
Abstract
Mucopolysaccharidosis type IIIA (MPS IIIA) is predominantly a disorder of the central nervous system, caused by a deficiency of sulfamidase (SGSH) with subsequent storage of heparan sulfate-derived oligosaccharides. No widely available therapy exists, and for this reason, a mouse model has been utilized to carry out a preclinical assessment of the benefit of intraparenchymal administration of a gene vector (AAVrh10-SGSH-IRES-SUMF1) into presymptomatic MPS IIIA mice. The outcome has been assessed with time, measuring primary and secondary storage material, neuroinflammation, and intracellular inclusions, all of which appear as the disease progresses. The vector resulted in predominantly ipsilateral distribution of SGSH, with substantially less detected in the contralateral hemisphere. Vector-derived SGSH enzyme improved heparan sulfate catabolism, reduced microglial activation, and, after a time delay, ameliorated GM3 ganglioside accumulation and halted ubiquitin-positive lesion formation in regions local to, or connected by projections to, the injection site. Improvements were not observed in regions of the brain distant from, or lacking connections with, the injection site. Intraparenchymal gene vector administration therefore has therapeutic potential provided that multiple brain regions are targeted with vector, in order to achieve widespread enzyme distribution and correction of disease pathology.
Introduction
L
MPS IIIA patients appear normal at birth, but usually present clinically between the age of 2 and 6 years with a range of symptoms such as hyperactivity, sleep disturbance, delayed development, and loss of learned skills; the disorder is associated with progressive central nervous system (CNS) degeneration and mild somatic effects, 2 and the median age of death is 18 years. 3
Therapy is currently unavailable for MPS IIIA, although several strategies are being trialed. In the clinic, unmodified bone marrow transplantation has been successful in treating several neuropathic LSD, including early presenting MPS I (Hurler), late-onset metachromatic leukodystrophy, and early onset Krabbe disease 4 ; however, cognitive improvement in MPS IIIA patients (and MPS IIIA mice) is not seen. 5,6 Enzyme replacement therapy (ERT) is utilized in several nonneurodegenerative LSD. 7 –9 However, because of the protective nature of the blood–brain barrier, intravenous administration of enzyme at conventional doses is unlikely to treat the CNS. This can be circumvented by intracerebrospinal fluid (CSF) administration of enzyme, which is currently under clinical investigation in MPS IIIA patients. However, this therapeutic approach requires repeated administration and with this comes the risk of infection, potentially limiting this strategy to the medium-term.
Gene therapy has the capacity to overcome the challenges faced by ERT, where once-off vector administration could theoretically achieve widespread CNS transduction, long-term gene expression, and subsequent cross-correction of neighboring nontransduced cells by endocytosis of secreted mannose-6-phosphated enzyme, increasing the therapeutic sphere of treatment. A clinical trial using an adeno-associated virus (AAV) serotype 2-based vector to deliver the human ceroid lipofuscinosis neuronal 2 (CLN2) gene, 10 encoding tripeptidyl-peptidase I (TPP-I) to patients with late infantile neuronal ceroid lipofuscinosis (LINCL), has been conducted. The trial demonstrated slower (albeit not statistically significant) rates of neurological decline using magnetic resonance imaging and the modified Hamburg LINCL clinical rating scale. 11 This suggests potential clinical relevance for this treatment type. A further study in LINCL patients using an AAVrh10 serotype vector to deliver the human CLN2 gene is now in the recruitment phase.
A mouse model for MPS IIIA has been described 12 with pathological changes and disease course resembling the human condition. This model has been used to test the therapeutic efficacy of gene therapy using an AAV2/5-CMV-SGSH-IRES-SUMF1 vector to deliver recombinant human SGSH (rhSGSH) and the sulfatase modifying factor (SUMF1; essential and limiting in the activation of all sulfatases), 13 bilaterally to the lateral ventricles of newborn presymptomatic mice. A significant reduction in neuronal and glial vacuolization was observed in the brain regions examined, and AAV2/5-CMV-SGSH-IRES-SUMF1-treated MPS IIIA mice demonstrated behavioral improvements compared with control MPS IIIA mice. 14 Similarly, a single unilateral intrastriatal (putamen) injection of AAV serotypes 2 or 5 encoding α-N-acetylglucosaminidase (NaGlu) in 6-week-old MPS IIIB mice significantly improved behavioral outcomes and reduced secondary GM2 and GM3 ganglioside storage in areas where NaGlu was detected. NaGlu was detected in both hemispheres of all AAV5-NaGlu-injected mice. 15
AAV serotype selection is an important factor to consider for optimal cellular transduction, vector and enzyme distribution, and immune system evasion. Cellular tropism of various AAV serotypes in the CNS has been investigated, with AAV 1, 2, 7, 8, 9, and rh10 transducing neurons specifically, 16 –18 while AAV5 may also conditionally transduce astrocytes. 19 Of these serotypes, transgene expressed from AAVrh10 exhibits the largest rostral-caudal distribution, ahead of AAV9. 18 Axonal transport of AAV vectors is evident for several serotypes, including AAVrh10, and hence their administration can be tailored to take advantage of established neural circuitry. 20 The striatum has various efferent neuronal connections, predominantly to the ipsilateral external globus pallidus, globus pallidus pars interna, substantia nigra pars reticulate, and ventral tegmental area, 21 the latter of which has substantial connections throughout the brain. 22 The striatum is also connected via afferent neurons to several of these regions in addition to the thalamus, cortex, and amygdala. 21,23 Of important note is that, in a rat study, AAVrh10 evades preexisting immunity to the common AAV serotypes naturally infecting humans (AAV 1, 2, and 5). 24
We took advantage of the AAVrh10 vector characteristics and used it to deliver the human SGSH and SUMF1 genes (AAVrh10-SGSH-IRES-SUMF1; commercially referred to as SAF-301 and subsequently referred to here as AAV-SGSH) into the striatum of presymptomatic 5-week-old MPS IIIA mice. 12 We then examined the efficacy of this vector construct in mediating long-term gene expression and subsequent reduction of primary and secondary neuropathology in MPS IIIA mice, with the aim of providing preclinical data as to the suitability of this therapeutic approach for use in MPS IIIA children.
