurkat cells but significantly less than that of TK-SR39 cells (Fig. 9D). Interestingly, this reduction in virus replication was accompanied by a preservation of cell viability in Jurkat-SR39 cells compared with Jurkat-TK-WT or parental Jurkat cells (Fig. 9C). Moreover, HIV-induced cytopathic effect as evidenced by syncytium formation was also significantly diminished in Jurkat-SR39 and Jurkat-TK-WT cells (Fig. 9E). These data suggest that the TK-SR39 gene can inhibit HIV replication in the presence of GCV, which translates to preservation of cell viability and reduction in cytopathic effects as a result of virus inhibition.
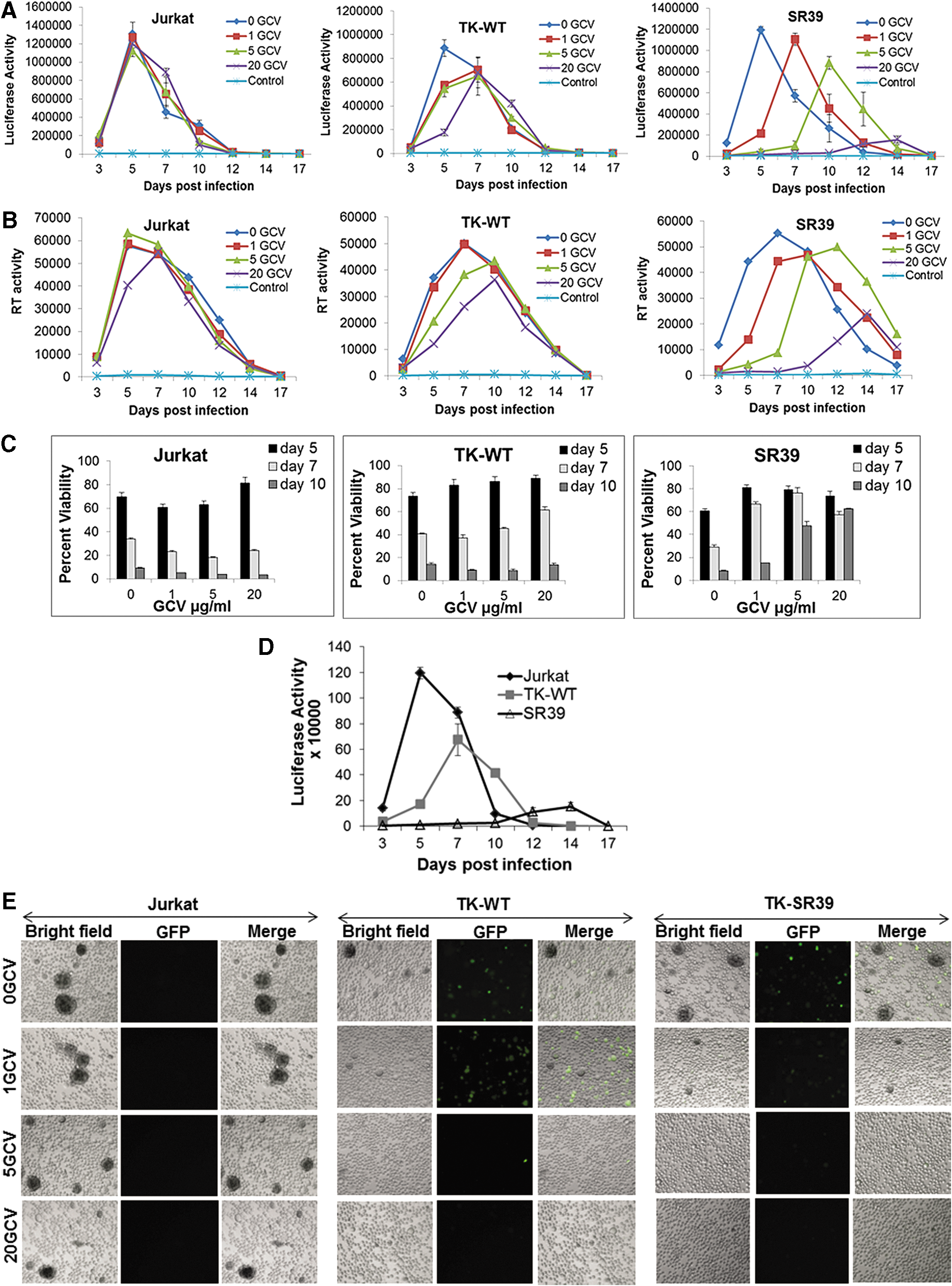
TK-SR39-modified cells inhibit HIV replication in the presence of GCV, allowing for preservation of cell viability in infected cultures. Parental Jurkat cells or Jurkat cells lentivirally transduced with TK-WT or TK-SR39 and then sorted were infected with HIV-Lai virus followed by culture in the presence of the indicated concentrations of GCV.
