
Abstract
In a model of growth-restricted sheep pregnancy, it was previously demonstrated that transient uterine artery VEGF overexpression can improve fetal growth. This approach was tested in guinea-pig pregnancies, where placental physiology is more similar to humans. Fetal growth restriction (FGR) was attained through peri-conceptual nutrient restriction in virgin guinea pigs. Ad.VEGF-A165 or Ad.LacZ (1 × 1010vp) was applied at mid-gestation via laparotomy, delivered externally to the uterine circulation with thermosensitive gel. At short-term (3–8 days post surgery) or at term gestation, pups were weighed, and tissues were sampled for vector spread analysis, VEGF expression, and its downstream effects. Fetal weight at term was increased (88.01 ± 13.36 g; n = 26) in Ad.VEGF-A165-treated animals compared with Ad.LacZ-treated animals (85.52 ± 13.00 g; n = 19; p = 0.028). The brain, liver, and lung weight and crown rump length were significantly larger in short-term analyses, as well as VEGF expression in transduced tissues. At term, molecular analyses confirmed the presence of VEGF transgene in target tissues but not in fetal samples. Tissue histology analysis and blood biochemistry/hematological examination were comparable with controls. Uterine artery relaxation in Ad.VEGF-A165-treated dams was higher compared with Ad.LacZ-treated dams. Maternal uterine artery Ad.VEGF-A165 increases fetal growth velocity and term fetal weight in growth-restricted guinea-pig pregnancy.
Introduction
F
More than half of FGR occurs secondary to placental insufficiency, which is characterized by reduced uterine blood flow (UBF). During pregnancy, UBF increases tremendously as a result of increased maternal cardiac output, increased uterine artery (UtA) diameter, and trophoblast-driven modification of the uterine spiral arteries. 5,6 Reducing UBF is known to cause FGR in animals such as the pregnant rat, guinea pig, or sheep, emphasizing that fetal growth is dependent on UBF. 7
The therapeutic use of vascular endothelial growth factor (VEGF) has been explored. VEGF is a potent vascular endothelial cell mitogen with powerful vasodilatory and pro-angiogenic effects. These are mediated in part through the ability of VEGF to stimulate endothelial production of nitric oxide and prostacyclin via its major signaling receptor KDR (VEGF Receptor 2). 8 VEGF plays an important role in uteroplacental development, angiogenesis, and blood flow, and in FGR, maternal serum levels of VEGF are significantly lower than in normal pregnancy. 9,10 A targeted approach has been developed using an adenovirus vector (Ad) to overexpress VEGF locally in the uteroplacental circulation transiently. It has been demonstrated that this increases UBF, 11 –13 and is associated with improved fetal growth velocity in a sheep paradigm of FGR. 14 The placental physiology of sheep differs substantially from humans. 7 The guinea pig was selected for this study for a variety of reasons, including its hemomonochorial placentation, which is most similar to third-trimester human placentation, 15 homologous process of trophoblast invasion 15 and trophoblast cell proliferation, 16 and a longer gestation than other rodents, making it thus more suited for therapeutic evaluation. 7 Finally, periconceptual nutrient deprivation in the guinea pig impairs placental functional development, reduces the placental exchange and trophoblastic surface, increases the thickness of the exchange barrier, and causes a 40% reduction in fetal weight with brain sparing. 17,18 Ad.VEGF-A165-induced reversal of FGR in this model, where there is a general impairment of placental function, would make these studies relevant to human FGR where the lesion is rarely simply due to reduced UBF.
The primary aim of this study was to determine the effect of short-term overexpression of VEGF-A165 applied locally to the uteroplacental site on fetal growth velocity and weight at term. Secondary aims were to assess safety aspects and possible mechanisms of action of the therapy.
Materials and Methods
Animals
To induce FGR, female virgin Dunkin Hartley guinea pigs received 70% normal nutrient intake for 4 weeks before time-mating. 18 Pregnancy was confirmed using ultrasound. 18 Nutrient restriction was continued at 70% until day 35 of gestation, when it was increased to 90% until term at 60–65 days. Dams were given unlimited access to food during recovery from surgery. All procedures on animals were conducted in accordance with UK Home Office regulations and the Guidance for the Operation of Animals (Scientific Procedures) Act (1986), under Home Office Project Licence.
Animal surgery and vector injection
At mid-gestation (range 29–39 gestational days), under a general anesthetic (44 mg/kg ketamine and 5 mg/kg xylazine intramuscularly, maintained by 1.5–2.0% isoflurane inhalation), fetal measurements were taken 18 and a laparotomy was performed. Vector (1 × 1010 particles per animal), either control (Ad.LacZ) or treatment (Ad.VEGF-A165), suspended in freshly made thermosensitive gel (F-127; Sigma) was applied under direct vision to exposed UtA and radial arteries (RAs) supplying both uterine horns as described. 19 Dams were randomly assigned to VEGF or control vector and subsequent analyses were performed blinded to study group assignation. After a short period (3–8 days), or at term (60–64 gestational days), animals were killed for postmortem examination. Maternal and fetal measurements were recorded using electronic calipers (biparietal diameter [BPD], occipital snout length [OSL], crown rump length [CRL], and femur length [FL]) and a measuring tape (abdominal circumference [AC]), and tissues were extensively sampled. Fresh tissues were flash frozen in liquid nitrogen and stored at −80°C. Tissue for histological and immunohistochemical studies were fixed in 4% paraformaldehyde before being embedded in paraffin wax.
Enzyme Linked Immunosorbent Assay
Quantitative determination of VEGF protein expression in UtAs, RAs, and serum was performed using enzyme-linked immunosorbent assay (ELISA), as recommended by the manufacturer (DVE00; R&D Systems). Tissue samples were solubilized by homogenization in Radio ImmunoPrecipitation Assay (RIPA) Buffer (R0278; Sigma Aldrich). Protein content was determined using a Protein Assay Kit (500-0001; Bio-Rad). Serum was assayed undiluted.
