
Abstract
Adeno-associated virus (AAV) gene transfer is a promising treatment for genetic abnormalities. Optimal AAV vectors are showing success in clinical trials. Gene transfer to skeletal muscle and liver is being explored as a potential therapy for some conditions, that is, α1-antitrypsin (AAT) disorder and hemophilia B. Exploring approaches that enhance transduction of liver and skeletal muscle, using these vectors, is beneficial for gene therapy. Regulating hormones as an approach to improve AAV transduction is largely unexplored. In this study we tested whether insulin therapy improves liver and skeletal muscle gene transfer. In vitro studies demonstrated that the temporary coadministration (2, 8, and 24 hr) of insulin significantly improves AAV2-CMV-LacZ transduction of cultured liver cells and differentiated myofibers, but not of lung cells. In addition, there was a dose response related to this improved transduction. Interestingly, when insulin was not coadministered with the virus but given 24 hr afterward, there was no increase in the transgene product. Insulin receptor gene (INSR) expression levels were increased 5- to 13-fold in cultured liver cells and differentiated myofibers when compared with lung cells. Similar INSR gene expression profiles occurred in mouse tissues. Insulin therapy was performed in mice, using a subcutaneously implanted insulin pellet or a high-carbohydrate diet. Insulin treatment began just before intramuscular delivery of AAV1-CMV-schFIX or liver-directed delivery of AAV8-CMV-schFIX and continued for 28 days. Both insulin augmentation therapies improved skeletal muscle- and liver-directed gene transduction in mice as seen by a 3.0- to 4.5-fold increase in human factor IX (hFIX) levels. The improvement was observed even after the insulin therapy ended. Monitoring insulin showed that insulin levels increased during the brief period of rAAV delivery and during the entire insulin augmentation period (28 days). This study demonstrates that AAV transduction of liver or skeletal muscle can be improved by insulin therapy.
Introduction
A
Insulin signaling occurs through the insulin receptor (IR) and triggers downstream intracellular events that regulate glucose metabolism, fat metabolism, glucose uptake, and mitogenic responses. 25 –29 After insulin binding, the liganded insulin receptor is internalized into the cell through dynamin-dependent clathrin-mediated endocytosis. 30 –34 Previous studies have shown that this form of endocytosis is one of the mechanisms used by AAV2 and AAV5 to enter the cell. 35 –38
All cells demonstrate basal glucose uptake. 20 Yet, glucose uptake can be variable and, in some cases, regulated by insulin. Insulin-dependent glucose uptake regulates the rate of glucose consumption by skeletal muscle. Conversely, glucose uptake in lung epithelial cells is insulin independent and the rate is not altered by insulin levels. Insulin-dependent and insulin-independent glucose uptake occur through glucose transporters (GLUTs). 39 Ubiquitous GLUT1 is associated with the basal rate of glucose uptake in all tissue and is independent of insulin. 40 Glucose uptake is not restricted to GLUT1 but is accompanied by other GLUTs; for example, GLUT3 is associated with glucose uptake in neurons, and GLUT2 and GLUT5 are associated with polarized lung epithelial cells. 41,42 Insulin-dependent glucose uptake is facilitated by GLUT4. Insulin-dependent glucose uptake through GLUT4 occurs in cardiac muscle, skeletal muscle, and adipose tissue. 43 –49 Therefore, the rate of glucose uptake in these tissues depends on physiological conditions that impact insulin levels (i.e., exercise and nutritional states). It is also well established that skeletal muscle and liver are the primary tissues that store glucose, which is regulated by insulin. 50 –52 In addition, the role of insulin activity in liver functions also involves glucose metabolism, which is critical for balancing glucose levels. 21,53 To summarize, liver, skeletal muscle, cardiac muscle, and adipose are considered insulin-responsive tissues, but lung tissue is not.
Acute hyperinsulinemia occurs during the fed state. 54,55 Yet, prolonged hyperinsulinemia can occur and has been observed in patients with type II diabetes. 56 In addition, congenital hyperinsulinemia has been reported in children. 57 Furthermore, fasting and other stress states such as exercise result in both a decrease in insulin activity and insulin levels. 54,58 The primary hormone responsible for both antagonizing insulin activity and lowering insulin levels is cortisol, an adrenal cortical steroid (glucocorticoid). In fact, long-term treatment with glucocorticoid antiinflammatory drugs has been linked to the development of type II diabetes. In addition, sustained hypercortisolism can lead to Cushing's disease, which mimics type II diabetes. 59 –63
In the event that insulin activity influences AAV gene transfer, the variability of insulin and cortisol levels due to normal and abnormal physiological conditions and the primary role of insulin in glucose and fat metabolism could have a latent impact on the current AAV gene therapy strategies that involve skeletal muscle and liver tissue gene correction. Therefore, investigating the effects of insulin activity on AAV transduction is critical in determining whether there is an undiscovered factor that is influencing the outcomes of AAV gene therapy. In this study we explored a novel approach, independent of modifying the AAV vector, that improves AAV gene transfer to skeletal muscle and liver tissue by augmenting insulin levels through exogenous insulin supplementation or by means of a high-carbohydrate diet.
