
Abstract
Osteoporosis is a common health problem severely affecting the quality of life of many people, especially women. Current treatment options for osteoporosis are limited due to their association with several side-effects and moderate efficacy. Therefore, novel therapies for osteoporosis are needed. This study tested the feasibility of adipose tissue–derived mesenchymal stem cell (ATMSC)-based human alpha-1 antitrypsin (hAAT, SERPINA1) gene therapy for the prevention of bone loss in an ovariectomized (OVX) mouse model. Eight-week-old female C57BL6 mice underwent ovariectomy and were treated with hAAT (protein therapy), ATMSC (stem-cell therapy), ATMSC + hAAT (combination of ATMSC and hAAT therapy), and ATMSCs infected with lentiviral vectors expressing hAAT (ATMSC-based hAAT gene therapy). The study showed that lenti-hAAT vector–infected ATMSCs (ATMSC-LV-hAAT) produced high levels of hAAT. Transplantation of these cells significantly decreased OVX-induced serum levels of interleukin 6 and interleukin 1 beta, and receptor activator of nuclear factor kappa B gene expression levels in bone tissue. Immunohistological analysis revealed that transplanted cells migrated to the bone tissue and secreted hAAT. Importantly, bone microstructure analysis by microcomputerized tomography showed that this treatment significantly protected against OVX-induced bone loss. The results suggest a novel strategy for the treatment of osteoporosis in humans.
Introduction
O
With respect to bone formation and remodeling, adipose tissue–derived mesenchymal stem cells (ATMSC) have received significant interest over the past few years due to their easy isolation, abundance, and ability to differentiate into lineages of mesodermal cells and tissues such as adipocytes, bone, cartilage, and skeletal muscles. 1,5 Cho et al. showed that human adipose-derived stromal cell therapy protected against ovariectomy (OVX)-induced bone loss. 2 Another group demonstrated that transplantation of human adipose–derived stem cells promoted osteoblast and osteoclast function and ultimately stimulated bone repair. 6 Moreover, MSC-mediated gene therapy has been investigated in a mouse model of osteoporosis, and it was shown that transplantation of bone-marrow MSCs overexpressing receptor activator of nuclear factor kappa B (RANK)-Fc or CXCR4 increased in vivo cell trafficking to bone in OVX mice and reduced bone loss. 7
In osteoclast-associated bone resorption, human and animal studies showed that proinflammatory cytokines such as tumor necrosis factor alpha (TNF-α), interleukin (IL)-6, and IL-1β played critical roles by regulating receptor activator of NF-κB (RANK) ligand/RANK/osteoprotegerin (OPG) pathways. 8 –11 Therefore, a targeting strategy that reduces proinflammatory cytokines TNF-α, IL-6, and IL-1β is one of the logical approaches to reducing osteoclast-associated bone resorption. Human AAT (hAAT) is a multifunctional protein with both proteinase inhibitory and anti-inflammatory activities. It inhibits lipopolysaccharide-induced release of TNF-α and IL-1β from human monocytes, and enhances the production of the anti-inflammatory cytokine IL-10. 12,13 Importantly, in clinical studies, it was found that there is a high risk of osteoporosis in hAAT-deficient patients. 14 It has been shown that hAAT reduces OVX-induced bone loss in mice. 15 It has also been shown that rAAV vector-mediated hAAT gene therapy ameliorated bone loss in an OVX-induced osteoporosis mouse model. 16
This study investigated the combined effects of ATMSC and hAAT by either co-administration of ATMSC and hAAT (ATMSC + hAAT) or transplantation of ATMSCs expressing hAAT (ATMSC-mediated hAAT gene therapy) in an OVX-induced mouse model of osteoporosis. It was found that combination therapy (ATMSC + hAAT) and ATMSC-mediated hAAT gene therapy showed greater protective effects than individual therapies (only hAAT or only ATMSC), and intriguingly it was observed that ATMSC-mediated gene therapy showed efficient protection, although the number of transplanted ATMSCs expressing hAAT was lower than that of cells transplanted in combination therapy.
Materials and Methods
Mouse ATMSCs isolation and culture
ATMSCs were isolated from C57BL/6 mice, as previously described. 17 These cells (1–2 × 106) were plated on a 100 mm tissue culture plate and cultured in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St. Louis, MO) supplemented with 10% fetal bovine serum (FBS; Corning, Manassas, VA) and 1% penicillin/streptomycin (Corning) for 12–16 h in a humidified atmosphere of 5% CO2. Non-adherent cells were washed with phosphate-buffered saline (PBS; Corning) and cultured in DMEM supplemented with 10% FBS and 1% penicillin/streptomycin. Cell culture medium was changed every 3 days. When 70–80% cell confluency was achieved, the cells were harvested with 0.25% trypsin-EDTA solution (Sigma-Aldrich), and 1–2 × 106 cells were cultured on a 100 mm tissue culture plate. Cells at passage 4 were taken for transplantation or lentiviral transduction.
Flow cytometry
Flow cytometric analysis of ATMSCs at passage 4 was carried out with FACSCaliber Cell Quest Pro v5.2.1 (BD Biosciences, San Jose, CA), and data were analyzed using FCS Express v4 (De Novo Software, Los Angeles, CA) at the University of Florida Flow Cytometry Core. Phycoerythrin (PE) conjugated anti-mouse CD105, CD 44, CD 31, and CD45 were used. All were purchased from e-Bioscience (San Diego, CA). Dead cells stained with 7-amino-actinomycin D (7-AAD; e-Bioscience) viability staining solution were excluded from the analysis. A gate was set to live cells, and the percentage of cells stained with specific antibodies was analyzed.