TK-SR39-transduced cells inhibit HIV replication by killing HIV-infected cells accompanied by preservation of live cell population
We next determined whether expression of the TK-SR39 gene in the presence of GCV led to a specific reduction in HIV-infected cells. For this, TK-WT- and TK-SR39-transduced Jurkat cells infected with HIV Lai were monitored for the presence of GFP-positive cells as well as a change in live versus dead cell population by forward scatter and side scatter flow cytometric analysis. As shown in Fig. 10A, Jurkat-SR39 cells showed a significant reduction in the percentage of GFP+ cells when compared with Jurkat-TK-WT over a period of 10 days in the presence of GCV. Because GFP is expressed by our vector only in the presence of HIV Tat, the reduction in GFP+ cells is indicative of a directed reduction in the number of HIV-infected cells. These finding correlated with a higher percentage of live cells for the TK-SR39 population in the presence of GCV. As shown in Fig. 10B, Jurkat-SR39 cells, when infected with HIV and treated with GCV, showed a percentage of cells in the live gate that was remarkably similar to that of the uninfected cells (70.6% for uninfected vs. 64.9% for infected in the presence of GCV). However, in the absence of GCV, there was a marked increase in cell death (79%) accompanied by a concomitant decrease in the live cells (16.5%). Clearly, with increasing GCV concentrations, in the SR39-transduced Jurkat cells infected with HIV there was a decrease in dead cells accompanied by preservation of the live cell population (Fig. 10C), indicating that a suppression of virus replication and virus-induced cytopathic effects can be achieved by this method. These data suggest that the TK-SR39 gene expressed in a conditional vector selectively limits HIV-infected cells in the presence of GCV, thereby preventing virus replication and spread and consequently limiting virus-induced CD4+ T cell depletion.
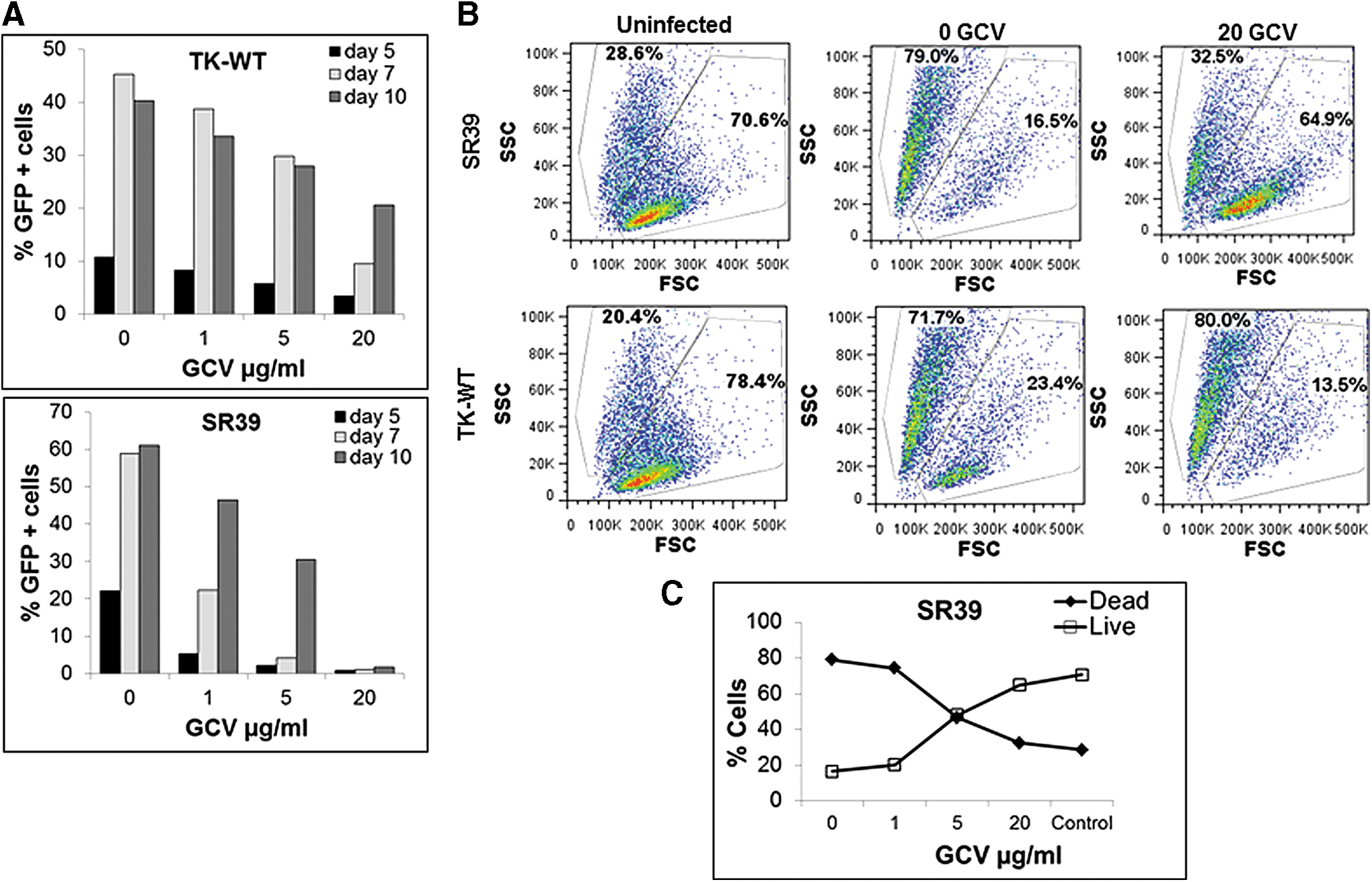
TK-SR39-modified cells inhibit HIV replication by killing HIV-infected cells accompanied by preservation of the live cell population. Sorted TK-WT- or TK-SR39-transduced Jurkat cells were infected with wild-type HIV-Lai virus followed by culture in the presence of various GCV concentrations.
Discussion
Successful treatment of the Berlin patient, using CCR5-negative cells from a CCR5Δ32 homozygous donor, has renewed interest in HIV gene therapy. 1,2 Although proof of principle for CD34+ HSC-based reconstitution of an HIV-proof immune system has been demonstrated, the applicability of this approach remains limited. For one, finding tissue-matched CCR5Δ32 homozygous donors for a large number of patients infected with HIV is impractical. Second, reconstituting with Δ32 cells leaves the possibility of virus switching to CXCR4 usage, which would essentially make the virus more pathogenic, a fear that has been realized in one study. 57