Materials and Methods
Mice
Congenic C57BL/6 MPS IIIA or heterozygous/normal littermates (referred to as “unaffected”) were genotyped after extraction of genomic DNA in 50 μl of 50 mM Tris pH 8.0, 2 mM NaCl, 1 mM EDTA, and 0.5% (v/v) Tween-20 supplemented with 1.2 mg/ml proteinase K. After heating at 95°C for 5 min, 1–2 μl of clarified lysate was amplified with forward primer 5′-NNT CTG TCT TCC TCA GCG-3′, reverse primer 5′-GAT AAG GCT GTG GCG GGA CAG GG-3′ (final concentration of 4 ng/μl of each primer), and HotStarTaq DNA Polymerase (Qiagen, Doncaster, Australia) in a 50 μl reaction by denaturing at 94°C for 15 min followed by 35 cycles of 94°C for 45 sec, 55°C for 45 sec, and 72°C for 40 sec, and a 5 min final extension at 72°C. After visualization of the 105 bp PCR product, amplified DNA was digested with 5 U AciI (New England Biolabs, Ipswich, MA) before electrophoresis through a 4.5% (w/v) agarose gel. As the G91A mutation abolishes an AciI site, wild-type mice display bands at 74, 16, and 15 bp, whereas MPS IIIA mice display bands at 90 and 15 bp.
All experimental procedures were undertaken with the approval of the WCHN Animal Ethics Committee, in accordance with the guidelines of the National Health and Medical Research Council of Australia on the use and care of experimental animals. The congenic strain was created after 10 backcrosses to inbred C57Bl/6 mice. 12 Animals received food and water ad libitum and were housed in a constant temperature/humidity, with a 14 hr light:10 hr dark cycle. Housing was enriched using sterilized toilet rolls, yoghurt containers, and shredded paper as nesting material.
Vector
Laboratory grade vectors were supplied by the Belfer Gene Therapy Core Facility, Weill Medical College Cornell University. AAVrh10 vectors carrying a defective AAV2 genome containing either the GFP gene driven by the cytomegalovirus (CMV) promoter or SGSH-IRES-SUMF1 genes driven by the phosphoglycerate kinase (PGK) promoter were utilized. Briefly, vectors were produced by transfection of 293 ORF6 cells. Three days posttransfection, the vector was released by three cycles of freeze–thaw, benzonase treatment, and an iodixanol step gradient. Pooled iodixanol fractions were applied to an ion-exchange column and the purified flow through was collected, and purity was assessed by SDS-PAGE. AAV2/rh10-CMV-GFP (control) and AAV2/rh10-PGK-SGSH-IRES-SUMF1 (AAV-SGSH) titers were 2.5 ± 0.7 × 1010 gc/μl and 3.0 × 1010 gc/μl, respectively. Vectors were diluted 1/10 in sterile PBS (Sigma-Aldrich, Castle Hill, NSW, Australia) and re-frozen in small aliquots before use in mice. Clinical-grade preparations of this AAV-SGSH vector have been used in the human clinical trial NCT# 01474343.
Intracerebral injections
Mice received intraparenchymal injections at 5 weeks of age according to the scheme shown in Fig. 1. A total of 96 mice were utilized and there were four treatment groups: unaffected + AAV-GFP (unaffected control); unaffected + AAV-SGSH (unaffected SGSH); MPS IIIA + AAV-GFP (MPS IIIA control); and MPS IIIA + AAV-SGSH (MPS IIIA SGSH), with 24 mice (15 males and 9 females) per treatment group.
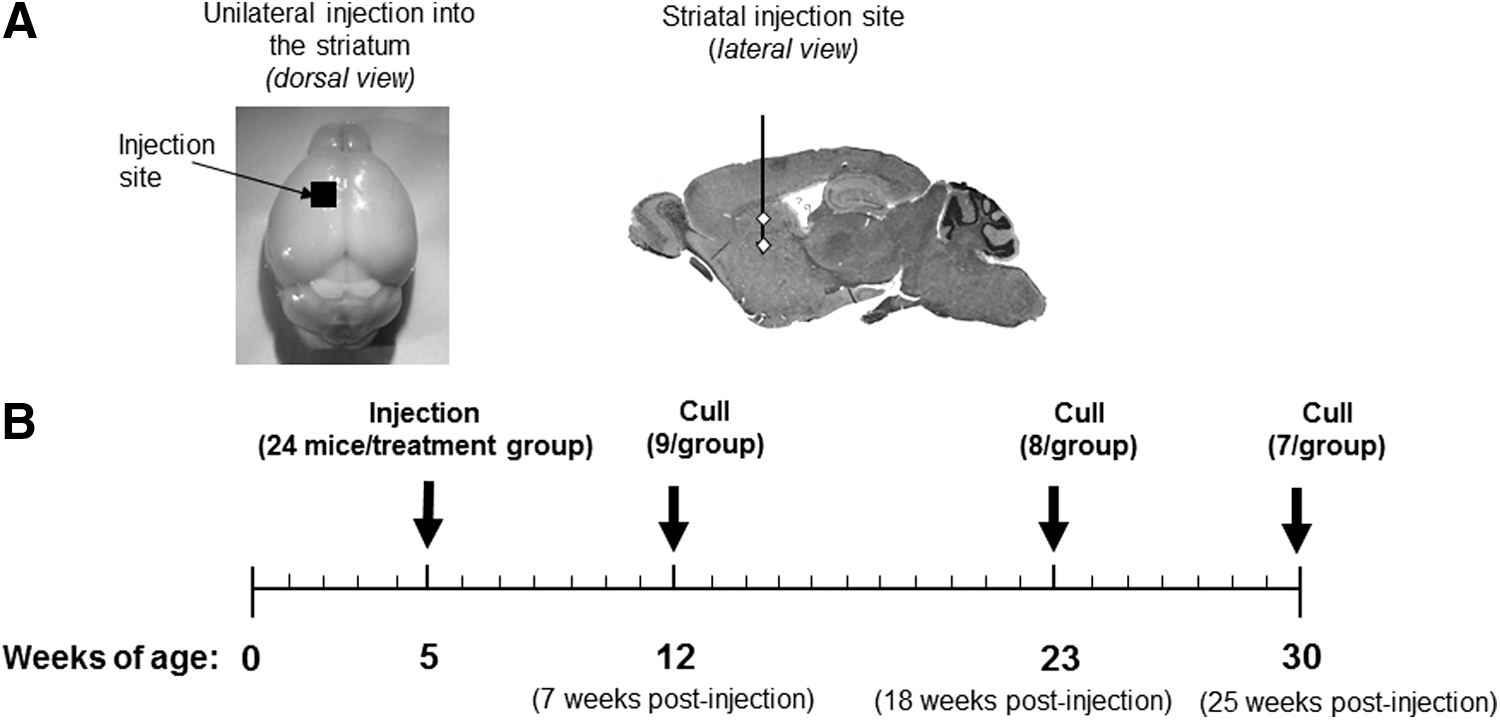
Experimental plan.
