
Abstract
Mesenchymal stem cells (MSCs) represent a promising approach for the treatment of acute respiratory distress syndrome (ARDS). However, their low efficiency in homing to injured lung tissue limits their therapeutic effect. Prostaglandin E2 (PGE2) biosynthesis substantially enhances the inflammatory response of the tissue. Moreover, it also facilitates the migration of MSCs by activating the E-prostanoid 2 (EP2) receptor in vitro. Given these observations, it would seem reasonable that PGE2 might act as a chemokine to promote the migration of MSCs through activation of the EP2 receptor. Herein, we confirmed that PGE2 was significantly increased in lung tissue as a result of stimulation by LPS. In addition, we constructed a lentiviral vector carrying the EP2 gene, which was successfully transduced into MSCs (MSCs-EP2). Near-infrared imaging and immunofluorescence showed that compared with MSCs-GFP, MSCs-EP2 significantly enhanced MSC homing to injured lung tissue. Moreover, the diminished amounts of Evans blue in homogeneous lung parenchyma in vivo indicated, in comparison with MSCs-GFP, that MSCs-EP2 significantly decreased LPS-induced pulmonary vascular permeability. In addition, administration of MSCs-EP2 largely decreased the levels of interleukin-1β and tumor necrosis factor-α compared with that observed after administration of MSCs-GFP at both 24 and 72 hr. Our results suggested that treatment with MSCs-EP2 markedly enhanced MSC homing to damaged lung tissue and, in addition, improved both lung inflammation and permeability. Thus, MSCs and EP2 combination gene therapy could markedly facilitate MSC homing to areas of inflammation, representing a novel strategy for MSC-based gene therapy in inflammatory diseases.
Introduction
A
In experimental animal models following lung injury, mesenchymal stem cells (MSCs) have alleviated inflammation of the injured lung structure by homing to the injured tissue, releasing paracrine factors, and differentiating into lung epithelial cells. 6,7 A major challenge of the current MSC treatment strategies for ARDS is poor MSC retention in injured tissue, which limits its therapeutic efficacy. Several studies have established that facilitating MSC homing to injured tissue can incrementally benefit acute lung injury (ALI). 8,9 Thus, facilitating MSC homing to target tissue represents a promising therapeutic strategy for ARDS.
Prostaglandin E2 (PGE2) is the major product of arachidonic acid during inflammation via the action of cyclooxygenase and PGE2 synthases. 10,11 Nonsteroidal antiinflammatory drugs inhibit PGE2 synthesis, and it is thus designated as a potent proinflammatory modulatory molecule. 12 Emerging evidence now suggests that PGE2 has antiinflammatory properties. 13 These opposing effects are mediated by E-prostanoid (EP) 1, 2, 3, and 4 receptors. The synergistic effect of PGE2 on macrophages, dendritic cells, and T and B lymphocytes during an inflammatory insult depends on the activation of specific EP receptors. 14 In addition to its role in the pathogenesis of the inflammation reaction, PGE2 has been proposed to participate in the enhancement of hematopoietic stem cell (HSC) homing by increasing HSC CXCR4 mRNA and surface expression, as well as facilitating HSC migration to stromal cell-derived factor-1 (SDF-1) and homing to bone marrow. 15 Nevertheless, Yun and colleagues demonstrated that PGE2 induced increased MSC migration. 16 This study also showed that of the four EP receptors, EP2 was responsible for PGE2-induced MSC migration.
In the current study, we hypothesized that the combination of MSC transplantation and EP2 gene transfection may augment MSC homing to damaged lung tissue. This would result in supplementary benefits in the treatment of tissue injury brought about by lipopolysaccharide (LPS) in the mouse model system. Therefore, the aims of our study were to elucidate the homing capacity and the therapeutic effect of MSC-EP2 gene expression on LPS-induced lung injury.
Materials and Methods
Transduction of lentiviral vectors into MSCs
MSCs isolated from the bone marrow of C57BL/6 mice were obtained from Cyagen Biosciences (Guangzhou, China) as previously reported. 17 Cell surface markers (CD73, CD90, CD105, and CD45) of passage 3 MSCs were determined by flow cytometry. In addition, the differentiation ability of the MSCs to the chondrogenic, osteogenic, and adipogenic lineages was also characterized.
Lentiviral expression vectors (pLV-EP2-GFP and pLV-null-GFP) and packing plasmids (pGag/Pol, pReV, and pVSV-G) were purchased from GenePharma (Shanghai, China). NotI and BamHI restriction enzymes were used to construct plasmids expressing the target genes. Subsequently, human embryonic kidney 293T cells (Cell Bank of the Chinese Academy of Sciences, Shanghai, China) were cotransfected with lentiviral vectors and the other three packaging plasmids. High-titer recombinant EP2 or GFP lentiviral vectors were collected. MSCs from passage 4–6 were transduced with recombinant lentiviral vectors. MSCs carrying the GFP gene (GFP-MSCs) and MSCs carrying both the EP2 and GFP genes (EP2-MSCs) were harvested. We then evaluated the adipogenic differentiation ability of these MSCs after EP2 overexpression, as previously described. 18
Transduction efficiency was evaluated by detecting GFP expression using a fluorescence microscope and through flow cytometry. MSC EP2 expression was assessed by Western blotting and PCR. TRIzol (Invitrogen, Carlsbad, CA) reagent was used to isolate total RNA, and a first-strand cDNA synthesis kit (Thermo Scientific, Wilmington, DE) was used to synthesize cDNA. Levels of mouse EP2 mRNA and endogenous control glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were determined by reverse transcription-polymerase chain reaction (RT-PCR). The following primers were used: GAPDH (133 bp), sense 5′-ACAACTTTGGCATTGTGGAA-3′ and antisense 5′-GATGCAGGGATGATGTTCTG-3′; and EP2 (140 bp), sense 5′-GTGCTGCCTGTCATCTATGG-3′ and antisense 5′-GTGGCGTACAGCTGAAGGTA-3′. The conditions applied for amplification were as follows: 95°C for 5 min, 40 cycles of 95°C for 15 sec, 60°C for 20 sec, and 72°C for 40 sec, followed by gradual extension at 0.1°C/sec from 45°C to 95°C. All samples were analyzed in triplicate.
