
Abstract
Liver fibrosis results in cirrhosis, liver cancer, and liver failure, which is a major cause of mortality worldwide. Gene therapy is a relatively new paradigm in medicine, with enormous therapeutic potential. The development of an efficient and safe delivery system is essential for clinical gene therapy. In the present study, we evaluated augmenter of liver regeneration/growth factor ERV1-like (ALR/GFER) gene therapeutic effect mediated by a novel minicircle vector (MC-hALR). The results in liver fibrotic rats that received MC-hALR through hydrodynamics-based transfection (HBT) for 8 weeks indicated that the minicircle DNA vector produced a more effective gene therapy effect than traditional plasmids (pcDNA3.1-hALR). Even when we reduced the treatment dose of MC-hALR to 30% (w/w) and the treatment frequency from weekly to biweekly, the in vitro and in vivo results still demonstrated that higher ALR gene expression significantly blocked increases in transforming growth factor-β1 (TGF-β1), platelet derived growth factor-BB (PDGF-BB), and α-smooth muscle aorta (α-SMA) levels; effectively suppressed the production of collagens, especially collagen I; and effectively alleviated liver injury and fibrosis in rats, thereby improving the survival rate of liver fibrotic rats. It is preliminarily concluded that the relative overexpression of MC-hALR inhibits the activation of hepatic stellate cells (HSCs), thereby alleviating liver fibrosis in rats.
Introduction
L
Gene therapy is a relatively new paradigm in medicine, with enormous therapeutic potential, and liver-targeted gene delivery has attracted a great deal of attention. 2 In addition to well-known growth factors, such as hepatocyte growth factor (HGF) and epidermal growth factor (EGF), augmenter of liver regeneration (ALR), also known as growth factor ERV1-like (GFER), is another cytokine of vital importance to liver. 3,4 ALR is a member of the newly discovered ALR/Erv1 protein family with FAD-linked sulfhydryl oxidase activity that catalyzes disulfide bond formation. 5,6 The CXXC motif of ALR is essential for cell survival 7 and the biogenesis of cytosolic Fe/S proteins. 8 Indeed, ALR has been demonstrated to function as a survival factor for hepatocytes, and the depletion of ALR protein by antisense oligonucleotides leads to hepatocyte cell death. 7 ALR promotes the growth of hepatocytes in the regenerating or injured liver, 3,7 plays a pivotal role as an immunoregulator and as an enzyme activator, 9,10 and protects hepatocytes against apoptosis. 4 Furthermore, various reports indicate that ALR appears to be a novel important cytokine with potential clinical significance in the gene therapy of liver fibrosis. 11,12
Nevertheless, one of the major barriers to ALR gene therapy is the safety and efficiency of the vectors used for gene transfer in eukaryotic cells. Compared with viral vectors, nonviral vectors are safe and simple to prepare and modify. 13 Plasmid DNA is frequently used for nonviral gene therapy, but the presence of CpG motifs and their associated sequences in bacterial DNA causes an immunotoxic response after the delivery of these plasmid vectors to mammalian hosts, which is a potential risk for safe clinical applications and reduces transgene expression. 14 Moreover, the results of our previous experiment demonstrated low-level and short-term expression in response to plasmid-mediated ALR gene therapy in rats, with large individual differences (unpublished). Therefore, the development of an efficient and safe delivery system is essential for ALR gene therapy. Minicircles (MCs), novel plasmid-based vectors that are composed solely of eukaryotic sequences do not carry either antibiotic resistance genes or bacterial origins of replication; thus, MCs induce markedly reduced immune and inflammatory responses and effectively protect transgene expression from being silenced because of the presence of bacterial backbone sequences. 15 The use of MCs is an interesting approach to increase the safety and efficiency of plasmid-based vectors for clinical gene therapy. 16
Therefore, in the present study, an ALR expression cassette in a minicircle DNA vector was prepared for the first time, the expression level of the ALR gene mediated by a minicircle DNA vector was compared with that of conventional plasmids both in vitro and in vivo, and the 8-week gene therapy effect in fibrotic rats was compared.
Materials and Methods
Parental plasmid construction
The human ALR gene expression fragment containing a CMV promoter, human ALR coding sequence (GenBank accession number: NM_005262), and BGH polyadenylation signal sequence was PCR-amplified with a forward primer (5′-GACTGAAGATCTATGTACGGGCCAGATATA-3′), reverse primer (5′-ATACCGCTCGAGGGTTCTTTCCGCCTCAGGA-3′), and PrimeSTAR Max DNA Polymerase (Takara), using the previously constructed pcDNA3.1-hALR 12 as a template. The amplified fragments were excised with BglII and XhoI (NEB) and then ligated between attB and attP sites (Fig. 1A, Parental Plasmid).