Mice were pretreated with glycopyrrolate (Robinul, 0.01 mg per mouse i.m.; Wyeth Ayerst, Baulkham Hills, NSW, Australia) and anesthetized using ketamine (87 mg/kg; Parnell Laboratories, Alexandria, NSW, Australia) and xylazine (13 mg/kg, Xylazil-20; Troy Laboratories, Sydney, NSW, Australia). Once mice were unresponsive, they were secured in a stereotaxic frame (David Kopf Instruments, Tujunga, CA). After resection of the scalp, a burr hole was made using a hand drill fitted with a 0.5 mm drill bit (Flintware, Adelaide, SA, Australia). Mice received an injection of vector (0.5 μl/min) via a Hamilton syringe at two depths into the left striatum (coordinates in reference to bregma and the midline: 0.75 mm anterior; 1.5 mm lateral; −2.0 mm and −3.5 mm ventral). 25 At each of the two ventral depths either 1.5 μl of AAV-GFP or 1.25 μl AAV-SGSH was delivered (7.5 × 109 g.c. delivered per mouse). All mice received warmed dextrose (4%) in NaCl (0.18%; 3 ml/100 g body weight) plus butorphanol tartrate for analgesia (2 mg/kg; Fort Dodge, Baulkham Hills, NSW, Australia) after surgery.
Necropsy and sample collection
Mice were euthanized by CO2 asphyxiation at 12 weeks (n = 36), 23 weeks (n = 32), or 30 weeks of age (n = 28). Blood samples were taken via cardiac puncture; some mice in each group were then PBS-perfused (pH 7.4) and 2 mm hemicoronal brain slices collected using a brain blocker (Braintree Scientific, Braintree, MA). Tissues were frozen for subsequent measurement of heparan sulfate-derived oligosaccharide storage and SGSH content. The remaining mice were fixation-perfused in 4% paraformaldehyde in PBS for light microscopic examination. The brains were sliced sagittally along the midline and also at 2 mm lateral to the midline on both the left (injected) and right (contralateral) hemispheres. The two medial slices were paraffin-embedded and the remaining more lateral tissue was snap-frozen for cryosectioning.
Detection of the SGSH protein in brain homogenates
The SGSH protein was quantified using a DELFIA detection system according to a previously published method 26 using an rhSGSH (Shire Human Genetic Therapies Inc., Cambridge, MA) standard curve range of 0–0.625 ng. Readings greater than two standard deviations above the mean background were considered significant. Two- to three replicates of a quality control (QC) sample, which consisted of an MPS IIIA dog brain homogenate known to contain rhSGSH, were used for each plate. The intraassay coefficient of variation of the QC was 4.4%, whereas the interassay coefficient of variation of QC was 15.2% (n = 8). The limit of detection of rhSGSH per well was determined to be 4.9 pg. Supernatant protein concentrations were determined with a micro-BCA protein assay kit (PIE23235, Pierce Protein Research Products; ThermoFisher Scientific, Rockford, IL).
Detection of SGSH activity in brain homogenates
The method of Hopwood and Elliott 27 was used to assay SGSH activity in brain homogenates.
Tandem mass spectrometric analysis of heparan sulfate-derived glucosamine-N-sulfate [α-1,4] hexuronic acid (GlcNS-UA)
Samples were prepared essentially as reported previously, 28 except that 100 μg of homogenate protein was assessed; centrifugation post-protein precipitation was at 14,600 g for 4 min, and the resulting supernatants (200 μl) were collected and acidified with 400 μl of 0.4 M formic acid.
Mass spectrometric analysis of disaccharide was performed in negative ion mode as previously described. 28 The relative peak area of disaccharide (GlcNS-UA) measured in each sample was determined by relating the peak area of derivatized disaccharide to the derivatized internal standard (4-deoxy-L-threo-hex-4-enopyranosyluronic (1 → 3) N-acetyl-galactosamine-4-sulfate; C4045; Sigma-Aldrich). Tissues were processed in batches to minimize the impact of batch-to-batch variation: a batch consisted of a complete time-point group, that is, 12, 23, or 30 weeks of age, containing the four brain slices. The intrabatch coefficient of variation was 9.0% (n = 7) and interbatch coefficient of variation was 17.3% (n = 3), determined using MPS IIIA mouse brain QC samples.
Immunohistochemistry and histochemistry
All procedures and poststaining image analyses were undertaken by an experimenter blinded to genotype and treatment status. Six-micrometer-thick sagittal paraffin sections of both the left and right brain hemispheres were cut at levels between 1.32 and 1.56 mm lateral to the midline for 12- and 23-week-old mouse groups, and 1.08 to 1.2 mm for 30-week-old mice (based on stereotaxic coordinates in a mouse brain atlas). 25
Immunohistochemistry was performed on deparaffinized sections. Primary antibodies to GFAP (Z334) and ubiquitin (Z458) were purchased from Dako (Glostrup, Denmark). Antibodies to SGSH (2C7) and endo/lysosomes (LIMP-II) were sourced and used as previously described. 26 ,55 P62/SQSTM1 was detected with a 1:2000 dilution of the p62/anti-SQSTM1 monoclonal antibody, clone 2C11 (H00008878-M01; Abnova, Taipei, Taiwan). Histochemical staining of activated microglia by incubation in a peroxidase-conjugated isolectin-B4 from Bandeiraea (Griffonia) simplicifolia (BSI-B4) (L5391; Sigma-Aldrich) was performed as previously described. 29
Six-micrometer-thick sagittal sections of cryoprotected brain tissue were cut on a cryostat (Shandon Cytochrome E; Thermo Scientific, Cheshire, England) at a lateral position from the midline of between ∼1.92 and 2.4 mm for detection of GM3 or filipin-positive inclusions. Immunodetection of GM3 was performed essentially using the method of McGlynn et al. 30 (anti-GM3 antibody; 370695; Seikagaku Biobusiness Corporation, Tokyo, Japan). Histochemistry was used to qualitatively assess filipin-positive inclusions, as previously described (Streptomyces filipin, F9765; Sigma-Aldrich). 6,30
Sections from 12- and 23-week-old mice were batch-stained before being viewed on an Olympus BX41 microscope. Digital images were collected using an Olympus CVIII camera at 200× magnification. AnalySIS Lifescience software (version 2.8, Build 1235; Olympus Soft Imaging Solutions, Munster, Germany) was used for threshold analysis based on the optical density of positive immunostaining, and was reported as % GM3 ganglioside, % LIMP-II, % GFAP, and % ubiquitin immunoreactivity. Manual counts were made of activated microglia and p62-positive inclusions, reported as number/mm2. Filipin-positive inclusions were semiquantitatively assessed with the number of positively stained cells scored on a −, −/+, +, ++, +++, and ++++ scale.