Transwell migration assay
We then assessed the migration ability of MSCs, MSCs-GFP, and MSCs-EP2, using Transwell chambers. Transwell inserts (pore size, 8 μm) were set up in a 24-well plate. MSCs, MSCs-GFP, or MSCs-EP2 were suspended in serum-free medium and adjusted to a density of 1 × 105 cells/ml; 100 μl of cell suspension was added to the upper chamber of the migration well. Serum-free medium and PGE2 were loaded into the lower chamber (PGE2, 1 μmol/liter). After a 6-hr incubation at 37°C, the cells from the top of the filter were excluded with a cotton swab, and the cells that had migrated into the lower chamber were fixed with 4% paraformaldehyde for 10 min and stained with crystal violet (Beyotime, Haimen, China) for 20 min. Cells in five random microscope fields were counted.
LPS-induced ALI mouse model
Male C57BL/6 mice (6 to 8 weeks old) were obtained from Southeast University (Nanjing, China). The mice were acclimatized under specific pathogen-free conditions for 1 week before the experiments. Lung injury was induced via the intratracheal administration of LPS (Sigma-Aldrich, St. Louis, MO) from Escherichia coli as previously described. 16 The concentration of PGE2 in lung homogenates from ALI and normal mice was estimated with an ELISA kit (Cusabio Biotech, Wuhan, China) 0, 2, 4, 6, 8, 10, and 12 hr after LPS administration. Four hours after LPS stimulation, mice were intravenously administered phosphate-buffered saline (LPS+PBS group), MSCs expressing GFP reporter (MSC-GFP group), or MSCs expressing EP2 (MSC-EP2 group); 5 × 105 cells suspended in 100 μl of PBS were injected via the tail vein. Mice that were not subject to LPS challenge were injected with PBS (NS+PBS group). The mice were sacrificed 24 or 72 hr after MSC administration. For survival assessment, 40 mice were randomly allocated to each group and observed for 168 hr. The animal experiments were approved by the Committee of Animal Care and Use of Southeast University.
Detection of engrafted MSCs in lung
Before being injected in vivo, MSCs-EP2 cells (5 × 105) were labeled with CellVue NIR815 dye (eBioscience, San Diego, CA). The lungs of six mice were excised 24 and 72 hr postinjection and imaged with a Maestro In Vivo optical imaging system (Cambridge Research & Instrumentation, Woburn, MA), and fluorescence in the regions of interest (ROIs) was imaged and quantified.
Lung tissue samples from the right lower lobe were used for immunofluorescence staining. Prepared sections were incubated for 12–16 hr at 4°C with anti-GFP primary antibody (diluted 1:100; Abcam, Cambridge, MA), followed by conjugation with secondary goat anti-mouse antibody (diluted 1:200; Abcam). The sections were subsequently stained with 4′,6-diamidino-2-phenylindole (Sigma-Aldrich) for 5 min to acquire images with a fluorescence microscope (Olympus, Tokyo, Japan).
Assessment of lung edema
Lung edema in mice from the NS+PBS, LPS+PBS, MSC-GFP, and MSC-EP2 groups (six mice in each group) was estimated on the basis of the lung wet weight-to-body weight ratio (LWW/BW), which was used to assess pulmonary microvascular permeability. 19
Measurement of Evans blue leakage
In the NS+PBS, LPS+PBS, MSC-GFP, and MSC-EP2 groups (six mice in each group), Evans blue (20 mg/kg; Sigma-Aldrich) in 80 μl of saline were injected via the tail vein. Thirty minutes later, the right ventricle was filled with 10 ml of heparinized saline to flush the intravascular dye out of the lungs. Complete perfusion was confirmed when all of the blood had been removed from the lungs. Right lung tissue (100 mg) was then immersed in formamide (Sigma-Aldrich) for 24 hr at 60°C. The extracted Evans blue was determined by spectrophotometry at 630 nm. 20
Lung histopathological assessment
Lung tissue samples from right upper lobe were fixed in 4% paraformaldehyde and embedded in paraffin. Sections (thickness, 5 μm) were incised from the paraffin block and stained with hematoxylin and eosin (H&E). A pathologist, blinded to the identity of the slides, used a light microscope to evaluate lung injury scores according to the following five criteria: edema, alveolar and interstitial inflammation, alveolar and interstitial hemorrhage, atelectasis, and hyaline membrane formation. 21
Measurement of inflammatory cytokines
Levels of proinflammatory cytokines tumor necrosis factor (TNF)-α, and interleukin (IL)-1β, and antiinflammatory cytokine IL-10, in the lung homogenates were evaluated by ELISA.
