
Abstract
Gene therapy has potential to treat rheumatic diseases; however, the presence of macrophages in the joint might hamper adeno-associated viral vector–mediated gene delivery. Here we demonstrate that in arthritic, but also in healthy, mice administration of agents that influence macrophage activity/number and/or addition of empty decoy capsids substantially improve the efficacy of recombinant adeno-associated viral vector 5 transgene expression in the joint. Pretreatment with triamcinolone or clodronate liposomes improved luciferase expression over a period of 4 weeks. Similar results were seen when empty decoy capsids were added to full genome containing capsids in a 5:1 ratio. In a study to assess the duration of expression as well as to investigate the combination of these two approaches, we observed a synergistic enhancement of gene expression, sustained for at least 12 weeks. The enhancement of gene expression was independent of the route of administration of triamcinolone (intra-articular or intramuscular). In healthy mice it was demonstrated that the combination improved expression of the transgene significantly, in a serotype independent manner. These data have implications for future applications of gene therapy to the joint and for other tissues with an abundance of macrophages.
Introduction
R
Several biological therapies are now available for RA, significantly improving the outcome of the disease in many, but not all, patients. 3 Over 50% of patients will still have active disease, and even the majority of clinical responders to biological treatment will have one or more actively inflamed joints. There remains an urgent need for the development of safe, efficacious, and cost-effective treatments to specifically target actively inflamed joints. Intra-articular gene therapy has the potential to treat arthritis, as the joint is an excellent target for the local delivery of gene therapy vectors.
There have been very few gene therapy clinical trials in RA, using either injection of cells that were genetically modified with a retrovirus expressing interleukin-1 receptor antagonist into the joint (ex vivo gene therapy), or injection of a recombinant adeno-associated viral vector (rAAV) encoding a tumor necrosis factor (TNF) antagonist (in vivo gene therapy). 4,5 While intra-articular administration of a recombinant adeno-associated virus 2 (rAAV2) vector encoding the TNF receptor immunoglobin G1 fusion gene (tgAAC94) 6,7 was well tolerated, the treatment was not effective. It is likely that lack of efficacy was due to insufficient levels of gene expression. This notion is supported by the fact that local gene expression, as determined by a radioimmunoassay for tumor necrosis factor receptor Fc (TNFR:Fc) protein in synovial fluid and an RT-PCR assay for mRNA specific to rAAV2–TNFR:Fc in synovial tissue, was not detected in the very limited number of samples that became available during the study. 7 Moreover, we have shown that intra-articular injection of etanercept into the inflamed joint of RA patients leads to clinical improvement, validating the therapeutic target when administered to the site of inflammation. 8 Importantly, this trial provided encouraging data on safety and feasibility for further development of AAV-based intra-articular gene therapy in RA.
We are developing intra-articular gene therapy for RA using rAAV5. 9 –11 Comparing AAV serotype 1 through 5 we have previously shown that AAV5 was the most efficient in transducing synovial tissue after intra-articular injection in two different animal models of RA. 12,13 The main target cells for rAAV5 in the joint are FLS. AAV5 is effective in in vitro transduction of both human and rodent FLS 14 as well as in in vivo transduction in rat and mouse models of RA. 12,13 AAV5 has the added benefit that the majority of the human population is seronegative for AAV5. 15
In an effort to improve transduction efficiency, we have investigated factors that can influence in vivo transduction of rAAV vectors after intra-articular injection in animal models of RA and in healthy animals. Here we present data that uncover an unexpected barrier to efficient gene transfer to the joint, namely, that synovial macrophages significantly inhibit AAV-mediated gene delivery, and we present strategies for overcoming this barrier. These data have implications for future applications of local gene therapy to the joint, or to other tissues that are abundantly infiltrated by macrophages.
Materials and Methods
Vector and empty capsid production
An rAAV5 and rAAV2 vector was produced coding for firefly luciferase (Fluc) with a cytomegalovirus (CMV) promoter (rAAV5.CMV.Fluc and rAAV2.CMV.Fluc; Children's Hospital of Philadelphia, Philadelphia, PA) as described previously. 16 In brief, the plasmid encodes the Fluc gene under the control of the CMV promoter and a human growth hormone polyadenylation signal. The transgene cassette is flanked by AAV2 inverted terminal repeats and is packaged in capsid from AAV5 or AAV2. 17 Empty capsids were generated in a protocol identical to the full capsids; however, the AAV genome plasmid was left out of the initial transfection. The genome containing vector and empty AAV capsid particles were purified by combined polyethylene glycol precipitation and empty capsids were separated from full capsids by cesium chloride density gradient centrifugation. 18 Vector titers (empty and full) were determined by silver stained sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) using internal AAV reference standards, and vector genome titers were verified by quantitative PCR and expressed as viral genomes per milliliter (vg/mL).
Vector administration in mice
Intra-articular rAAV expression was investigated in male Dilute Brown Non-Agouti (DBA) mice (8–12 weeks old; Harlan Sprague Dawley, Horst, the Netherlands). Groups consisted of 5 to 18 animals per experiment (indicated per experiment in the legends of the figures). Mice were injected with rAAV5.CMV.Fluc or rAAV2.CMC.FLuc in one or both knee joints and monitored periodically for luciferase expression (as indicated in the legends of the figures, from 3 days, weekly up till 4 weeks and monthly up till a maximum of 6 months). For animals without arthritis the vector was administered on day 1; in arthritic animals the vector was injected on day 17 or 24 after immunization (before or after the onset of arthritis). Animals received 1.26 × 1010 to 1.65 × 1010 vg of rAAV per knee joint (in a volume of 5 μL) in healthy mice or mice with collagen-induced arthritis (CIA) as indicated in the legends of the figures. Empty capsids were co-administered with the genome containing particles in several groups in a 5:1 or 20:1 empty-to-full ratio, as indicated.