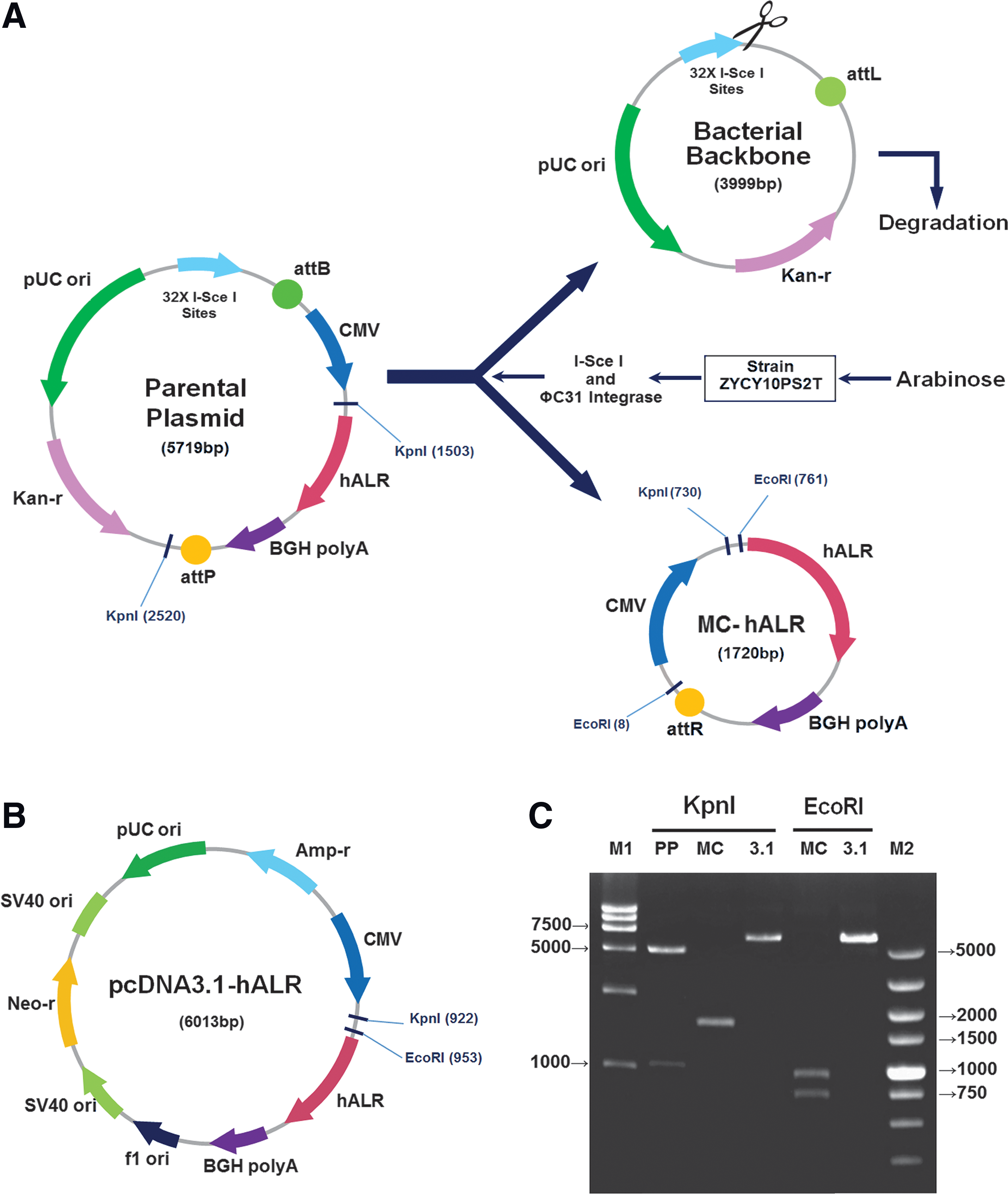
Comparison of minicircle- and pcDNA3.1-mediated human augmenter of liver regeneration (hALR) gene vectors.
Plasmid preparation
A previously reported protocol 17 for minicircle DNA vector production was used with some modifications. Briefly, Escherichia coli strain ZYCY10P3S2T (a kind gift from Dr. Zhiying Chen, Stanford University) was transformed with the parental plasmids. One transformant was picked and grown overnight (OD600 4.5–5.0) in 5 ml TB broth (Invitrogen) containing kanamycin (50 μg/ml) at 37°C with shaking at 250 rpm, followed by amplification with 0.1–0.3% of the inoculum in TB broth containing kanamycin (50 μg/ml) for 15 hr. Then, after the addition of a 1.2-fold volume of LB broth (Invitrogen) containing 0.03% L-arabinose (Sigma-Aldrich) and 3% 1 N NaOH, the incubation was continued for an additional 6 hr at 32°C with shaking at 250 rpm.
E. coli strain DH-5α (Invitrogen) was transformed with pcDNA3.1-hALR. One transformant colony was picked and grown overnight (OD600 4.5–5.0) in 5 ml LB broth (Invitrogen) containing ampicillin (50 μg/ml) at 37°C with shaking at 250 rpm.
All of the plasmids were purified using an E.N.Z.A. Endo-Free Plasmid Maxi Kit (Omega) according to the manufacturer's instructions.
Cell culture and transfection
The human hepatoma cell line SMMC-7721 and rat hepatic stellate cell line HSC-T6 were obtained from the Cell Bank of Type Culture Collection Chinese Academy of Sciences (Shanghai, China) and cultured in Dulbecco's modified Eagle's medium (DMEM; GIBCO) containing 10% fetal bovine serum (FBS; GIBCO) at 37°C in 5% CO2.
The SMMC-7721 and HSC-T6 cells (5 × 105 cells/well) were cultured in 6-well plates for 24 hr before transfection. The cells were transfected with 2.5 μg pcDNA3.1-hALR or 0.75 μg MC-hALR per well using X-tremeGENE HP DNA Transfection Reagent (Roche) according to the manufacturer's protocol. The cells were harvested 36 or 72 hr after transfection. At least three independent cultures were performed.
Animal model of liver fibrosis
Male Sprague-Dawley rats, weighing 120 ± 10 g, were obtained from the Medical Laboratory Animal Center of Guangdong (Guangzhou, China). All animals were maintained in a standard laboratory of 458th Hospital of PLA with a 12 hr light–12 hr dark cycle and free access to water and food. All animal procedures were conducted in accordance with the Declaration of Helsinki and with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the U.S. National Institutes of Health.
The animal model of liver fibrosis was described previously 18 and used with some modifications. Briefly, beginning 2 weeks before treatment with CCl4, the rats were allowed unlimited access to standard laboratory chow and drinking water containing 35 mg/dl phenobarbital (Sigma-Aldrich) and then received ∼400 mg CCl4/kg BW in olive oil (0.08 ml CCl4/ml olive oil) intragastrically twice a week for 5 weeks.
