
Abstract
Allergic airway inflammation driven by T helper 2 (Th2)-type immunity is characterized by airway hyperresponsiveness, eosinophilic infiltration, and elevated IgE production. Various novel strategies for managing asthma have been explored, such as DNA vaccines, T-cell peptides, and allergen-specific immunotherapy. A principal goal of most immunotherapeutic approaches is active and long-term allergen-specific tolerance. Liver-specific gene transfer using adeno-associated virus (AAV) has been shown to favorably induce tolerogenic responses to therapeutic products in various experimental models. AAV8 has strong liver tropism and induces immune tolerance in mice. The present study aimed to determine whether hepatocyte-specific allergen expression by pseudotyped AAV2/8 alleviates asthmatic symptoms in ovalbumin (OVA)-sensitized mice. Mice were intravenously injected with AAV2/8 vector carrying membrane-bound OVA transgene under transcriptional control of a hepatocyte-specific alpha 1 antitrypsin promoter (AAV2/8-OVA) and then sensitized with OVA. AAV2/8-OVA specifically transduced the OVA transgene in the liver. Airway hyperresponsiveness, eosinophilia, mucus hypersecretion, and Th2 cytokines were significantly suppressed in both the lungs and secondary lymphoid organs of asthmatic mice infected with AAV2/8-OVA. Significant reduction of OVA-specific antibodies was detected in the bronchoalveolar lavage fluid from AAV2/8-OVA-treated mice. Moreover, AAV2/8-OVA treatment prominently promoted the expression of Foxp3, IL-10, and TGF-β in the liver. Enhanced Foxp3 expression was also detected in the lungs of asthmatic mice after AAV2/8-OVA treatment. Taken together, these results suggest that the induction of immune tolerance by hepatic AAV gene transfer may be beneficial for modulating allergic asthma.
Introduction
A
The liver preferentially facilitates local or systemic immune tolerance via unique antigen-presenting cells (APCs) and the cytokine milieu. 5 The advantages of sustained transgene expression and the tolerogenic immunity of the liver are widely utilized and investigated for the treatment of hepatitis, hemophilia, and inherited metabolic disorders by gene therapy. 6 T-cell activation in the liver promotes tolerant immune responses by inducing anergy, clonal deletion, or regulatory T-cell (Treg) generation. 7 –9 Accumulated evidence indicates that in vivo hepatocyte-specific gene transfer contributes to the induction of immune tolerance and has therapeutic potential in mice. 6 Some reports have proposed that antigen-specific Foxp3+ Tregs are generated after hepatic gene transfer, which prevents or suppresses the development of experimental autoimmune encephalomyelitis (EAE). 7,10 Luth et al. confirmed that specific expression of neural autoantigen in hepatocytes, instead of other tissues, protects against neuron inflammation in EAE. 10 CD4+CD25+Foxp3+ Tregs mainly engage in negative control of autoimmune responses and allergic diseases, such as asthma. 11 However, whether hepatocyte-specific allergen expression modulates allergic airway inflammation has not yet been reported.
Adeno-associated virus (AAV), a single-stranded DNA virus with low immunogenicity, consists of numerous serotypes that target various tissues and establish long-term transgene expression. 12 AAV1, AAV5, AAV8, and AAV9 efficiently infect muscle, retina, liver, and lung tissue, respectively. 13 Recombinant AAV (rAAV) vectors derived from rep of AAV2 and cap of different serotypes would enhance gene transduction efficacy, change tissue tropism, and evade preexisting neutralizing antibodies. 13 Gene therapy with rAAV vectors has been employed for studies in animal models and clinical trials. Several reports have suggested that rAAV vectors carrying immunomodulatory transgenes, such as T-bet 14 and Clara cell 10 kDa protein (CC10), 15 or antisense sequences against Th2 cytokines 16 and chemokines, 17 attenuate allergen-induced inflammation in asthmatic models. Because of the potential tolerogenic immune responses induced by hepatocyte-specific gene expression, we investigated whether Th2 cell-driven inflammation in allergic asthma is modulated by rAAV carrying an allergen transgene targeting hepatocytes. As the AAV8 serotype had been shown to superiorly transduce liver tissue and lead to immune tolerance in mice, 18,19 we applied ovalbumin (OVA) as a model antigen and examined the immunomodulatory effects of AAV2/8 vector carrying a liver-specific membrane-bound OVA transgene (AAV2/8-OVA). Significant suppression of asthmatic symptoms was achieved in OVA-sensitized mice treated with AAV2/8-OVA. These results suggest that hepatocyte-specific allergen gene expression may be considered a novel approach for alleviating allergic asthma.
Materials and Methods
Construction and preparation of pseudotyped AAV2/8 vector
A recombinant AAV (rAAV) construct was created with the coding sequence of chicken OVA or enhanced green fluorescence protein (EGFP) as the vector control. The full-length OVA DNA fragment (386 amino acids) was derived from pAc-neo-OVA (gift from Dr. M.J. Bevan). 20 The OVA386 fragment was fused with 178 COOH terminal amino acids of B7-1 domain including the transmembrane domain by PCR to generate membrane-bound OVA. Next, the OVA/B7-1-fused fragment was subcloned into pAAV vector containing the inverted terminal repeats (ITRs) of AAV serotype 2 at both ends. The expression cassette comprised a hepatocyte-specific alpha 1 antitrypsin (hAAT) promoter, a 9-amino-acid influenza virus HA tag, and a polyadenylation signal sequence.