Lentiviral vector production and transduction to ATMSCs
Lentiviral vector–expressing hAAT (lenti-hAAT) was generated, as previously described. 18 ATMSCs at passage 4 were infected with 6.6 multiplicity of infection (MOI; 3.3 × 106 virus for 0.5 × 106 cells) for 2 h with serum-free DMEM (Sigma-Aldrich). After 2 h of incubation in a humidified atmosphere of 5% CO2, cells were cultured with DMEM supplemented with 10% FBS and 1% penicillin/streptomycin for 2 days. Lentiviral-transduced ATMSCs expressing hAAT (ATMSC-LV-hAAT) were trypsinized with 0.25% trypsin-EDTA solution and resuspended 0.6 × 106 cells in 200 μL of 0.9% saline solution. This cell suspension was freshly produced immediately before the intraperitoneal (i.p.) injection to the experimental mice.
Animals
Seven-week-old bilaterally OVX and sham-operated C57BL/6 mice were purchased from Charles River Laboratories (Wilmington, MA). One week after the ovariectomy, the mice were shipped to the University of Florida animal care facility. They were housed in a specific pathogen-free conditions under a 12 h light/dark cycle. The University of Florida Institutional Animal Care and Use Committee (IACUC) approved all of the animal protocols for this study.
Human AAT (hAAT), ATMSC, and ATMSC-LV-hAAT administration
The mice were housed for 1 week for acclimatization after which they were randomly divided into five groups: PBS-injected OVX group (n = 10; i.p. injection of 200 μL of PBS every 3 days for 8 weeks), which served as the osteoporosis model group; hAAT protein–injected OVX group (n = 9; Prolastin C®; Telecris Biotherapeutics, Inc., Research Triangle Park, NC; i.p. injection of 2 mg/mouse in 200 μL of PBS every 3 days for 8 weeks); ATMSC-transplanted OVX group (n = 10; two i.p. injections every 4 days: 1 × 106 cells in 200 μL of PBS in 1st injection and 0.8 × 106 cells in 200 μL of PBS in 2nd injection; all cells were freshly harvested); ATMSCs and hAAT (ATMSC + hAAT)–injected OVX group (n = 10; two i.p. injections for ATMSCs every 4 days: 1 × 106 cells in 200 μL of PBS in 1st injection and 0.8 × 106 cells in 200 μL of PBS in 2nd injection; all cells were freshly harvested; 2 mg/mouse hAAT in 200 μL of PBS every 3 days for 8 weeks), and ATMSC-LV-hAAT-injected OVX group (n = 3; one i.p. injection: 0.6 × 106 cells in 200 μL of 0.9% saline). Sham-operated mice (n = 10) were i.p. injected with PBS (200 μL/mouse) and used as a model control. In the hAAT-injected group, one mouse died at week 2, and data from this mouse were excluded from analysis.
Sample collection and preparation
Blood samples were collected biweekly by facial vein bleeding, and samples were stored at −80°C until further analysis. After 8 weeks of treatment, all animals were sacrificed by cervical dislocation. The vertebrae and right femur of each mouse were removed and cleaned of adherent tissue. The right femur was flash-frozen in liquid nitrogen and stored at −80°C until further analysis. These femurs were pulverized with a liquid nitrogen–cooled steel mortar and pestle in order to isolate the total RNA, as described below. Before scanning with the microcomputerized tomography (μCT) scanner to evaluate the bone microstructure, the vertebrae were stored at −20°C.
Detection of serum proinflammatory cytokines (IL-1β, IL-6, and TNF-α), phosphate, calcium, and alkaline phosphatase
Using a murine enzyme-linked immunosorbent assay (ELISA) kit (Peprotech, Rocky Hill, NJ), serum levels of the proinflammatory cytokines IL-1β, IL-6, and TNF-α were measured. Serum calcium and phosphate levels were determined by calcium and phosphate colorimetric detection assay kits (Abcam, Cambridge, United Kingdom; catalog no. ab102505 for calcium and ab65622 for phosphate), and serum alkaline phosphatase (ALP) was measured by ALP assay kit (Takara, Otsu, Japan; catalog no. MK301). The manufacturers' instructions were followed in all the detection procedures.
Detection of serum levels of hAAT and anti-hAAT antibodies
Serum hAAT and anti-hAAT levels were detected by ELISA, as previously described. 19 –21
Real-time polymerase chain reaction
For the determination of RANK (TNFSF11A), RNAKL (TNFSF11), integrin β3 (TGB3), and GAPDH (Gapdh) gene expression, total RNA from the right femur bone was purified, and cDNA was synthesized and subjected to real-time polymerase chain reaction (PCR), as previously described. 16,22
Determination of bone microstructure by μCT
The 2nd lumbar vertebrae (L2) from each mouse were cleaned of adherent tissue, and the bone microstructure was determined, as described previously. 16,22 Different bone-morphometric indexes were considered to determine the bone microstructure: bone volume/total volume (BV/TV), which indicates the fraction of a given volume of interest (TV) that is occupied by mineralized bone (BV); connectivity density (Conn.Dn), which is a measure of the degree of connectivity of trabeculae, normalized to the TV; trabecular number (Tb.N), which is a measure of the average number of trabeculae per unit length; bone mineral density (BMD), which is the amount of bone mineral in bone tissue; trabecular thickness (Tb.Th), which is the mean thickness of trabeculae; and structure model index (SMI), which is an estimation of the plate-rod characteristic of the structure.
