
Abstract
Adenovirus (Ad) infection in humans is associated with inflammatory responses and thrombocytopenia. Although several studies were conducted in mice models to understand molecular and cellular mechanisms of Ad-induced inflammatory responses, only few of them turned their interest toward the mechanisms of Ad-induced thrombocytopenia. Using different depletion methods, the present study ruled out any significant role of spleen, macrophages, and vitamin K-dependent factor in Ad-induced thrombocytopenia. Interestingly, mice displaying thrombocytopenia expressed high levels of cytokines/chemokines after Ad administration. Most importantly, pseudotyping adenovirus with the fiber protein from other serotypes was associated with reduction of both cytokine/chemokine production and thrombocytopenia. Altogether, our results suggest that capsid fiber protein (and more precisely its shaft) of Ad serotype 5 triggers the cytokine production that leads to Ad-induced thrombocytopenia.
Introduction
A
Direct vector administration into the bloodstream is associated with several toxicities and acute responses. For example, a strong hepatic tropism leads to liver toxicity and damages. 3,4 The liver transduction was linked to the ability of Ad hexon protein to interact with coagulation factor X (FX), allowing subsequent virus binding to heparan sulfate proteoglycans expressed on hepatocytes. 5 –7 Interestingly, several studies have reported that mutations of the main capsid components or their replacement by proteins from other Ad serotypes also result in reduced hepatocyte transduction. 8 –12 Besides liver toxicity, the triggering of an acute inflammatory response and cytokine production within hours after virus administration represents one of the major hurdles to Ad gene therapy. 6
Once into the bloodstream, Ad can interact with many circulating blood cells, including human erythrocytes, 13 peripheral blood mononuclear cells, 14 neutrophils, 15 dendritic cells, 16 and platelets. 17 Particularly, an important decrease in platelet count known as thrombocytopenia was associated with Ad infection, both in preclinical studies 18 and in clinical trials. 19 –21 How Ad trigger thrombocytopenia is still poorly understood. Some studies have demonstrated that Ad can bind directly to platelets and trigger their activation and aggregation both in vitro and in vivo. 17,22 In contrast, another study reported that Ad is not able to interfere with or potentiate platelet aggregation. 23 The involvement of Ad primary receptor CAR and integrins in platelet interaction is also controversial. Indeed, two conflicting results were obtained regarding CAR expression on human platelets. 22,24 Finally, although different integrin ligands were able to block Ad binding to platelets in vitro, 22 a recent study questioned the role of integrins because blocking αIIbβ3 and αvβ3 integrins does not modify Ad binding and activation of platelets. 25 Studies of Ad-induced thrombocytopenia in different knockout (KO) mouse models helped to identify the role of specific soluble host factors. For example, Ad administration in Von Willebrand factor (VWF) KO mice did not trigger a significant thrombocytopenia compared with wild-type (wt) mice, thereby suggesting a key role of VWF in Ad-associated thrombocytopenia. 26 Also, the lack of thrombocytopenia in mice deficient for C3 and factor B proteins suggests a role for these complement factors in this phenomenon. 27 Finally, both genetic and acquired (pretreatment with neutralizing antibodies) deficiencies in TNF-α improved Ad-induced thrombocytopenia, suggesting a role for TNF-α pathway in this phenomenon. 28
In the present study, we seek to go deeper in the understanding of viral and host factors at the origin of Ad-induced thrombocytopenia. Most strikingly, we report a role of Ad5 fiber protein in Ad-induced thrombocytopenia.
Materials and Methods
Adenovirus
Adwt and AdF3 were previously described 9 and derived from Ad5 with E1 and E3 regions deleted. AdS3K5 was obtained by replacing Ad5 shaft (amino acid residues 47–400) by Ad3 shaft (amino acid residues 46–132). All viruses contain an expression cassette with the promoter/enhancer from the immediate early gene of human cytomegalovirus (CMV), the Escherichia coli lacZ gene with a nuclear localization signal, and the SV40 late polyA signal. eGFP recombinant and E1-deleted serotype 3 (Ad3) was described previously. 29 Luciferase recombinant Ad pseudotyped with fiber from serotype 35 (AdF35) was described previously. 30 All viruses were obtained using standard procedures as described, 31 stored at −80°C in PBS and 7% glycerol, and titrated by spectrophotometry (1 OD260 = 1.1 × 1012 viral particles [vp]/ml).
In vivo experiments
All animal experimental procedures were conducted in accordance with the institutional guidelines of Gustave Roussy Institutional Animal Care and Use Committee. Six- to eight-week-old female C57BL/6 (Harlan, France) mice were intravenously (i.v.) injected into the retro-orbital plexus with 5 × 1010, 1011, or 2 × 1011 vp of corresponding vectors. Blood from the retro-orbital plexus of mice was collected in citrated tubes at the indicated time points after virus administration. Platelets were counted using an automated counter of blood cells (MS9; Schloessing Melet).