Materials and Methods
AAV vector production, purification, and titration
An AAV serotype 1 or 8 vector with a self-complementary human factor IX coding region and a cytomegalovirus promoter (AAV1-sc-hFIX or AAV8-sc-hFIX) was used in animal studies and an AAV serotype 2 vector with a β-galactosidase (LacZ) coding region and a cytomegalovirus promoter was used for in vitro studies (AAV2-CMV-LacZ). The cis pITR2-CMV-sc-hFIX plasmid was provided by R. Herzog (University of Florida, Gainesville, FL). The cis pITR2-CMV-LacZ plasmid was purchased from Cell Biolabs (San Diego, CA). The trans pDP helper plasmids (pDP1, pDP2, and pDP8) were provided by T.E. Golde (University of Florida, Gainesville, FL) and P. Cruz (University of Florida, Gainesville, FL). Virus was prepared as previously described, using HEK293 cells factors and calcium phosphate transfection with cis and trans plasmids. 64 A cleared lysate containing AAV vector was prepared by using citric acid buffer as the suspension buffer as previously described. 65 Empty capsids were separated by iodixanol separation as previously described. 64 After iodixanol separation, the purified vector with packaged vector genome was rinsed with 5 × phosphate-buffered saline (PBS) and concentrated to 2 ml by tangential flow filtration, using a 300-kDa MicroKros hollow fiber filter with a modified polyethersulfone membrane (Spectrum Labs, Rancho Dominguez, CA). The virus was titered by quantitative PCR with primers for the CMV region of the recombinant viral genome (forward, 5′-ACGCCAATAGGGACTTTCC-3′; reverse, 5′-TCAATAGGGGGCGTACTTG-3′), using SYBR green, and reported as vector genomes (VG) per milliliter. The Applied Biosystems StepOnePlus instrument (Thermo Fisher Scientific, Waltham, MA) was used. The titering was done in quadruplicate wells and with two different dilution factors (1:103 and 1:104). The titering was done twice for precision and accuracy.
Animal protocol for insulin augmentation
All animal studies performed were approved by the institutional animal care and use committee. Male and female C57BL/6J mice (Jackson Laboratory, Bar Harbor, ME) were used. Male mice at 22 g and female mice at 18 g were used with an age range of 12–14 weeks. First, mice were deprived of food for 1 hr. Mice were divided into three groups: 1, control; 2, an insulin pellet insulin augmentation protocol; or 3, a high-carbohydrate diet (HCD) insulin augmentation protocol. For the HCD arm, control mice received a normal diet. For the insulin pellet arm control mice received a blank (no insulin) palmitic acid microcrystal pellet implanted subcutaneously (LinShin Canada, Scarborough, Canada). Insulin pellet mice received a normal diet, and an insulin pellet that delivers about 0.1 U of insulin per day for about 28 days (LinShin Canada) was implanted subcutaneously. HCD mice received a high-carbohydrate diet (77.5% carbohydrate, 9.8% fats, 13.7% proteins; TestDiet, St. Louis, MO) for 28 days. Thirty minutes after the insulin augmentation protocol began, the experimental and control groups of mice were injected with virus. AAV1-sc-hFIX vector at 3.0 × 1010 VG was delivered by intramuscular injection. AAV8-sc-hFIX vector at 5.0 × 1010 VG was delivered to the liver by splenic capsule injection as previously described. 66 Mice were kept on the insulin augmentation protocol for 28 days. The mice were euthanized 2 weeks (+42 days) after the end of insulin therapy.
Whole blood collection and glucose monitoring
About 150 μl of whole blood was collected at various time points (time zero [T0], +30 min, +60 min, +120 min, +240 min, +24 hr, and weekly up to 6 weeks). Blood was collected via retro-orbital bleeding, using a heparinized capillary tube. Whole blood (10 μl) was used to measure glucose using a TRUEtrack glucometer (Thermo Fisher Scientific). Serum was separated by refrigerated centrifugation at 4°C for 10 min at 8000 × g from the remaining whole blood to measure insulin, cortisol, and human factor IX (hFIX) levels.