Recombinant AAV2/8 vectors were generated using a tri-plasmid system as described previously. 17 Briefly, the pAAV/hAAT plasmid with the OVA or EGFP transgene was co-transfected into HEK 293T cells with an adenovirus helper plasmid and a packaging chimeric plasmid encoding the AAV2 rep and AAV8 cap genes. Recombinant single-strand AAV vectors were purified by cesium chloride sedimentation. Genome copies (GCs) of AAV2/8-OVA or AAV2/8-EGFP were determined by quantitative PCR (qPCR) using SYBR Green Master mix (Roche Diagnostics GmbH).
Animals and OVA-sensitization protocol
Three-week-old female BALB/c mice were purchased from National Laboratory Animal Center (Taipei, Taiwan, ROC) and acclimatized for 1 week. Four-week-old mice were administered 1 × 1011 GCs per mouse in 200 μl of normal saline via intravenous injection. Four weeks after virus treatment and before the sensitization protocol, blood samples were collected under anesthesia from the retro-orbital venous plexus using heparinized capillary tubes. All mice (8 weeks old) were sensitized by intraperitoneal injection of 50 μg OVA (grade V; Sigma-Aldrich Inc.) mixed with 0.8 mg aluminum hydroxide (Pierce Biotechnology Inc.) on days 56, 57, 58, and 70 after virus administration. These mice were challenged by inhaling 2% aerosolized OVA for 20 min using an atomizer (Pulmo-aide; DeVilbiss Health Care) on days 70, 73, 77, 80, and 84. AHR was analyzed on day 85, and mice were sacrificed 24 hr later. The care and handling of mice in all experimental procedures was approved by the Institutional Animal Care Committee of Chang Gung University and complied with the National Institutes of Health (NIH) Guidelines for the Care and Use of Laboratory Animals.
Assessment of AHR and cell composition in bronchoalveolar lavage fluid
AHR was assessed on day 85, twenty-four hours after the last challenge, using methacholine (Mch; Sigma). Mice were exposed to normal saline or increasing concentrations of Mch (10, 20, 30, and 40 mg/ml) for 3 min and then underwent whole-body plethysmography while conscious. The enhanced pause (Penh) value was measured for 3 min, and pulmonary airway obstruction analyzed by BioSystem XA software (Buxco Electronics, Inc.). As described in detail in our previous reports, the sensitized mice had significantly enhanced Penh values compared with the normal healthy control mice. 21,22
After the measurement of AHR, the tracheas of mice from different groups were cannulated and the lungs lavaged three times with 1 ml phosphate-buffer saline (PBS) supplemented with 0.1 mM EDTA. Cells in the bronchoalveolar lavage fluid (BALF) were collected and counted using trypan blue dye exclusion. For differential cell counts, BALF cells were centrifuged (Shandon Cytospin 4; Thermo Scientific) and stained with Liu staining solution. Different cell types were distinguished under light microscopy by counting at least 200 cells based on their morphological profile.
Lung histology
On day 86, nonlavaged lung tissues were removed and fixed with 4% formaldehyde for 24 hours at 4°C for histological examination. After dehydration, the tissues were embedded in paraffin and sliced into 4 μm sections. The tissue sections were subjected to hematoxylin and eosin (H&E) staining and periodic acid-Schiff (PAS; Sigma) staining. Inflammatory cell infiltration and goblet cell hyperplasia were examined by automated MetaMorph microscopy and image analysis software (Molecular Devices).
Measurement of OVA-specific IgE
The titers of OVA-specific IgE antibodies in the BALF and serum were determined by ELISA. Briefly, the plates (Costar) were coated with 10 μg/ml OVA at 37°C for 1 hr. The OVA-coated plates were incubated with diluted BALF or serum samples at 37°C for 1 hr. Next, biotinylated rat antimouse IgE monoclonal antibodies (BD PharMingen) and streptavidin-conjugated horseradish peroxidase (HRP; BD PharMingen) were sequentially added to the plates. Finally, the plates were incubated with substrate solution (TMB; BD Biosciences) for 20 min and the reaction was stopped with 2N H2SO4. Absorbance was measured on an ELISA reader at 450 nm (SpectraMax M2; Molecular Devices).
Cell culture and cytokine assays
Spleens and mediastinal lymph nodes (mLNs) were removed and single-cell suspensions prepared. Cells (5 × 106 cells/ml) were cultured in RPMI 1640 medium (Gibco, Life Technologies) containing 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin (Gibco) with 100 μg/ml OVA. Culture supernatants were harvested on day 6 and the levels of interleukin (IL)-4, IL-13 (DuoSet ELISA kit; R&D Systems), and IL-5 (BD OptEIA set; BD Pharmingen) determined by ELISA according to the manufacturer's instructions.
RNA isolation and qPCR
Total RNA was extracted from homogenized lung and liver tissues using TRIzol reagent (Invitrogen, Life Technologies) and treated with DNase I (Fermentas; Thermo Scientific). The cDNA were generated using random primers and M-MLV reverse transcriptase (Invitrogen). The sequences of specific primers for qPCR of CD4 and CCL22 are listed in Supplementary Table S1 (Supplementary Data are available online at
Statistical analysis
Results are presented as mean ± SEM. Significance was assessed using a two-tailed p-value calculated by the Mann–Whitney U test. p < 0.05 was considered significant. All graphs were generated and statistical analyses performed using GraphPad Prism 5.0 software.