Collagen-induced arthritis
Arthritis was induced and evaluated as previously described. 19–20 Bovine type 2 collagen (2 mg/mL in 0.05 M acetic acid; Chondrex, Redmond, WA) was mixed in an equal volume of Freund's complete adjuvant (2 mg/mL of Mycobacterium tuberculosis; Chondrex). The mice were immunized intradermally at the base of the tail with 100 μL of emulsion (100 μg collagen) on day 0. On day 20, mice received an intraperitoneal booster injection of 100 μg type 2 collagen in NaCl. The severity of arthritis was assessed using an established semiquantitative scoring system (0–4; 0 = normal, 1 = swelling and/or erythema of 1 joint, 2 = swelling in >1 joint, 3 = swelling of all joints, and 4 = swelling of the entire paw and at least one of the following symptoms: ankylosis, loss of grip). The cumulative score for all four paws of each mouse was used to represent overall disease severity and progression. In the long-term follow-up study presented in Fig. 1, due to technical problems of the in vivo imaging system, data are not available for the time point at 8 weeks. In addition, a total of 15 animals (2–4 per group) were sacrificed prior to the end of this experiment due to reaching a humane endpoint (clinical score >12).
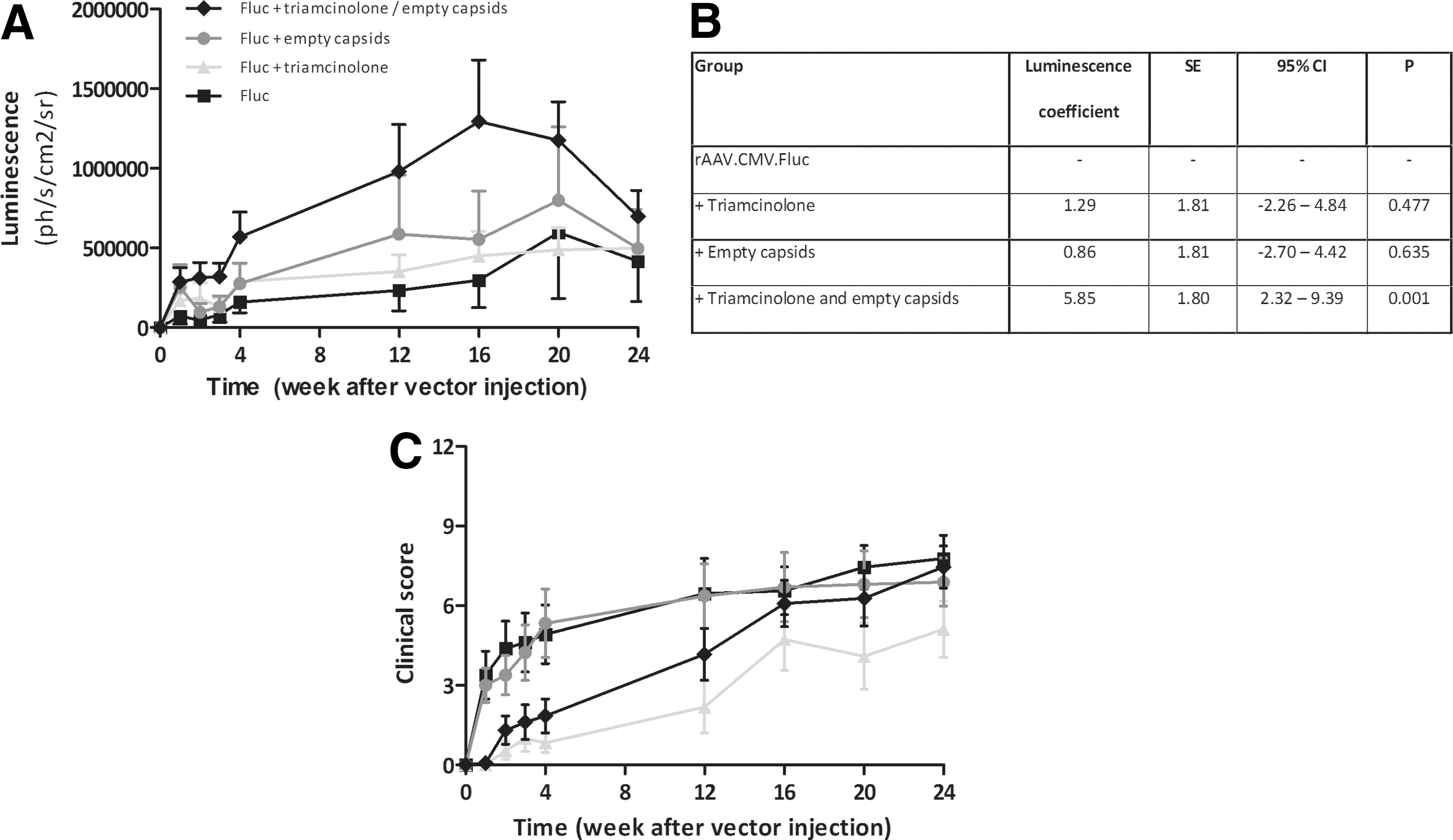
Improvement of intra-articular recombinant adeno-associated viral vector 5 (rAAV5)–luciferase expression by addition of empty capsids and/or triamcinolone. Arthritic mice (n = 13 mice per group, 26 injected joints per group) were injected intra-articularly (i.a.) with 1.65 × 1010 viral genomes (vg) per joint of rAAV5 firefly luciferase (Fluc) with cytomegalovirus (rAAV5.CMV.Fluc) vector with or without the addition of empty capsid (5:1 empty to full) or triamcinolone (5 mg/kg intramuscularly [i.m.]). The vector was administered on day 24 after onset of arthritis, and mice were followed weekly for 1 month and thereafter monthly up till 6 months.