For mice splenectomy, C57BL/6 mice were anesthetized and the lateral area of the animal left side was shaved. A 3–4 cm incision was made and the spleen was removed from the visceral cavity and the blood vessels were cauterized. Five days later, mice were i.v. injected with Ad and platelets were counted 2 days later as described above.
Kupffer cell and coagulation factor depletion
Two approaches were used to induce Kupffer cells (KC) depletion. The predosing method consists of injection of 3 × 1010 vp of AdGFP 4 hr before vector administration. 32 KC depletion was alternatively induced with intravenous administration of 200 μl of clodronate-liposomes or PBS-liposomes as negative control 24 hr before Ad administration. Transient KC elimination was confirmed by liver section immunolabelling with F4/80 antibody. For coagulation factor depletion, mice were injected subcutaneously with 133 μg of warfarin in 100 μl of PBS at 3 and 1 day before Ad delivery as previously described. 5
Cytokine and chemokine measurement
Spleens harvested 1 hr after intravenous Ad administration (1011 vp) were homogenized in sample buffer according to manufacturer's instructions. Lysates were incubated overnight with membranes from Proteome Profiler Antibody array (Mouse Cytokine Array Panel A; R&D System) according to manufacturer's instructions. Membranes were developed with HRP substrate and revealed using ChemiDoc MP System (BioRad).
Interleukin-6 measurement
Blood samples were collected 6 hr after Ad administration and serum interleukin-6 (IL-6) was quantified using a Duo-Set ELISA assay according to manufacturer's instructions (R&D).
FX measurement
Endogenous murine FX activity was determined by an amidolytic assay using the Russel's Viper Venom-X enzyme (RVV-X). In this assay, FX activation into FXa by the RVV-X leads to the cleavage of the chromogenic substrate S-2765. Briefly, plasma samples or a pool of plasma from wt C57Bl/6 mice (used as standard) were diluted in TBS/BSA (0.1%)/CaCl2 (5 mM) and incubated with 0.5 μM RVV-X enzyme (Kordia Life Science) 5 min at 37°C. The chromogenic substrate S-2765 (Chromogenix) was added at 0.2 mM and the kinetic reading at 405 nm was immediately started with a reading every 30 sec over 20 min. The V max (maximum speed of chromogenic conversion) was calculated for the linear part of the kinetic curve and plasma sample values were normalized to the standard. FX activity was expressed in units/ml.
Statistical analysis
Results were expressed as means ± SEM. ANOVA analysis followed by Tukey's post-hoc test was used to compare sets of data. Variations were considered statistically significant when p < 0.05. All graphs and statistical tests were performed using GraphPad Prism software.
Results and Discussion
Kinetics and dose dependency of Ad-induced thrombocytopenia