Statistical analysis
Results are presented as means ± SD. Data from multiple groups were analyzed by one-way analysis of variance (ANOVA) with Bonferroni's post hoc test. Differences between two groups were analyzed by unpaired Student t test. Kaplan–Meier curves were used to describe the survival rate of mice in each group, and log-rank tests were performed to analyze the significance of differences. p < 0.05 was considered statistically significant. All statistical analyzes were performed with SPSS 17.0 (IBM, Armonk, NY) and GraphPad Prism 6 (GraphPad Software, San Diego).
Results
MSCs are successfully transduced with recombinant EP2 plasmids
MSCs at passage 4 were stably transduced with one of the lentiviruses, expressing GFP alone or EP2 with GFP, over a range of multiplicities of infection (MOI); an MOI of 20 was considered adequate for downstream tests. The transduction efficiency in both the infections was estimated as 97% on day 7 (Fig. 1A and B) by GFP fluorescence. EP2 expression in MSC-EP2-transduced cells was assessed by RT-PCR and Western blotting. EP2 protein expression in EP2-MSCs was superior to that in MSCs that received empty vector (MSC-vec) and to that in control cells (Fig. 1C). Similarly, RT-PCR demonstrated an approximately 23-fold higher EP2 mRNA level (Fig. 1D; p < 0.01) than that in MSCs-GFP or control MSCs. Also, EP2 overexpression did not alter the adipogenic differentiation of the MSCs (Fig. 1E). Flow cytometric analysis revealed that MSCs expressed CD73 (98.9%), CD90 (98.3%), and CD105 (97.1%), whereas CD45 was negligibly expressed (0.2%). The results of flow cytometry are presented in Supplementary Fig. S1 (supplementary data are available online at
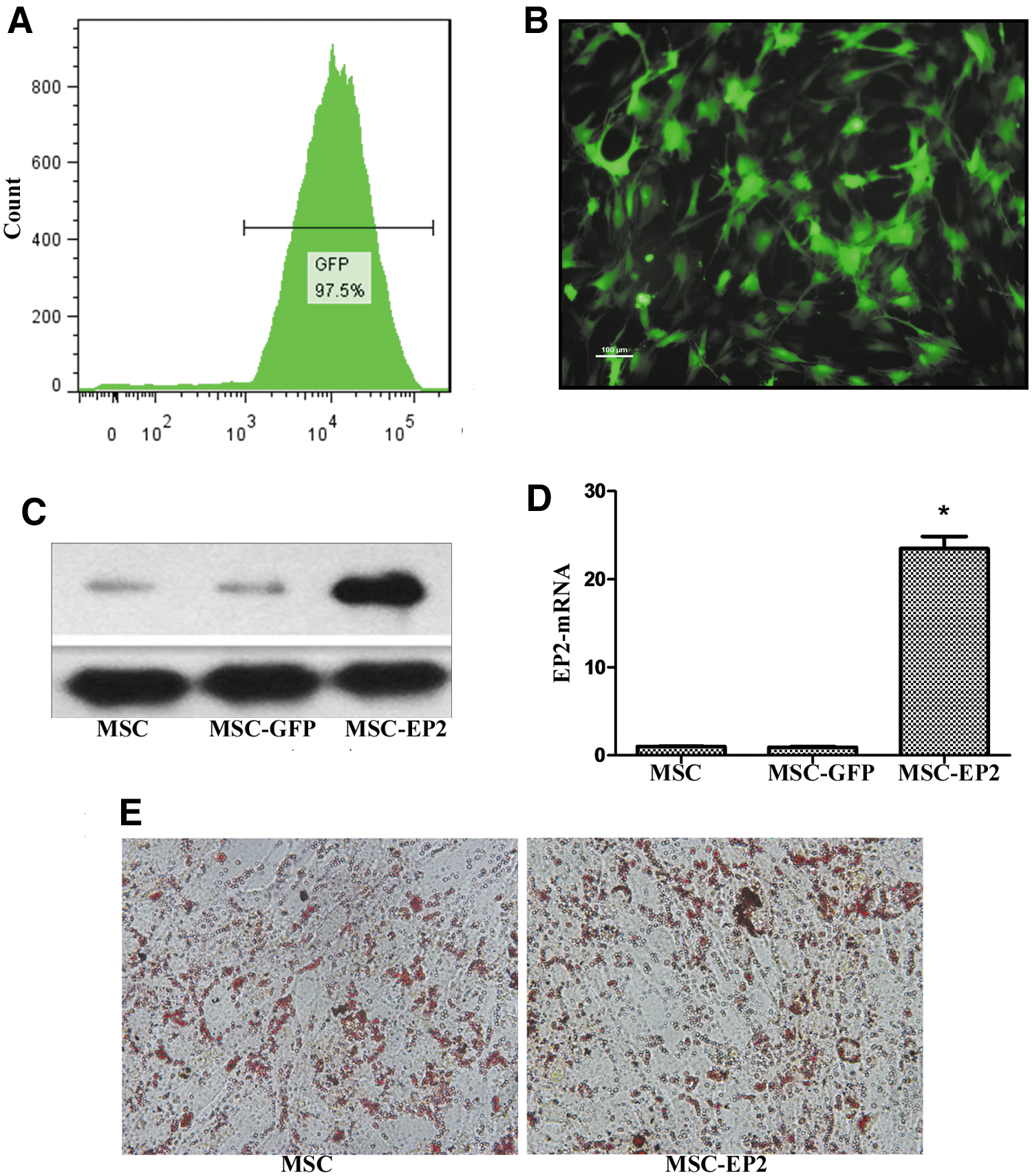
Transduction of E-prostanoid 2 (EP2) into mesenchymal stem cells (MSCs), using a lentiviral vector.