Molecular analyses
Ad.VEGF-A165 vector expression was assessed using semi-nested reverse transcription polymerase chain reaction (PCR) in maternal and fetal tissues, with method and target primers as described. 12 Primers designed to amplify sequences within the guinea pig β-actin gene were used as a positive control. 20 The reaction conditions for both rounds were initial denaturation at 94°C for 4 min, followed by 30 cycles of denaturation at 94°C for 60 s, annealing for 64°C for 60 s, and extension for 72°C for 60 s, and a final extension period of 72°C for 10 min. PCR products were analyzed on a 1.2% agarose gel using GelRed fluorescent nucleic acid gel stains (VWR).
Biochemistry, hematology, and histological analysis
Blood was collected from dams and pups at necropsy. Standard small mammal biochemistry and hematology panels were performed by Royal Veterinary College Clinical Services Diagnostic Laboratories. Control blood was from ad libitum–fed dams from the same animal colony. Stained hematoxylin and eosin (H&E) maternal, fetal, and placental tissue sections were reviewed by an expert perinatal and placental pathologist who was blinded to experimental group. Trophoblast invasion was scored from 0 (no invasion) to 3 (maximal invasion), in UtAs and RAs.
Intima-to-media ratios
H&E-stained sections of the UtAs were imaged using a digital slide scanner (Nanozoomer; Hamamatsu). The relative thicknesses of the intimal and medial layers were determined with NDP.view software and used to calculate the intima:media ratios.
Immunohistochemistry
To evaluate neovascularization around the UtA and RAs, artery sections were immunostained with Anti-Von Willebrand Factor (1:200; A0082; Dako) using an ABC kit (Vector Laboratories), 3,30-diaminobenzidine (Sigma Aldrich), and a hematoxylin counterstain. Slides were imaged as above, and positively stained adventitial blood vessels were counted. To assess apoptosis and proliferation, placental sections were stained with anti-cleaved caspase 3 (1:800; 9664S; Cell Signaling) or anti-Ki-67(1:125; M724029-2; Dako), respectively. Stained placental sections were imaged using a digital slide scanner, as above. To obtain cell counts, each image was divided into six sections, and a field of view at 20 × magnification randomly captured from each section. Total positive cells per field of view were counted in the ImageJ program (National Institutes of Health) using the Cell Counter function.
Western blot
Protein extracts from the snap-frozen UtA tissues were used to estimate eNOS, iNOS, and phosph-eNOS levels by Western blotting. Tissue was lysed by sonication in RIPA buffer (R0278; Sigma), and the resulting lysates were analyzed by SDS-PAGE on 4–20% Tris-glycine gels (Novex EC6025BOX; Invitrogen), followed by electrotransfer onto nitrocellulose membranes (LC2001; Invitrogen). Membranes were blocked with 5% (w/v) bovine serum albumin (BSA; A9418; Sigma) in phosphate-buffered saline (PBS-T) for 1 h at room temperature, before being probed with the primary antibody by overnight incubation at 4°C, followed by incubation for 1 h at room temperature with a horseradish peroxidase (HRP)-linked secondary antibody (1:8,000; Santa-Cruz Biotechnology) and detection using enhanced chemiluminescence (ECL) reagents (GE Healthcare), following the manufacturer's protocol. Immunoblots were quantified by scanning of films with a calibration strip and analysis by densitometry using ImageJ (National Institutes of Health). The primary antibodies used were monoclonal mouse anti-eNOS/iNOS Type III antibody (1:3,000; 610296; BD Transduction Laboratories), monoclonal mouse anti-iNOS/NOS Type II antibody (1:3,000; 610328; BD Transduction Laboratories). GAPDH was probed using a goat polyclonal, GAPDH antibody (V-18)-HRP conjugate (1:1,000; 20357; Santa Cruz Biotechnology).
Myography
Cleared UtA segments 1–2 mm in length were mounted in a two-chambered wire myograph (410A; Danish Myotechnology) run using Myodaq.2.vi software, and maintained according to standard protocols. 21 Concentration response curves to phenylephrine (PE) and acetylcholine (ACh) were constructed for each vessel.
Statistics
Sample size calculations were based on the primary outcome of fetal birthweight, and a sample data set of birth-weight data was obtained from 29 normal dams and 89 offspring. A generalized linear mixed models (GLMM) approach was used for statistical analysis as described, 22 with dam and treatment (Ad.LacZ or Ad.VEGF-A165) set as a factor, with fetus sex, litter size, and gestational age (gestational days) at postmortem as covariates. Fetus sex was found to have no effect and was removed as a covariate. GLMM statistical analysis was needed for the set of data, as the model is predicting random effects in addition to the usual fixed effects, with non-normal data. Statistical significance was considered to be achieved if p < 0.05.
Results
Seven dams treated with Ad.LacZ delivered of 19 pups and eight dams treated with Ad.VEGF-A165 delivered of 26 pups were included in the study. A further 10 females underwent surgery but were excluded from the study (Supplementary Tables S1 and S2; Supplementary Data are available online at
Fetal measurements
There was no significant difference between the groups in any fetal measurements taken at mid-gestation by detailed fetal ultrasound before Ad vector treatment. Treatment with Ad.VEGF-A165 resulted in a significant increase in fetal weight at term (88.01 ± 13.36 g) compared with control Ad.LacZ-treated fetuses (85.52 ± 13.00 g; p = 0.028; Fig. 1 and Tables 1 and 2). There was no effect of treatment on placental weight or diameter, although placental depth was increased (Table 1 and Fig. 1). Placental efficiency, as indicated by the placental:fetal weight ratio, was not significantly different (Table 1). Other fetal growth measurements were significantly increased by Ad.VEGF-A165 treatment (Table 2), including the brain, liver, and lung weight and CRL. Kidney weight, heart weight, and BPD were not affected. Ad.VEGF-A165 increased the fractional growth rate of the BPD and OSL head measurements and the brain-to-fetal weight ratio at term compared with Ad.LacZ treatment (Table 2). The fractional growth rate of the AC, CRL, and FL, and the placental diameter and thickness were not different between the groups (data not shown).