Results
Liver-specific OVA expression by AAV2/8 vector virus suppressed AHR and OVA-specific IgE in OVA-sensitized mice
We constructed DNA encoding the membrane-bound OVA sequence using the rAAV2/8 vector with hepatocyte-specific alpha 1 antitrypsin promoter (Fig. 1a) to achieve hepatocyte-specific OVA expression. The expression of OVA on the cell surface was confirmed in pAAV/hAAT-OVA-transfected Huh7 cells (Supplementary Fig. S1a). Furthermore, mice infected with AAV2/8-OVA exhibited significant OVA transgene expression in the liver but not in the lung (Supplementary Fig. S1b). To examine whether hepatic OVA expression alleviates the development of airway inflammation in vivo, mice were infected with AAV2/8-OVA or AAV2/8-EGFP control viruses and then sensitized with OVA (Fig. 1b). After the final challenge, AHR was assessed in response to increasing concentrations of inhaled methacholine (Mch). When mice were stimulated with 40 mg/ml Mch, the enhanced pause (Penh) value was significantly lower in AAV2/8-OVA-treated mice than in the nontreatment and AAV2/8-EGFP control groups (Fig. 1c) and was comparable to healthy normal controls in our previous studies. 21,22 Decreased Penh values were also measured in the AAV2/8-OVA versus AAV2/8-EGFP control groups when mice were stimulated with 30 mg/ml Mch.
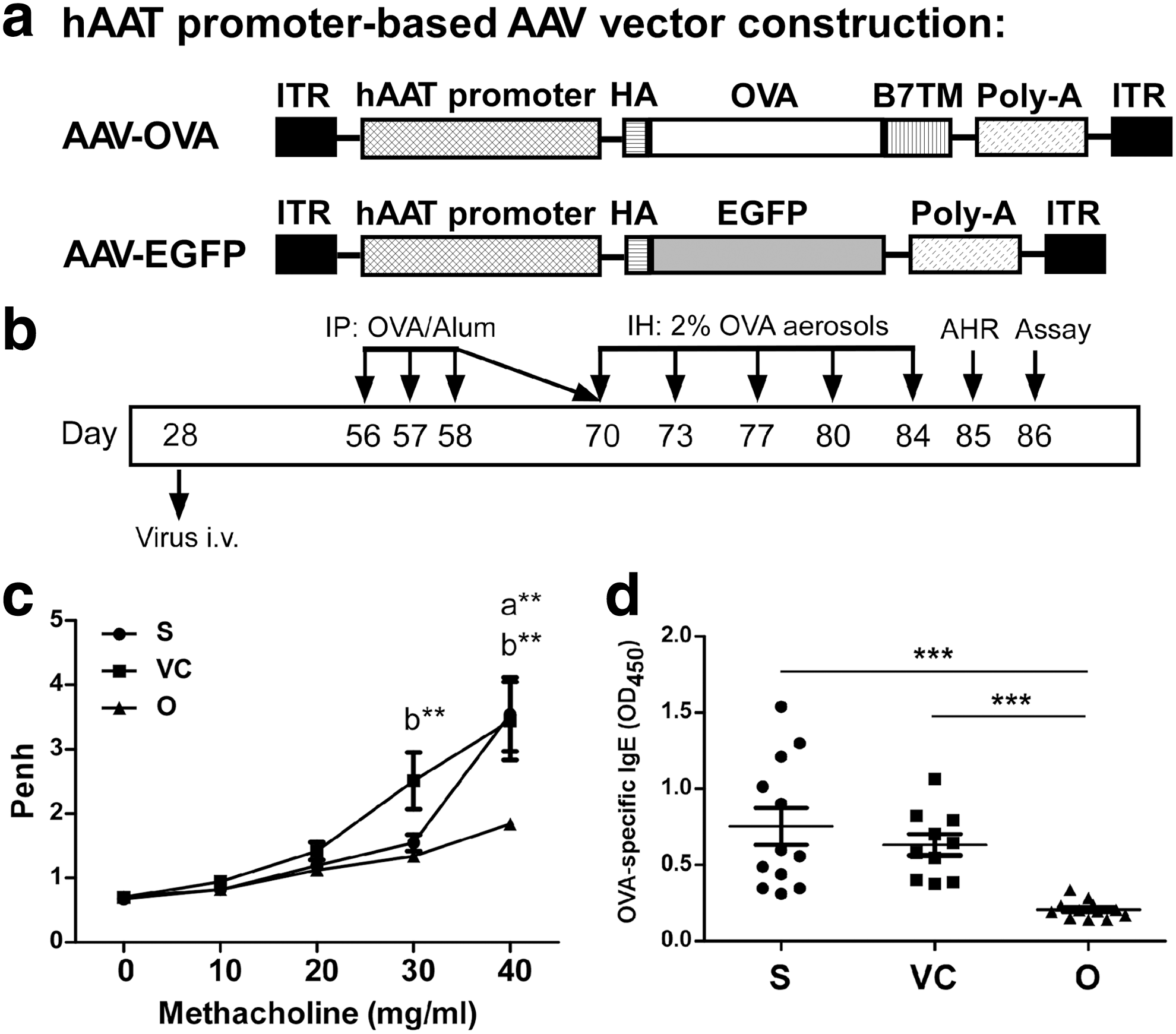
AAV2/8-OVA ameliorated airway hyper-responsiveness (AHR) and inhibited OVA-specific antibody secretion in OVA-sensitized mice.