Expression of pcDNA3.1-hALR versus MC-hALR in liver fibrotic rats
Liver fibrotic rats (n = 44) were randomly assigned to the pcDNA3.1-hALR group (n = 18), MC-hALR group (n = 21), or control group (n = 5) and were treated by the hydrodynamics-based transfection (HBT) method. 19 More specifically, the animals were injected via the tail vein with pcDNA3.1-hALR (500 μg/kg BW), MC-hALR (150 μg/kg BW), or sterile saline in a volume equal to 5.25% (v/w) body weight and over a period of 5–8 sec using a 22 G injection needle. All of the fibrotic rats were dosed intragastrically twice a week with ∼200 mg CCl4/kg BW until the end of the experiment.
On days 3, 6, 9, 12, 15, and 18, three rats in the MC-hALR group were sacrificed; the other 3 rats were treated with MC-hALR again on day 15 and sacrificed on day 18. On days 1, 3, 5, 7, 9, and 11, three rats in the pcDNA3.1-hALR group were sacrificed. All of the control rats were sacrificed within 4 hr of injection with sterile saline. The right lobes of the rat livers were collected and preserved at −80°C for Western blot and quantitative real-time PCR analysis.
ALR gene therapy for liver fibrosis
The protocol of ALR gene therapy for liver fibrosis is shown in Table 1. The healthy rats (n = 20) were randomly divided into two groups: the healthy control group (group I, n = 10) and the HBT method control group (group II, n = 10) treated with sterile saline by the HBT method described above. The fibrotic rats (n = 60) were randomly assigned to four groups: the liver fibrosis model group (group III, n = 15) dosed with sterile saline, the pcDNA3.1-hALR group (group IV, n = 15) treated with pcDNA3.1-hALR (500 μg/kg BW) every week, the MC-hALR low-dose group (group V, n = 15) treated with MC-hALR (150 μg/kg BW) every 2 weeks (biweekly), and the MC-hALR high-dose group (group VI, n = 15) treated with MC-hALR (500 μg/kg BW) every 2 weeks (biweekly). All of the fibrotic rats were treated by the HBT method described above and were dosed intragastrically with ∼200 mg CCl4/kg BW twice a week until the end of the experiment. After an 8-week treatment, all of the animals were sacrificed on day 60 (day 4 after last treatment); serum samples were collected to measure biochemical parameters; and the right lobe of the livers was taken for histopathology, immunohistochemistry, Western blot, and quantitative real-time PCR analyses.
The protocol of ALR gene therapy for liver fibrosis
The fibrotic rats were dosed intragastrically with ∼200 mg CCl4/kg BW twice a week until the end of the experiment.
ALR, augmenter of liver regeneration; HBT, hydrodynamics-based transfection; NS, normal saline.
Serum biochemical parameters
Blood samples were drawn from the inferior vena cava of each rat anesthetized with sodium pentobarbital (65 mg/kg, i.p.; Sigma-Aldrich) on day 60. After coagulation, the serum was obtained by centrifugation of the whole blood at 1000 g for 15 min and was stored at −20°C for further analysis. The biochemical levels, including alanine albumin (ALB), aminotransferase (ALT), aspartate aminotransferase (AST), and alkaline phosphatase (ALP) levels, were assayed using an automatic biochemical analyzer (Cobas 6000; Roche).
Western blotting
Western blot analysis was performed as described previously. 20 Briefly, the protein concentration was quantified using a BCA protein assay kit (Pierce). The samples were probed with primary rabbit anti-ALR antibody (Proteintech; ∼23 kDa) diluted 1:1000, rabbit anti-PCNA antibody (Santa Cruz; ∼30 kDa) diluted 1:2500, or rabbit anti-β-actin antibody (Santa Cruz; ∼45 kDa) diluted 1:8000. The immune complexes were visualized using goat antirabbit IgG secondary antibody (Bioworld) diluted 1:8000, followed by a chemiluminescent reagent (Millipore), and the complexes were then detected on photographic film. Densitometric analysis of the protein levels was performed with ImageJ 1.44p software (Wayne Rasband, National Institutes of Health).
Histopathology and immunohistochemistry
The liver was fixed with 4% paraformaldehyde overnight and then embedded in paraffin, after which tissue sections (5 μm) were obtained using a microtome (Leica, Germany) and were processed by routine procedures. For Sirius red staining, the sections were stained with 0.1% Sirius red in picric acid (Sigma-Aldrich) for 1 hr at room temperature (RT); for hematoxylin and eosin (H&E) staining, the sections were stained with H&E (Sigma-Aldrich); for immunohistochemistry, the sections were incubated in 3% hydrogen peroxide for 10 min at RT, subjected to immunostaining with primary rabbit anti-ALR antibody (Proteintech) diluted 1:100 overnight at 4°C, measured to determine the protein activity using the EnVision horseradish peroxidase rabbit/mouse reagent kit (DAKO K5007), and then washed and counterstained with hematoxylin. Afterward, all of the sections were washed and mounted in neutral balsam.
Quantitative real-time PCR
The ALR, TGF-β1, PDGF-BB, α-SMA, COL1A1, COL3A1, COL4A1, and β-actin (endogenous control) transcript levels were quantified using quantitative real-time PCR technology. The total RNA was prepared from cells or tissues using TRIzol Reagent (Invitrogen). cDNA for each RNA sample was synthesized using a miScript Reverse Transcription Kit (Qiagen). Quantitative real-time PCR was performed using a miScript SYBR Green PCR Kit (Qiagen) with a real-time PCR System (CFX96; Bio-Rad) according to the manufacturer's instructions. The amplification step consisted of 44 cycles of 94°C for 10 sec, 60°C for 20 sec, and 72°C for 30 sec. Relative quantification was performed according to the ΔΔCt method.