Macrophage inhibition/depletion
Macrophage inhibition was initiated by two different compounds, clodronate containing liposomes (ordered from
Imaging of luciferase expression
Luciferase expression was measured at different time points after vector administration, from day 3 up till 6 months depending on the experiments. Luciferase expression was detected as described previously. 14 In brief, D-luciferin potassium-salt substrate (Caliper Life Sciences, Hopkinton, MA) was injected intraperitoneally (150 mg/kg of body weight, in a volume of approximately 200 μL). Photon counts were acquired 10 min after substrate administration for 5 min using a cooled charge-coupled device camera system (Photon Imager, Biospace Lab, Paris, France) and image processing, signal intensity quantification and analysis were performed using M3 Vision (Biospace Lab). The number of photons emitted per second per square centimeter per steradian was calculated as a measure of luciferase activity.
Determination of genome copy per injected joint
Knee joints were harvested at the end of study (week 12) and flash frozen using liquid nitrogen. Frozen knees were cut into fragments and pulverized using a mortar and pestle using dry ice and liquid nitrogen. Genomic DNA was isolated using a DNeasy Blood and Tissue DNA isolation kit (Qiagen) and used for quantitative PCR using a polyA signal specific primer:probe set. A standard curve using a polyA signal containing AAV5 was generated and was used to determine the vector copy numbers in the tissue. Vector copies were normalized to the total amount of genome DNA isolated and corrected for differences in knee weight.
General animal conditions and ethics statement
Immunization, intra-articular injections and in vivo imaging were performed under isoflurane anesthesia (3% isoflurane and oxygen). At the end of the experiments, animals were sacrificed by cardiac puncture under isoflurane anesthesia, followed by cervical dislocation. The studies were reviewed and approved by the animal care and use committee of the University of Amsterdam (Amsterdam, the Netherlands; permit numbers ART 102881, ART 102656, ART 103021, and ART 111AB) and carried out in strict accordance with the recommendations in the Dutch Law on Animal Welfare (Dutch: “Wet op Dierproeven”). Animals were maintained under pathogen-free conditions in the animal facility of the University of Amsterdam.
Statistical analysis
For the experiments presented in Figs. 2 –6 statistics were analyzed using where appropriate the t-test or Mann Whitney test in Graphpad Prism. For all tests, differences with a p-value of <0.05 were considered significant. In the experiments presented in Figs. 2 –4 and Fig. 6, both hind paws were injected and they were included as separate measurements in the analysis.
In the experiment presented in Fig. 1, luminescence over time was investigated using a multilevel longitudinal analysis with generalized estimating equations (GEEs). This is a suitable technique for longitudinal analysis, making use of all available data and adjusting for the “within subject” correlation. GEEs also allows one to use an unequal number of repeated measurements, which was the case here. Every observed knee was nested within mice, making it a multilevel analysis (with two levels) and appropriately adjusting for the fact that some observations came from the same mouse (Fig. 1). 22 In this model, we investigated the effect of different interventions (empty capsids and/or triamcinolone) on luminescence.
Results
Inflammation affects intra-articular rAAV5 gene expression
A hallmark of synovial inflammation is the proliferation of FLS in the synovial tissue of RA patients. 2 In a healthy joint the intimal lining layer is 1–2 cell layers thick, whereas in the inflamed joint this is increased to 8–10 cell layers. This is also true in mouse models of RA, including in the CIA model. As FLS are the primary target cells for rAAV5 in the joint, we hypothesized that administration of rAAV5 after the onset of inflammation in the CIA model would lead to higher expression, due to a higher number FLS present in the joint at the time of inflammation. To test this hypothesis, we administered an rAAV5 vector encoding the firefly luciferase gene (rAAV5.CMV.Fluc) intra-articularly in mice with CIA, before (day 17) or after (day 24) the onset of arthritis. Surprisingly, we found that administration of an rAAV5 vector after the onset of inflammation (day 24) resulted in lower expression compared to vector administration before the onset of inflammation (day 17) (Fig. 2).
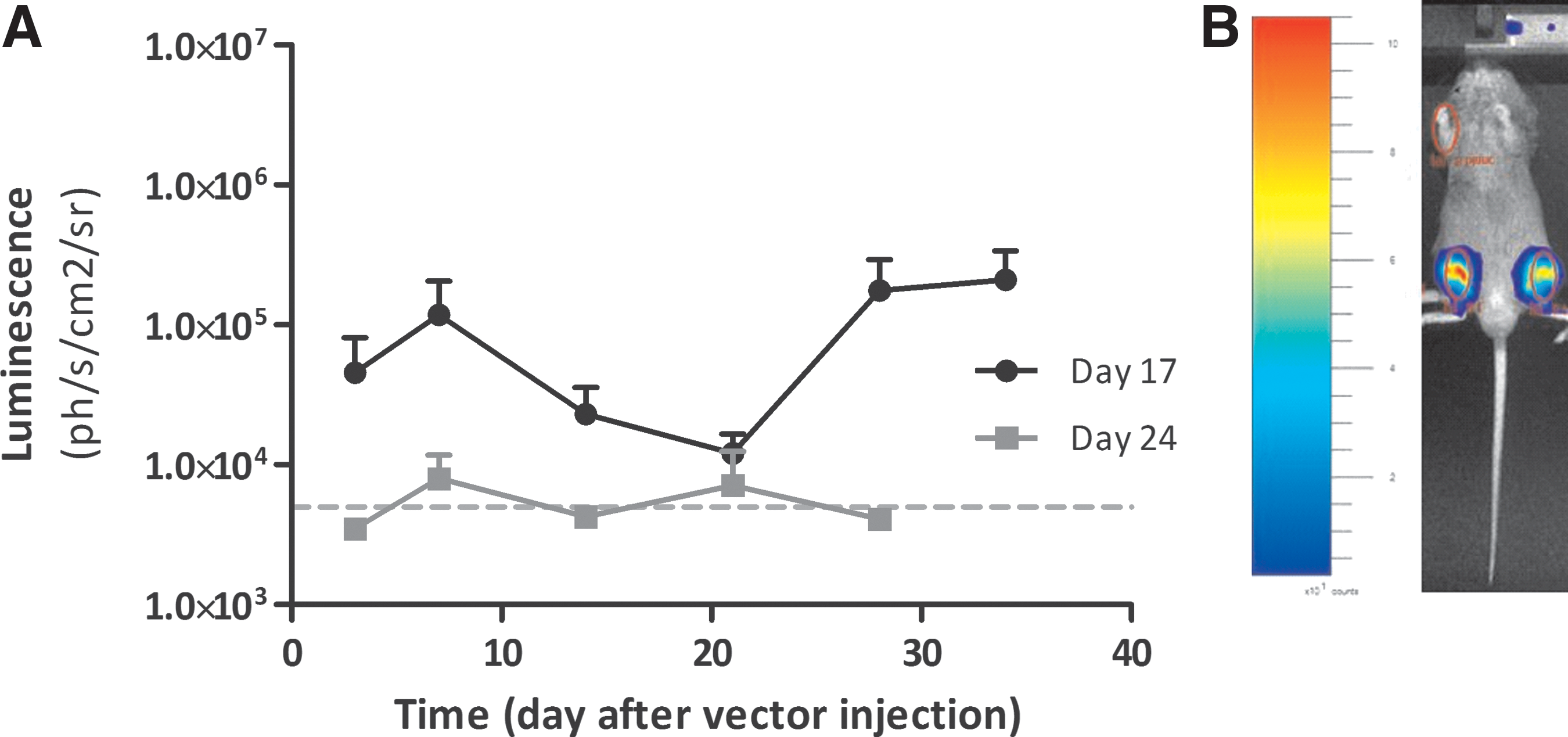
Luciferase expression is influenced by vector administration before or after the onset of arthritis.