EP2 overexpression facilitates MSC migration to PGE2
The Transwell migration assay was done to assess the migratory response of MSCs to PGE2. In comparison with MSCs-GFP and control MSCs, MSCs-EP2 exhibited significantly more migration in response to PGE2 in vitro after 6 hr (Fig. 2; p < 0.01).
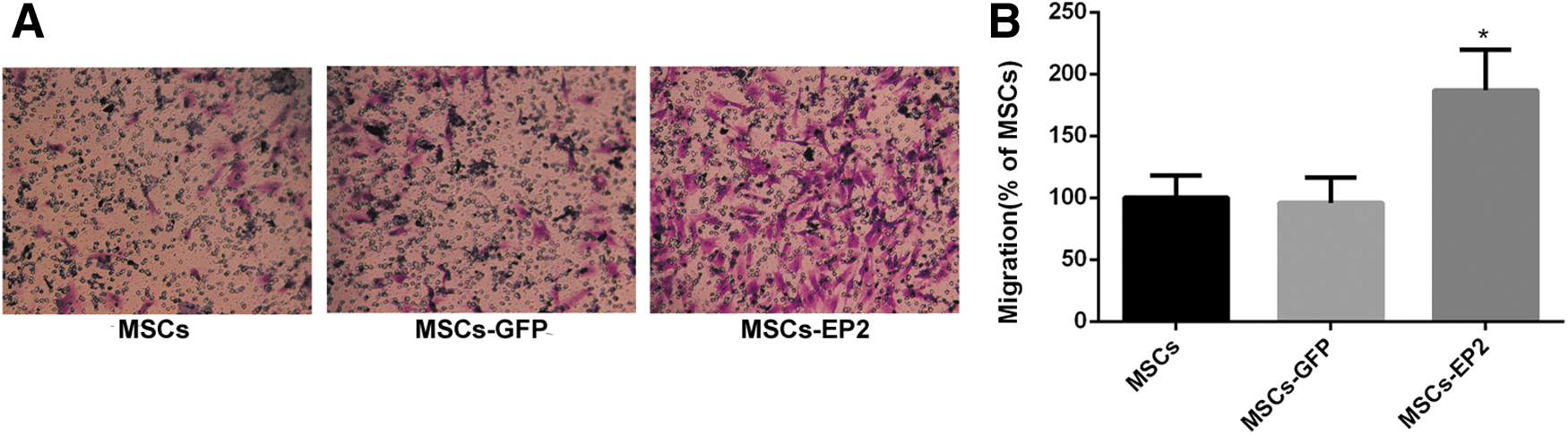
EP2 overexpression enhances the migration of MSCs in response to prostaglandin E2 (PGE2).
PGE2 is increased after LPS challenge
The concentration of PGE2 in lung homogenates from ALI mice and normal mice was measured by ELISA. PGE2 was significantly increased 2, 4, 6, 8, 10, and 12 hr after LPS administration (Fig. 3).
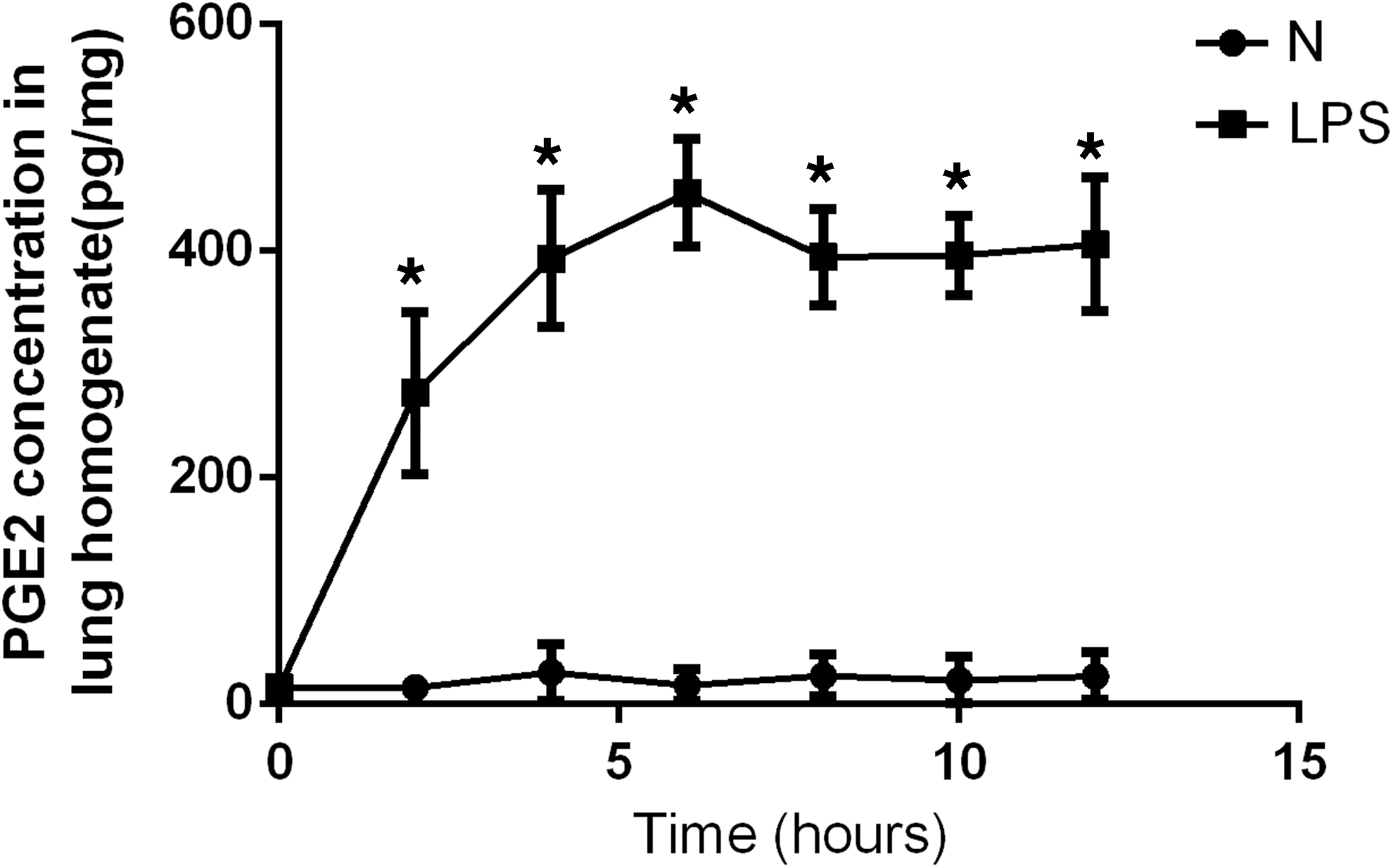