Histopathological examinations and immunohistochemistry for hAAT
Immediately after euthanasia, multiple tissues, including the heart, liver, muscle, brain, gastrointestinal tract, pancreas, lung, kidney, uterus, spleen, and left femur, were collected from each mouse. One age-matched healthy female mouse was used as a normal control for histopathological analysis. Tissues were fixed in 10% neutral-buffered formalin, processed for paraffin blocks, sectioned at 5 μ, and stained with hematoxylin and eosin. Tissue sections from one ATMSC and one ATMSC-LV-hAAT-injected OVX mouse were also immunostained with antibodies directed to hAAT, as described previously. 18 Tissues were examined microscopically, and tissue architecture was scored using a scoring system of 0–5, where 0 = no lesion, 1 = minimal change, 2 = mild change, 3 = moderate change, 4 = severe change, and 5 = markedly severe change. Digital images were captured using a Nikon Eclipse Ci microscope equipped with a Nikon DS-Fi2 digital camera.
Statistical analysis
For statistical data analysis, GraphPad Prism5 software was used. One-way analysis of variance followed by Dunnett's multiple comparison test was used to determine the statistical significance between treatment group and PBS-injected group. An unpaired two-tailed Student's t-test was also used to test for differences between two samples. Data are expressed as mean ± standard error of the mean (SEM). p-Values of <0.05 were considered as statistically significant.
Results
Characterizations of AT-MSCs
In previous studies, a detailed characterization of ATMSCs has been performed, and it has been shown that these cells have adipogenic and osteogenic potential, and can differentiate into hepatocyte-like cells after transplantation. 17,21 In addition, these cells can be reprogrammed to iPSCs. 23 Although the same isolation procedure has been used in the present study, ATMCSs were characterized by examining the lineage-specific cell surface markers by fluorescence-activated cell sorter (Fig. 1A–F) to ensure the quality of the cells. The ATMSCs expressed stromal-associated markers CD105 (68%) and CD44 (95.25%), and they were negative for the endothelial cell marker CD31 (2.5%) and hematopoietic marker CD45 (2.4%). The percentage of CD105-positive cells indicated that high-quality ATMSCs were obtained.
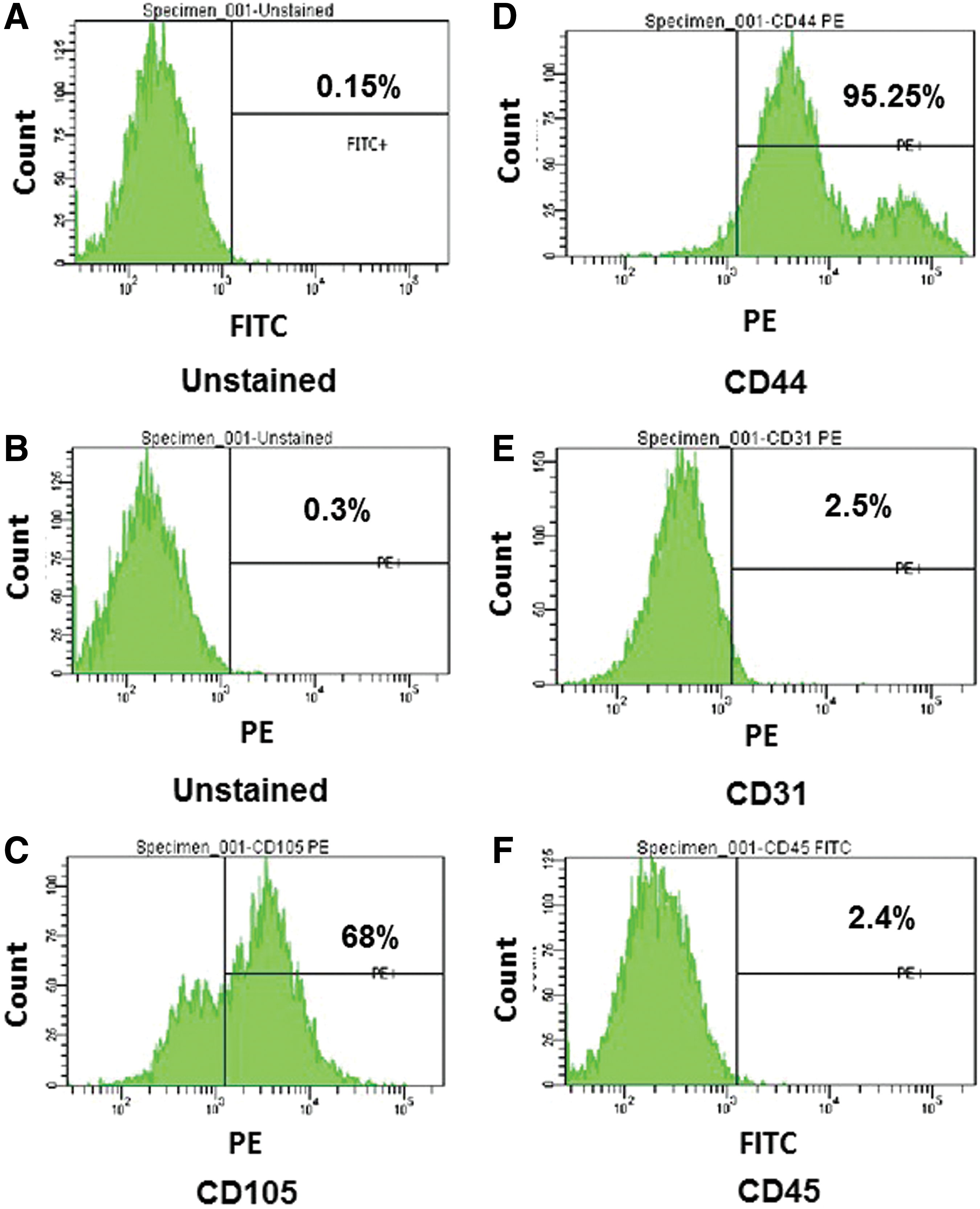
Characterization of adipose tissue–derived mesenchymal stem cells (ATMSCs). ATMSCs from C57BL/6 mouse at passage 4 were stained with phycoerythrin (PE)-conjugated CD105, CD44, and CD31, and fluorescein isothiocyanate (FITC)-conjugated CD45 antibodies. The cells were also stained with 7-aminoactinomycin D (7-AAD) in order to remove the dead cells population. (
In vitro transduction of ATMSCs with lenti-hAAT vector
This study sought to use ATMSCs as a platform for hAAT (SERPINA1) gene delivery. 17 ATMSCs were infected with a lentiviral vector expressing hAAT (LV-hAAT). The efficiency of transduction was evaluated 48 h after infection by detecting the hAAT transgene expression using hAAT-specific ELISA. As shown in Fig. 2A, infected cells (ATMSC-LV-hAAT) produced hAAT in the culture medium.