Immunosuppressive agents improve rAAV5 transgene expression
An explanation for decreased expression in animals with inflamed joints could be degradation or neutralization of the vector before it is able to transduce the target cells. During inflammation, there is not only an increase in the number of FLS, but there is also an increase in the number and activation of macrophages. We hypothesized that the decreased expression could be due to vector neutralization by macrophages (for example through phagocytosis or opsonization by soluble factors (e.g. complement)). To investigate this possibility we examined whether administration of agents that influence macrophage activity and/or number had an effect on rAAV5 expression.
Triamcinolone, a corticosteroid, acts by inhibiting the activation and proliferation of immune cells, including macrophages. 23 Alternatively, clodronate containing liposomes—a well-established method for depleting macrophages (including synovial macrophages)—were used to deplete macrophages. 21 The two agents were administered 48 hours before vector administration. Both triamcinolone and clodronate liposomes improved rAA5.CMV.Fluc luciferase expression over a period of 4 weeks, showing that either depletion or inhibition of macrophages leads to an increase in gene expression (Fig. 3A).
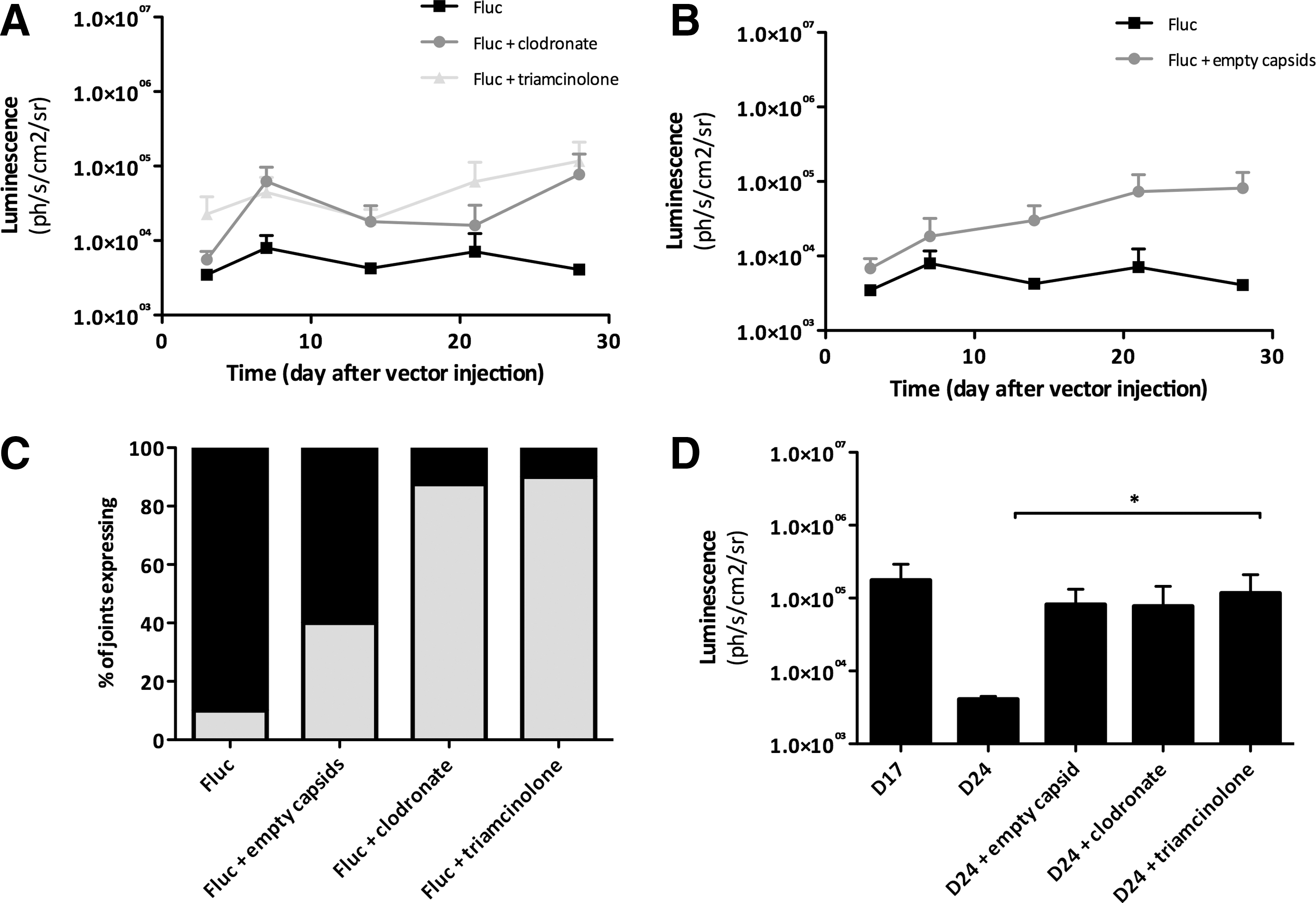
Effect of addition of clodronate, triamcinolone and empty capsids on rAAV5 luciferase expression. After induction of arthritis, mice (n = 5 mice per group, 10 injected joints per group) were i.a. injected with 1.65 × 1010 vg/joint of rAAV5.CMV.Fluc vector. Imaging was performed 3 days after vector injection and thereafter weekly up to 4 weeks.