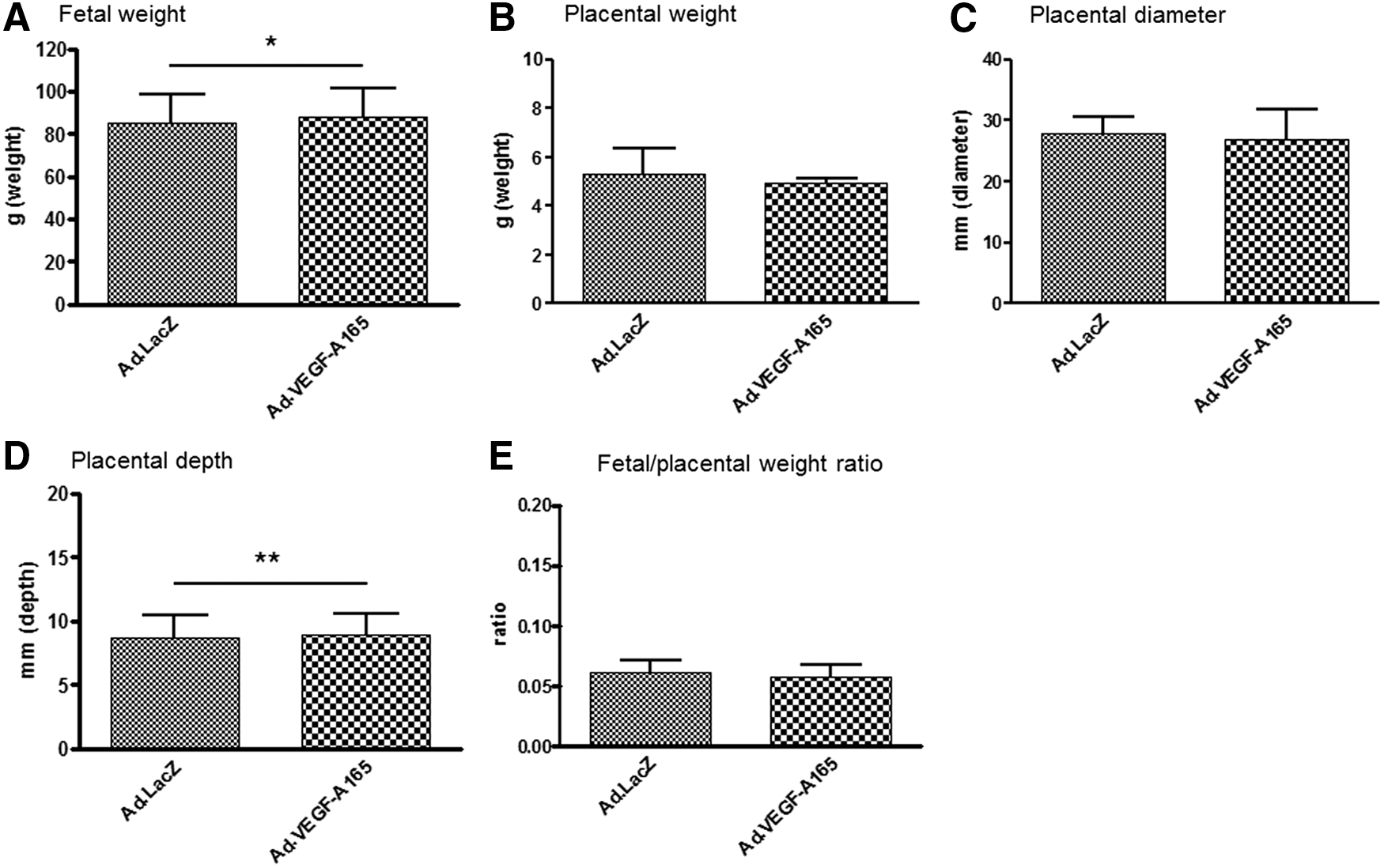
Fetal weight
Fetal weight and placental measurements of FGR pregnancies following mid-gestation treatment with Ad.LacZ or Ad.VEGF-A165
All data are means ± standard deviation. Data were analyzed by generalized linear mixed models.
n = 19 pups from seven dams.
n = 26 pups from eight dams.
p < 0.05; ** p < 0.01.
FGR, fetal growth restriction.
Fetal size characteristics and fractional growth rate in pregnancies affected by FGR following mid-gestation maternal uterine artery treatment with Ad.LacZ or Ad.VEGF-A165
All data are means ± standard deviation. Data were analyzed by generalized linear mixed models.
n = 19 pups from seveb dams.
n = 26 pups from eight dams.
p < 0.05; ** p < 0.01.
BPD, biparietal diameter; CRL, crown rump length; OSL, occipital snout length; AC, abdominal circumference.
VEGF protein expression
VEGF expression following treatment with Ad.VEGF-A165 was confirmed by ELISA in collected samples. Protein was detected in maternal and fetal serum, with higher levels detected in fetal serum from dams treated with Ad.VEGF-A165 (maternal n = 6, fetal n = 5) than those treated with Ad.LacZ (maternal n = 3, fetal n = 4; Fig. 2A). VEGF protein was also detected in UtA samples (Ad.VEGF-A165 n = 9 samples from six animals), but levels were not significantly higher compared to Ad.LacZ samples (n = 3 samples from six animals; Fig. 2B). Results were similar in the RA samples (Ad.VEGF-A165 n = 7 samples from six animals; Ad.LacZ n = 4 samples from three animals; Fig. 2C).