The concentration of PGE2 in lung homogenates from acute lung injury (ALI) mice and normal (N) mice at 2, 4, 6, 8, 10, and 12 hr. Data are expressed as means ± SD; *p < 0.01 versus group N.
MSCs-EP2 exhibit improved retention in lung
To evaluate the homing efficiency of implanted MSCs to injured lung tissue, MSCs-GFP and MSCs-EP2 were injected into mice via the tail vein. NIR (near-infrared) fluorescence images showed that fluorescence signals were detected in the lungs of four representative animals in each of the MSC-GFP and MSC-EP2 groups 24 and 72 hr after intravenous lentivirus-transduced MSC administration (Fig. 4A). The quantified fluorescence signals were significantly higher from the MSC-EP2 group as compared with the MSC-GFP group (six mice in each group) (see Fig. 4B).
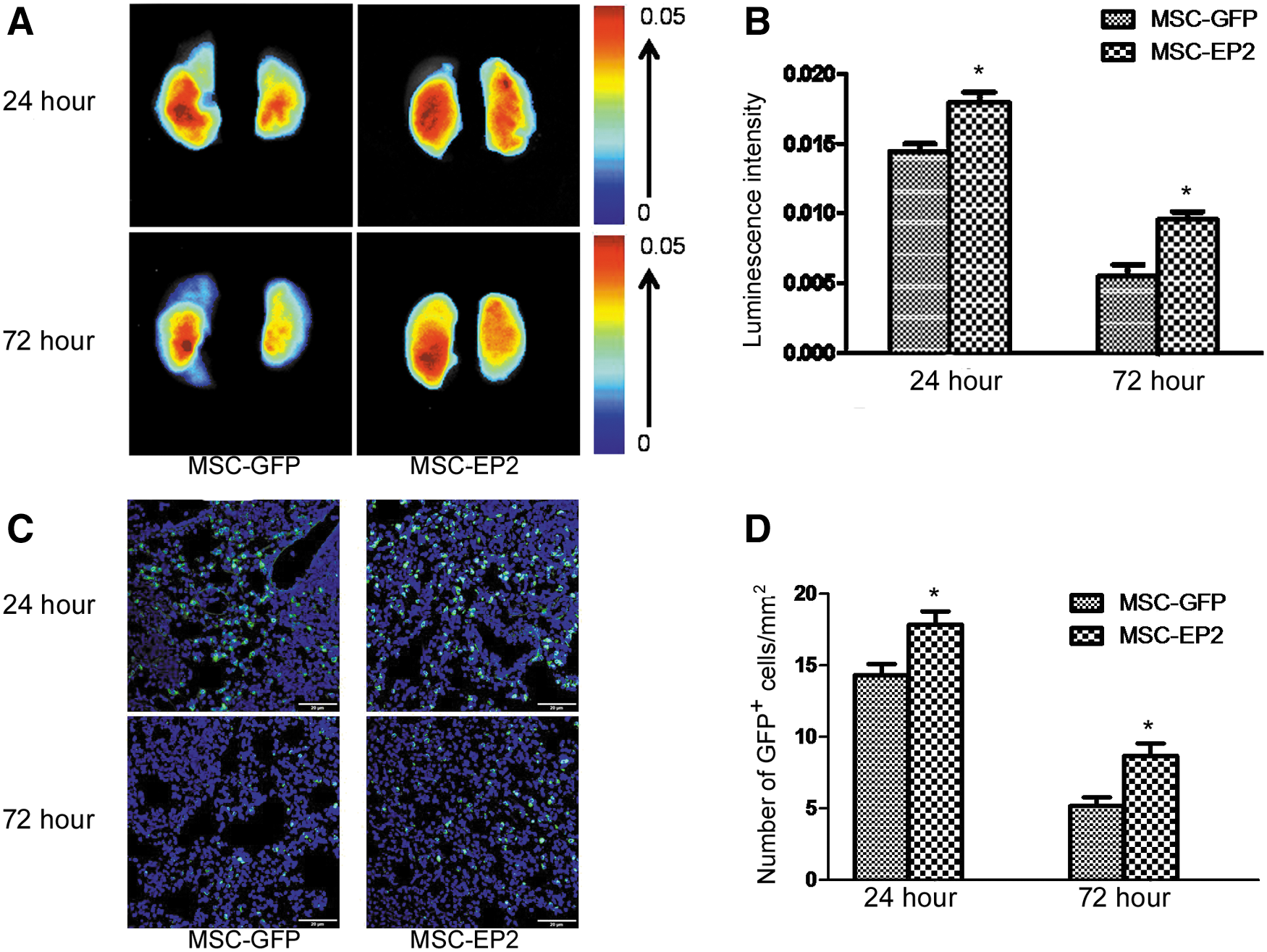
Detection of transplanted MSCs in injured lungs.