First, we sought to determine the kinetics of thrombocytopenia induction in response to Ad administration. C57BL/6 mice were i.v. injected with saline or two different doses of Ad (1011 and 2 × 1011 vp), and platelet counts were determined every day during the first week and at day 15 postinfection (p.i.) (Fig. 1). Ad injection caused a rapid decrease in platelet count within 24 hr p.i. with a nadir at 48 hr p.i. Of note, the reduction in platelet count was more severe with the higher viral dose, demonstrating the dose dependency of this phenomenon (Fig. 1). However, the thrombocytopenia was transient and mice recovered a normal platelet count by day 5 p.i. The kinetic of thrombocytopenia induction and the dose dependency observed in our study are in accordance with previous results obtained by others in mice 33 but also in rabbits, 33 baboons, 34 and rhesus monkeys. 18
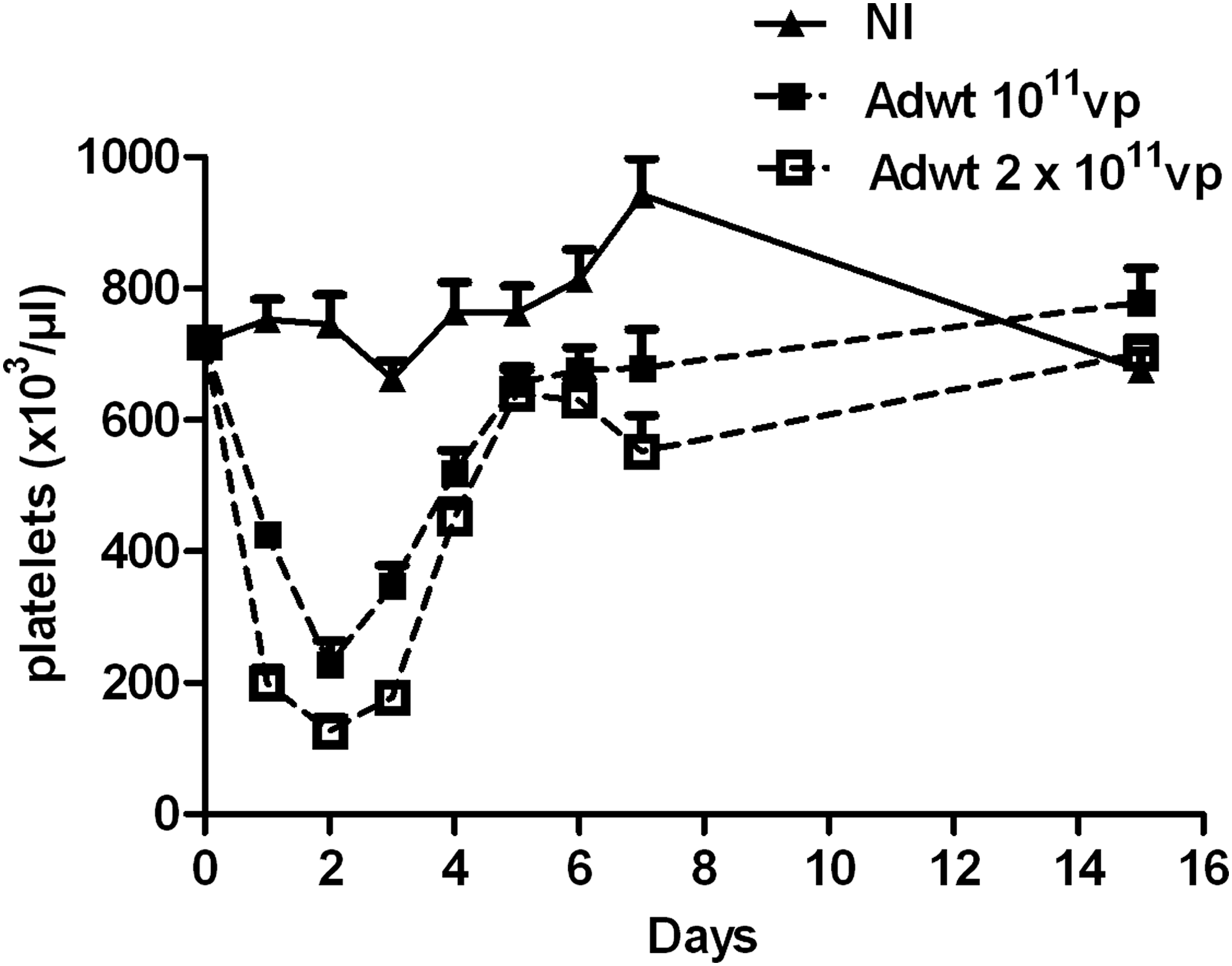
Kinetics of Ad-induced thrombocytopenia. Mice were injected intravenously with PBS or with different doses (1011 and 2 × 1011 vp) of Adwt. Platelet counts were determined at different time points thereafter. n = 5 mice; means + SEM. Ad, adenovirus; Adwt, Ad bearing a wild-type capsid.
Blood coagulation factors and spleen are not involved in Ad-induced thrombocytopenia
Coagulation factors and platelet activation are complementary and mutually dependent processes participating in hemostasis. 35 Because Ad is able to interact with different blood factors such as coagulation factor IX and X (FX), 5 we evaluated whether these factors play a role in Ad-induced thrombocytopenia. To answer this question, C57BL/6 mice were pretreated with warfarin or saline at days 3 and 1 before Ad administration, as described previously. 5 Blood factor depletion was confirmed by the measurement of FX activity in mice sera harvested before virus delivery (12.8 ± 3.2 in warfarin-treated mice vs. 187.9 ± 54.7 in PBS-treated mice). After Ad administration of 2 × 1011 (Fig. 2a) or 1011 vp (Fig. 2b), no statistically significant difference in platelet counts was observed between saline- or warfarin-pretreated mice. These results ruled out any significant role of coagulation factors in Ad-induced thrombocytopenia and are in accordance with the fact that hexon-modified Ad that are disabled in FX interaction triggered thrombocytopenia as efficiently as their wt counterparts. 8 Then, we analyzed the spleen involvement in Ad-induced thrombocytopenia. Indeed, the spleen is well-known as a site of platelet storage able to release them upon request. 36 In addition, this organ removes old platelets from the bloodstream. 37 To assess the role of the spleen in Ad-induced thrombocytopenia, C57BL/6 mice were splenectomized or sham-treated 5 days before Ad systemic administration, and platelet counts were determined 48 hr p.i. Both splenectomized and nonsplectomized mice displayed a similar reduction in platelet numbers in response to Ad administration (Fig. 2c). In parallel, because the spleen can store excessive numbers of platelets leading to splenomegaly, 37 we carefully examined the size of the spleens of nonsplectomized mice and did not observe signs of splenomegaly at day 2 after Ad administration (data not shown). Taken together, these results demonstrated no significant role of the spleen in Ad-induced thrombocytopenia. In contrast, Hofherr et al. reported that splenectomy protected mice from Ad-induced thrombocytopenia. 3 The discrepancy between our results and Hofherr et al. could be because of differences in mice strains (C57BL/6 in our study and outbreed ICR mice in the study of Hofherr et al.) and viral doses used.