The corresponding PCR primer sequences were as follows: ALR forward primer, 5′- ATACGCTGGCCGCCTATTAC-3′; ALR reverse primer, 5′-TCTGGCTGGCTCCTGTCTAT-3′; TGF-β1 forward primer, 5′-AGGGCTACCATGCCAACTTC-3′; TGF-β1 reverse primer 5′- CCACGTAGTAGACGATGGGC-3; PDGF-BB forward primer, 5′-ATGACCCGAGCACATTCTGG-3′; PDGF-BB reverse primer 5′-ACACCTCTGTACGCGTCTTG-3′; α-SMA forward primer, 5′-GGAGTGATGGTTGGAATGG-3′; α-SMA reverse primer 5′- GGTGATGATGCCGTGTTC-3′; COL1A1 forward primer, 5′-GCGAAGGCAACAGTCGATTC-3′; COL1A1 reverse primer 5′-ACCAGATTGGGATGGAGGGA-3′; COL3A1 forward primer, 5′-TTCCTGGGAGAAATGGCGAC-3′; COL3A1 reverse primer 5′-CCAGGATAGCCACCCATTCC-3′; COL4A1 forward primer, 5′- TCTGCTCTATGTCCAAGG-3′; COL4A1 reverse primer 5′- CAGCCAGTAGGAGTAGTC-3′; β-actin forward primer, 5′-CACCCGCGAGTACAACCTTC-3′; β-actin reverse primer 5′-CCCATACCCACCATCACACC-3′.
Statistical analysis
Each of the experimental values was compared with its corresponding control value using SPSS 17.0 software (SPSS Inc.). Continuous variables are presented as the mean ± standard deviation (SD) and were analyzed using one-way analysis of variance (ANOVA), followed by post hoc tests. Survival was analyzed using Kaplan–Meier curves, with differences evaluated by the log-rank (Mantel–Cox) test. p < 0.05 was considered statistically significant.
Results
Preparation and amplification of MC-hALR with relatively high quality and yield
As shown in Fig. 1A, the human ALR gene expression fragment containing a CMV promoter, human ALR cDNA (Gene Bank Accession Number: NM_00526), and the BGH polyadenylation signal sequence was ligated between the attB and attP sites to construct the human ALR gene parental plasmid (5719 bp). For high quality and yield, the protocol for MC-hALR production was modified slightly according to a previously described method.
17
The inoculum size was increased from 0.025% to 0.1% for amplification; for production, instead of an equal volume of LB broth, a 1.2-fold volume of LB broth containing 0.06% L-arabinose and 3% 1 N sodium hydroxide was added to the cultures, which generated a product with relatively high yield (9–11 mg/liter). Analysis by agarose gel electrophoresis indicated that almost no parental plasmid or bacterial backbone remained in the culture (Fig. 1C). MC-hALR and pcDNA3.1-hALR in their supercoiled and linear forms were prepared (Supplementary Fig. S1; Supplementary Data are available online at
High-level expression of ALR in SMMC-7721 and HSC-T6 cell lines transfected with MC-hALR
To compare the ALR gene expression levels in vitro, the human hepatoma cell line SMMC-7721 and rat stellate cell line HSC-T6 were cultured and transfected with MC-hALR or pcDNA3.1-hALR. The ALR was measured by Western blotting and quantitative real-time PCR (not shown) at 36°C or 72 hr after transfection. As shown in Fig. 2A, at 36 hr, the relative ALR protein levels in SMMC-7721 and HSC-T6 cells treated with MC-hALR were ∼5-fold and ∼4-fold higher (p < 0.001), respectively, than those in cells treated with pcDNA3.1-hALR; at 72 hr, the expression levels were more than 3-fold higher (p < 0.001). Furthermore, there was no statistically significant differences (p > 0.05) in the relative ALR protein level between the pcDNA3.1-hALR group and the control group in either cell line at 36 hr, indicating the knockdown pcDNA3.1-hALR expression level in 36 hr after transfection. By contrast, MC-hALR showed higher and more sustained expression level in 72 hr after transfection despite using one-third (w) of pcDNA3.1-hALR transfection dose.
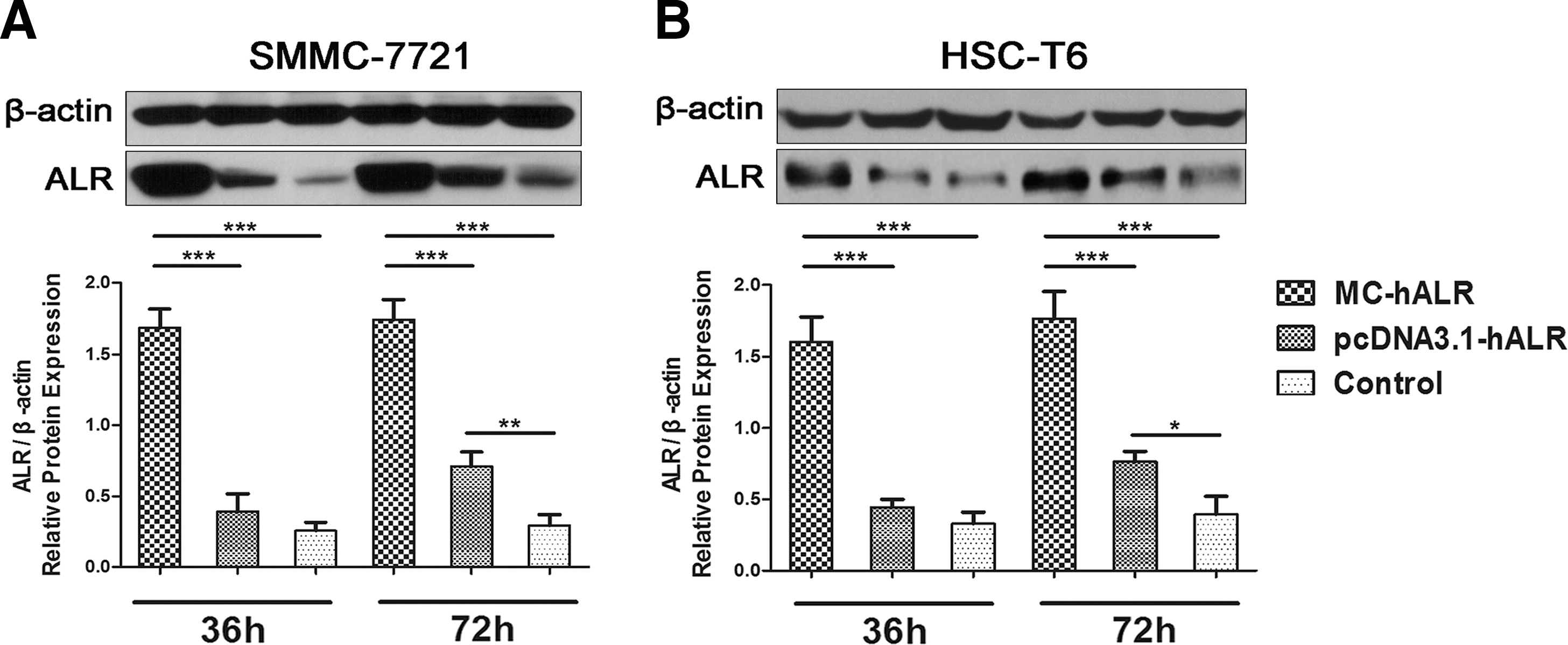
Comparison of MC-hALR and pcDNA3.1-hALR expression in the SMMC-7721 and HSC-T6 cell lines.