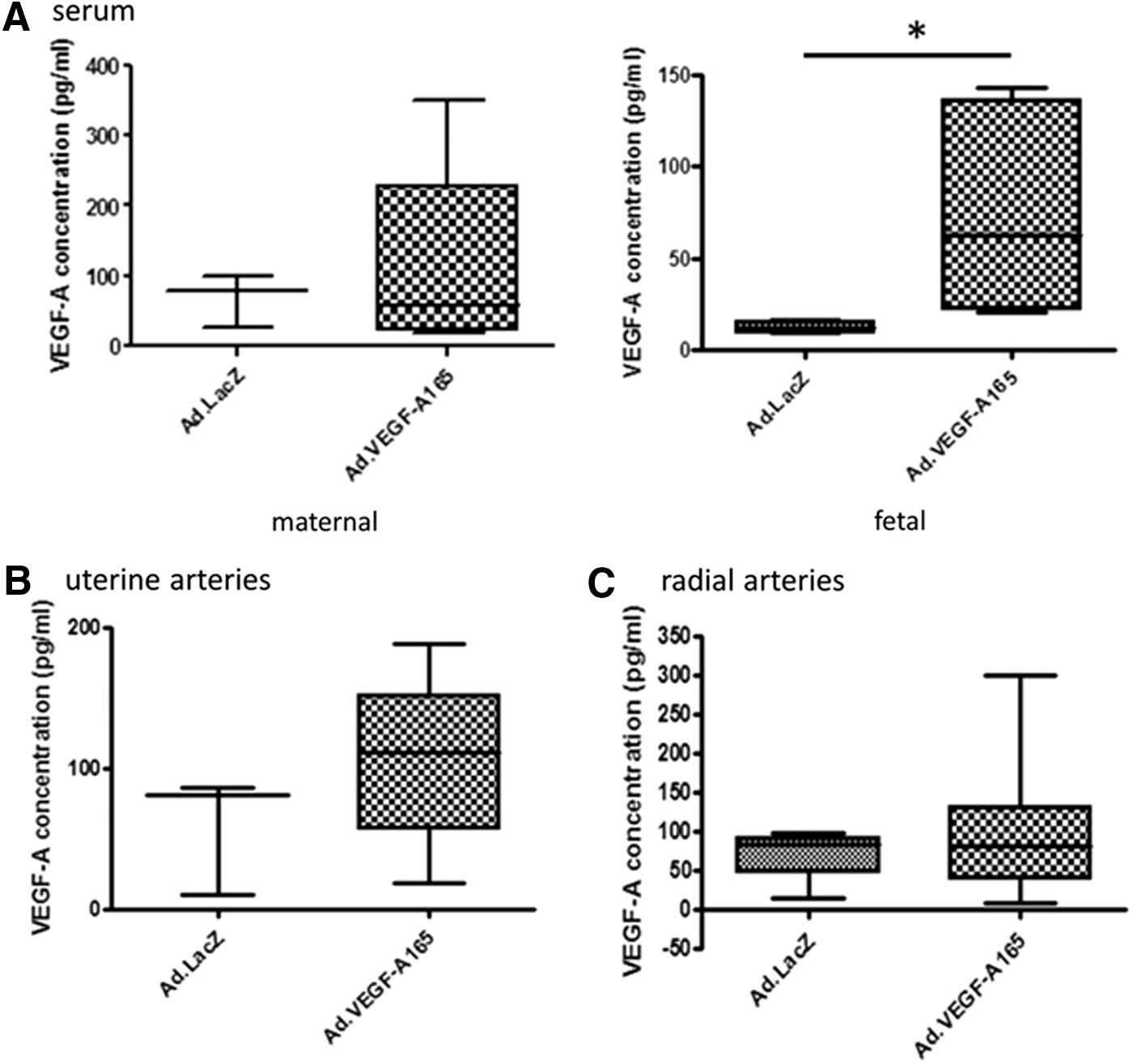
Enzyme-linked immunosorbent assay measurement of VEGF-A in the serum (
Immunohistochemical analysis for VEGF expression, also at 3–8 days post therapy, showed specific staining of VEGF around the target vessels in both the Ad.LacZ- and the Ad.VEGF-A165-treated animals, with a visible increase in Ad.VEGF-A165-treated UtAs (Fig. 3A and D). No detectable difference was found in VEGF receptor 1 or 2 expression in the UtAs of Ad.LacZ- and Ad.VEGF-A165-treated animals (Fig. 3B, C, E, and F). Overall immunohistochemical staining was more intense for all three proteins in RAs, with no clear difference between Ad.LacZ- and Ad.VEGF-A165-treated animals (data not shown).