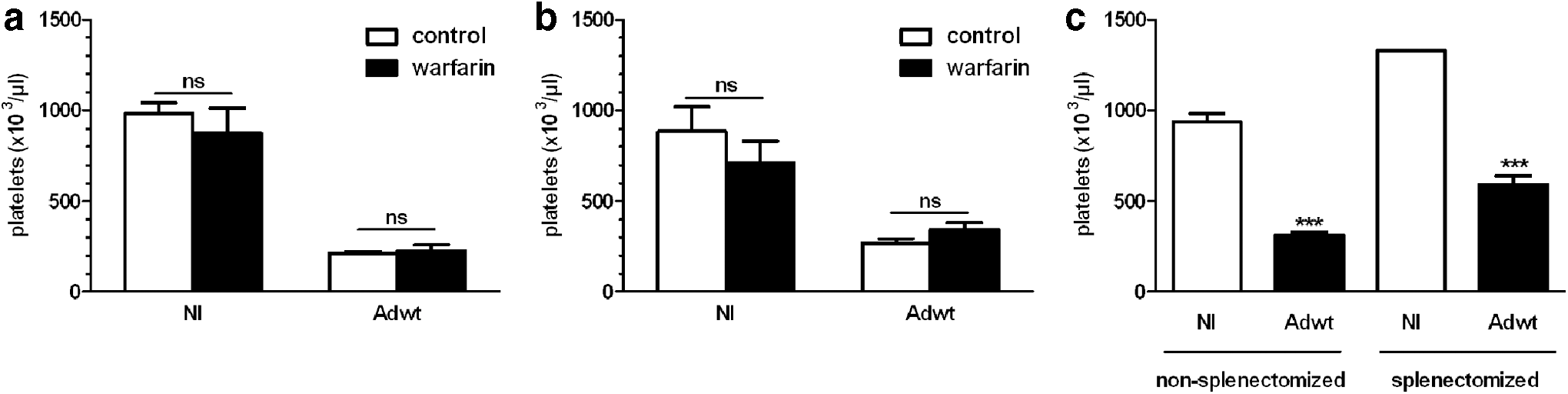
Coagulation factor and spleen involvement in Ad-induced thrombocytopenia. Mice were mock-treated (noninfected, NI) or treated with warfarin before injection of 2 × 1011
Requirement of the fiber shaft in Ad5-induced thrombocytopenia
To address the question of viral proteins involved in Ad-induced thrombocytopenia, we focused on capsid proteins known to participate in Ad infection process. These proteins correspond to hexon, penton base, and fiber proteins able to bind to coagulation FX, integrins, CAR and heparan sulfates, respectively. First, we ruled out a role for hexon–FX interaction in Ad-induced thrombocytopenia because we reported previously that impairing hexon binding to FX has no impact on platelet counts.
8
Second, previous data from our laboratory based on the use of Ad disabled in integrin interaction ruled out any role for the penton base protein in Ad-induced thrombocytopenia (data not shown). Then, taking advantage of the two already described serotype 5 recombinant Ad pseudotyped with the fiber from serotype 3 (Vigne et al.