Detection of anti-rhSGSH antibodies in serum
Plasma samples were assayed as previously described, 31 using a horseradish peroxidase-based assay to detect antibodies to SGSH. Antibody titers are expressed as the lowest serum dilution giving an absorbance greater than two standard deviations above the blank.
Statistical analysis
All data were assessed using GraphPad Prism version 5.02 for Windows (San Diego, CA). Biochemical (heparan sulfate-derived oligosaccharide and SGSH) data were assessed using one-way ANOVA followed by post-hoc testing using a Bonferroni correction to adjust for multiple comparisons. Histological data were log-transformed (Y = log [Y + 1]) to better satisfy normality assumptions, and were subsequently examined using one-way analysis of variance (ANOVA) with post-hoc testing using a Bonferroni correction to adjust for multiple group comparisons. Data are shown as mean ± SEM and p < 0.05 was considered to be statistically significant. Symbolic representation of statistics in figures is as follows: *, MPS IIIA controls versus unaffected controls or MPS IIIA-SGSH versus unaffected-SGSH; ^, MPS IIIA controls versus MPS IIIA-SGSH; °, unaffected controls versus unaffected-SGSH; *, ^, or °, p < 0.05; **, ^^, or °°, p < 0.01; ***, ^^^, or °°°, p < 0.001. Treatment group comparisons were consistently made between equivalent regions in the same hemisphere.
Results
SGSH expression in brain after intrastriatal injection of AAV-SGSH
Immunohistochemical localization of rhSGSH revealed intracellular punctate staining in what appeared to be the lysosomal compartment (based on morphology) in soma and axons (Fig. 2A). Immuno-positive staining was observed only in the ipsilateral hemisphere of AAV-SGSH-treated mice at 12 and 23 weeks of age (7 and 18 weeks p.i., respectively). Staining was widespread along the injection tract, striatum, frontal association, and rostral (secondary motor) cortex and anterior olfactory nucleus. Less frequently, positive staining was seen in the cingulum and olfactory bulb.

Sulfamidase distribution.
Quantification of SGSH in mouse brain
Supraphysiologic levels of SGSH (i.e., 113- to 337-fold unaffected) were detected in the brain slice containing the injection site (L2) of both MPS IIIA-SGSH and unaffected-SGSH mice, compared with control MPS IIIA or unaffected mice (respectively) at 12, 23, and 30 weeks of age (Fig. 2C–E). On average, 185-fold more SGSH was observed in L2 from AAV-SGSH-treated mice compared with control mice. In MPS IIIA-SGSH mice, enzyme levels in slice L4 (Fig. 2C–E) were greater than or equal to unaffected control mice at each age, and SGSH levels in R2 and R4 were generally less than those found in unaffected control mice. At 30 weeks of age, a trend toward raised levels of SGSH was observed beyond the injection site, with L2 > L4 > R2 > R4 in AAV-SGSH-treated mice, although statistical significance was not achieved.
SGSH activity in mouse brain
When we examined the activity of SGSH in these same brain slices, we found that brain slice L2 (containing the injection site of MPS IIIA-SGSH mice) exhibited high activity levels at all times postinjection (11–23-fold unaffected-GFP; data not shown). In contrast, brain slices further from the injection site exhibited much lower amounts of SGSH activity. In MPS IIIA-SGSH brain slice L4, we observed 15–57% unaffected-GFP enzyme activity. In brain slice R2 we recorded 6.5–54% unaffected mouse SGSH activity and in slice R4 there was 6.5–30% of unaffected levels of SGSH recorded.
Effect of treatment on primary lysosomal storage of heparan sulfate
To determine the biochemical efficacy of the expressed SGSH, the relative level of a heparan sulfate-derived disaccharide (GlcNS-UA) was measured by tandem mass spectrometry (Fig. 3). MPS IIIA control mice exhibited statistically significant elevations of GlcNS-UA compared with unaffected control mice in each slice, at each time point. Statistically significant reductions in GlcNS-UA were observed in L2 and L4 at 12 weeks of age in MPS IIIA-SGSH mice compared with MPS IIIA control mice (Fig. 3A). However, no statistically significant reduction was observed in slices R2 and R4 (Fig. 3A). At 23 (Fig. 3B) and 30 (Fig. 3C) weeks of age, statistically significant reductions in GlcNS-UA were observed in all brain slices in MPS IIIA-SGSH mice compared with MPS IIIA control mice. Remaining GlcNS-UA levels in slice L2 of MPS IIIA-SGSH mice were no longer significantly different from levels recorded in unaffected-SGSH mice at both later time points (23 and 30 weeks of age). The general trend of storage reduction was consistently L2 > L4 > R2 > R4, particularly evident at 23 and 30 weeks of age (Fig. 3D).

Primary heparan sulfate storage.
Effect of treatment on secondary pathological changes
Endosomal/lysosomal system expansion
Significant elevations of lysosomal integral membrane protein II (LIMP-II) staining were observed in all brain regions examined in MPS IIIA control mice compared with unaffected control mice (Supplementary Figs. S1 and S2; Supplementary Data are available online at
GM3 ganglioside accumulation
Accumulation of GM2 and GM3 gangliosides has been reported in several of the MPS, including MPS IIIA. 30 Here we observed a significant elevation in GM3 in many regions of the brain (Fig. 4 and Supplementary Fig. S3), including the rostral cerebral cortex, caudal (retrosplenial) cortex, and amygdala of MPS IIIA control mice (compared with unaffected mice) by 12 weeks of age. No statistically significant treatment effect was observed in MPS IIIA-SGSH mice at 12 weeks of age in any region assessed. However, by the time mice reached 23 weeks of age, statistically significant reductions in GM3 (to levels approaching those seen in unaffected mice) were observed in the rostral cortex in both hemispheres. These reductions were maintained to 30 weeks of age.