Measuring hFIX levels
Circulating hFIX was measured by sandwich ELISA as previously described. 67 This is a well-established ELISA that measures only hFIX levels (not endogenous murine factor IX). The wells of a 96-well plate were coated with monoclonal anti-human factor IX clone HIX-1 antibody (Sigma-Aldrich, St. Louis, MO) at 1:880 dilution overnight (18 hr) at 4°C. Wells were blocked with 5% nonfat milk for 1 hr at room temperature. The standard curve and serum samples were prepared in 5% nonfat milk and incubated for 4 hr at room temperature. The standard curve was prepared with recombinant hFIX protein (Pierce-Thermo Scientific, Waltham, MA) by 2-fold serial dilution from 200 to 1.56 ng/ml, and serum samples were prepared as 1:5 dilutions. A secondary goat anti-human factor IX affinity-purified horseradish peroxidase (HRP)-labeled antibody (Affinity Biologicals, Ancaster, Canada) was added at 1:1000 dilution in 2% bovine serum albumin (BSA) for 1 hr at room temperature. HRP developer (ortho-phenylenediamine [OPD] at 1.5 mg/ml in peroxide buffer) was added for 8 to 15 min at room temperature and HRP development was measured with a ELx800 plate reader (BioTek, Winooski, VT).
Measuring cortisol levels
A cortisol ELISA was performed as previously described, using a competitive ELISA method. 68 The wells of a 96-well plate were coated with monoclonal anti-cortisol antibody (EastCoast Bio, North Berwick, ME) at 6.2 μg/ml overnight (18 hr) at 4°C. Wells were blocked with 0.1% BSA in PBS for 1 hr at room temperature. The standard curve and serum samples were prepared in 0.1% BSA in PBS and incubated for 3 hr at room temperature. The standard curve was prepared using cortisol reference solution (Sigma-Aldrich) by 2-fold serial dilution from 40 to 0.31 ng/ml, and serum samples were prepared as 1:10 dilutions. Cortisol–HRP (EastCoast Bio) was prepared as a 1:80 dilution in 0.1% BSA in PBS and was added (at a ratio of 1:1) to the standard curve points and diluted serum samples. HRP developer (OPD at 1.5 mg/ml in peroxide buffer) was added for 6–10 min at room temperature and HRP development was measured with a BioTek ELx800 plate reader.
Measuring insulin levels
A direct ELISA was developed to measure insulin levels. Wells of a 96-well plate were coated with reference standard curve or serum samples at 1:10 dilution. Both were prepared in coating buffer (12 mM sodium carbonate, 88 mM sodium bicarbonate in water) and incubated overnight (18 hr) at 4°C. To prepare the standard curve, recombinant human insulin (Sigma-Aldrich) was aliquoted in a range of 100–1.56 pmol/liter using 1:2 serial dilutions. Wells were blocked with 6% BSA in T-PBS (0.05% Tween in PBS) for 1 hr at room temperature. A biotin-labeled anti-insulin/proinsulin antibody (GeneTex, Irvine, CA) was added at 100 ng/ml (1:20,000) in dilution buffer (2% BSA in T-PBS) for 3 hr at room temperature. A streptavidin–HRP secondary antibody (Pierce-Thermo Scientific) was added at 500 ng/ml (1:1000) in dilution buffer for 1 hr at room temperature. HRP developer (OPD at 1.5 mg/ml in peroxide buffer) was added for 5–8 min at room temperature, and HRP development was measured with a BioTek ELx800 plate reader.