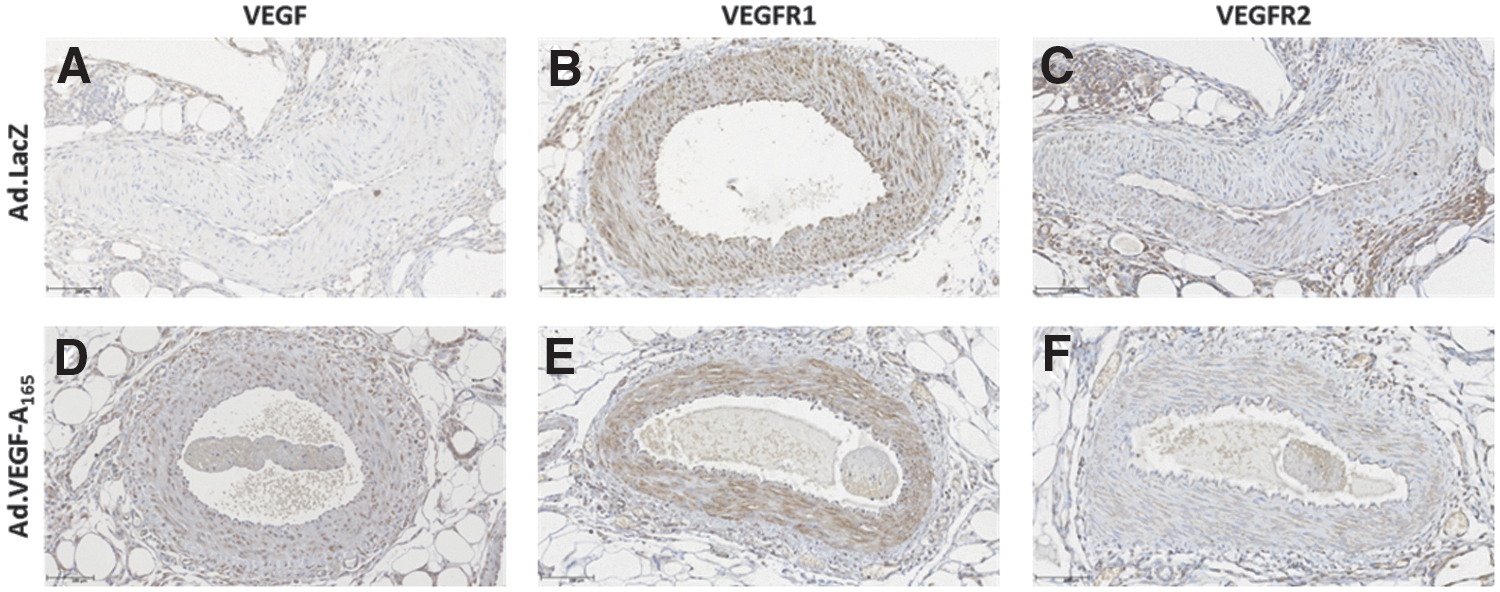
Immunohistochemistry for localization of expression of VEGF (
Vector spread
RT-PCR analysis showed the presence of the transgene in the target UtAs and RAs of three of four Ad.VEGF-A165-treated animals (Fig. 4). All other tissues tested (maternal: UtA, RAs, uterus, lung, heart, ovary, liver, adrenal gland, kidney, spleen, brain, retina, and thymus; fetal: placenta, membranes, umbilical cord, lung, thymus, heart, spleen, liver, gonads, adrenal gland, kidney, muscle, skin, brain, and bone marrow) were negative, with the exception of one maternal ovary sample.

Reverse transcription polymerase chain reaction (RT-PCR) to examine vector spread. (
Histological, hematological, and biochemical evaluations
A standard small mammal hematology panel returned similar values for all term maternal serum submitted (VEGF n = 3, LacZ n = 1, ad libitum n = 3), within the standard range for the panel (Pathology Department, Royal Veterinary College, Hawkshead, United Kingdom) and comparable to published guinea-pig values. 23 Fetal values differed as expected from maternal values, but there were no differences between values from fetuses of ad libitum (n = 3 from three dams), Ad.VEGF-A165 (n = 7 from four dams), and Ad.LacZ (n = 2 from one dam) subjects, with the exception of mean corpuscular volume (MCV), which was slightly elevated in the Ad.VEGF-A165-treated fetuses (93.1 ± 8.0 fL) compared to Ad.LacZ-treated and ad libitum fetuses (82.0 ± 6.8 fL). In biochemistry tests, there were decreased levels of total protein and globulin and increased levels of potassium compared with the standard ranges for small mammals and guinea pigs as before. 23 However, these differences were preserved across all groups (Ad.VEGF-A165 n = 5, Ad.LacZ n = 1, ad libitum n = 5). As with the hematology, there were no difference between values from fetuses of ad libitum (n = 4 from four dams), Ad.VEGF-A165 (n = 8 from five dams), and Ad.LacZ (n = 2 from one dam) subjects.
Microscopic histological examination of dams (n = 2 each group), fetuses (n = 2 each group), and placentas (control n = 6, Ad.VEGF-A165 n = 7) did not reveal any pathological observations. The proportion of animals with moderate or maximum (scored 2 or 3) trophoblast invasion in the uterine spiral arteries did not differ between dams treated with Ad.VEGF-A165 (4/10 dams) and Ad.LacZ (2/6 dams; p = 1.00). Neither was there a difference between treatment groups in the proportion of dams with moderate or maximal trophoblast invasion in the RAs (Ad.LacZ 4/4 dams, Ad.VEGF-A165 3/5 dams; p = 0.44).
Mechanism of action
Myography experiments on UtA segments sampled at postmortem examination 38 ± 5 days post vector application showed no change in the mean contractile response to PE in the Ad.VEGF-A165 (n = 7) transduced vessels when compared with Ad.LacZ vessels (n = 5; Fig. 5A). Neither was there an overall change in the relaxation of vessels in response to ACh as analyzed by two-way analysis of variance with repeated measures (Fig. 5B). However, at high concentrations (ACh = 3 × 10–8, 1 × 10–9 M) prior to saturation, there was significantly more relaxation, with Emax values of 72.7 ± 5.8 for Ad.VEGF-A165 compared with 50.5 ± 3.5 for Ad.LacZ (p = 0.050).