Modification of CD34+ HSCs with lentiviral vectors ex vivo before reconstitution in patients with HIV is an attractive approach. However, for HIV gene therapy to be successful, certain fundamental criteria need to be met: (1) The gene therapy approach should be safe and effective against a variety of HIV strains, and the possibility of development of virus resistance should be minimal; and (2) a method needs to be developed whereby the modified stem cells can be selectively enriched before transplantation. 31 This will enhance the reconstitution/engraftment efficiency and provide the recipient with an “HIV-proof” immune system. For safety concerns either transient methods such as zinc finger and/or CRISPR may provide the necessary safety for ex vivo modifications. Furthermore, safety concerns with lentiviral vectors can be overcome by using conditional vectors that limit the expression of the protein of interest in HIV-infected cells. 19 This can be achieved by using the viral LTR promoter that is dependent on the HIV-1 Tat protein 58,59 for activity, hence restricting the expression of proteins to infected cells and limiting the toxicity and side effects of the vector. Several HIV LTR-based Tat- and Rev-dependent lentiviral vectors have been developed by numerous groups. 8,10,24,36,42 Another important aspect of using conditional vectors is that the anti-HIV gene needs to target late stages of the virus, as Tat-mediated activation in infected cells would not prevent early steps in the HIV life cycle. In this regard, we have previously described the use of viral dominant-negative proteins that target the late stages of HIV life cycle. 14
In this study we have used the TK-SR39 gene in a conditional Tat-dependent vector for elimination of HIV-infected cells. Our strategy fulfills both the previously mentioned requirements by using a conditional vector and eliminating infected cells by Tat-dependent expression of the TK-SR39 gene, which induces cell death in the presence of ganciclovir. In some respects our approach reflects natural infections, in which CD8+ T cells can specifically eliminate HIV-infected cells by apoptosis. In fact, cytotoxic T lymphocyte (CTL)-mediated elimination of infected cells remains one of the most efficient ways to control HIV, often seen in long-term nonprogressors. 60,61 The same principle was used to develop a chimeric TCR by fusing the CD4 extracellular domain with the intracellular signaling domain of the TCR. These chimeric TCR-bearing CD8+ cells would potentially eliminate HIV-infected cells. 16
The efficacy of TK-SR39 and the superior activity of the mutant form in comparison with TK-WT 25,30 are clearly confirmed in our study. In fact, other studies have also demonstrated the efficacy of TK-SR39 in tumor models in mice 62,63 and elimination of autologous HSC-derived gene-modified cells in rhesus macaques. 43 The successful reconstitution of HSCs modified with lentivirus-delivered TK-SR39 in humanized mice suggests that our approach is feasible. 40 It has been reported that the TK-SR39 gene in the presence of GCV may have a bystander effect on unmodified neighboring cells. 64,65 However, considering the phenomenon of HIV transmission and the importance of cell-to-cell contact in this process, this may work to our advantage as bystander killing may prevent cell-to-cell transmission by eliminating the neighboring recently infected cells.