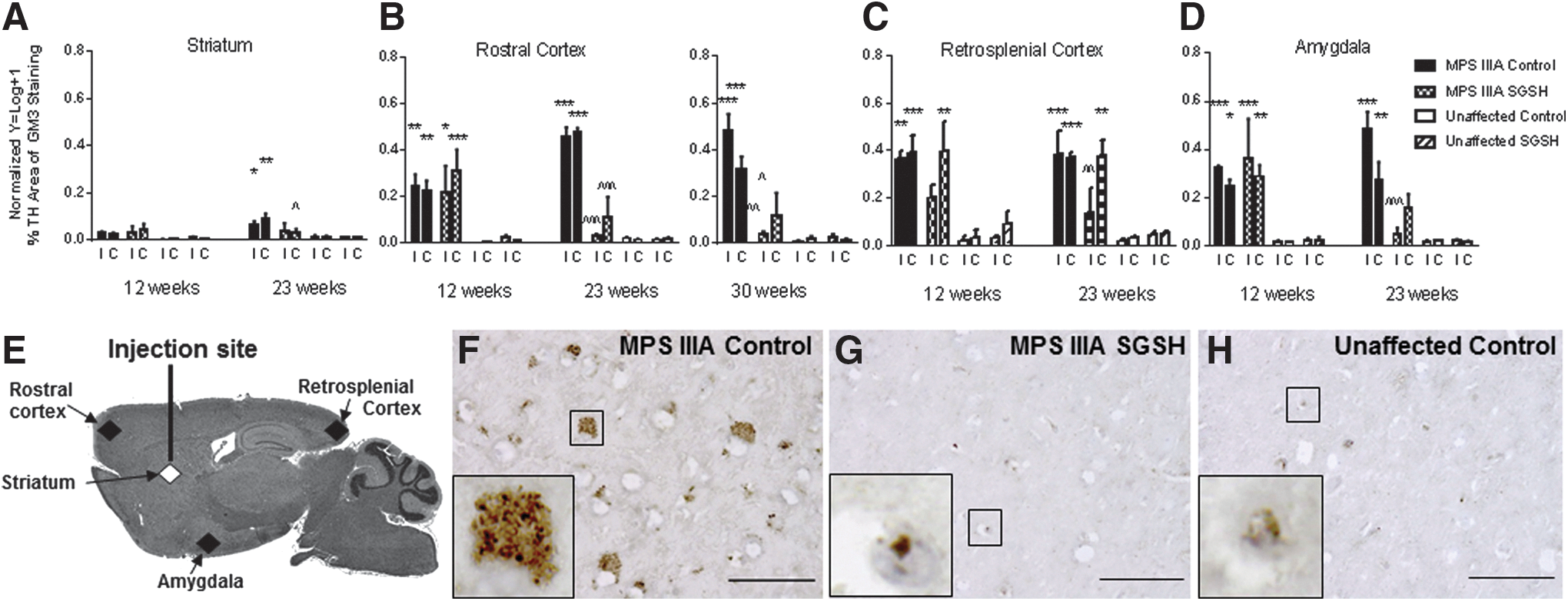
GM3 ganglioside immunohistochemistry.
Filipin-positive inclusions
Filipin-positive inclusions have previously been observed in MPS IIIA mouse brain. 30 A semiquantitative assessment of staining revealed elevations in MPS IIIA control mouse brain at all time-points assessed, compared with the staining seen in similarly treated unaffected mice (Supplementary Table S1). There was little impact of the treatment on the number of filipin-positive inclusions, when sections from 12-week-old mice were examined. However, by the time MPS IIIA-SGSH mice reached 23 weeks of age, the staining pattern was essentially normalized in all assessed regions of the ipsilateral hemisphere. Normalization was also observed in the rostral cortex of the contralateral hemisphere. Inconsistent improvements were observed in retrosplenial cortex and amygdala of the contralateral hemisphere. The apparent normalization of staining in the rostral cortex of the injected hemisphere was maintained with time, as assessed in 30-week-old mice.
Neuroinflammation
Glial fibrillary acidic protein
MPS IIIA mice have previously been found to exhibit significant elevations in glial fibrillary acidic protein (GFAP) compared with unaffected mice. 28 Here, we observed elevations in GFAP expression in brain regions close to/at the injection site in both MPS IIIA and unaffected mice treated with both vector types. This was presumptively caused by the formation of a glial scar and made interpretation of the impact of treatment impossible at early time points postinjection. AAV-SGSH treatment of MPS IIIA mice resulted in the first statistically significant reductions in GFAP expression in the ipsilateral medial cortex and both hemispheres of the more distal thalamus at 23 weeks of age (Fig. 5A–F and Supplementary Fig. S4). Structures situated caudal to the injection site, for example, the inferior colliculus, exhibited no reduction in GFAP expression in MPS IIIA-SGSH mice.
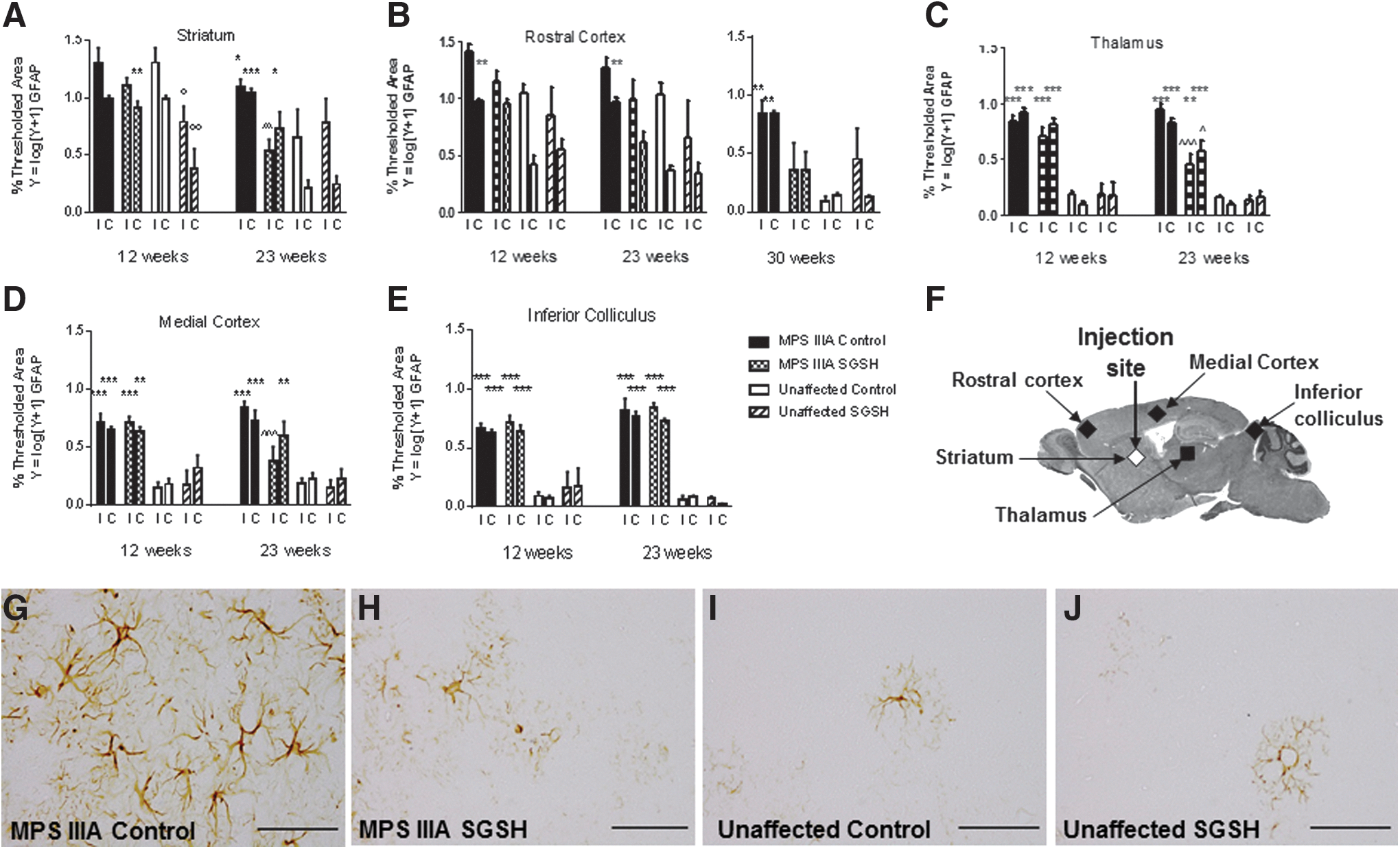
GFAP immunohistochemistry.