We hypothesized that an alternative approach to avoid macrophage vector neutralization would be to add empty capsid particles upon vector administration. These empty capsids could act as a decoy and therefore increasing the chances that full virus particles will be able to reach the target cells. This strategy is similar to the one previously used to avoid neutralizing anti-AAV capsid antibodies. 24 When empty capsids were added to full genome containing capsids in a 5:1 ratio, expression improved significantly (Fig. 3B). These data supported our hypothesis that the vector is likely being neutralized by macrophages. In support of this hypothesis, we observed that all three treatment groups (triamcinolone, clodronate liposomes, empty capsid) also showed an increased percentage of positive joints (Fig. 3C). In Fig. 3D the luminescence 28 days after vector administration is shown, demonstrating a significant difference between administration of the vector alone after onset of disease compared to the groups that received treatment. As triamcinolone is an anti-inflammatory agent, arthritis activity was closely monitored. As expected, mice treated with triamcinolone or clodronate liposomes showed a delayed onset of arthritis (results not shown).
Triamcinolone and decoy capsids have a synergistic effect on rAAV5 expression
As we had found that pharmacological inhibition of macrophage activity or addition of empty decoy capsids resulted in increased gene expression, we performed a long-term follow-up study to assess the duration of improved gene expression, as well as to investigate the combination of these two approaches. As triamcinolone is a therapeutic compound that is already approved for the treatment of RA, we selected this compound for the follow-up studies. The effect of addition of triamcinolone and/or empty capsids was analysed in a multilevel longitudinal analysis using GEEs, allowing us to include all available longitudinal data, allowing unequal numbers of repeated measurements and taking into account the within-subject correlation (over time and between the two knees of the same mouse). Similar to the previous study, we observed increased expression using triamcinolone or empty capsid alone. As can be seen from Fig. 1, it was clear that the combination of macrophage inhibition and decoy capsids resulted in a synergistic enhancement of gene expression (Fig. 1A, B; 5.85-fold enhancement, p < 0.001) and that this enhancement was sustained for up to 16 weeks post vector administration. After 16 weeks, the effect was diminishing but increased expression of the transgene was observed for up to 6 months. As expected, due to its anti-inflammatory effect, arthritis activity was lower in groups treated with triamcinolone until week 4 (Fig. 1C); however, long-term arthritis activity was comparable between groups.
Local triamcinolone administration is as effective as systemic administration on increased gene expression of rAAV5
In the clinic, triamcinolone is usually administered locally by intra-articular injection into the inflamed joint; therefore, we compared the efficacy of local versus systemic administration of triamcinolone on enhancement of gene expression. To test this, triamcinolone (or saline) was administered systemically (intramuscular) or locally (intra-articular) 2 days prior to intra-articular administration of rAAV5.CMV.Fluc + empty capsid (empty to full ratio of 5:1). Luciferase expression was monitored over time by the in vivo imaging system imaging up till 112 days. We observed similar increases in gene expression when triamcinolone was administered locally or systemically, indicating that local administration of triamcinolone was as effective as systemic administration (Fig. 4).

Improvement of intra-articular rAAV5-luciferase expression by addition of empty capsids and/or triamcinolone. Mice with CIA (n = 18 per group) were injected with triamcinolone either systemically (i.m.) or locally (i.a.) two days prior to i.a. administration of rAAV5.CMV.Fluc (1.65e10 vg/joint) and empty capsid (5:1 empty to full). Luciferase expression was monitored over time by IVIS imaging for a follow-up of 30 days after vector administration. Luminescence is shown per group as average; error bars, SEM.
AAV empty capsids improve transgene expression of rAAV5 in the absence of inflammation
All previous experiments were performed in CIA models, in which animals experienced significant inflammation in the joint at the time of vector administration. As even healthy synovium contains a significant percentage of macrophages, we next decided to investigate whether empty decoy capsids could enhance gene expression in a healthy joint. When empty capsids were added to genome containing particles in two different ratios (5:1 and 20:1) we observed a dose dependent increase in gene expression (Fig. 5). The increase was 4.8-fold on average in animals injected with empty capsids in a 5:1 ratio to full capsids (p < 0.01), and 20-fold when a 20:1 empty to full ratio was used (p < 0.05).