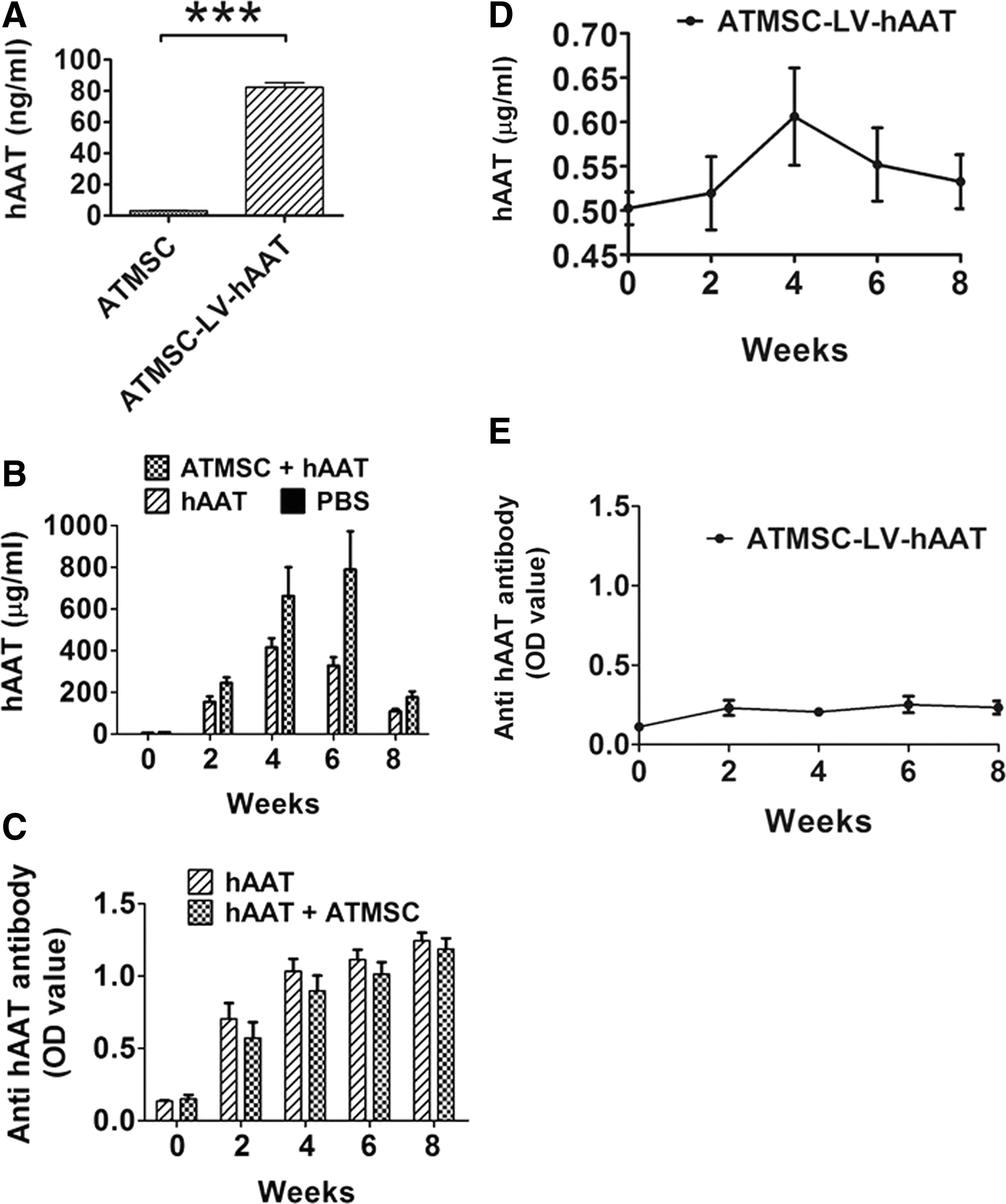
Serum levels of human AAT (hAAT) and anti-hAAT antibodies. (
ATMSC-LV-hAAT produced sustained levels of hAAT and a negligible amount of anti-hAAT antibodies
Cohorts of OVX mice were treated with PBS (200 μL every 3 days for 8 weeks), hAAT protein injection (Prolastin C®, 2 mg/mouse in 200 μL of PBS), ATMSCs transplantation (1 × 106 cells at day 4 and 0.8 × 106 cells at day 8 in 200 μL of saline solution), and ATMSC-LV-hAAT (0.6 × 106 cells in 200 μL of saline solution for one time only) transplantation. Sham-operated mice were used as controls. As expected, hAAT protein injection resulted in high levels of hAAT and anti-hAAT antibodies in the serum (Fig. 2B and C). Transplantation of ATMSC-LV-hAAT produced a detectable and sustained level of hAAT, but negligible levels of anti-hAAT antibodies (Fig. 2D and E).