Isolectin B4
Statistically significant elevations in the number of isolectin B4 (IB4)-stained activated microglia were observed in MPS IIIA control mice compared with unaffected control mice in all brain areas and at each time point assessed (Fig. 6A–F and Supplementary Fig. S5). AAV-SGSH treatment of MPS IIIA mice resulted in a significant reduction in the number of activated microglia in the striatum and rostral cortex of the injected hemisphere at 12 weeks of age (compared with MPS IIIA controls); however, no reductions were observed in other brain regions assessed at this time point. By 23 weeks of age, significant reductions in the number of activated microglia were observed in the striatum, rostral cortex, and thalamus of both hemispheres (with highly significant reductions seen in the injected hemisphere), as well as in the medial cortex of the injected hemisphere (data not shown). The impact of AAV-SGSH treatment on the number of activated microglia in MPS IIIA mouse rostral cortex was maintained to 30 weeks in the injected hemisphere. AAV-SGSH treatment did not, however, reduce the number of IB4-stained microglia in the dentate gyrus of the hippocampus or the inferior colliculus in either hemisphere at either time point.
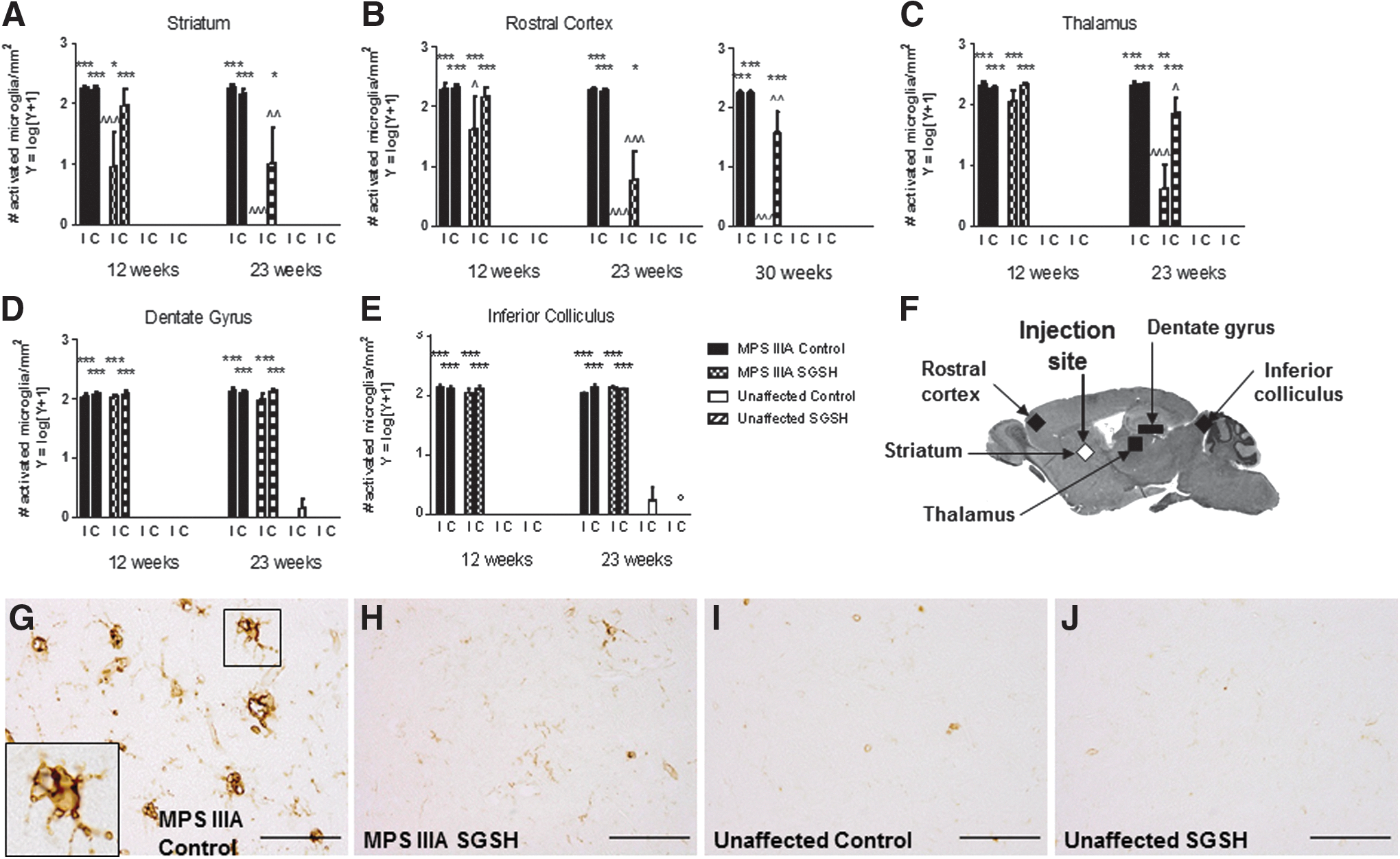
Isolectin B4 histochemistry.
Ubiquitinylated proteins
Ubiquitin-positive lesions have previously been reported in MPS IIIA mouse brain. 32 Here, we observed these inclusions in the tissues from MPS IIIA control mice (Supplementary Fig. S6A–E and S7). The number of lesions increased with age and AAV-SGSH treatment significantly reduced lesion number, particularly in the 23-week-old cohort. In the substantia innominata of the injected hemisphere (a region close to the injection site), a statistically significant reduction in the number of ubiquitin-positive inclusions was observed at both 12 and 23 weeks of age; a treatment effect was also seen in the contralateral hemisphere at 23 weeks of age. As these lesions are apparent by approximately 9 weeks of age in this brain region (unpublished observations), AAV-SGSH treatment from 5 weeks of age appears to have slowed but not completely prevented lesion formation. The halt in further lesion formation from 12 to 23 weeks of age also appears to have occurred in the periaqueductal gray and brainstem (spinal trigeminal region). Ubiquitin-positive lesions first appear in these regions at 4 and 6 weeks of age, respectively (unpublished observations).