GFP-positive cells (green) observed by fluorescence microscopy in both the MSC-GFP and MSC-EP2 groups confirmed the retention of MSCs in the lung after tail vein injection. Representative images of four mouse lungs 24 and 72 hr after GFP-MSC and EP2-MSC administration are shown in Fig. 4C. Evaluation of engrafted MSC fluorescence further established that the number of GFP+ cells was significantly greater in the EP2-MSC group than in the GFP-MSC group at both 24 and 72 hr (Fig. 4D).
MSCs-EP2 reduce LPS-induced pulmonary vascular permeability
ALI is most commonly characterized by pulmonary edema. Animals treated with MSCs-EP2 significantly attenuated the increase in lung wet weight-to-body weight ratio as compared with the GFP conjugation 24 and 72 hr after MSC administration (Fig. 5A). Moreover, pulmonary vascular leakage in ALI was much lower in the MSC-EP2 group as compared with the MSC-GFP group 24 and 72 hr after MSC administration (assessed by Evans blue concentration) (Fig. 5B).

Effects of MSC-GFP and MSC-EP2 injections on pulmonary vascular permeability.
MSCs-EP2 improve lung histopathology
Lung sections in the LPS+PBS group were infiltrated with numerous granulocytes and macrophages. Strikingly, the alveolar walls also showed increased inflammation. However, the total severity of the lung histopathology was significantly improved after MSC-EP2 administration as compared with that of the MSC-GFP group. This indicated that MSCs-EP2 may have an additional beneficial effect on LPS-induced lung injury (Fig. 6).

Histopathological evaluation of the therapeutic potential of MSCs-EP2 in LPS-induced murine acute lung injury.
MSCs-EP2 alter inflammatory cytokine levels in injured lung tissues
Levels of IL-1β, TNF-α, and IL-10 were measured in lung homogenates. MSC-EP2 treatment markedly decreased IL-1β and TNF-α levels compared with MSC-GFP treatment, as observed at both time points, 24 and 72 hr. Conversely, the level of IL-10 was substantially increased under similar conditions (Fig. 7).
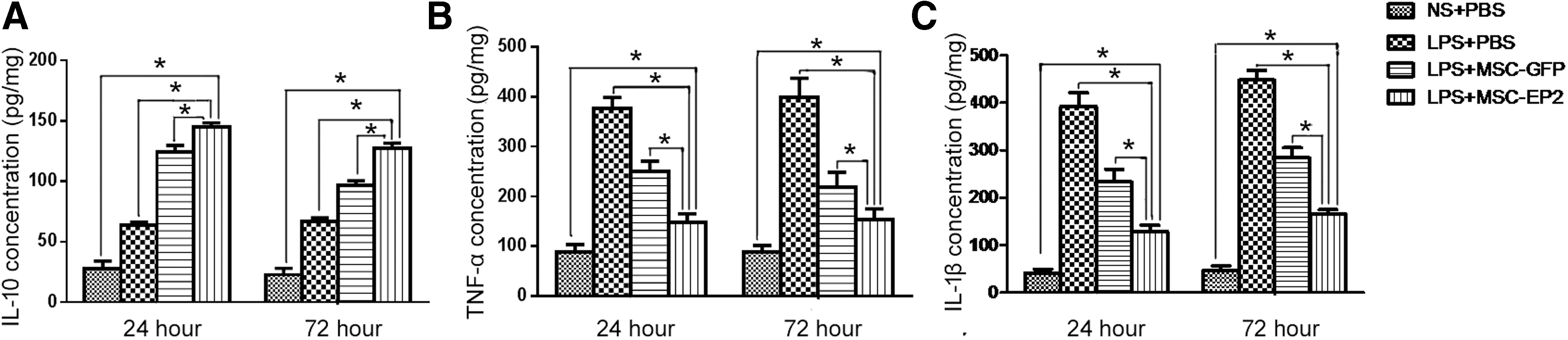
Assessment of lung inflammation after MSCs-EP2 administration.
EP2-MSC therapy improves mouse survival
Mice that received EP2-MSC treatment showed a greater, but statistically insignificant, survival rate at 168 hr than those that received GFP-MSC treatment (p > 0.05). Also, the survival rate in the LPS+PBS group was significantly lower than that in the GFP and EP2 groups (Fig. 8).