9
and Raddi et al., unpublished manuscript), we investigated the role of the fiber protein. C57BL/6 mice were injected i.v. with a high (Fig. 3a), intermediate (Fig. 3b), or low (Supplementary Fig. S1; Supplementary Data are available online at
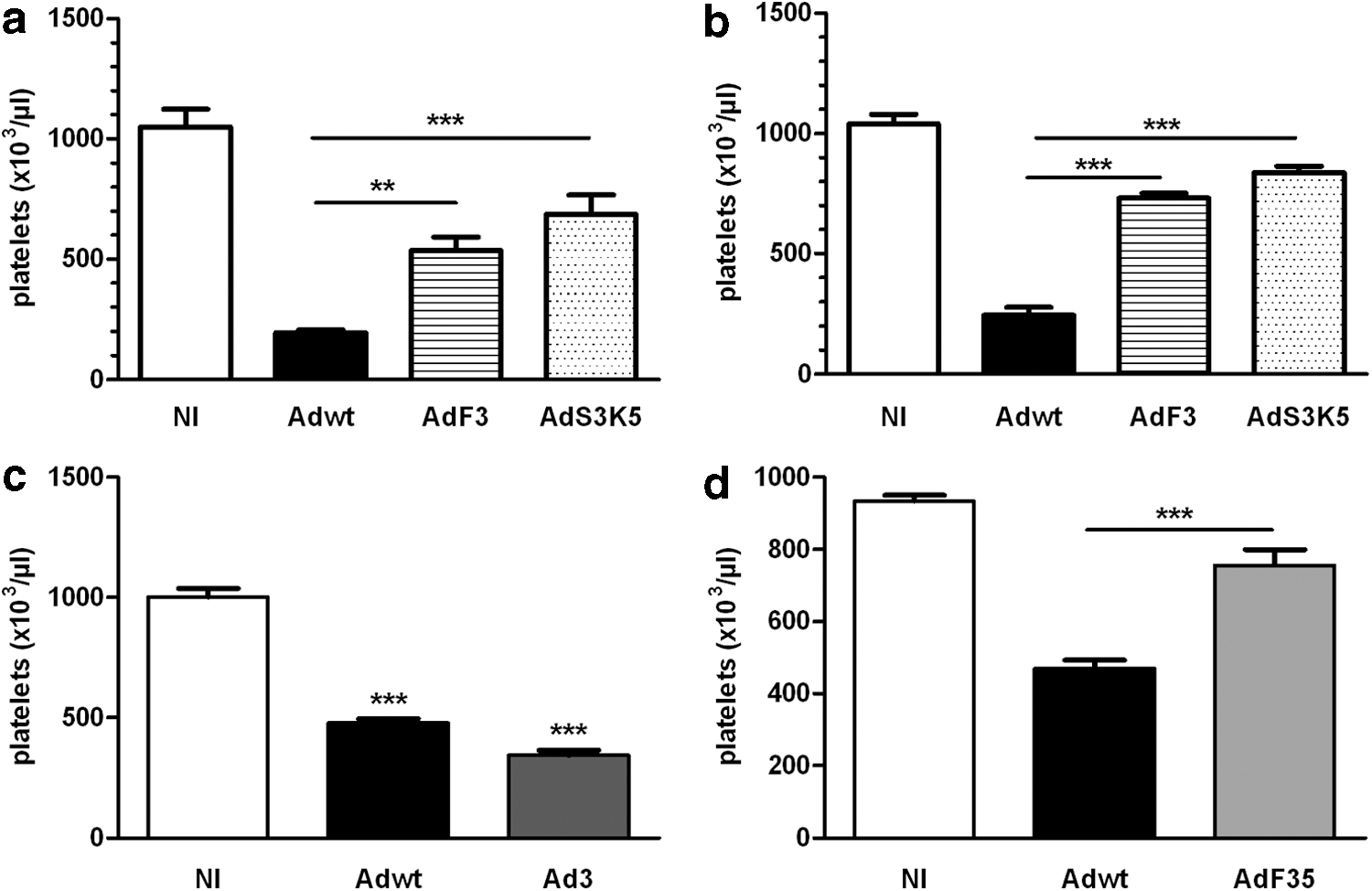
Thrombocytopenia induction by fiber-modified Ad5 and Ad3. Mice were intravenously injected with 2 × 1011
These results suggest that either Ad5 fiber was responsible for the induction of the thrombocytopenia or Ad3 fiber has an intrinsic property to mitigate thrombocytopenia. To decipher between these two hypotheses, C57BL/6 mice were injected with 1011 vp of Adwt or a recombinant Ad3. Figure 3c shows that Ad3 induces thrombocytopenia to the same extent as Adwt does, thus ruling out a modulatory role of Ad3 fiber. On the other hand, pseudotyping Ad5 with a fiber from serotype 35 (AdF35) strongly reduced virus-induced thrombocytopenia (Fig. 3d). Altogether, these results suggest that Ad5 fiber was involved in Ad-induced thrombocytopenia. More precisely, because AdS3K5 also displayed a reduced thrombocytopenia, this indicates a role for the Ad5 shaft in this phenomenon. The fact that thrombocytopenia was observed in clinical trials after administration of an Ad pseudotyped with a chimeric fiber containing Ad5 shaft and Ad3 knob further supports our conclusions. 38 However, it should be underlined that Ad displaying a targeting peptide within the HI loop also led to a reduced thrombocytopenia. 39 The discrepancy between our results (role of the shaft) and their results (role of the knob) may be linked to the different strains of mice (C57BL/6 and BALB/c, respectively) used or alternatively to the targeting ligand used in Coughlan et al. 39
Role of macrophages in Ad5-induced thrombocytopenia
Different pathogens are able to interact with platelets, leading to their uptake by liver KC. 40 Whether or not Ad interact with platelets 17,22,23 and whether KC play any role in platelet clearance during virus infection 17,41 are still a matter of debate. Therefore, we investigated the potential role of KC in the development of Ad-induced thrombocytopenia using two depletion methods: predosing, 32 which consists of preadministration of a low dose of Ad (3 × 1010 vp) able to specifically eliminate KC, the resident liver macrophages, or pretreatment with clodronate-liposomes that depletes any macrophages. 42 Regardless of the depletion method, comparison of platelet counts in control (Fig. 4a and c) and in macrophage-depleted mice (Fig. 4b and d) did not show any significant modification of Adwt-induced thrombocytopenia. However, both methods of depletion were able to abrogate the modest reduction in platelet counts observed after intravenous AdF3 or AdS3K5 delivery. Thus, although macrophages do not appear to play a significant role in Adwt-induced thrombocytopenia, they may play a role in the modest thrombocytopenia triggered by Ad pseudotyped with Ad3 fiber.