Markers of autophagy
p62 binds LC3 in order to target proteins for autophagic degradation, and may target ubiquitinated proteins to this system as well. 33 Dark p62-stained intracellular inclusions were observed in the rostral cortex in MPS IIIA control mice from 12 weeks of age (Fig. 7A). Additional brain regions, such as the substantia innominata, inferior colliculus, and brainstem exhibited p62-positive inclusions from 23 weeks of age (Fig. 7B–D and Supplementary Fig. S8). Similar lesions were not seen in these areas in the unaffected mice. MPS IIIA-SGSH mice exhibited significantly fewer p62-positive inclusions in the rostral cortex at 23 and 30 weeks of age (compared with MPS IIIA control mice), but there was little change in lesion number in areas distant from the injection site.
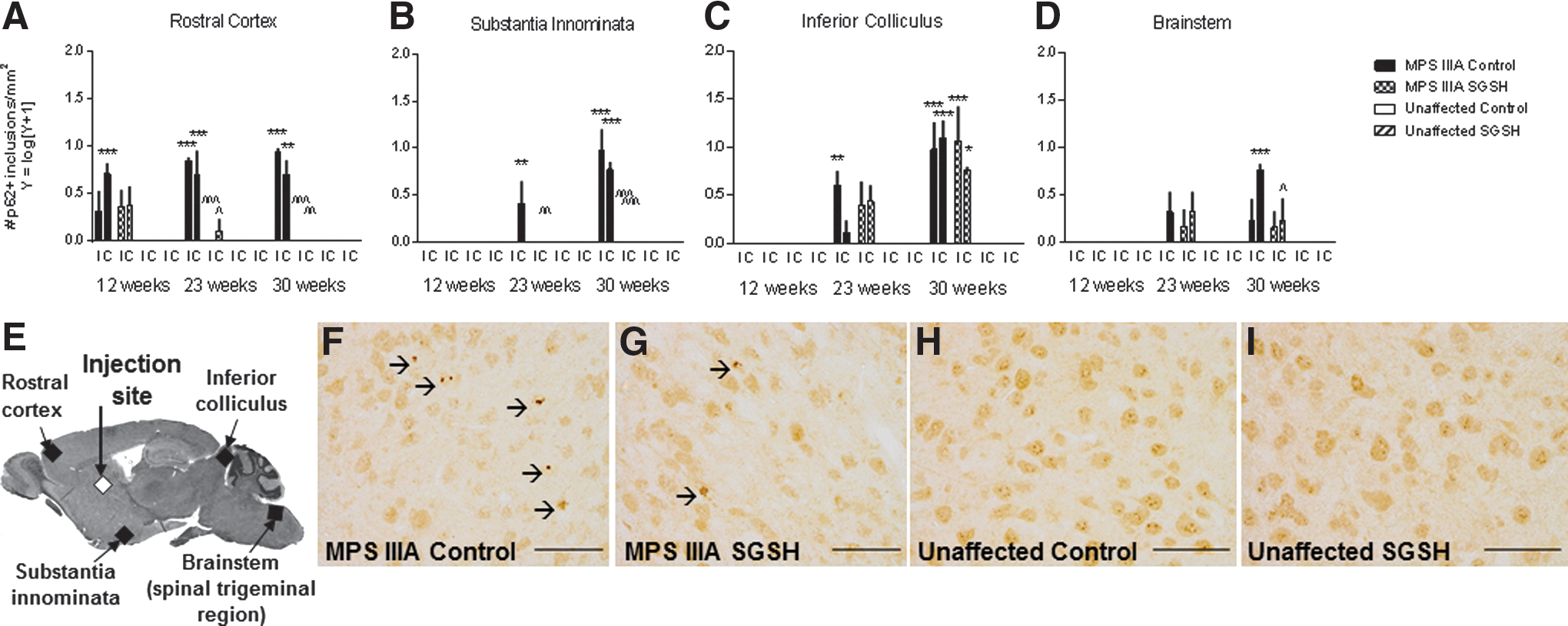
p62 immunohistochemistry.
Measurement of anti-SGSH antibodies
As shown in Supplementary Table S2, the sera from mice treated with AAV-SGSH did not contain anti-SGSH antibodies, when titers were compared with those in control mice.
Discussion
Previously, we administered an AAV2/5-based vector to newborn MPS IIIA mice and observed improvements in behavior. 14 Based on brain development, the time of instigation of that treatment is approximately equivalent to the second trimester of gestation in humans 34 and the outcomes are therefore considered as providing proof-of-principle. In the present study, we undertook an evaluation of AAV-SGSH treatment of young adult (5-week-old) MPS IIIA mice. The outcomes of the current study are more clinically relevant, given the fact that the condition is rarely diagnosed in utero.
The therapeutic potential of a gene therapy vector for use in a neurodegenerative LSD that exhibits brain-wide neuropathology relies on its ability to mediate diffuse lysosomal enzyme production on a long-term basis. Although very high amounts of active SGSH were found proximal to the injection region (brain slice L2), we did not observe brain-wide wild-type-level replacement of SGSH, although the enzyme was detected (although at lower protein/activity levels), in some more distant regions in the ipsilateral hemisphere, and in the contralateral hemisphere. Our experiments document SGSH expression up to 25 weeks postinjection (i.e., at 30 weeks of age); however, we note that AAVrh10-mediated galactocerebrosidase (GALC) expression was maintained until approximately 35 weeks postinjection in mice with Krabbe disease (8 months) 35 and expression of TPP-1 was observed at 54 weeks postinjection in LINCL mice. 36
Given the location of AAV-derived SGSH within the brain, not surprisingly, statistically significant improvements in both primary and secondary pathological changes were only seen in regions local to and/or in connection with the striatum. Reductions in primary storage of heparan sulfate-derived oligosaccharides were observed consistently in the pattern of L2 > L4 > R2 > R4, and the impact of enzyme delivery on primary lysosomal storage became more significant with time, when tissues were compared with those from MPS IIIA control mice. Similarly, amelioration of pathologically elevated endosomal/lysosomal membrane protein LIMP-II, secondarily stored GM3 ganglioside, and filipin-positive inclusions, in addition to neuroinflammatory markers, for example, isolectin B4, were observed and became more significant with time.