Analysis of ALR gene expression in fibrotic rats treated with MC-hALR by HBT
To compare the ALR gene expression in vivo, animal experiments were performed. The dose of pcDNA3.1-hALR was 500 μg/kg BW, as described previously, 12 and the dose of MC-hALR was reduced to150 μg/kg BW. All the liver fibrotic rats were treated only once.
The relative ALR protein expression levels over time are shown in Fig. 3A, and the densitometry results from analysis using ImageJ 1.44p software are shown in Fig. 3B. The relative ALR protein level in the MC-hALR group reached a maximum value at day 3 and was sustained at a level ∼2-fold higher than that in the pcDNA3.1-hALR group until day 9. The ALR expression declined to baseline levels on day 18 for the MC-hALR group but on day 11 for the pcDNA3.1-hALR group. Three fibrotic rats were treated with MC-hALR again on day 15, and the protein level increased significantly (p < 0.001 vs. day 0) on day 18. Taken together, these results clearly demonstrate that, compared with conventional pcDNA3.1-hALR, in vivo MC-hALR delivery by HBT led to ALR detection possible for longer period because of initial high level, and they provide evidence supporting the following experiments in ALR gene therapy.

Comparison of ALR level in livers of fibrotic rats treated with MC-hALR and pcDNA3.1-hALR.
Analysis of therapeutic effects in fibrotic rats treated with MC-hALR by HBT for 8 weeks
To further substantiate the antifibrotic effects of the ALR gene in vivo, 80 rats were used: 20 healthy rats divided into 2 groups (groups I and II) and 60 fibrotic rats divided into 4 groups (groups III–VI). The treatment protocol is presented in Table 1.
As shown in Fig. 4A, the liver morphology of the rats in the liver fibrosis model group (III) showed a rough and granulated liver surface, and H&E staining revealed that the normal structure of the hepatic lobule was markedly damaged, with massive neutrophil infiltration. By contrast, the liver morphology of rats in the ALR treatment groups (IV, V, and VI) showed a relatively smooth liver surface, especially for the MC-hALR high-dose group (VI), and H&E staining revealed less necrosis and inflammation in the liver tissue of rats in the MC-hALR groups (V, VI), although necrosis and neutrophil infiltration were still observed for pcDNA3.1-hALR group (IV).

Analysis of the therapeutic effects in fibrotic rats treated with MC-hALR or pcDNA3.1-hALR for 8 weeks.
As shown in Fig. 4B, the ALT, AST, and ALP values were lower in the MC-hALR groups (V, VI) than in the pcDNA3.1-hALR group (IV). The survival rate improved for the MC-hALR low-dose group (V) was higher (p < 0.05 vs. group III), and the survival rate was 100% for the MC-hALR high-dose group (VI) (p < 0.01 vs. group III), with a statistically difference between group VI and group IV (p < 0.05) (Fig. 4C).
These findings indicate that the results for MC-hALR were superior to those obtained using pcDNA3.1-hALR; minicircle DNA vector was able to induce the expression of the functional ALR protein to protect hepatocytes and ameliorate liver fibrosis, and exhibited better therapeutic effects against liver fibrosis even when we reduced the treatment dose of MC-hALR from 500 to 150 μg/kg BW and the treatment frequency from weekly to biweekly.
The expression of the ALR gene in fibrotic rats treated with MC-hALR by HBT for 8 weeks
To further research the relationship between the expression and therapeutic effect in fibrotic rats treated with the ALR gene over an extended time period, the ALR liver expression was evaluated using immunohistochemistry and Western blotting. The result of immunohistochemistry was shown in Fig. 5A; in the ALR treatment groups (VI, V, and VI), some cells expressed the ALR protein (massive brown spots) around the portal area. To quantify the result shown in Fig. 5A, the relative IOD was analyzed by IPP 6.0 software (Fig. 5B). The expression level in the MC-hALR high-dose group (VI) was ∼2-fold higher than that in the pcDNA3.1-hALR treatment group (IV) (p < 0.001), which was consistent with the result of Western blotting (Fig. 5C). Western blot analysis of relative ALR protein expression levels in the heart, lungs, liver, and kidneys was shown in Fig. 5D. After 8 weeks of treatment with MC-hALR by HBT, ALR proteins of ∼23 kDa were expressed at the highest level in the liver (p < 0.0001), somewhat in the kidneys (p > 0.05), and at barely detectably levels in the heart and lungs (p > 0.05) relative to those in the healthy control, indicating the relative overexpression of MC-hALR in liver using the HBT method.