A significant reduction in OVA-specific IgG1, IgG2a (Supplementary Fig. S2), and IgE (Fig. 1d) was detected in the BALF from OVA-sensitized mice treated with AAV2/8-OVA compared with the nontreatment and AAV2/8-EGFP control groups. The data indicate that administration of AAV2/8-OVA attenuated AHR and inhibited OVA-specific antibody production in the lungs of OVA-sensitized mice.
AAV2/8-OVA significantly decreased cell infiltration and mucus hypersecretion in the lungs of OVA-sensitized mice
Because the infiltration of eosinophils and lymphocytes into lung tissue is one of the main characteristics of asthmatic patients, we analyzed the inflammatory cell composition of BALF from OVA-sensitized mice. A markedly decreased percentage of eosinophils was found in mice treated with AAV2/8-OVA compared with the nontreatment and AAV2/8-EGFP control groups (Fig. 2a). The absolute numbers of total leukocytes, eosinophils, and lymphocytes were significantly lower in the BALF from AAV2/8-OVA-treated mice than that from the two control groups (Fig. 2b–d). Therefore, AAV2/8-OVA reduced the infiltration of inflammatory cells into the lungs of OVA-sensitized mice.

AAV2/8-OVA reduced the infiltration of eosinophils and lymphocytes in the BALF from OVA-sensitized mice. Lung tissues were lavaged with PBS, and
We further analyzed whether AAV2/8-OVA can suppress cell infiltration and mucus secretion in the airways based on histological examination. Massive inflammatory cells infiltrated peribronchial and perivascular sites in the lungs of untreated and AAV2/8-EGFP control mice, whereas the number of inflammatory cells was much lower in AAV2/8-OVA-treated mice (Fig. 3a). Quantitative analysis demonstrated a significant reduction in inflammatory cells in the lungs of AAV2/8-OVA-treated mice compared with the nontreatment and AAV2/8-EGFP control groups (Fig. 3c). Similarly, decreased goblet cell hyperplasia and mucus production were observed in the airways of AAV2/8-OVA-treated mice compared with the two control groups (Fig. 3b and d). To confirm this result, gob5, a molecule associated with mucin gene regulation and goblet cell hyperplasia, and Muc5ac expression were analyzed by qPCR (Fig. 3e and f). Mice treated with AAV2/8-OVA expressed lower levels of gob5 and Muc5ac than mice in the nontreatment and AAV2/8-EGFP control groups. Thus, AAV2/8-OVA administration was able to attenuate the accumulation of inflammatory cells and decrease mucus production in the lungs of asthmatic mice.

Lung histopathology of OVA-sensitized mice treated with AAV2/8-OVA. Lung tissues were harvested and fixed in formaldehyde at the end of the sensitization protocol. Paraffin-embedded lung tissues were sliced and subjected to
Significant reduction of Th2 cytokine and chemokine gene expression in the lungs of AAV2/8-OVA-treated mice
Because of the substantial suppressive effect of AAV2/8-OVA on lung inflammation, we analyzed the expression levels of cytokine and chemokine genes in the lungs. OVA-sensitized mice treated with AAV2/8-OVA expressed significantly lower levels of Th2 cytokines compared with untreated and AAV2/8-EGFP control mice (Fig. 4a). However, the RNA expression of Th1-related cytokines, such as IFN-γ, IL-12p35, and IL-12p40, was unchanged (Fig. 4a). To determine whether the decreased number of lymphocytes in the lungs is attributed to the abolishment of Th2 cell recruitment, we measured CCL17 and CCL22, which selectively induce the chemotaxis of CD4+ Th2 cells into inflamed tissues in allergic diseases. The lungs of untreated and AAV2/8-EGFP control mice expressed large amounts of CCL17 and CCL22 compared with mice receiving AAV2/8-OVA (Fig. 4b). The expression of the main chemokines for eosinophil recruitment (CCL11 and CCL24) was also reduced in AAV2/8-OVA-treated mice (Fig. 4b). These results indicate that reduced chemokine production in response to AAV2/8-OVA treatment may prevent inflammatory cells from migrating into the lungs and alleviate the pathological features of OVA-sensitized mice.