One of the major obstacles for ex vivo modification of HIV for transplantation is to either generate enough modified stem cells for reconstitution or develop a method to select modified stem cells to increase the chances of successful reconstitution with modified stem cells alone. In this regard, our study demonstrates that transient expression of Tat in vector-transduced cells, using either Tat RNA or Tat expressed by a nonintegrating lentiviral (NIL) 66 vector, can provide a means to transiently express GFP and sort the modified cells. We demonstrate here that both these methods can be used for selection of vector-modified cells with high efficiency, with Tat NIL vector superior compared with RNA transfection by limiting toxicity associated with electroporation in lymphocytic cell lines. The modified cells have stable integration of the conditional vector and do not express the TK genes in the absence of viral infection or Tat expression. Our study also demonstrates that in these selected cells, infection with HIV-1 results in rapid upregulation of TK-SR39 that mediates cell toxicity only in the presence of GCV and eliminates infected cells. This is manifested as efficient control of virus replication in multiple cell lines.
As a proof of principle our study shows that the TK-SR39 plus GCV (Fig. 11) is an efficient way to eliminate HIV-infected cells and suppress virus replication. Our study also develops a unique method for transiently marking the modified stem cells for selection before reconstitution in humans, thus significantly increasing the chances of reconstitution with an HIV-proof immune system. The use of GCV in patients with HIV is already approved and the safety profile of the drug is impeccable. 67 As our approach is directed toward targeting HIV-infected cells and not a particular HIV protein, the therapy is effective against both CXCR4-tropic and CCR5-tropic viruses as seen in our study. Also, the chances of development of resistance against the therapy are minimal as no viral proteins are being targeted. This study characterizes the safety and efficacy of TK-SR39 conditional vector modification of cells and subsequent control of virus replication, thereby paving the way for adaptation of this platform to other cell types such as peripheral blood mononuclear cells (PBMCs) and CD34+ HSCs.
In summary, we have developed a novel and safe conditional cytotoxic gene therapy approach for HIV by use of a vector that incorporates multiple safety features as well as a selection feature. We used a mutant form of the HSV-TK gene (TK-SR39 mutant) that is highly potent at killing cells in the presence of small amounts of GCV (Fig. 11). Our HIV-derived conditional vector expresses the TK-SR39 gene only in HIV-infected cells (in the presence of the viral protein Tat). Moreover, the vector has an added unique feature in that it incorporates a Tat-inducible GFP gene (Fig. 11). Thus, in vitro expression of Tat by RNA transfection or nonintegrating lentiviral transduction resulted in GFP expression in gene-modified cells that can then be sorted by flow cytometry. These gene-modified cells exhibited suppression of HIV replication by virus-specific cytotoxicity in the presence of GCV.