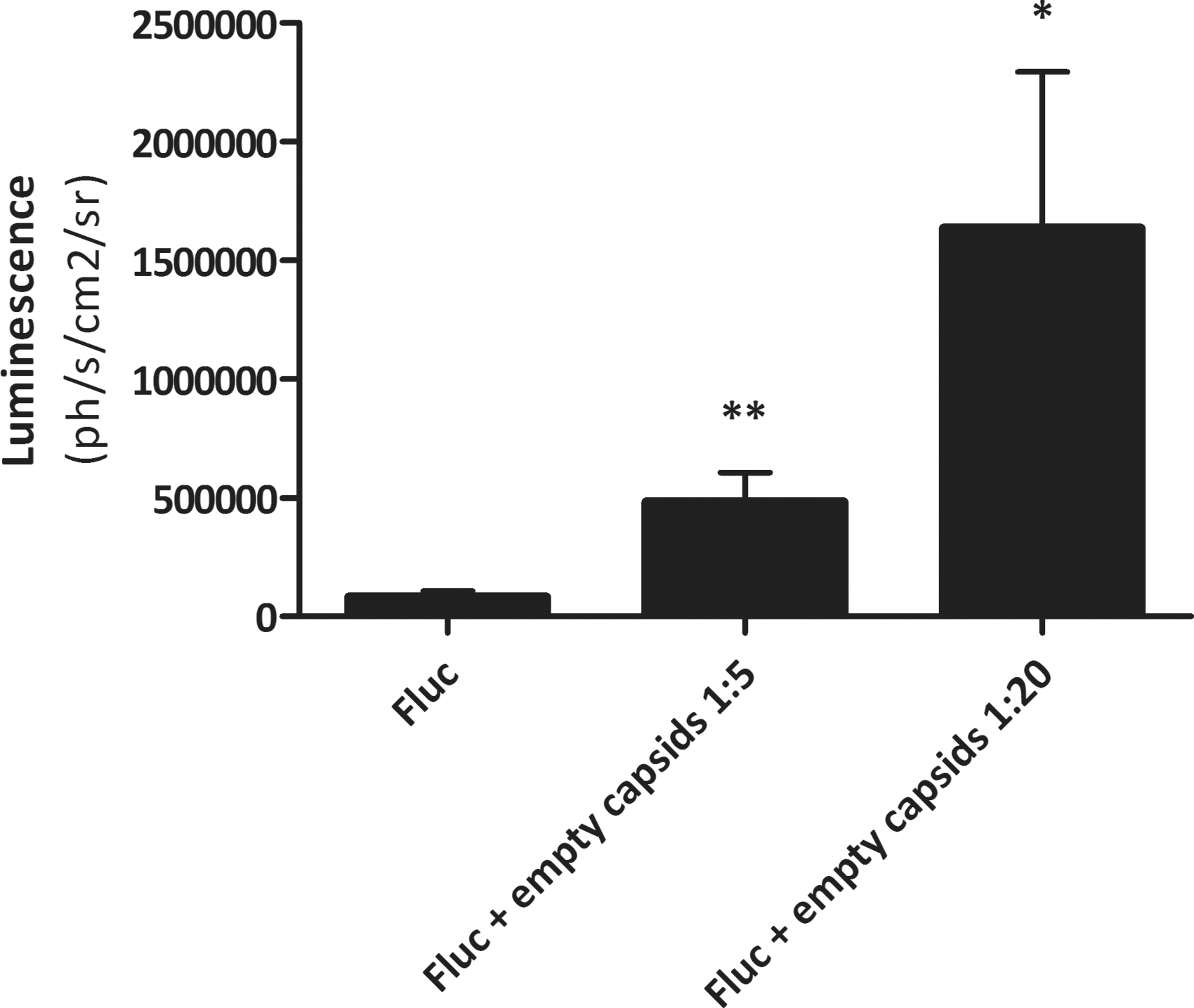
Addition of empty capsids in 2 different ratios (5:1 and 20:1 empty to full) improves intra-articular rAAV5-luciferase expression in healthy mice. Healthy mice (n = 7 mice per group, one joint injected per animal) were injected i.a. with 1.65 × 1010 vg/joint rAAV5-CMV-FLUC. In two groups, empty capsids were added in different ratios. Luciferase expression was measured weekly until mice were sacrificed after 4 weeks. Luminescence at week 4 is shown per group as averages; error bars, SEM. *p < 0.05 and **p < 0.01 in groups with empty capsid addition versus the control group that only received genome containing vector (one-tailed, unpaired t-test).
Next, we investigated the effect of the combination of the two approaches on the expression of the transgene in healthy joints. As can be seen in Fig. 6A and B, both empty decoy capsid and triamcinolone treated animals showed increased gene expression compared with vector alone animals, but the combination of empty capsid and triamcinolone gave rise to the highest increase in gene expression levels, similar to what was observed in inflamed (CIA) joints. To further investigate the possible mechanism of increased expression, genomic DNA was extracted from knees from triamcinolone + empty capsid and control mice and was used to determine the vector genome copy number. It was found that the treated group had significantly higher numbers of vector genomes per joint than the control treated animals (Fig. 6C). These results support the hypothesis that macrophages in the intimal lining layer inhibit AAV-mediated expression, even in healthy joints, and that either adding decoy capsid particles and/or inhibiting macrophage activity can overcome this inhibition, leading to increased gene expression and number of vector genomes per joint.