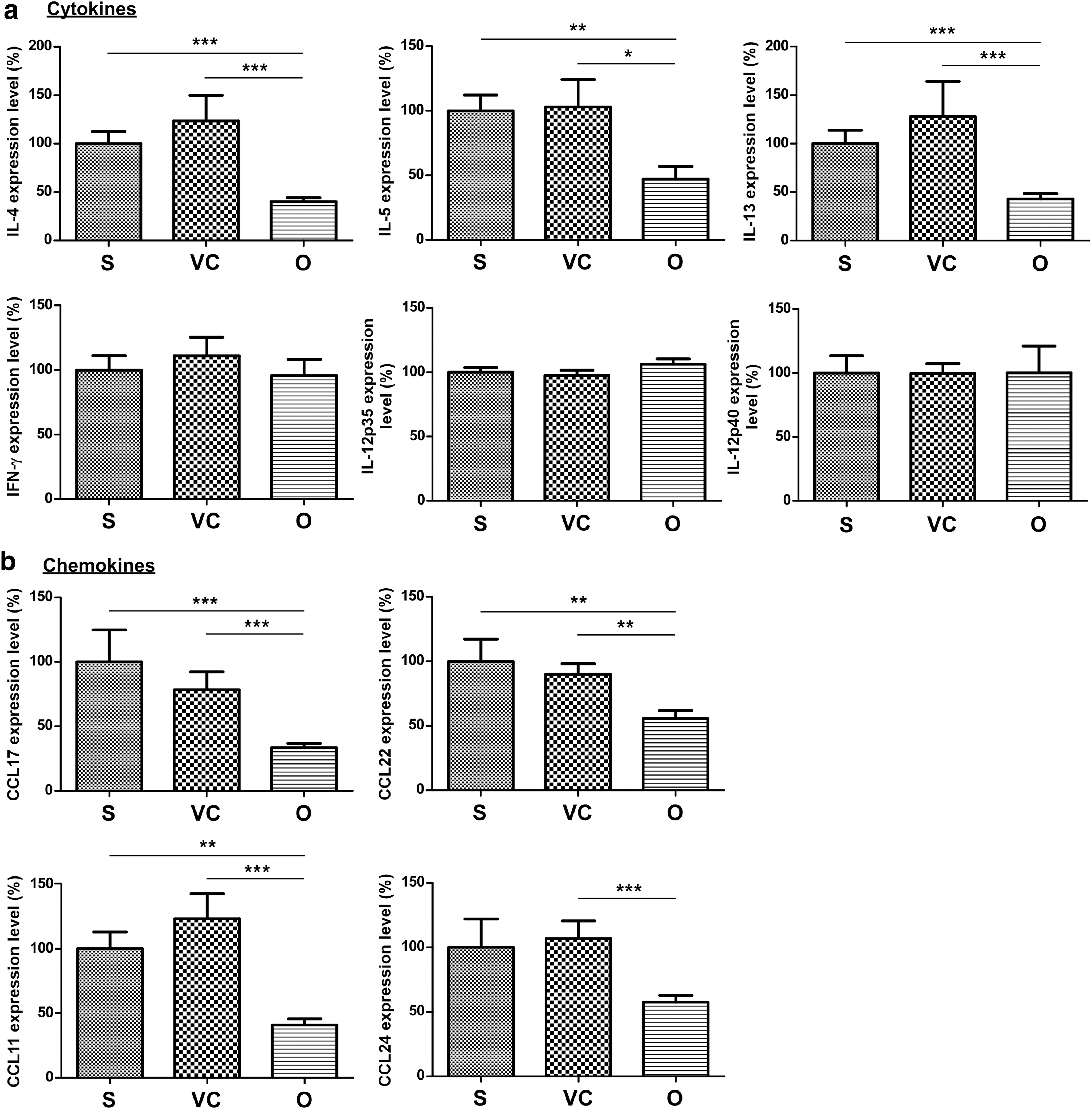
Expression of Th2-related cytokines and eotaxin chemokines was suppressed in the lungs of OVA-sensitized mice treated with AAV2/8-OVA. Total RNA was extracted from homogenized lung tissues and the RNA expression levels of
AAV2/8-OVA repressed regional and systemic Th2-driven immune responses in the lungs of OVA-sensitized mice
To determine whether AAV2/8-OVA treatment suppressed local Th2 cell-derived cytokines, culture supernatants of OVA-stimulated lymphocytes from mediastinal lymph nodes (mLNs) were examined. OVA-stimulated lymphocytes from mice receiving AAV2/8-OVA secreted less IL-4, IL-5, and IL-13 than lymphocytes from untreated and AAV2/8-EGFP control mice (Fig. 5a). Similarly, OVA-stimulated splenocytes from AAV2/8-OVA-treated mice expressed significantly lower levels of Th2 cytokines compared with untreated and AAV2/8-EGFP control mice (Fig. 5b). No significant differences were observed in IFN-γ secretion by OVA-stimulated lymphocytes from mLNs and spleens among the different groups (data not shown). Thus, AAV2/8-OVA inhibited Th2 cell-mediated immune responses and ameliorated the pathogenesis of allergic asthma.