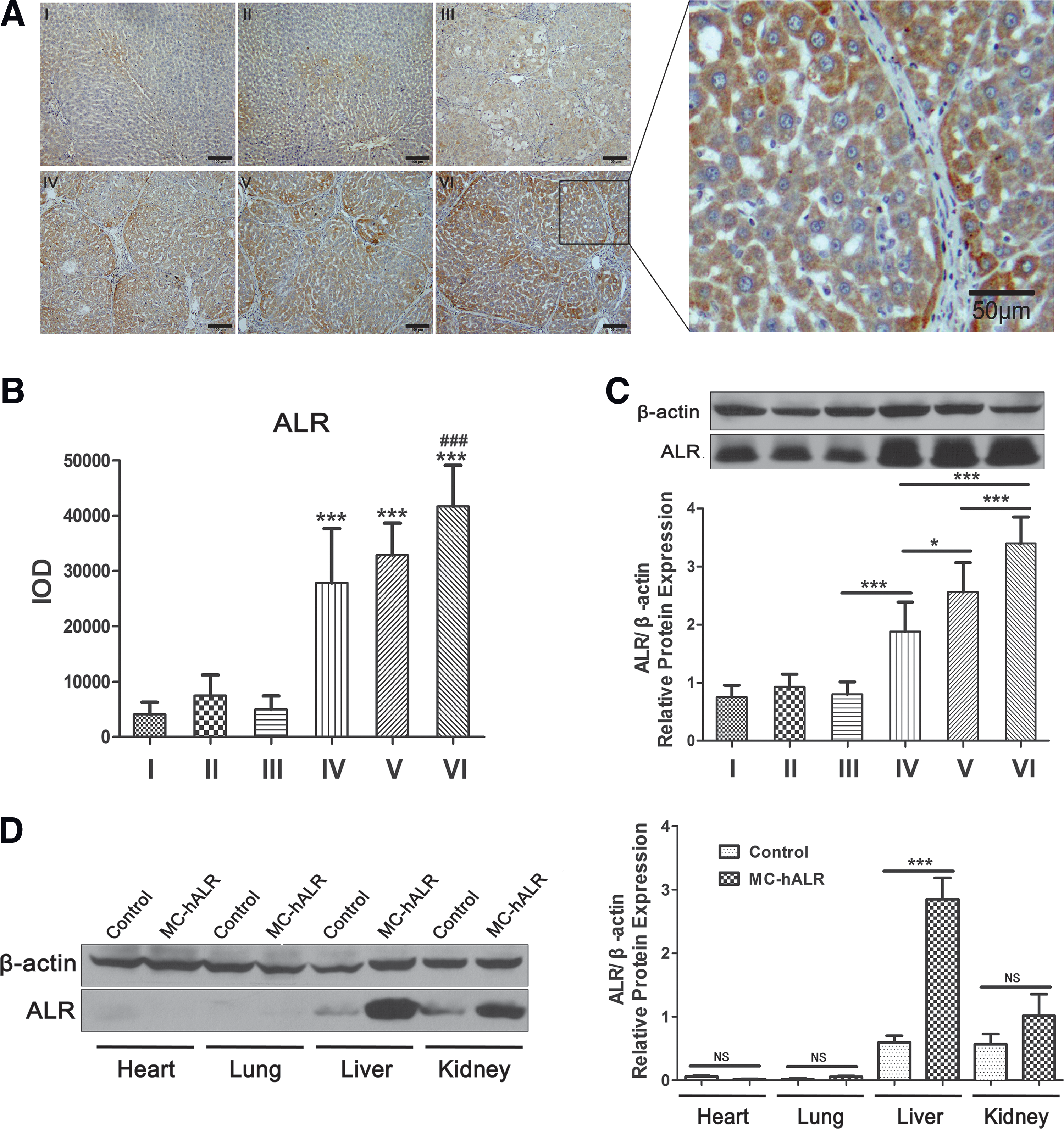
Analysis of the ALR expression in the fibrotic rats treated with MC-hALR or pcDNA3.1-hALR for 8 weeks.
Analysis of liver collagens and profibrogenic factors in fibrotic rats treated with MC-hALR for 8 weeks
To explore the therapeutic mechanism of ALR gene therapy, liver collagens and profibrogenic factors, such as TGF-β1, PDGF-BB, and α-SMA, were measured using immunohistochemistry or quantitative real-time PCR. Liver sections were stained with Sirius red to detect liver collagens and were compared with sections from the healthy control group (I). The liver sections from the fibrotic model group (III) exhibited prominent red staining in the fibrotic septa (Fig. 6A, left), indicating a high level of collagen deposition. The MC-hALR treatment groups (V, VI) demonstrated remarkably reduced staining by Sirius red. For quantification, the relative IOD was analyzed by IPP 6.0 software (Fig. 6A, right); a statistically significant difference was observed between group VI and group IV (p < 0.001).