When ubiquitin-positive lesions were assessed, regional reductions were observed from 12 weeks of age (7 weeks postinjection) and it appeared that AAV-SGSH treatment of MPS IIIA mice halted further lesion formation. Detection of p62-positive inclusions typically occurred after that of ubiquitin-positive inclusions, potentially indicating an initial dysfunction in protein degradation at the proteasome. p62-positive inclusions in the rostral cortex and substantia innominata were responsive to treatment with time. These regions are local to or in connection with the injection site.
Our observations were somewhat unexpected given the brain-wide enzyme distribution and pathology improvements reported by Cressant et al., 15 who also used a unilateral injection strategy. In that study an AAV5 vector encoding human NaGlu, the enzyme defective in MPS IIIB, was administered to 6-week-old mice. 15 When mice were examined 16 weeks later, NaGlu was detected throughout the rostro-caudal axis, bilaterally in most mice (far beyond the region of transduced cells), with enzyme expression maintained to at least 38 weeks of age. Possible differences in enzyme distribution patterns between the current study and that of Cressant et al. 15 may be because of differences in the secretion of the respective enzymes and/or uptake by nontransduced cells.
Multiple-site intracerebral injection of AAVrh10-CLN2 in LINCL mice demonstrated widespread TPP-I enzyme distribution and resulted in improved survival. 36 Phase I trials of this therapy in children affected with LINCL are in the recruitment phase. Similarly, brain-wide expression of GALC significantly increased survival and improved behavioral/functional parameters in Krabbe mice that received both intracerebroventricular and intracerebellar injections of AAVrh10-GALC at birth. 35 On the basis of those studies and our own observations here, it is our firm belief that brain-wide high-concentration SGSH replacement will be needed in order for robust pathological/biochemical and clinical improvements to be achieved in MPS IIIA mice (and therefore patients), and that multiple-site injections of viral vector are required, even in the small murine brain.
An alternative to multiple-site injections may be convection-enhanced delivery (CED), a clinically relevant technique for CNS gene therapy, 37 which can enhance AAV vector distribution in brain. 38 Although AAV2 has a well-documented small volume of distribution compared with other AAV vectors, 39,40 administration of an AAV2 vector to as few as three sites using the CED regime provided broad transgene expression in the nonhuman primate brain. 41 The use of CED would be expected to significantly reduce the predicted 40–60 injection sites that have been suggested as being needed to provide global coverage (by conventional injection) of the infant brain with AAV1 vector-derived lysosomal enzyme. 42 AAVrh10-derived gene products have one of the largest volumes of distribution compared with other AAV vectors administered conventionally. 18 CED of AAVrh10 may therefore result in an enhanced vector/transgene distribution profile and require fewer injection sites, particularly if it was applied to brain regions with substantial axonal connectivity with other nuclei, for example, ventral tegmental area. 20,22
Other vector types under consideration for use in neurodegenerative LSD such as MPS IIIA include canine adenovirus, which has been assessed in MPS IIIA mice with transgene expression found to be short-lived, 26 potentially because of an immune response to the vector. A lentiviral vector has also been used to deliver an SGSH-SUMF1 construct bilaterally into the lateral ventricles of 6-week-old MPS IIIA mice. 43 The maximum SGSH activity detected in the lentivirus-injected slice appeared to be ∼43% of normal, substantially less than that observed in our study (average 16-fold increase above unaffected; data not shown). Similarly to the current study, histological improvements correlated with SGSH distribution; however, lentiviral-mediated SGSH distribution in the brain was less extensive.
AAV9 administered intravenously has been shown to cross the blood–brain barrier and provide widespread SGSH and NaGlu delivery in MPS IIIA and MPS IIIB mice, respectively. 44,45 An alternate regime involving intracerebroventricular administration of very high-dose AAV9-SGSH resulted in full-body correction of disease parameters in MPS IIIA mice 46 ; however, to achieve such favorable CNS results from intravenous delivery, significant systemic exposure to high doses of the vector is required, which may increase the likelihood of carcinogenic adverse events. 47 Finally, intravenous administration of AAV8-SGSH (or AAV8-mod SGSH, where the transgene had been modified to enable blood–brain barrier transmission) has also been shown to mediate significant improvements in brain disease in MPS IIIA mice. 48,49
Although we did not observe an immune response to the SGSH transgene in mice (as determined by measurement of anti-SGSH antibodies in sera), this does not preclude an immune response to SGSH in patients or an immune response to the vector itself in either patients or mice. Preexisting immunity to particular AAV serotypes in patient populations risks AAV vector or transduced cell elimination. Naturally acquired human AAV immunity (AAV1, 2, and 5) does not confer immunity toward AAVrh10, 24 making this AAV serotype particularly relevant to human application and possible re-application if required. Although AAVrh10 is not expected to provoke immune system elimination of transgenic cells, immunosuppression should be considered in patients with null mutations to prevent a humoral immune response being mounted against the expressed transgene. 50
In summary, intraparenchymal AAVrh10-SGSH administration has therapeutic potential for use in MPS IIIA children; however, multiple-site administration of vector will be required to achieve widespread rhSGSH distribution. Multi-injection site studies in MPS IIIA mice are planned, along with phenotypic characterization of treated animals. The addition of a CED regime may improve the efficiency of vector/transgene distribution, thereby improving the therapeutic outcomes.
On the basis of data reported here, a phase I/II clinical trial using this vector construct in four MPS IIIA children was recently undertaken in France, and 1-year follow-up assessments are complete. 51 A total of 12 injections were made in the brain, delivering a total of 7.2 × 1011 vector genomes per child. Although no change in CSF levels of heparan sulfate was observed, imaging and behavioral test outcomes indicated that the treatment may have stabilized disease progression in some participants. The outcome of further follow-up studies will be of great interest to the field. A phase II/III clinical trial is now planned.
Footnotes
Acknowledgments
We thank Karen Aiach, Prof. Olivier Danos, and Celine Schiff (Sanfilippo Alliance) for their contributions to discussions regarding study design. Prof. Andrea Ballabio and colleagues (TIGEM, Italy) are acknowledged for provision of plasmids/cDNA used in vector construction, and we thank Dr. Neil Hackett and colleagues (Belfer Gene Therapy Core Facility, Cornell University) for constructing the vectors used in this study. We thank Hanan Hannouche for genotyping the mice and Dr. Maria Fuller for assistance with mass spectrometry. Funding support came jointly from not-for-profit organizations Alliance Sanfilippo (France), AFM–Telethon (France), and LDRU (Australia).
Author Disclosure
An international patent is held by J.J.H. and others for mammalian sulfamidase and genetic sequences encoding it, for use in the investigation, diagnosis, and treatment of subjects suspected of suffering from sulfamidase deficiency (U.S. Patent No. 5,863,782).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.