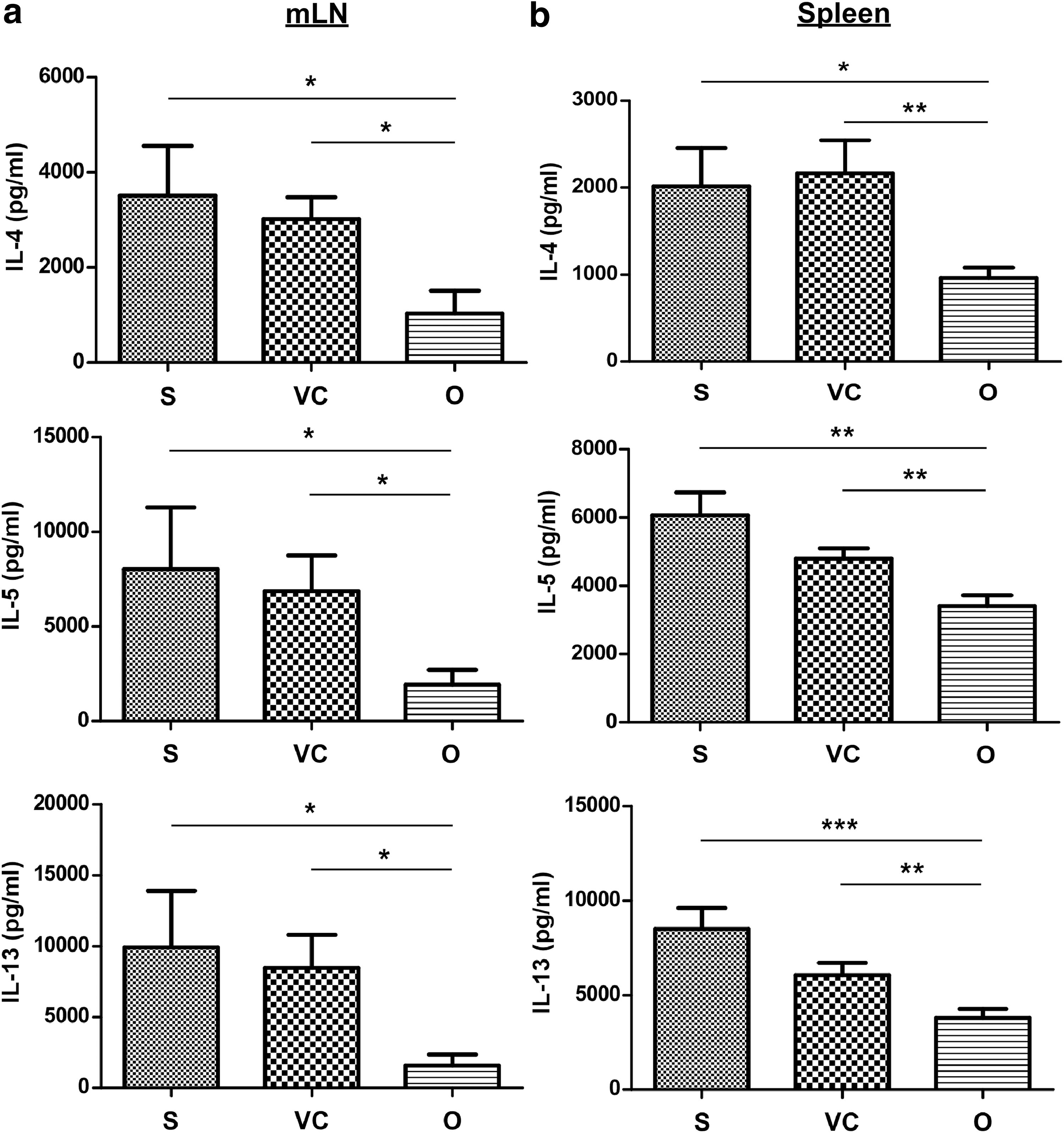
Inhibited Th2 cytokine secretion by OVA-stimulated lymphocytes from the mediastinal lymph nodes (mLNs) and spleens of OVA-sensitized mice treated with AAV2/8-OVA. Lymphocytes from
We also examined whether AAV2/8-OVA administration inhibited antibody production against OVA in serum. Before encountering allergen, serum OVA-specific antibodies were hardly detected in all mice (data not shown), even in mice infected with AAV2/8-OVA. Significantly lower levels of serum OVA-specific IgG1 and IgG2a were detected in OVA-sensitized mice treated with AAV2/8-OVA than untreated or AAV2/8-EGFP control mice (Supplementary Fig. S3a and S3b). The ratio of OVA-specific IgG1 to IgG2a was also significantly reduced (Supplementary Fig. S3d). However, hepatic OVA expression did not seem to affect serum OVA-specific IgE in asthmatic mice (Supplementary Fig. S3c). These data suggest that the administration of AAV2/8-OVA suppressed the production of OVA-specific antibodies in OVA-sensitized mice.
Higher Foxp3 expression was detected in liver and lung tissues from AAV2/8-OVA-treated mice
To explore the possible mechanisms involved in modulating the hypoimmune responses to allergen, we examined the proportions and activation phenotypes of various lymphocyte subpopulations in the liver, spleen, liver-draining lymph nodes, and mLNs. The frequencies of NK, NKT, CD3+, CD4+, and CD8+ T-cells were similar in all groups (Supplementary Fig. S4a). The activation profiles of CD4+ and CD8+ T-cells were comparable among the different groups (Supplementary Fig. S4b). Hepatocyte-restricted transgene expression was previously reported to promote immune tolerance by inducing Foxp3 expression in CD4+CD25+ Treg cells. 7,10 Thus, we investigated whether Foxp3 expression by CD4+ T-cells from asthmatic mice was affected by AAV2/8-OVA treatment. The frequency of CD4+CD25+Foxp3+ T-cells did not differ significantly in AAV2/8-OVA-treated mice and control mice (data not shown). The mean fluorescence intensity (MFI) of Foxp3 in liver-draining lymph node-isolated CD4+ T-cells was elevated by AAV2/8-OVA treatment compared with the nontreatment group (Supplementary Fig. S5). We further applied qPCR, a more sensitive method, to detect the Foxp3 RNA expression levels in CD4+ cells. The CD4+ cells from mice treated with AAV2/8-OVA expressed significantly increased levels of Foxp3 in the liver and lungs (Fig. 6a and b). Higher TGF-β and IL-10 RNA levels were also observed in the livers of mice treated with AAV2/8-OVA compared with control groups (Fig. 6c and e). In the lungs, IL-10 expression was greatly decreased in mice treated with AAV2/8-OVA compared with control groups (Fig. 6f), but the level of TGF-β was not different between the groups (Fig. 6d). These results imply that liver-specific allergen expression may create a tolerogenic microenvironment through enhanced amounts of TGF-β and IL-10 in the liver, promoting Foxp3 expression in CD4+ T-cells.