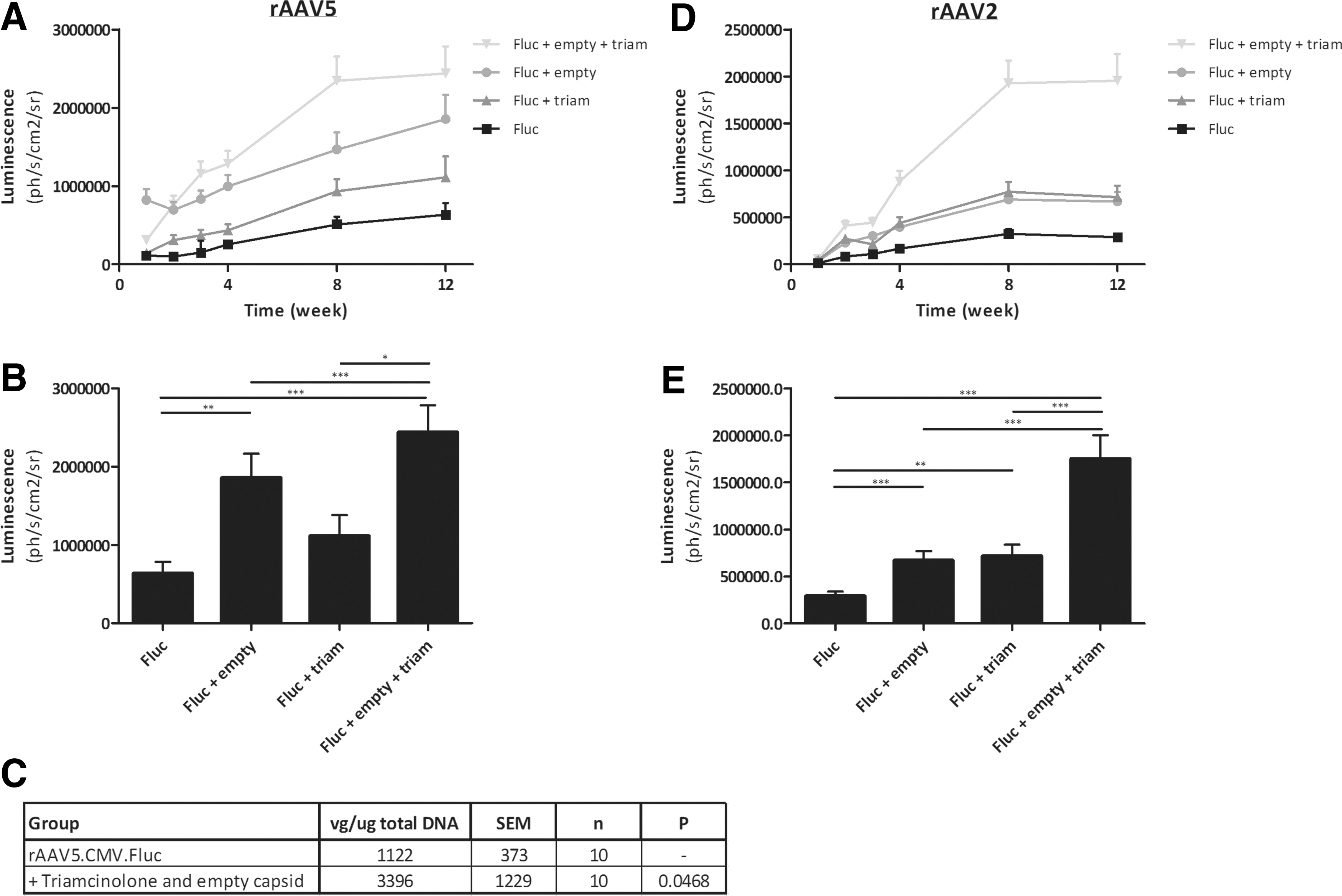
Effect of empty capsid and triamcinolone on intra-articular rAAV5 and rAAV2 gene expression in healthy mice. Healthy mice (n = 17 mice per group, 34 total injected joints) were injected with 1.26 × 1010 vg/joint rAAV5-CMV-Fluc or rAAV2-CMV-Fluc intra-articularly +/− empty capsid (5:1 empty to full ratio) preceded 2 days prior with i.m. administration of either saline (NaCl) or triamcinolone. Luciferase expression was measured weekly by IVIS up to week 12.
Combination of empty capsid and triamcinolone is not specific for rAAV5 but enhances intra-articular gene expression using other serotypes as well
Our studies thus far focused on AAV5, as this serotype is very effective in transducing FLS; however, we hypothesized that the effect of macrophage neutralization on gene transfer using AAV is not serotype specific. AAV uptake by macrophages is a general phenomenon utilizing scavenger receptors, 25 and thus this should not be limited to one specific serotype, or any specific virus as macrophages are known to take up a wide range of viruses and bacteria. To test this hypothesis, we performed an experiment in which we evaluated whether triamcinolone and empty capsid could enhance gene expression from a serotype that is very different from AAV5, this being AAV2. AAV5 and AAV2 share only 57% homology at the amino acid level, making them two of the most diverse serotypes of AAV known. To test this, we performed an experiment identical to that described above (Fig. 6A, B), but now with rAAV2. Similar to results seen with rAAV5, both empty decoy capsid and triamcinolone treated animals showed increased gene expression compared with vector alone animals following rAAV2 administration (Fig. 6D, E). The combination of empty capsid and triamcinolone gave rise to the highest increase in gene expression levels, similar to what was observed in rAAV5 treated animals, providing further validation of this approach.
Discussion
While there have been many advances in the treatment of RA, there remains an urgent need for safe and effective therapies targeting inflamed joints that are unresponsive to systemic therapies. To address this need, we and others have proposed local delivery of rAAV as a potential platform for the long-term expression of therapeutic genes. 14,26 –28 Recombinant AAV vectors have been extensively used for in vivo gene therapy and have been shown to be safe and effective in prelinical models as well as in clinical trials. 4,29
In the course of developing an rAAV-based gene therapy product for the treatment of RA, we have used rAAV5 expressing firefly luciferase combined with in vivo imaging of mice to investigate factors affecting intra-articular gene expression. Of importance, we found that, following intra-articular administration of rAAV5 in arthritic animals, not all joints are effectively transduced (usually <50%). To ensure sustained local production of effective doses of therapeutic proteins in the joint, in particular in the rheumatoid synovium, an optimized gene delivery system is clearly needed.
Here we present data showing that synovial macrophages represent a previously unrecognized barrier for efficient intra-articular gene transfer. Our data suggests that agents that can deplete and/or inhibit activation of macrophages (clodronate liposomes or triamcinolone), as well as the addition of empty decoy capsids to the vector preparation, can improve luciferase expression after local injection in the joint. Using a combination of both approaches resulted in a synergetic effect. Importantly, this effect was not limited to inflamed joints and was AAV serotype independent, having clear implications for all intra-articular gene transfer applications.