ATMSC-LV-hAAT reduced inflammatory cytokines (IL-6 and IL-1β) and serum phosphate, and downregulated RANK (TNFSF11A)and RANKL (TNFSF11) gene expression
Previous studies suggested that the proinflammatory cytokines IL-6, IL-1β, and TNF-α are the primary mediators of postmenopausal osteoporosis by regulating RANK/RANKL/osteoprotegerin (OPG) pathways. 8,10,11 In order to evaluate the anti-inflammatory effect of hAAT treatments, serum levels of IL-6, IL-1β, and TNF-α were detected, and it was found that all treatments reduced the OVX-induced increase in IL-6 and IL-1β (Fig. 3A and B) but had no significant effect on TNF-α production (Fig. 3C). Eight weeks after the initiation of the treatments, all animals were sacrificed and subjected to molecular and pathological examinations. RANK (TNFSF11A), RANKL (TNFSF11), and integrin β3 (ITGB3) gene expression was detected in the bone tissue. As shown in Fig. 3D, hAAT and ATMSC combination therapies (ATMSC + hAAT and ATMSC-LV-hAAT) significantly decreased RANK (TNFSF11A) gene expression, while hAAT or ATMSCs alone did not. In addition, a decreasing trend of OVX-induced RANKL (TNFSF11) gene expression was observed in ATMSC + hAAT and ATMSC-LV-hAAT groups (Fig. 3E). The study also showed that hAAT alone and ATMSC + hAAT treatments significantly reduced integrin β3 (ITGB3) gene expression (Fig. 3F). Serum levels of calcium (Ca++), phosphate (PO4 3–), and ALP were detected. Compared with the sham-operated group, the PBS-injected OVX group had significantly lower levels of serum phosphate. ATMSC and combination treatments (ATMSC + hAAT and ATMSC-LV-hAAT) significantly increased serum phosphate level (Fig. 3G). No significant changes were found in serum calcium or ALP between PBS-injected OVX (osteoporosis disease model group) mice and the sham-operated groups (Fig. 3H and I).
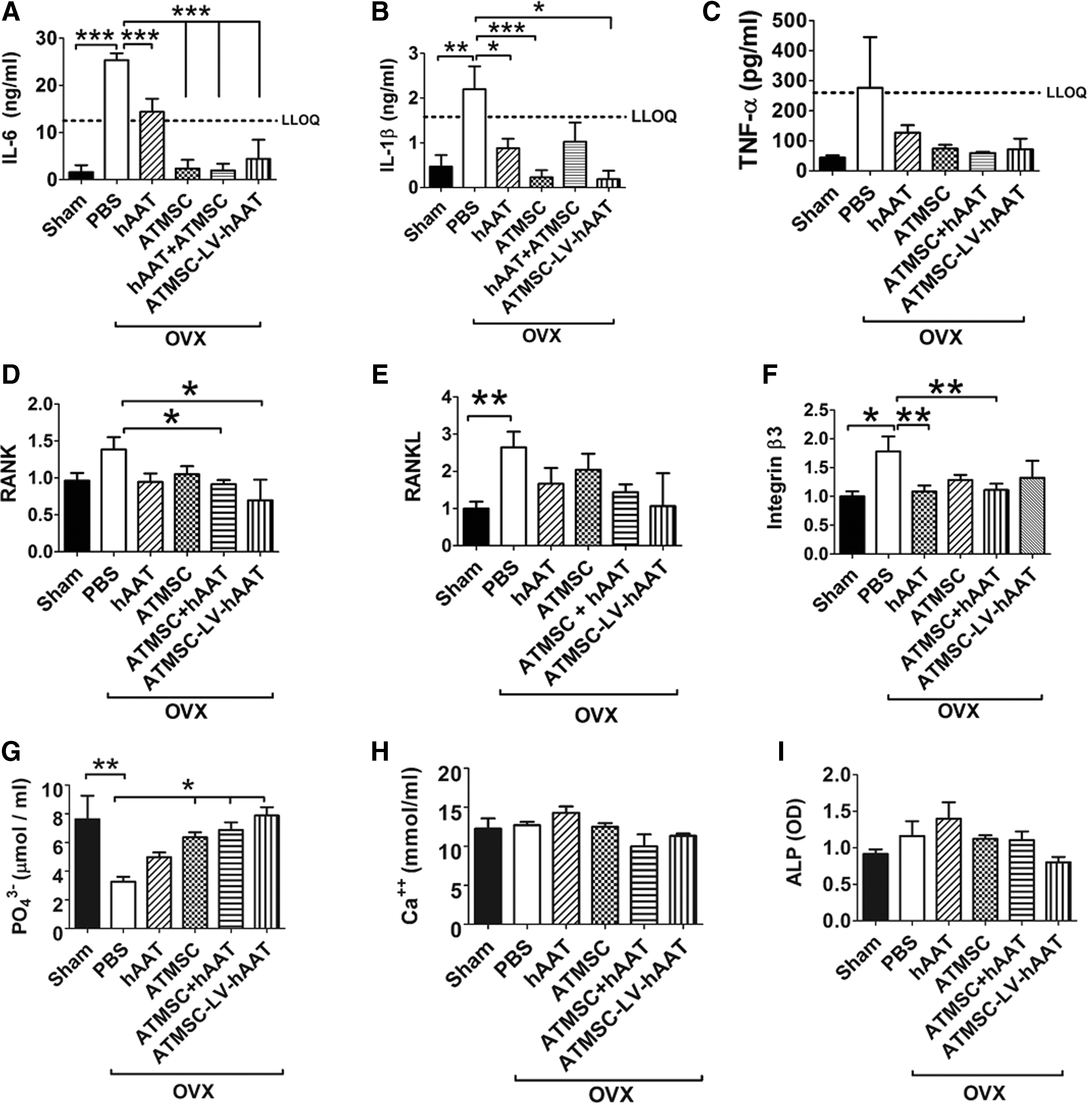
ATMSC-LV-hAAT reduced serum levels of proinflammatory cytokines (interleukin [IL]-6 and IL-1β) and phosphate, and downregulated bone tissue gene expression of RANK, RANKL, and integrin β3. Serum levels of (
ATMSC-LV-hAAT protected against OVX-induced bone loss
To evaluate the treatment effect of hAAT on bone microstructure, μCT scanning analysis was performed. As shown in Fig. 4, hAAT and ATMSCs combination therapies (ATMSC + hAAT or ATMSC-LV-hAAT) significantly improved bone microstructure parameters (BV/TV, BMD, SMI, and BV) compared with the PBS-injected OVX group.