AAV2/8-OVA significantly enhanced Foxp3 expression in CD4+ cells from the livers and lungs of OVA-sensitized mice. The livers (n = 6–8 per group) and lungs (n = 10 per group) were homogenized for RNA extraction at the end of the sensitization protocol. Expression levels of
Discussion
Allergic asthma has become a global public health issue that has been considerable burden on health services and incurs expensive social costs. Despite a growing number of pharmaceutical and immunotherapy strategies for asthma, the development of an active, effective, and long-term immunomodulatory approach has been difficult. Here, our results demonstrate for the first time that specific expression of allergen in hepatocytes by rAAV-mediated gene transfer induces tolerogenic immunity, which alleviates allergic airway inflammation and inhibits Th2 cell-driven immune responses. Pseudotyped AAV2/8 vector with hepatocyte-restricted promoter specifically transduced the allergen gene in the liver. OVA-specific Th2 immune responses and antibody production were significantly suppressed in asthmatic mice receiving AAV2/8-OVA. These results suggest that ectopic allergen expression in hepatocytes resulted in local and systemic hypoimmune responses to allergen re-challenge. Consequently, the application of the liver tolerance effect may be a novel strategy for therapeutic modulation and intervention in allergic asthma.
Antigen-specific tolerance has been demonstrated by transgene expression in hematopoietic stem cells, immature APCs, and tolerogenic tissues, such as the liver and thymus. 5 Gene transfer-mediated immune tolerance has been applied in the treatment of hereditary protein deficiencies 25 and autoimmune diseases 10 in animal models and clinical trials. 26 In a murine model of hemophilia B, hepatocyte-restricted factor IX expression sustained therapeutic protein levels and evaded unwanted immune responses. 27 In addition, hepatocyte-specific gene transfer of α-galactosidase and acid sphingomyelinase has been shown to trigger tolerogenic immunity and improve pathological features of Fabry and Nieman-Pick disease. 18,28 In an animal model of human multiple sclerosis, neural autoantigen-expressing hepatocytes prevented and repressed CD4+ T-cell-driven neuroinflammation. 10 Some potential mechanisms of immune tolerance by hepatic gene transfer have been proposed that involve the elimination and unresponsiveness of effector T-cells or the induction of Tregs. 7,8,10 The development of transgene-specific CD4+CD25+Foxp3+ Tregs has also been reported after hepatic gene transfer. 7,10 For allergic disease, the current study demonstrates that allergen-expressing hepatocytes effectively protect against asthma development.
How lung inflammation was alleviated by allergen expression in the liver is currently not clear. The phenomenon of “liver tolerance” has been shown to be local and systemic tolerance of self and foreign antigens presented by nonparenchymal cells or expressed in hepatocytes. Several nonparenchymal cells are involved in the tolerogenic properties of the liver, including dendritic cells, 29 liver sinusoidal endothelial cells (LSECs), 30 Kupffer cells, 31 and hepatic stellate cells. 32 These unique cells regulate tolerant responses via the production of anti-inflammatory cytokines, such as IL-10, TGF-β, or programmed cell death ligand-1. 33 Furthermore, hepatocytes directly contact circulating T-cells through the specialized fenestrations of LSECs. The immunological functions of hepatocytes are facilitated by the polarized expression of MHC and ICAM-1 molecules toward the sinusoidal lumen. 34 A previous study suggested that T-cells activated by hepatocytes die prematurely, leading to immune tolerance. 35 Thus, it is likely that antigen presented by hepatocytes or other cell populations is able to modulate dysregulated immune responses.
The induction of CD4+CD25+ Tregs in tolerance to the transgene product or autoantigen has been reported after hepatic in vivo gene transfer. 7,10 Based on the aberrant number of Tregs and Foxp3 mRNA levels in patients allergic to house dust mites (HDMs), 36,37 the increased Foxp3 gene expression in the liver and lungs of AAV2/8-OVA-treated mice suggested the possibility of the development of Tregs. Liver-generated CD4+Foxp3+ cells are recruited to or circulate in the inflamed lung tissue to suppress Th2 cell-driven inflammation. Thus, the increased Foxp3 gene expression levels in the liver and lungs (Fig. 6a and b) suggest that hepatic gene transfer-induced CD4+Foxp3+ Tregs partially contributed to the tolerogenic effect. Similar to oral tolerance, the reduction of AHR, lung inflammation, and Th2 cytokines is associated with an increased proportion of airway CD4+Foxp3+ Tregs in asthmatic mice. 38 IL-10 and TGF-β are well documented to be crucially involved in the maintenance of immune tolerance to transgene products and peripheral Treg conversion. 10,39 In the allergen milieu of the liver, significantly increased expression of TGF-β and IL-10 was detected in OVA-sensitized mice treated with AAV2/8-OVA (Fig. 6). Allergen stimulation combined with immunosuppressive cytokines in the liver probably facilitated Foxp3 acquisition of CD4+ cells with both cytokines. Nevertheless, IL-10 expression was significantly decreased after AAV2/8-OVA treatment in the lungs. Although the reduction in IL-10 may be attributed to reduced lymphocyte recruitment into the lungs, whether IL-10 directly suppresses lung inflammation or indirectly targets allergen-mediated responses through enhanced Foxp3 expression in the lung tissues is not clear. Nevertheless, we cannot formally prove which mechanism is really involved in the suppression of lung inflammation in asthmatic mice, though it is clear that the alleviated allergic responses occurred when the allergen gene was expressed in the liver.