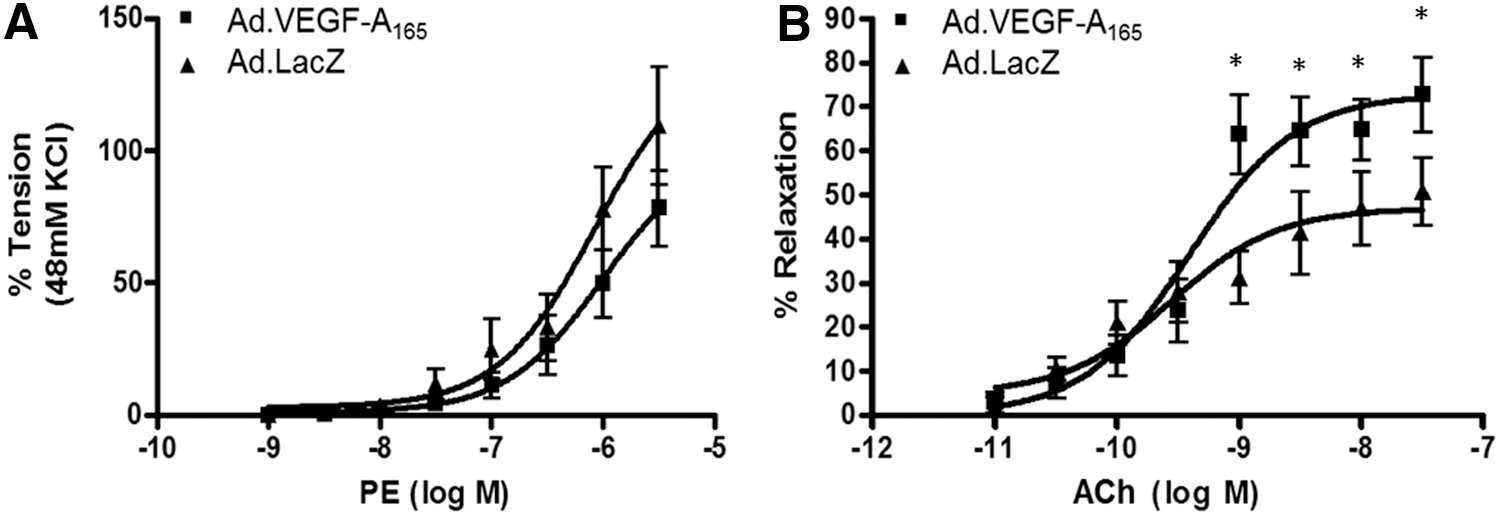
Vascular reactivity of uterine arteries 3–8 days after vector administration measured by wire myography. (
No difference was observed in intima:media ratios in UtAs between Ad.LacZ- (n = 7 arteries from four animals) and Ad.VEGF-A165-treated animals (n = 11 arteries from six animals), measured on H&E-stained sections (data not shown). The finding was the same in the RAs, where no differences between Ad.LacZ- (n = 24 arteries from four animals) and Ad.VEGF-A165-treated animals (n = 15 arteries from four animals) were seen (data not shown).
An increase was detected in the levels of eNOS and iNOS between the Ad.LacZ (n = 3) and Ad.VEGF-A165 (n = 3) transduced UtAs by Western blotting 3–8 days after gene transfer (Supplementary Fig. S1). eNOS and iNOS expression was significantly higher in UtAs treated with Ad.VEGF-A165 compared with Ad.LacZ (eNOS: 1.58 ± 0.2 [Ad.VEGFA165] vs. 0.69 ± 0.18 [Ad.LacZ], p < 0.001; iNOS: 2.9 ± 0.59 [Ad.VEGFA165] vs. 0.81 ± 0.1 [Ad.LacZ], p < 0.001).
Ad.VEGF-A165 transduction increased the average number of Von Willebrand Factor–stained adventitial blood vessels around the UtAs (77.5 ± 38.6, n = 12 arteries from eight animals) compared with Ad.LacZ transduction (32.5 ± 14.2, n = 8 arteries from five animals; p = 0.006; Fig. 6A). In the RAs, the difference in number of adventitial vessels failed to reach statistical significance (Ad.VEGF-A165: 26.4 ± 19.2, n = 28 arteries from eight animals; Ad.LacZ: 16.4 ± 13.5, n = 20 arteries from six animals; p = 0.051). The number of vessels per mm2 of adventitial tissue was lower in RAs compared with the UtAs (Fig. 6B).
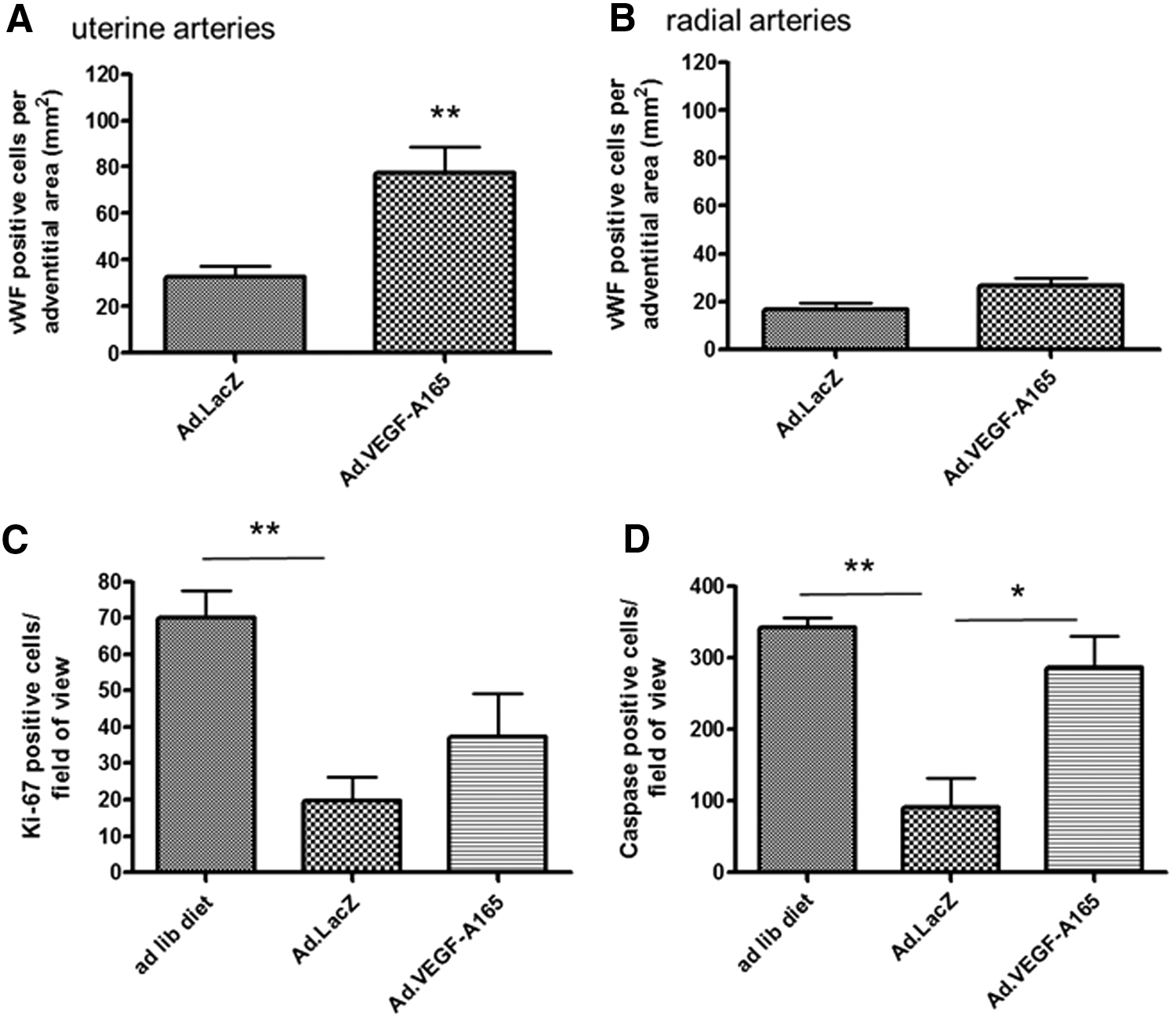
Immunohistochemistry analysis of endothelial cell proliferation and placental turnover. Von Willebrand Factor staining for vessels in the adventitia of uterine (
The restricted diet had a significant effect on both proliferation and apoptosis in the placenta. Proliferation was reduced in Ad.LacZ- (19.8 ± 20.4 cells/field of view, n = 5 placentas from five dams) and Ad.VEGF-A165-treated animals (37.3 ± 34.2 cells/field of view, n = 4 placentas from four animals) compared with ad libitum animals (70.1 ± 25.0 cells/field of view, n = 6 placentas from three dams), as measured by positive Ki-67 staining (p = 0.001; Fig. 6C). Similarly, apoptosis was reduced in Ad.LacZ (90.7 ± 128.5 cells/field of view, n = 5 placentas from five dams) nutrient restricted animals compared with ad libitum animals (342.8 ± 45.4 cells/field of view, n = 6 placentas from three dams), as measured by positive cleaved caspase-3 staining (p = 0.001; Fig. 6D). Following Ad.VEGF-A165 treatment, however, there was a significant increase in the number of cleaved caspase-3-stained cells (287 ± 114.8 cells/field of view, n = 4 placentas from four animals; p = 0.025), such that there was no difference between ad libitum and Ad.VEGF-A165-treated groups (p = 0.179).
Discussion