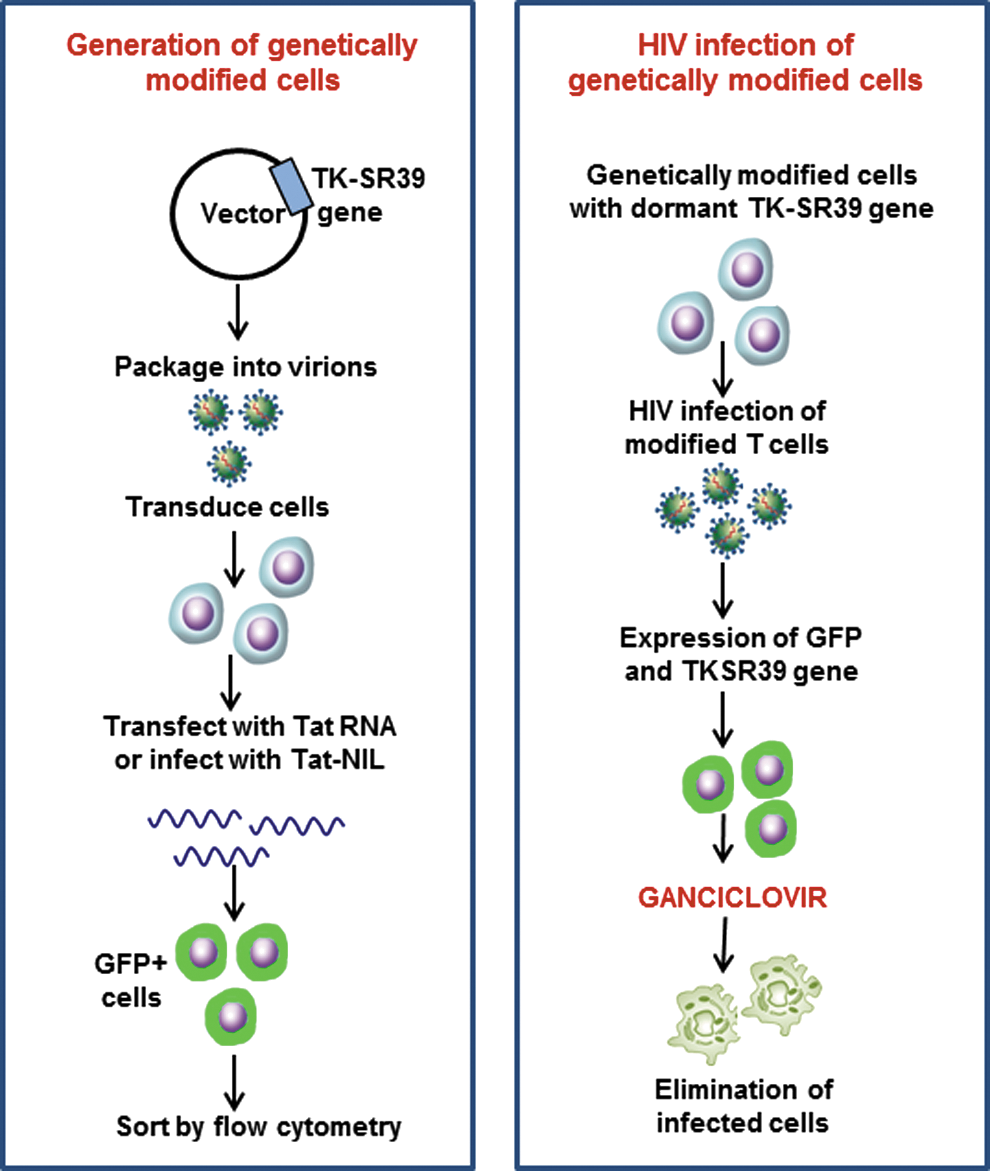
Overall schematic of strategy for generation of genetically modified stem cells, using the TK-SR39 lentiviral vector, and the outcome of genetically modified cells on HIV infection. The conditional cytotoxic lentiviral vector pNL-GFPRRESA-TK-SRR9 expresses the TK gene only in the presence of HIV Tat. On Tat expression, the cells also express GFP. The strategy can be exploited in vitro for sorting/enrichment of genetically modified cells. During HIV infection, Tat is provided by the virus leading to TK-SR39 gene expression. When treated with GCV, the TK-expressing, HIV-infected cells are marked for elimination by apoptosis. Color images available online at
Footnotes
Acknowledgments
The authors thank Dr. Margaret Black (Washington State University) for providing the TK-WT and TK-SR39 plasmids, and Dr. Yuntao Wu (George Mason University) for providing the pNL-GFPRRESA vector. The authors also thank the NIH AIDS Reference and Reagent Program for providing the valuable reagents essential for this study. This study was supported in part by NIH R03 grant 1R03AI095040-01A1 (A.J.) and the Texas Tech University's intramural seed grant program.
Author Disclosure
Texas Tech University had filed a patent application on behalf of H.G. and A.J., covering this technology.