Role of Kupffer cells (KC) in Ad-induced thrombocytopenia. Mice were pretreated with or without Ad (predosing,
Reduced inflammatory responses triggered by Ad5 pseudotyped with the fiber from other serotypes
Different studies have proposed that cytokine production participates to Ad-induced thrombocytopenia. 28,43,44 Therefore, we analyzed whether pseutotyping Ad5 with the fiber of serotype 3 modifies cytokine and chemokine production in response to intravenous Ad delivery. First, using antibody arrays, we measured cytokine and chemokine production in the spleen harvested 1 hr p.i. (Fig. 5a). Compared with PBS-injected mice, Adwt-injected mice displayed a strong production of cytokines (IL-1β and IL-6), chemokines (IP-10, KC, CCL2, CCL3, and CXCL2), as well as complement factor C5a and triggered receptor expressed on myeloid cells-1 (TREM-1). Interestingly, AdF3 and AdS3K5 induced a weak production of these cytokines and chemokines in the spleen, comparable to the one observed in the spleen from PBS-injected mice (Fig. 5a). Using ELISA, we observed a strong dose-dependent release of IL-6 in the blood 6 hr after Adwt administration. However, both AdF3- and AdS3K5-injected mice displayed a dramatic reduction in IL-6 production (Fig. 5b), at both viral doses. To exclude a putative inhibitory effect of Ad3 fiber (and more precisely Ad3 shaft) on inflammatory responses, C57BL/6 mice were injected with Adwt or Ad3. As shown in Fig. 5c, compared with Adwt, Ad3 led to a 20-fold higher production of serum IL-6 (Fig. 5c), ruling out a role for Ad3 fiber in decreasing cytokine and chemokine production. The fact that AdF3 and AdS3K5, on the one hand, and Ad3, on the other hand, present opposite capabilities to trigger cytokine production and thrombocytopenia is puzzling. One hypothesis would be that Ad3 shaft alone does not possess