Using a guinea-pig model of FGR, this study provides further evidence that mid-gestation maternal gene therapy giving transient local overexpression of VEGF in the uteroplacental circulation improves fetal growth rate and fetal weight at term. The results of animals treated with Ad.VEGF-A165 Ad.LacZ vector were compared with two contemporaneous groups of (1) normally fed non-FGR guinea pigs and (2) untreated maternal nutrient-restricted FGR guinea pigs, as reported previously. 18 Short-term VEGF expression was confirmed in fetal and maternal blood and in transduced maternal uterine vessels without detectable vector transmission to the fetus. There was upregulation in eNOS and iNOS detected in transduced vessels, and enhanced vascular relaxation short term. Guinea pigs have a hemomonochorial placentation, which is the most similar to human placentation in an animal model, with the exception of some non-human primates. In contrast to the work in sheep, this model is also closer to the human condition, in that the FGR is early rather than late onset, and therapy is applied mid-gestation at approximately the same stage as would be feasible in humans (therapy 29–39 gestational days, term 65 gestational days vs. therapy 20–26 weeks of gestation, term 40 weeks of gestation). Thus, experimental results obtained from pregnant guinea pigs are likely to hold great relevance for clinical translation.
Although the mean weight increase is a modest 3%, there was nevertheless a significant improvement in fetal growth rate in fetuses receiving Ad.VEGF-A165. Even for normally grown fetuses born very premature, small increases in fetal growth and gestation at birth are associated with major improvements in survival and morbidity. 3,4 Therapeutic interventions for severe FGR might therefore be successful by improving outcome if they lead to even small increases in fetal growth, or later gestation at delivery. Several other fetal measurements were significantly increased in response to Ad.VEGF-A165 gene therapy, with treated fetuses having an increased CRL and brain and liver weight, although no change in the AC was detected. This is in contrast to ultrasound measurement findings from previous research in sheep 14 and from other groups studying the FGR mouse. 22 Placental weight, diameter, and area were not affected by Ad.VEGF-A165 treatment, with only placental depth being increased. The gain in fetal weight did not therefore appear to be mediated through an increase in placental dimensions.
Delivery of Ad.VEGF-A165 to the UtA and RAs increased expression of VEGF shortly after vector delivery, showing efficient transduction of the vector and high concentration of VEGF as detected in fetal blood serum, as well as a mild increase in maternal blood serum. It is proposed that released VEGF-A165 protein promoted an increase of eNOS and iNOS levels via functional activation of VEGFR-2, potentially being the factor leading to angiogenesis and hypotension in the target vessels. 24 At this stage, it is not possible to speculate about the mechanism by which Ad.VEGF-A165 induces neovascularization in the UtA and RA system, as further studies are needed. Immunohistochemistry of the guinea-pig UtA and RAs showed increased VEGF staining in Ad.VEGF-A165-treated animals, supporting the ELISA data. There was also positive staining for VEGR-1 and VEGFR-2. Thus, either or both of these receptors could have been involved in mediating the effects seen following Ad.VEGF-A165 treatment.