Effect of ATMSC-based hAAT gene therapy on vertebral bone microstructure. (
ATMSCs-LV-hAAT migrated into bone marrow and expressed hAAT after transplantation
Previous studies have shown that ATMSCs migrated into bone marrow after liver-directed transplantation. To test if IP-injected ATMSC-LV-hAAT can migrate into bone marrow, immunostaining for hAAT (a marker for transplant cells) was performed. As shown in Fig. 5, hAAT-positive cells were detected in the bone marrow of the ATMSC-LV-hAAT transplanted group (Fig. 5C and G) but not in the ATMSC group, in which transplant cells had no hAAT gene and served as a negative control (Fig. 5B). As expected, some hAAT-positive cells were also detected within the liver, heart, pancreas, skeletal muscle, uterus, and mesentery (Fig. 5F and H–L) in the same group. Features of acute or chronic toxicity associated with transplanted ATMSC or ATMSC-LV-hAAT were not observed in any of the examined tissue sections (Fig. 6). Likewise, features of neoplasia were not observed in this 8-week study.
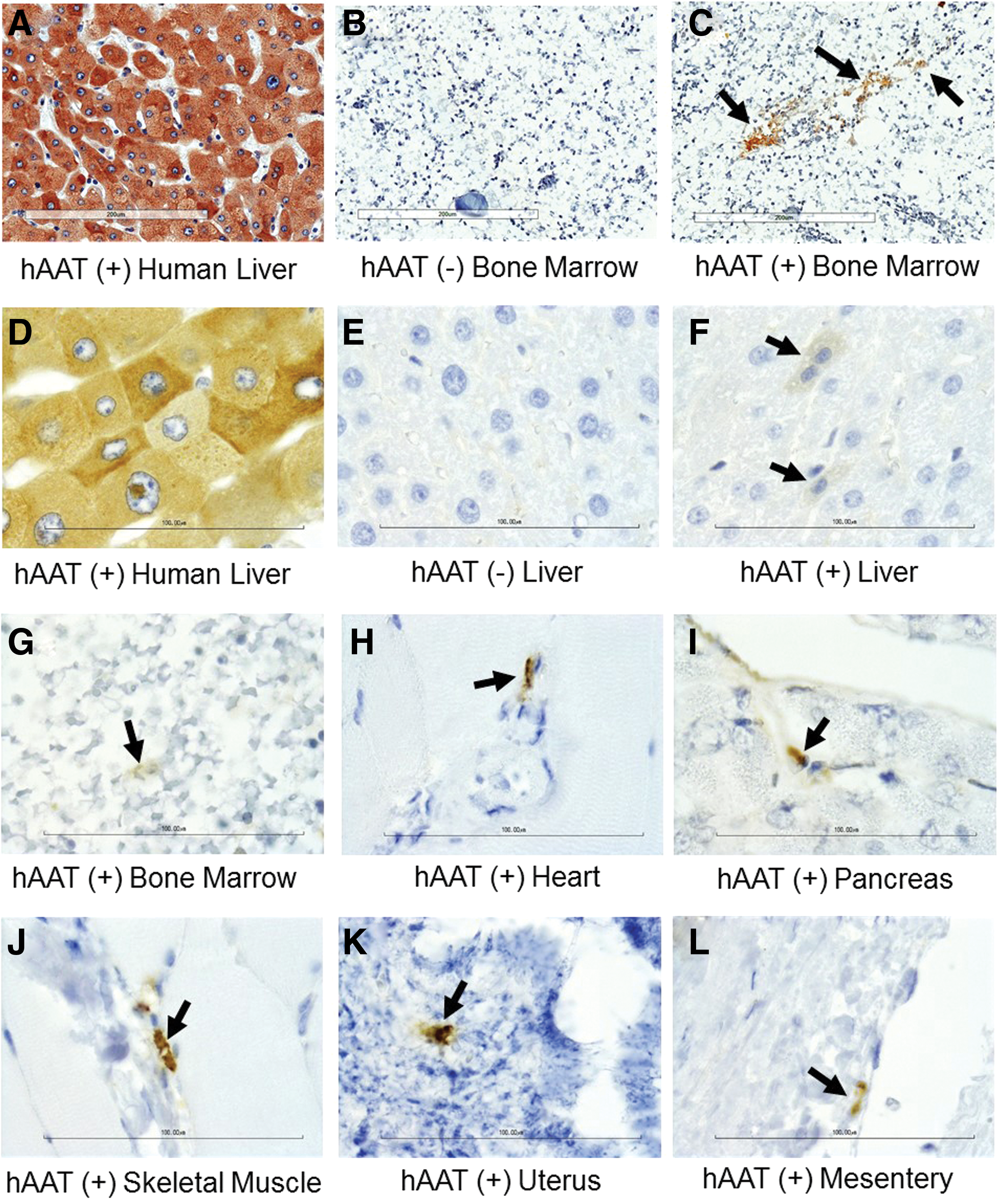