the intrinsic property to trigger cytokine production and thrombocytopenia but needs other Ad3 capsomers. An alternative hypothesis would be that insertion of fiber from serotype 3 in Ad5 backbone led to a particle trafficking different from Ad5, thus impairing cytokine production. Interestingly, intravenous administration of AdF35 led to serum IL-6 levels comparable to noninfected mice (Fig. 5d). Taken as a whole, our results argue for a role of Ad5 shaft in cytokine and chemokine production. These results are further supported by a previous work showing that pseudotyping Ad5 with fiber from serotypes 11 or 35 reduced IL-6 and TNF-α production. 45
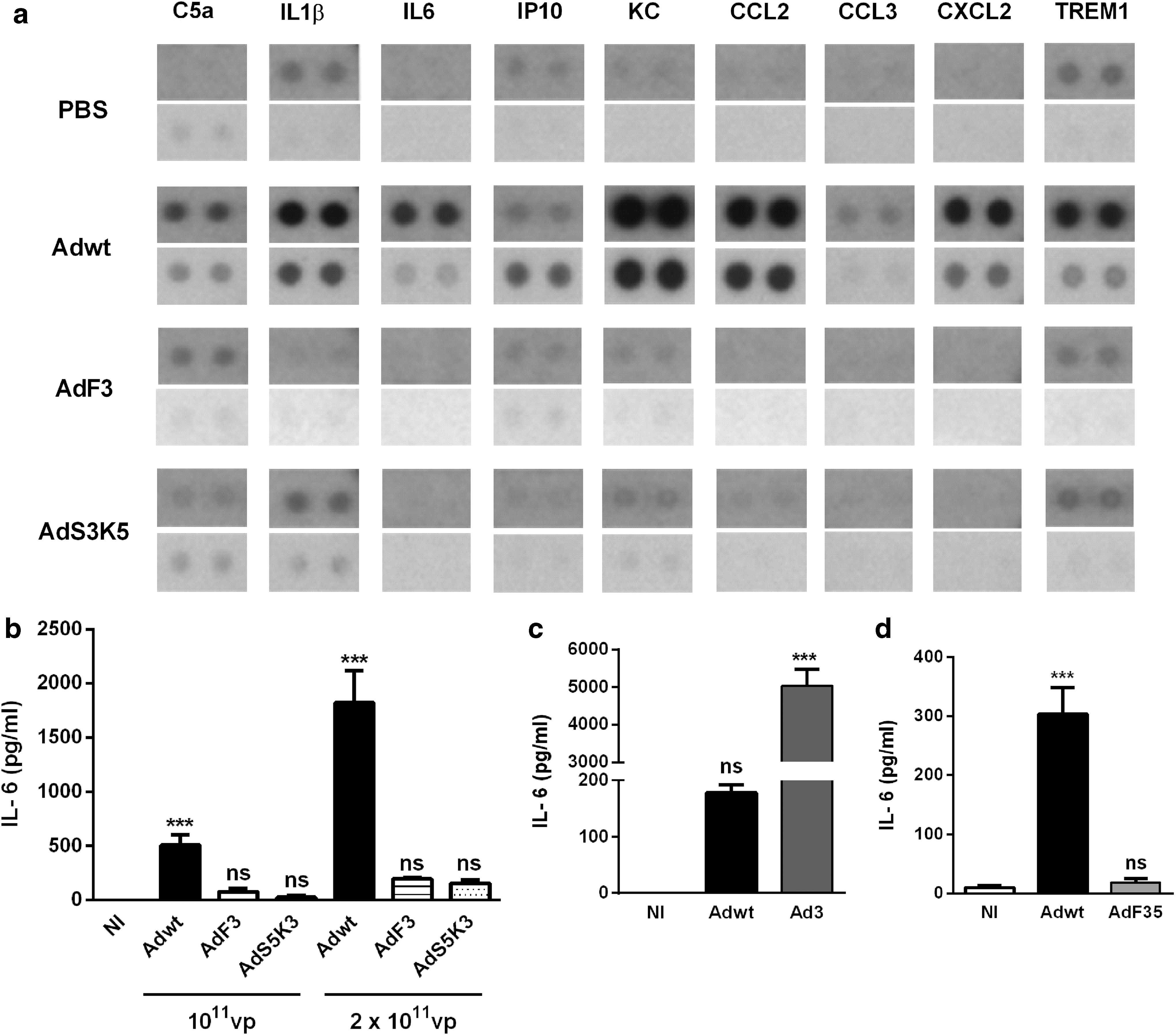
Cytokine and chemokine production in response to Ad injection. Mice were intravenously injected with 1011 vp.
Ad-induced thrombocytopenia is associated with cytokine production
As stated above, Ad was reported by different teams 46 to trigger inflammatory cytokines production as well as thrombocytopenia. Our present results uncover that pseudotyping Ad5 with fibers from other serotypes (Ad3 or Ad35) both reduces cytokine production (Fig. 5) and ameliorates the thrombocytopenia (Fig. 3). This suggests an unanticipated link between cytokine production and thrombocytopenia. Interestingly, a point-by-point comparison of IL-6 production and platelet counts in individual mice demonstrated that Adwt-injected mice displayed both high levels of serum IL-6 and a reduction in platelet counts, whereas mice injected with pseudotyped Ad exhibit reduced level of IL-6 and high platelet counts (Fig. 6a and b). However, our previous results showed that thrombocytopenia occurred even in Ad-injected IL-6-deficient mice. 47 Taken together, these results point out that IL-6 itself is not mandatory to this phenomenon and suggests a role for other cytokines.