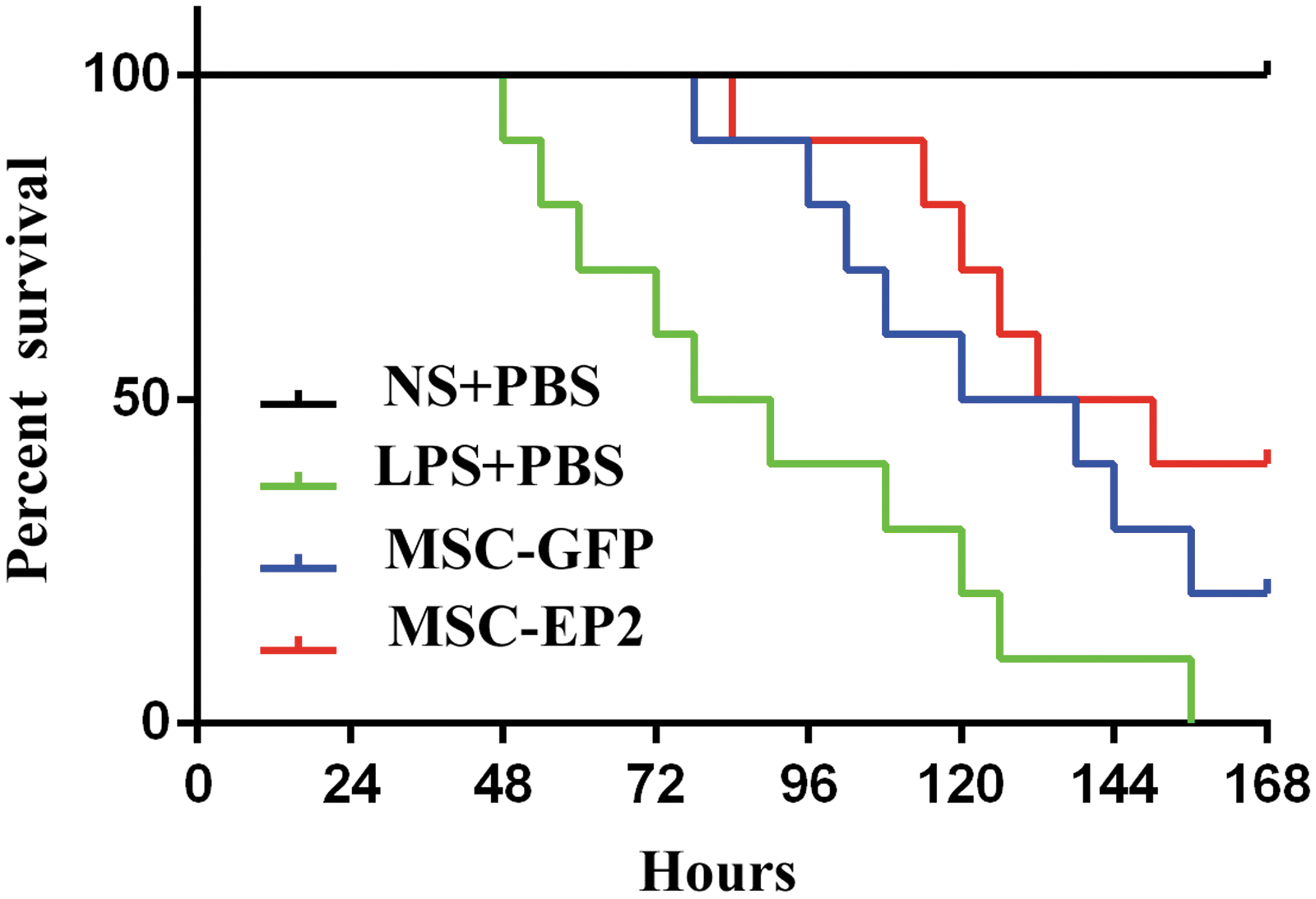
Kaplan–Meier survival curves. The survival rate at 168 hr was higher in mice that received MSCs-EP2 than in mice that received MSCs-GFP, but the difference was not statistically significant (p > 0.05). The survival rate was significantly lower in the LPS+PBS group than in the MSC-GFP and MSC-EP2 groups (p < 0.05).
Discussion
Herein, we demonstrated efficient transduction of MSCs with a recombinant lentivirus for stable EP2 gene expression without affecting the adipogenic differentiation of the cells. The in vivo data presented here suggested that the administration of EP2-overexpressing MSCs can significantly facilitate MSC homing to injured lung tissue. We also demonstrated that treatment with MSCs-EP2 improved both inflammation and vascular permeability, whereas MSCs-GFP exerted beneficial therapeutic effects to a lesser extent. In addition, we noticed a striking improvement in LPS-induced lung pathogenesis by MSCs-EP2 as compared with MSCs-GFP. Together, these results suggested that MSC-EP2 gene therapy is a potentially effective approach for treating ARDS in a clinical setting.