Transplantation of ATMSC-LV-hAAT-expressed hAAT in bone tissue and multiple organs. Tissue sections from ATMSC-LV-hAAT- and ATMSC-injected OVX group were subjected to immunostaining for hAAT at the same time. A human liver tissue section was used as a positive control. ATMSC (without lenti-hAAT infection)–injected mouse tissue sections were used as a negative control. (
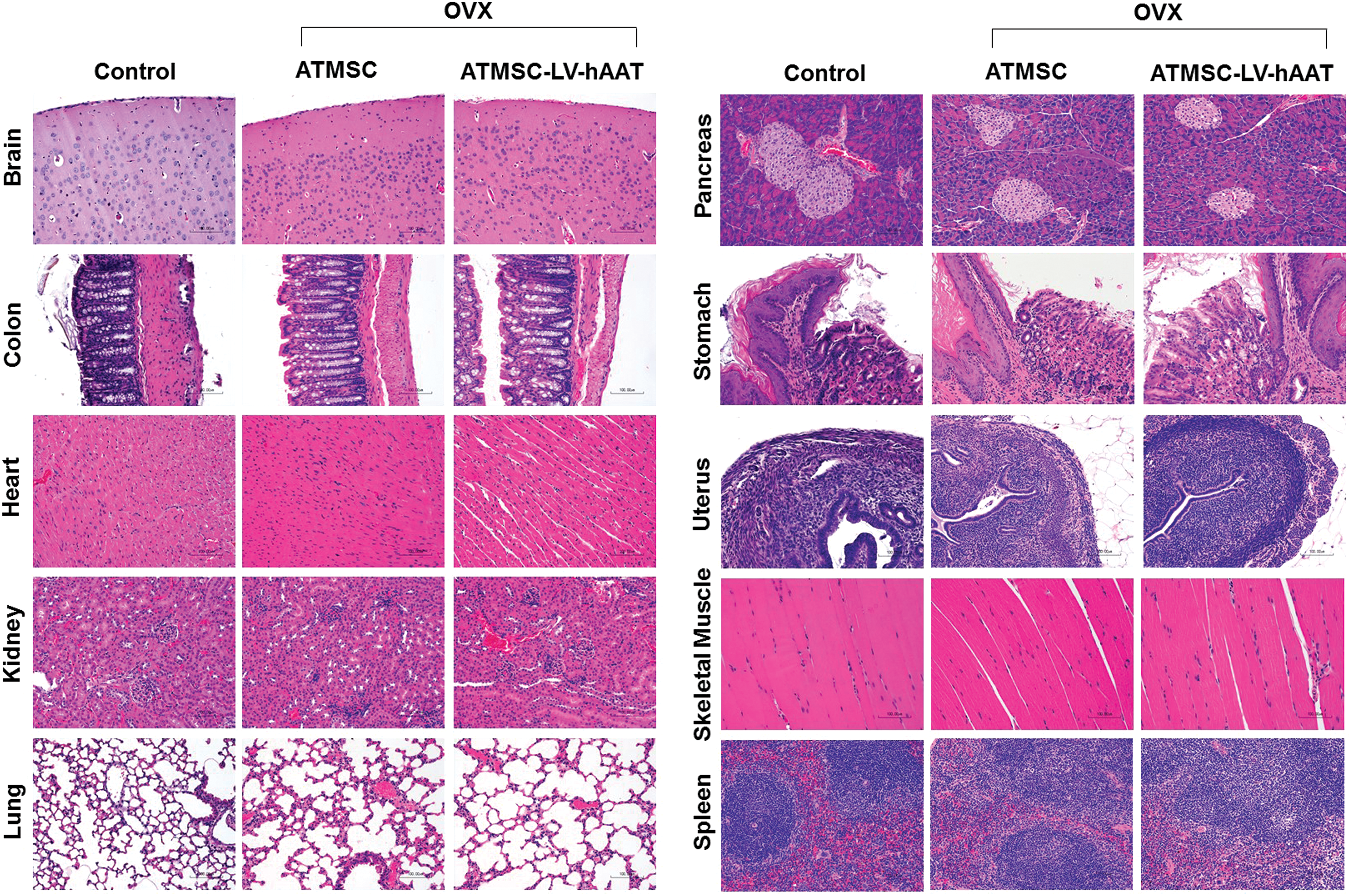
Histopathological examinations. Tissue sections from age-matched normal C57BL/6 mouse (control), and ATMSC- and ATMSC-LV-hAAT-injected OVX mice were subjected to hematoxylin and eosin staining. Color images available online at
Discussion
The present study has demonstrated that the combined administration of ATMSC and hAAT (ATMSC + hAAT), as well as the transplantation of genetically modified ATMSC with lentiviral vector–carrying hAAT (SERPINA1) gene (ATMSC-LV-hAAT), protected OVX-induced bone loss in a mouse model. This approach, combining the protective effects of hAAT and ATMSC, suggests a novel therapy for the treatment of osteoporosis in humans.