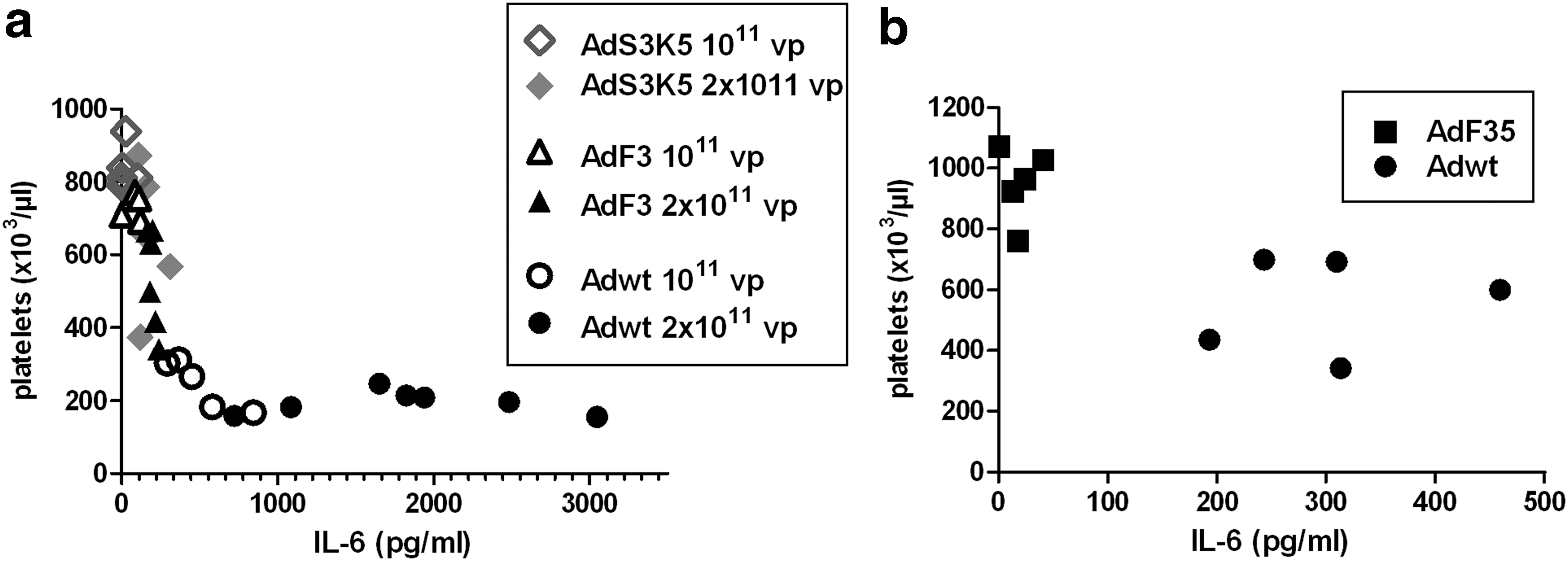
Relationship between IL-6 levels in mice sera and platelet counts.
To summarize, taking advantage of various vectors (serotypes or pseudotyped Ad), we demonstrated a role for Ad5-fiber protein and more precisely for its shaft in Ad5-induced thrombocytopenia. We also reported that coagulation factors, spleen, and macrophages were dispensable in this phenomenon. In parallel, our results suggest that Ad5 fiber—more precisely Ad5 shaft—plays a key role in induction of chemokines and cytokines. Interestingly, we draw a parallel between cytokine/chemokine production and thrombocytopenia. Indeed, using different Ads, our results show that Ads able to trigger high levels of cytokines do elicit strong thrombocytopenia. Mice injected with AdF3 and AdS3K5 displayed a reduction in C5a production (Fig. 5a), suggesting a reduced activation of the complement cascade. Since the complement activation was proposed by others to be involved in thrombocytopenia, 48 the low C5a production in the spleen of AdF3- and AdS3K5-injected mice could explain their inability to induce thrombocytopenia. Further experiments are needed to better understand the link between cytokine production, complement factors, and thrombocytopenia. By showing the key role of the Ad5 fiber protein in both cytokine production and thrombocytopenia, this study paves the way to a better understanding of molecular and cellular mechanisms involved in Ad-induced thrombocytopenia. Previous results of our laboratory have shown that fiber-pseudotyped Ad led to a reduced liver transduction upon intravenous delivery but to efficient tumor gene transfer upon direct intratumoral administration (Raddi et al. unpublished manuscript). Altogether, the results underline that fiber-pseudotyped Ad that elicit low cytokine/chemokine production and reduced thrombocytopenia constitute tools of valuable interest for safer gene delivery in tumors.
Footnotes
Acknowledgments
We are very grateful to the staff of the animal facility of Gustave Roussy for technical help. We also thank Olivier Christophe and Mohamad Kurdi for FX measurement. This work was supported by Centre National de la Recherche Scientifique, University Paris-Sud, INCa (PIMRANIS project), and EDF. N.R and F.V. received fellowships from the Ministère de la Recherche et de la Technologie. N.R. received an additional fellowship from University Paris-Sud, and F.V. additional fellowships from the AFM and the Société Française d'Hématologie.
Author Disclosure
The authors have no competitive financial interests to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.