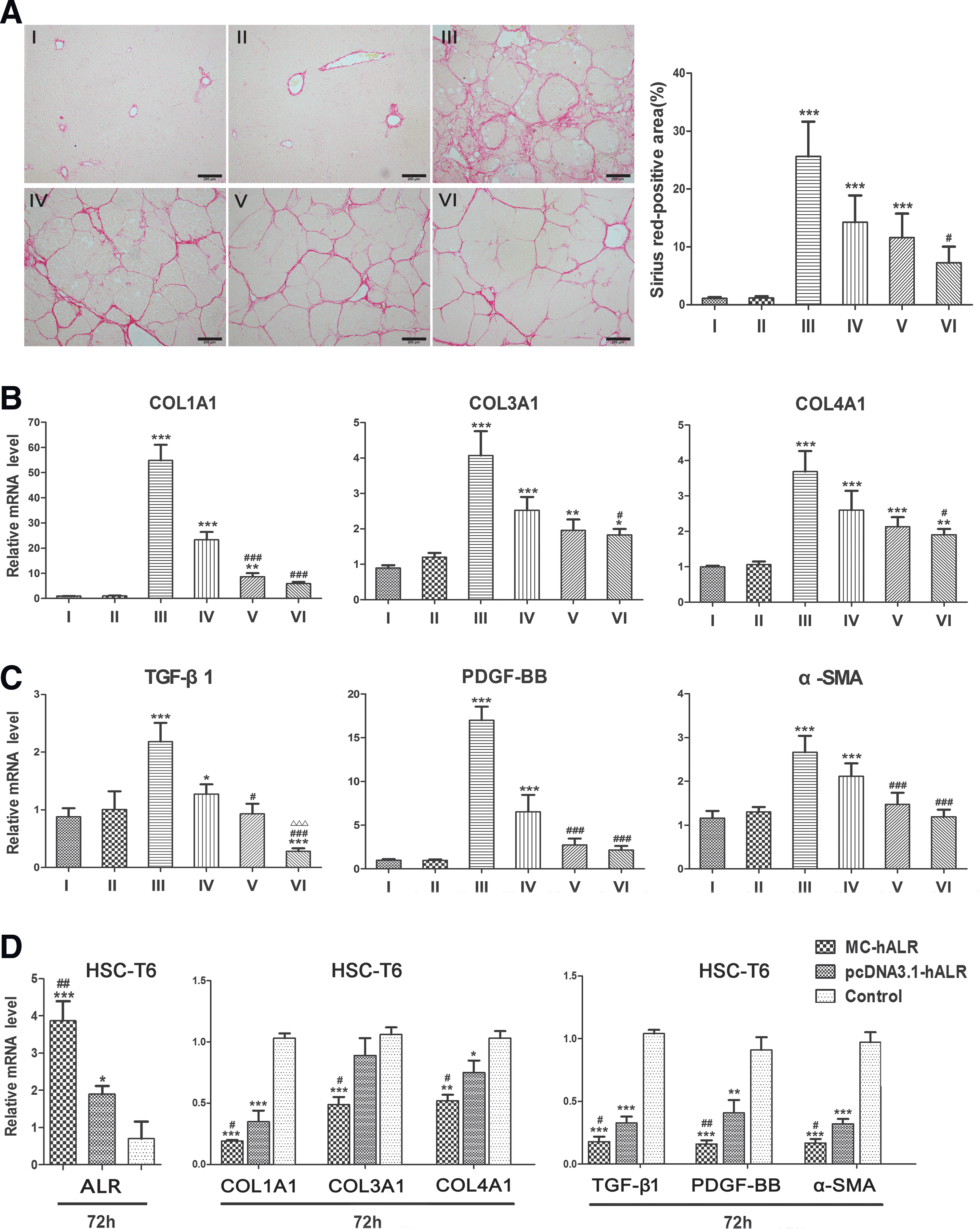
Analysis of the liver collagens and profibrogenic factors in the fibrotic rats treated with MC-hALR or pcDNA3.1-hALR for 8 weeks.
The mRNA expression levels of COL1A1, COL3A1, and COL4A1 were reduced in the MC-hALR treatment groups (V, VI) compared with the pcDNA3.1-hALR group (IV) (Fig. 6B). In particular, the level of COL1A1 was dramatically decreased in group VI (p < 0.001 vs. group IV), which suggested that relatively high-level ALR gene expression mediated by MC-hALR suppressed the expression of collagen genes, especially collagen I, to protect the liver from fibrogenesis.
As shown in Fig. 6C, compared with healthy rats (I), the fibrotic rats (III) showed significantly higher mRNA expression of TGF-β1, PDGF-BB, and α-SMA (p < 0.001), perhaps suggesting the activation of HSCs after CCl4-induced injury. Nevertheless, the levels of TGF-β1, PDGF-BB, and α-SMA were significantly reduced by the administration of MC-hALR (p < 0.001 vs. group I and II) and almost declined to baseline levels (NS vs. group I and II). Interestingly, the TGF-β1 level was dramatically reduced below the baseline level in group VI, which indicated that TGF-β1 gene expression was suppressed by the high-level expression of MC-hALR.
Subsequently, we tried to transfect MC-hALR into the cell line HSC-T6 to further determine whether HSC activation was efficiently blocked. With an increase in the ALR mRNA level in the MC-hALR-treated cells, the COL1A1, COL3A1, and COL4A1 levels were significantly attenuated (p < 0.001 vs. control), especially COL1A1, and the levels of TGF-β1, PDGF-BB, and α-SMA in the MC-hALR group were one-fifth of that in the control group (Fig. 6D).
Discussion
To our knowledge, this is the first report about the preparation of an ALR gene expression cassette in a novel minicircle DNA vector (MC-hALR). For high quality and yield of MC-hALR, we properly adjusted the size of inoculum, L-arabinose, and sodium hydroxide in previous preparation protocol; almost no parental plasmid or bacterial backbone remained in the culture; and the yield of MC-hALR was over 10 mg/liter, similar to that for pcDNA3.1-hALR, whereas the DNA size was reduced to one-third of the pcDNA3.1-hALR size. During the preparation of MC-hALR, L-arabinose was the most important agent. Upon L-arabinose induction, E. coli strain ZYCY10P3S2T engineered to express ΦC31 integrase and I-SceI endonuclease, and then the attB and attP sites (the bacterial and phage attachment sites of the ΦC31 integrase) of the parental plasmid were recombined to generate MC-hALR using ΦC31 integrase. In the end, the bacterial backbone plasmids were linearized by I-SceI and degraded by bacterial exonucleases, and only the MC-hALR ultimately remained in the culture.
This is also the first report of the application of MC-hALR in gene therapy in fibrotic rats. ALR is significantly expressed in the liver, kidneys, and testes. 21 In those tissue cells, ALR is distributed in several subcellular compartments, including the mitochondrial intermembrane space, nucleus, and endoplasmic reticulum, 22 which suggests the functional diversity of ALR. Many studies, including our previous research, revealed that exogenous ALR administration ameliorates hepatic damage, enhances hepatic proliferative capacity, and improves the survival of animals with partial hepatectomy, acute liver disease, or liver fibrosis. 2,3,12,23,24 However, minicircle-mediated ALR gene therapy for liver fibrosis has not yet been studied.