ATMSCs have several unique features for therapeutic applications. First, they can differentiate into multiple cell types and home to different tissues, including fat, bone, the liver, and the lung. 17 Transplantation of ATMSCs may enhance tissue or organ regeneration. In osteoporosis, ATMSCs can differentiate into osteoblasts and inhibit overactive osteoclast actions to maintain balanced bone remodeling. Second, ATMSCs have immune suppressive functions. They secrete anti-inflammatory factors and produce exosomes carrying anti-inflammatory molecules, all of which may contribute to their ability to protect against osteoporotic bone loss. 24 Third, ATMSCs express low (or no) levels of MHC-II molecules. Thus, they can escape the cellular immune response and establish long-term grafts. This feature makes ATMSCs a unique carrier or platform for therapeutic gene delivery into tissues or organs. 17 This study used ATMSCs with the expectation that all three features of ATMSCs would help to prevent bone loss in OVX mice.
Estrogen withdrawal increases the production of pro-inflammatory cytokines, which are associated with increased activity of osteoclasts and loss of bone mass. Studies supported that pro-inflammatory cytokines, such as IL-6, IL-1β, and TNF-α, are the primary mediators of the accelerated bone loss at menopause. 8 Human AAT is a multifunctional protein with antiproteinase, anti-inflammatory, and cytoprotective properties. 25 It is possible that its anti-inflammatory properties could play a critical role in preventing OVX-induced bone loss in mice.
Although the contributions of each feature of hAAT and ATMSC mentioned above to the protective outcome remain for future investigation, the protective effects of combination therapy (ATMSC + hAAT or ATMSC-LV-hAAT) observed may be due to both the inhibition of osteoclastogenesis and the promotion of osteogenic capacity, thus reducing the bone turnover rate. The present study showed that both ATMSC + hAAT and ATMSC-LV-hAAT treatments inhibited RANK (TNFSF11A) gene expression. These results are consistent with a previous observation 16 indicating inhibitory activity of hAAT on osteoclastogenesis. In addition, it was shown that ATMSC-LV-hAAT inhibited inflammatory cytokines IL-6 and IL-1β, which are the major mediators for osteoclastogenesis. Ovariectomy is known to alter phosphate homeostasis. It decreases serum phosphate levels and increases ALP activity, suggesting that OVX increases the bone turnover rate. 26 Consistent with previous observations, the present results showed that serum level of phosphate in PBS-injected OVX mice were significantly lower than that of the sham-operated group. 26,27 Importantly, ATMSC + hAAT and ATMSC-LV-hAAT treatment significantly increased the serum phosphate level compared with the PBS-injected OVX group. In addition, there was a trend that ATMSC-LV-hAAT administration decreased OVX-induced ALP activity compared with the PBS-injected OVX group. Although phosphate and ALP homeostasis are complex processes, the results indicate the ATMCS and hAAT therapy decreased the bone turnover induced by OVX. Ovariectomy may alter calcium homeostasis in the rat, 26 –29 and no significant changes were observed between groups in this study with OVX mice, consistent with previous observations. 30,31
Nonetheless, the advantage of combining ATMSCs and hAAT was clearly observed. Importantly, with a lower number of transplanted cells, two injections of ATMSC-LV-hAAT showed the most impressive protective effect among all the treatments tested. Several factors may contribute to the efficiency of the protection. ATMSC-LV-hAAT can migrate to bone tissue and function in two ways: (1) differentiate into osteoblasts, which can enhance bone remodeling, as discussed above; and (2) express hAAT, which can inhibit inflammatory cytokine IL-6 and IL-1β production, RANK (TNFSF11A) gene expression, and tissue damage. Indeed, our immunohistological data showed that ATMSC-LV-hAAT homed to the bone marrow and expressed transgene hAAT. As expected, some transplant cells were also observed to have migrated into other tissues and expressed hAAT, indicating that these cells may contribute to the systemic inhibition of inflammation and that this cell delivery method may be used for other tissue regeneration (e.g., the liver). It should be noted that features of neoplasia or toxicity associated with transplanted ATMSC or ATMSC-LV-hAAT were not observed in this study.
This study was performed together with an rAAV-mediated hAAT (SERPINA1) gene therapy study, which has been published recently. 16 Although both therapies showed a protective effect, no statistical difference was observed between these two strategies. It is difficult to conclude which one is better, since both strategies have advantages and disadvantages. rAAV8-mediated hAAT (SERPINA1) gene delivery can result in systemic infection (multiple organs and cell types) and lead to long-term and high serum levels of hAAT. Transplantation of ATMSCs expressing hAAT has the advantage of combined effects of hAAT and ATMSCs. However, ATMSC transplantation may be limited by the number of cells transplanted depending on the models used. Nevertheless, both strategies have therapeutic potential and confirmed that hAAT is protective. Compared with hAAT protein injection, which resulted in variable serum levels of hAAT, the local expression of hAAT from ATMSCs in the bone marrow may be more effective due to the sustained expression at the site needed and perhaps less exposure to clearance processes. Future studies optimizing cell numbers and transgene expression may further improve the protective effect.
Footnotes
Acknowledgments
This work was supported by a grant from the University of Florida. We thank Dr. Jay J. Cao and his group in USDA (ARS Grand Fork Human Nutrition Research Center, North Dakota, USA) for their assistance in bone μCT scanning.
Author Disclosure
No competing financial interests exist.