Triamcinolone, a corticosteroid that is frequently used in the treatment of RA, had a positive effect on transgene expression in a similar order of magnitude as seen for clodronate liposomes. Systemic administration of corticosteroids is known to exert a local effect in the joint by decreasing the number of macrophages in synovial tissue of RA patients, 30 indicating that the effect of triamcinolone is at least partly related to the depletion of macrophages. 31 Both systemic and intra-articular administration of corticosteroids can be accompanied by side effects. 32 In cases of mono- or oligoarthritis, the preferred route is intra-articular. We demonstrated that intra-articular injection of triamcinolone was equally effective in increasing gene expression compared with systemic administration, which is an important finding in view of the possible application of this approach for intra-articular gene therapy in RA.
The findings reported in this manuscript may also be relevant for the lack of efficacy in the phase 1–2 clinical trial with tgAAC94. 7 In this study, adults with persistent moderate or severe inflammation in a clinically inflamed joint received a single intra-articular injection of rAAV2 expressing the human TNFR:Fc gene. Arthritis activity is strongly correlated with macrophage infiltration of the synovial tissue. 33 Based on the findings presented here, we postulate that the presence of macrophages at the site of injection may have diminished the transduction efficacy of the vector, thereby preventing efficient transgene expression to induce a therapeutic effect. Other factors, including neutralizing antibodies, may also have played a role.
One interesting observation was that the enhanced gene expression using triamcinolone and empty capsid is pronounced up to at least 84 days (12 weeks) after vector injection, but thereafter the difference between experimental and control animals decreases. This appears to be due to a slow and gradual increase in gene expression from the control groups. The reason for this delayed onset of gene expression following intra-articular gene expression is currently not clear; however, it should be noted that reducing the lag time between gene delivery and therapeutic levels of gene expression would be advantageous to affected patients.
Our study shows that the combination of triamcinolone and empty capsid is effective for increasing expression not only in diseased joints, but also in healthy joints. These results may be relevant for a wider range of applications using rAAV vectors delivered to tissue containing macrophages. This notion is supported by a previous study showing that blocking rAAV8 uptake by macrophages via scavenger receptors (using polynosinic acid [poly(i]] can reduce AAV endocytosis in vitro, and that poly(i) enhances liver targeted gene expression in vivo in a rat model of hyperbilirubinemia. 25 As expected, we found that enhancement of gene expression by macrophage inhibition is applicable to AAV serotypes other than AAV5 as well. On a related note, these studies used the CMV promoter to drive expression of luciferase, and as the regulation of expression from this promoter is complicated due to multiple transcription factor binding sites, it is possible that some of the enhanced expression that was observed was due to changes in promoter expression. Given that enhanced gene expression was observed in both inflamed (CIA) and healthy mice, and that previous studies have shown that inflammation can actually increase CMV expression in fibroblast-like synoviocytes, 34 it is likely that the enhanced gene expression seen here is not specific to the CMV promoter; however, future studies using different promoters should be performed.
Recently, it was reported that addition of a significant quantity of empty capsids to the AAV transgene composition after intravenous administration is able to overcome the inhibitory effect of (pre-existing) neutralizing antibodies following systemic delivery to the liver. 24 It was shown that addition of empty decoy capsid to the final vector formulation can adsorb these antibodies and thus overcoming their inhibitory effect. Based on these findings, we hypothesized that empty capsids could act in a similar manner as decoy for macrophages, thereby preventing uptake and degradation of the transgene-containing rAAV vector. Indeed, we found that addition of empty decoy vectors to the full vector preparation in a ratio of 5:1 or 20:1 (empty to full) has a positive effect on transgene expression in the joint. The exact mechanism of action of these decoys is currently not known, and it is unclear if empty capsids have a higher affinity for macrophages compared to full capsids, or if the abundance of total capsid particles is overwhelming the macrophage scavenger pathway, resulting in more rAAV particles able to transduce the synovial FLS.
Empty capsids can be part of the vector preparation or be produced independent of the rAAV vector batch, and if desired, added to AAV vector preparations, or administered separately to an individual. In future studies, the ratio of empty to full particles needs to be investigated in more detail, also in the context of the level of inflammation and number of macrophages present. Given that capsid specific immune responses have been reported in clinical trials using AAV, 35–36 increasing the total antigen load by using empty capsids may not be desirable. An interesting approach to further enhance the safety of the use of empty capsids to bind neutralizing antibodies is the mutation of the receptor binding site of AAV2 to generate an empty capsid mutant that can adsorb antibodies but cannot enter a target cell, 18 thereby also reducing the potential for targeted immunotoxicity. Whether such an approach is also feasible in reducing the detrimental effects of macrophages on transgene expression needs to be evaluated in future studies.
In conclusion, we provide evidence that intra-articular macrophages represent a barrier to efficient gene transfer, and that triamcinolone or empty decoy capsids alone result in improved gene expression in inflamed and healthy joints. Importantly, the combination of a corticosteroid and empty capsids led to a synergistic increase in gene expression. These data have implications for future applications of gene therapy to the joint and also to other tissues with an abundance of macrophages.
Footnotes
Acknowledgments
The authors want to thank L. Bevaart, S. Snoek, and F.A. Koopman for their contributions in writing the application for the animal ethical committee and their assistance in getting the studies initiated.
Author Disclosure
This research was supported by Arthrogen. Co-authors Caroline J. Aalbers, Margriet J. Vervoordeldonk, and Jonathan D. Finn were at the time of the experiments employed by Arthrogen BV. Niels Broekstra and Emiel Kramer are currently employed by Arthrogen BV and receive a salary. Paul P. Tak is inventor on the patent “AAV vectors for in vivo gene therapy of rheumatoid arthritis” (EP 2 322 638 A1) but does not receive any financial compensation for this patent from Arthrogen or any other institution.