UtAs from Ad.VEGF-A165-treated dams demonstrated greater relaxation in response to ACh when examined in a myograph. This complements the authors’ previous work in sheep, where the relaxation response in UtAs collected at short-term time points was significantly increased in Ad.VEGF-A165-transduced vessels. 11 Others have observed in vitro that VEGF-A165 is a powerful vasodilator of the fetoplacental vasculature, 25 and adult vascular tone is regulated via VEGF-A induced vasodilation. 26 ACh-induced relaxation of guinea-pig UtA is dependent on the release of nitric oxide from vascular endothelial cells. 27
Enhanced UtA vascularization was also observed via immunohistochemical staining of new blood vessels in the adventitia. In animal experiments, VEGF gene transfer is capable of inducing therapeutic angiogenesis in diverse tissues and organ systems. Overexpression of VEGF using viral vectors can stimulate significant neovascularization in skeletal muscles and myocardium (reviewed in Rissanen and Ylä-Herttuala 28 ). The authors’ own work suggests that an increase in UBF is likely to be mediated in the longer term through neovascularization within the perivascular adventitia. 12,13
Safety will be an important consideration for translation into the clinic. Reassuringly, there was no transgenic protein expression in any fetal tissues, as determined by RT-PCR. Only one maternal sample of non-target tissue tested positive, from an ovary sample, likely due to an inadvertent positioning of the gel/vector mixture proximal to the ovary during surgery.
Pluronic gel has been used previously as a delivery vehicle for adenoviral vectors to the carotid artery of rats. 29 –31 It has been demonstrated that external vascular application of Ad. vector combined with pluronic gel achieves very high levels of localized gene transfer and expression specifically at the site of administration in the UtAs in guinea pigs compared with direct intravascular injection. 19 The relatively smaller size of the UtA in guinea pigs compared with sheep limited the feasibility of intravascular injection. In pregnant women, however, transfemoral UtA catheterization with temporary balloon occlusion of the vessel lumen could be used as a minimally invasive technique.
FGR in sheep and humans is associated with smaller placentas and reduced cellular proliferation. 32 –34 Similarly, this study found a significant reduction of proliferation in the placenta of the nutrient-restricted guinea pigs. Treatment with Ad.VEGF-A165 did not affect the level of proliferation compared with Ad.LacZ-treated animals, as both remained lower than ad libitum–fed animals. FGR in humans is associated with increased placental apoptosis. In this study, however, the opposite effect was found. Diet-restricted animals treated with Ad.LacZ had significantly less apoptosis in the placenta than animals fed ad libitum. However, treatment with Ad.VEGF-A165 restored apoptosis back to ad libitum levels. Whether this is a phenomenon confined to the guinea pig is unknown. It is unlikely to be a product of the Ad. treatment, as no evidence of vector was found in any placental samples.
The degree of rescue of FGR seen in this study in nutrient-restricted FGR guinea pigs did not appear to be as much as in the previous study in sheep where FGR was created in the overnourished adolescent ewe. 14,35 There are differences in the animal models and experimental techniques that may explain this. The overnourished FGR sheep is a highly manipulated and controlled experimental model where exclusively singleton pregnancies with maximum genetic homogeneity are created by in vitro fertilization using eggs from superovulated donor ewes inseminated by a single sire with embryo transfer into adolescent ewes that are subsequently overfed. In contrast, the nutrient-restricted FGR guinea pig conceives over a 3–5 day period, carries between one and five pups, and is genetically heterogeneous, 7,18 resulting in a wider range of offspring weights and gestational ages than is found in the FGR sheep. The vector was applied externally to the UtAs and RAs of the guinea-pig circulation by necessity because of the small vessel size, using a thermolabile pluronic gel technique 19 rather than a direct UtA injection with stasis of blood flow used in the sheep study, which delivers the total dose concentrated to the vessel. In addition, the vector dose was double in the sheep experiments: 1 × 1010 vp per guinea pig (1 × 1010 per kg assuming a dam weight of 1 kg) compared with 1 × 1011 vp per sheep (2 × 1010 per kg assuming a ewe weight of 50 kg). To ensure optimum gene transfer, in the clinical trial, the plan is to use an intravascular injection technique with doses comparable to those used in the sheep study.
This study has confirmed that maternal gene therapy with Ad.VEGF-A165 is able to increase the fetal growth rate and term weight of fetuses in an animal model with placentation relevant to human FGR. Transduction with Ad.VEGF-A165 appears to achieve this outcome by increasing UtA vasorelaxation and promoting adventitial angiogenesis, with virtually no vector spread outside target tissues. Further work in this model to examine the long term effects of Ad.VEGF-A165 therapy in treated offspring will lend weight to applications for translation to the clinic.
Footnotes
Acknowledgments
We thank the BSU staff at the RVC and in particular Adnan Avdic-Belltheus for assistance with animal husbandry. We thank Paul Bassett for statistical advice. Funding for the study was received from the following sources: Action Medical Research (SP4409), UCL/UCLH NIHR Comprehensive Biomedical Research Centre (ALD, DMP), Rosetrees Trust and Magnus Life Science (grant to fund AS salary), The British Heart Foundation (ICZ), Ark Therapeutics Oy, Kuopio, Finland (supplied adenovirus vectors free of charge). The funders played no part in study design, data collection, data analysis, manuscript preparation, and/or publication decisions.
Author Disclosure
A.L.D. and D.M.P. are consultants for Magnus Growth, for which they receive a token consultancy payment and shareholding in the company. A.L.D. is director of Magnus Growth, part of Magnus Life Science, which is aiming to take to market a novel treatment for FGR. I.Z. is a consultant for Magnus Life, part of Magnus Life Science for which he receives a token consultancy payment and shareholding in the company. No competing financial interests exist for the remaining authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.