Before starting the gene therapy experiments, we compared the expression levels after MC-hALR or pcDNA3.1-hALR transfection in vitro and in vivo. In vitro, the relative ALR protein levels were ∼5-fold higher in SMMC-7721 and ∼4-fold higher in HSC-T6 cells treated with MC-hALR for 36 hr compared with those treated with pcDNA3.1-hALR (Fig. 2A). Subsequently, in liver fibrotic rats, even when we reduced the dose of MC-hALR to 30% (w/w) that of pcDNA3.1-hALR, the relative ALR protein level in the MC-hALR group was ∼2-fold higher and the protein remained for ∼7 days longer compared with that in the pcDNA3.1-hALR group (Fig. 3B). Taken together, the results clearly demonstrate that MC-hALR treatment resulted in relatively higher and more sustained ALR expression compared with that for pcDNA3.1-hALR. It is the novel minicircle technology 13,17,25 that removes large prokaryotic sequences to improve performance and to reduce the DNA size (Fig. 1A). As mentioned above, MC-hALR is composed only of the hALR gene expression cassette, without an origin of replication, drug selection markers, or expression-silencing sequences, especially CpG motifs and associated sequences that increase immune responses to the DNA and generate transgene silencing. 26 The elimination of the plasmid bacterial backbone, shortening of the spacer between the 5′ and 3′ ends of the transgene expression cassette, and a reduction in plasmid size could be the reasons for enhanced and persistent transgene expression mediated by MC-hALR. However, the expression of MC-hALR was relatively short-lived compared with other publications. 26,27 This is probably mainly because of the use of the CMV promoter that turned off soon after gene delivery in liver. 28,29 Thus, an optimal promoter should be chosen in the subsequent experiments.
In the 8-week gene therapy experiments using fibrotic rats, we were pleasantly surprised to find that MC-hALR exhibited greater potential for gene therapy for liver fibrosis, and minicircle DNA vector was able to induce the expression of the functional ALR protein to protect hepatocytes and ameliorate liver fibrosis. Even when we reduced treatment dose (30% [w/w] of the pcDNA3.1-hALR) and treatment frequency (half of the pcDNA3.1-hALR), MC-hALR exhibited better therapeutic effects against liver fibrosis because of its capability to mediate persistently high levels of ALR gene expression (Fig. 5A–C). In other words, MC-hALR is relatively safer and more efficient in gene therapy for liver fibrosis than pcDNA3.1-hALR because of persistently high levels of ALR gene expression, lower treatment dose, and lower treatment frequency. Additionally, even in the rats receiving 8 weekly hydrodynamic injections, necropsies revealed no apparent systemic anomalies, and liver histology confirmed the absence of malignant or premalignant lesions; most of the ALR proteins accumulated in the liver, with lower ALR protein expression found in the heart, kidneys, and lungs. These results demonstrate that HBT 30,31 is a more efficient method in ALR gene therapy for liver disease.
Analysis of the therapeutic effects in fibrotic rats treated with MC-hALR for 8 weeks (Figs. 4 –6) indicated that higher and more sustained ALR gene expression significantly blocked increases in the levels TGF-β1, PDGF-BB, and α-SMA, which play essential roles in the progression of liver fibrosis, and suppressed the production of collagens, especially collagen I (Fig. 6B). Liver fibrosis is a dynamic process characterized by the excessive accumulation of interstitial ECM distortion of the normal hepatic architecture, hemodynamic alterations, and hepatic dysfunction. 32 Liver injury elevates a variety of cytokines and growth factors, which can activate HSCs to produce ECM and cause liver fibrosis. 33,34 TGF-β1 is considered to be the key cytokine in accelerating cirrhosis progression, and it is critical for activation of HSCs in fibrotic liver. 35 PDGF-BB is another strong stimulator of HSCs proliferation and intracellular signaling, with concurrent predominant expression of PDGFR-β subunits in activated HSCs. 36 Once activated, HSCs will upregulate expression of ECM components, including collagen, TGF-β1, PDGF, and α-SMA, which all contribute to liver fibrosis, 37 and α-SMA is a marker of activated HSCs. 38 In our experiment, the fibrotic rats (Group III) showed significantly higher mRNA expression of TGF-β1, PDGF-BB, and α-SMA (p < 0.001) (Fig. 6C), suggesting the activation of HSCs after CCl4-induced injury. Whereas, as expected, TGF-β1, PDGF-BB, and α-SMA mRNAs were downregulated, and the production of collagens was partly inhibited in ALR treatment groups. We preliminarily concluded that the relative overexpression of MC-hALR inhibited the activation of hepatic stellate cells (HSCs), thereby alleviating liver fibrosis in rats. The subsequent experiments in HSC-T6 cells transfected with MC-hALR indicated that overexpression of ALR in HSC-T6 efficiently reduced the expression levels of TGF-β1, PDGF-BB, and α-SMA (Fig. 6D), and the collagen I, III, and IV levels were significantly attenuated, especially collagen I, which was in accordance with the in vivo results. These results may be correlated with a remarkable inhibition of HSCs proliferation.
Overall, MC-hALR gene delivery maximizes safety, efficacy, and sustained gene expression, and is a novel approach for the treatment of liver fibrosis. However, many challenges exist in terms of promoter optimization, safety evaluation, and underlying mechanisms research. In the subsequent experiments, we will try to target the ALR to liver cells and further improve the persistently high levels of ALR gene expression and determine the underlying mechanisms.
Conclusions
Compared with traditional plasmid pcDNA3.1-hALR, MC-hALR exhibits greater potential for gene therapy for liver fibrosis because of the persistently high levels of ALR gene expression, the lower treatment dose, and the lower treatment frequency. It preliminarily concluded that the relative overexpression of MC-hALR inhibits the activation of hepatic stellate cells (HSCs), thereby alleviating liver fibrosis in rats.
Footnotes
Acknowledgments
This work was supported by the Major Project of Chinese National Programs for Fundamental Research and Development (2010CB945502), Natural Science Foundation of Liaoning (2015020748), and Liaoning Education Department Scientific Research Project (L2015523).We thank Prof. Zhiying Chen, Prof. Chengyi He, and Dr. Tianyan Wang for their valuable suggestions on minicircle DNA preparation.
Author Disclosure
All authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.