Enhancing MSC homing to target tissues is considered a potential means of achieving improved therapeutic effects via paracrine mechanisms. Accumulating evidence suggests that exogenously delivered MSCs preferentially home to sites of inflammation or injury. 22,23 However, the percentages of infused MSCs that are ultimately found in injured lung tissue are quite low; researchers have found that after MSC transplantation, only 5–9% of the transplanted MSCs migrate to the injured lung tissue. 24,25 Mounting evidence indicates that MSCs exert their therapeutic benefits mainly by secreting a variety of paracrine factors. 26,27 However, most paracrine cytokines have a relatively short half-life, and their beneficial effects are confined to the tissue within which they are released. Thus, by enhancing MSC homing to target tissues, better therapeutic effects could be achieved through the increased release of paracrine factors within the target tissues. 28 Several studies have shown that modifying MSCs before their systemic infusion improved the homing of MSCs to injured tissues and led to better functional outcomes. 29,30 Therefore, facilitating MSC migration to injured lung tissue would likely improve the efficacy of MSCs in ARDS treatment through the increased release of paracrine factors within the injured lung tissue. Moreover, one study has demonstrated that facilitating the homing of MSCs to injured lung tissue led to incremental therapeutic benefits. 31
As a response to tissue inflammation, prostaglandin E2 (PGE2) biosynthesis is upregulated to a large extent. Also, PGE2 has been shown to promote the migration of MSCs in vitro by stimulation of the EP2 receptor. Given these observations, it would seem reasonable that PGE2 might act as a chemokine to promote the migration of MSCs through activation of the EP2 receptor. PGE2 is one of the most abundant prostaglandin derivatives of arachidonic acid through the cyclooxygenase reaction. 32 To date, four receptors for PGE2 have been identified: EP1, EP2, EP3, and EP4. 33 PGE2 contributes to sustaining homeostatic functions and mediating pathogenic mechanisms via signaling through these four distinct receptors. 34 Moreover, significantly enhanced PGE2 biosynthesis plays a vital role in the inflammatory response. 35 In pulmonary inflammation, PGE2 originates from airway epithelial cells, smooth muscle cells, dendritic cells, and macrophages. Researchers have found that PGE2 exerts protective effects in allergen-induced airway inflammation. 36 Evidence suggests that PGE2 may potentially restrict inflammation via increased IL-10 production from macrophages. 37 Experiments conducted in vivo and in vitro have suggested that PGE2 can promote the migration of various types of cells, including monocytes, vascular endothelial cells, and tumor cells. 38 –40 In addition, one study has suggested that EP2 signaling is responsible for the ability of PGE2 to promote MSC migration. 16 Taken together, it seems reasonable that PGE2 might act as a chemokine in order to enhance MSC migration through EP2 receptor activation.
In this study, we first confirmed that PGE2 was significantly higher in lung tissue after LPS challenge. Subsequently, an EP2 lentiviral construct was successfully transduced into MSCs, and EP2 expression did not affect MSC proliferation or differentiation. Near-infrared imaging showed that MSCs-EP2 essentially enhanced MSC homing to damaged tissue of the lung structure as compared with MSCs-GFP. The effect was confirmed by immunofluorescence evaluation. In contrast, LPS-induced pulmonary vascular permeability was reduced by MSCs-EP2, based on assessment of Evans blue levels in the lung parenchyma. Also, the levels of IL-1β and TNF-α were greatly decreased at 24 and 72 hr.
The Transwell assay revealed that proportionally more MSCs-EP2 than MSCs-GFP migrated to PGE2, indicating that EP2 overexpression significantly facilitated MSC migration in response to PGE2. The mechanism underlying this protective effect of EP2 overexpression may be that PGE2 is a chemotactic factor for MSCs expressing the EP2 receptor, resulting in increased MSC homing to injured lung tissue as well as incremental improvements in lung repair after systemic injection when compared with the effects of control MSCs. Yang and colleagues demonstrated that the chemokine SDF-1, and the corresponding chemokine receptor 4 (CXCR4), were involved in MSC migration into injured lung tissue and that the overexpressed CXCR4 in MSCs largely improved lung repair in ALI rat models. 41 The increased homing of MSCs to injured lung tissue is the fundamental mechanism of this incremental beneficial effect. However, the precise mechanism is yet an enigma that necessitates further studies.
Our findings suggested that MSC-based EP2 gene delivery could be advantageous over the intravenous administration of MSCs. While the cell-based gene strategy is of significant clinical and economic importance, we have to acknowledge certain limitations of this study. First, CellVue might leak or continue to be detected as a dye on cell death; therefore, the presence of fluorescence may not indicate viable biodistribution. A further limitation is related to the use of lentiviral vectors, which might lead to biological hazards although the virus is referred to as a “suicide” lentivirus. Finally, the LPS-induced lung injury mouse model does not completely recreate the complex clinical ARDS condition in human patients. Thus, it is imperative to reproduce the findings in animal models of sepsis and pneumonia. Furthermore, in our study, transduction of EP2 gene into MSCs was performed prior to LPS-induced ALI mouse model, which calls into question the effectiveness of a posttreatment strategy and its relevance for clinical therapy. Further studies will be needed to address these limitations.
In conclusion, our results suggested that EP2-based treatment significantly enhances MSC homing to damaged lung tissue, improving lung inflammation and permeability. Furthermore, histopathological evaluation suggested that MSCs-EP2 markedly repaired lung injury compared with that observed after administration of MSCs-GFP. These results suggest that a combination of MSC and EP2 gene therapy could markedly facilitate MSC homing to sites of inflammation and represent a potential novel strategy for MSC therapy in inflammatory diseases.
Footnotes
Acknowledgments
The current work was funded by the National Natural Science Foundations of China (81201489, 81372093, 81571874, 81501705, 81300060, 81471843) and the Graduate Innovation Project in Jiangsu Province of China (KYZZ_0072).
Author Disclosure
The authors claim no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.