Various factors, including dose, nature, and expressed form of the antigen, exposure route, delivery system, and mouse strain, determine the development of immune activation or tolerance. The location where antigen encounters APCs is also important. 40,41 We tested whether different subcellular antigen localizations will influence immune tolerance by hepatic gene transfer. Although both membrane-anchored and secreted antigens induce antigen-specific antibodies, 42,43 the activation of CD4+ T-cells was greater in mice receiving membrane-anchored antigen than those receiving secreted antigen. 42 Other studies have indicated that mice immunized with soluble antigens encoded by DNA-based vaccine provoke a strong IgG1 response and greater levels of IL-4, but this does not occur with the membrane-bound antigen. 44,45 Our results indicate that mice receiving secreted OVA (treated with AAV2/8-sOVA) had higher eosinophilia and Th2 cytokine RNA expression in the lungs than mice treated with vector carrying membrane-anchored OVA (Supplementary Fig. S6). Highly efficient immune tolerance was also induced by the transmembrane form of neural autoantigen in hepatocytes in autoimmune neuroinflammation. 10 On the other hand, low doses of antigen have been demonstrated to efficiently induce and expand Tregs compared with high doses of antigen. 46,47 Lower levels of OVA transgene expression were observed in the livers of mice receiving vector carrying membrane-anchored OVA compared with those receiving secreted OVA (data not shown). Thus, we speculate that membrane-bound OVA provides an environment for less effective activation of Th2 cells and less antigen exposure promotes Treg function compared with secreted OVA.
AAV is a favorable vector for gene transfer because of its nonpathogenic nature, widespread infectivity, and stable transgene expression. 12 Over the past few decades, pseudotyped AAV vectors have been developed extensively and exerted therapeutic efficacy in disease models. A growing number of successful gene therapies have also been reported in clinical trials of AAV for inherited disorders and acquired diseases. 26 Our previous studies successfully applied an AAV2/9 vector carrying CC10 to efficiently target lung epithelial cells and restore CC10 levels, alleviating lung inflammation in OVA-sensitized mice. 15 In addition, AHR, eosinophilia, and airway remodeling were significantly mitigated in HDM allergen-sensitized mice with AAV2/9-delivered CCL11 small hairpin RNA. 17 Compared with overall immune-suppressive chemokine-modulatory gene sequences, antigen-specific immune tolerance may have fewer side effects. In addition, the liver is a more tolerogenic organ and evidence has demonstrated that hepatic gene transfer using AAV vector may promote transgene-specific systemic immune tolerance. 7,48,49 The AAV8 serotype preferentially targets the liver to support sustained transgene products and induce active tolerogenic responses. 50,51 AAV2/8 vector carrying therapeutic transgenes predominantly assists tolerant responses to transgenes in mice with lysosomal storage disorders. 18,28 The livers of patients with hemophilia B have been transduced successfully by AAV2/8-Factor IX vectors with no lasting toxicity and transiently achieving therapeutic levels. 52 We established liver-restricted allergen expression with AAV2/8 vectors, which resulted in immune hyporesponsiveness to OVA-induced antibody production and Th2 cytokine secretion in OVA-sensitized mice. Thus, AAV2/8 vector combined with a hepatocyte-specific promoter was a beneficial vehicle for hepatic gene transfer to suppress hypersensitive lung responses to allergen. Similar strategies may be applied for other allergic diseases that do not have a localized target, such as atopic dermatitis. Furthermore, liver-mediated immune tolerance may be beneficial for systemic or organ-specific autoimmune diseases.
In conclusion, ectopic expression of antigen in the liver by AAV-directed gene transfer has been applied gradually to induce tolerant responses against transgene products. For the first time, our study indicates that hepatocyte-restricted allergen expression by pseudotyped AAV2/8 vector effectively suppressed asthmatic pathogenesis in allergen-sensitized mice. AAV2/8 vector carrying the OVA transgene upregulated the expression of immunosuppressive cytokines, which may facilitate Foxp3 acquisition of CD4+ cells in the liver. Therefore, targeting allergen expression to the liver serves as a novel approach to induce active tolerance for the treatment of allergic disease.
Footnotes
Acknowledgments
This work was supported, in part, by grants from the National Science Council (NSC101-2320-B-182-033) and Ministry of Science and Technology (MOST102-2320-B-182-030-MY3) of the Republic of China, as well as Chang Gung Memorial Hospital (CMRPD190513, CMRPG1A0071 ∼ 3, CORPD1F0021, and BMRP362).
Author Disclosure
None of the authors have any financial conflict of interest related to this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.