Cell culture for vector administration and insulin treatment
HuH7 (human liver) (Human Science Research Resources Bank, Tokyo, Japan), HepG2 (human liver) (HB-8065; American Type Culture Collection [ATCC], Manassas, VA), Hep3B (human liver) (HB-8064; ATCC), HEK293 (human kidney) (CRL-1573; ATCC), A549 (human lung) (CCL-185; ATCC), undifferentiated human myoblasts (Thermo Fisher Scientific), and undifferentiated C2C12 mouse myoblasts (CRL-1772; ATCC) were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with glucose (4.5 g/liter), sodium pyruvate, 10% fetal bovine serum (FBS), and penicillin–streptomycin antibiotics at 37°C in 5% CO2. For vector administration/insulin treatment, cells were grown to 70% confluency in 24-well tissue culture plates in triplicate. An established myoblast differentiation protocol was used to generate differentiated myofibers. 69 Specifically, DMEM with 2% horse serum was added to human undifferentiated myoblasts or undifferentiated C2C12 mouse myoblasts at approximately 50% confluency for 7 days. For transduction studies, AAV2-CMV-LacZ was added at a multiplicity of infection (MOI) of 104 to the cells in Opti-MEM medium for 8 or 2 hr depending on the study. After AAV infection, liver cells and lung cells were grown in DMEM with 5% FBS to reduce the rate of cellular proliferation, and differentiated myofibers were changed back to DMEM with 2% horse serum. LacZ staining was performed 96 hr after AAV infection. Three different recombinant human insulin cell culture studies were performed (Sigma-Aldrich). First, human insulin at a range of doses (0.001, 0.005, 0.03, 0.2, 1.0, 5.0, and 25.0 μg/ml) was coadministered during the AAV vector infection period and continued for 24 hr. The second study used only the 5- and 25-μg/ml doses of human insulin coadministered during the AAV vector infection period and continued for 24 hr. In the third study, the period of AAV2-CMV-LacZ infection was only for 2 hr and insulin was given at different time frames (2, 8, 24, 96, and +24 hr after infection). For all studies cells were rinsed once with PBS after all treatments. These studies were repeated three times. Control groups for all studies were cells infected with AAV2-CMV-LacZ without insulin.
LacZ staining in cultured cells
A standard LacZ staining protocol was used.
66
Briefly, 96 hr after AAV vector administration, cells were fixed with 0.5% glutaraldehyde for 5 min. Cell were rinsed twice with PBS and LacZ staining solution was added (20 mM K3[Fe(CN)6], 20 mM K4[Fe(CN)6], 1 mM MgCl2, and 5-bromo-4-chloro-3-indolyl-β-
Comparative analysis of INSR gene expression
For mice, mRNA was isolated from 30 mg of biopsied C57BL/6J mouse tissue (n = 4) and stored in RNAlater (Qiagen, Valencia, CA) at −80°C until performing first-strand synthesis. First-strand synthesis was performed to produce cDNA as previously described. 70 For cultured cells mRNA was also isolated from cells that were grown to about 90% confluency in 6-well plates in triplicate and stored in RNAlater at −80°C until performing first-strand synthesis. SYBR green RT-PCR (real-time polymerase chain reaction) was performed in triplicate, using 500-ng samples of cDNA pools generated from mouse tissue or culture cells, and was used to comparatively assess insulin receptor gene (INSR) expression. The primer pair used for mouse INSR was as follows: forward, 5′-ATGGCTCCTGGACAGAACC-3′; reverse, 5′-TCCGGCTGCCTCTTTCTC-3′; and compared with β-actin using the following primer pair: forward, 5′-ATCTGGCACCACACCTTCT-3′; reverse, 5′-GGGGTGTTGAAGGTCTCAAA-3′. The primer pair used for human INSR was as follows: forward, 5′-GACGTCCCGTCAAATATTGC-3′; reverse, 5′-TCTGGCTGCCTCTTTCTC-3′; and was compared with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Qiagen). Fold change was calculated by the 2ΔΔC t quantification method. ΔC t was calculated by comparing the mINSR C t value with that of β-actin or the hINSR C t value with that of GAPDH. ΔΔC t was calculated using the ΔC t value from lung data (for both mouse tissue and cultured cells) as the reference to compare with the ΔC t from the other samples. The Applied Biosystems StepOnePlus instrument from Thermo Fisher Scientific was used. These studies were done twice and samples were not pooled, so that a standard deviation could be calculated.
Statistical analysis
To determine the optimal insulin dose in in vitro studies, two-way analysis of variance (ANOVA) was used. To determine whether there was significant variation over time in blood glucose levels within groups, one-way ANOVA (repeated measures) was used. For all other tests a two-tailed, paired Student t test was used.
Results
INSR expression is higher in skeletal muscle and liver
INSR transcription levels were measured in cultured cells and mouse tissues to determine the range of gene expression between different tissues and cell lines and to show that differentiating cultured myoblasts into myofibers increases INSR transcription. A comparative fold change analysis was performed with cultured human liver cells (HuH7, HepG2, and Hep3B), cultured human myoblasts that were differentiated into human myofibers, and the HEK293 human kidney cell line; a human lung cell, A549, served as the control. The results show that the A549 cell line had the lowest amount of INSR transcription (Fig. 1A). With the exception of undifferentiated human myoblasts, all cell lines showed a substantial increase in INSR gene expression levels, with Hep3B cells showing the highest (14-fold) compared with A549 cells, followed by differentiated human myofibers (10-fold). In addition, differentiated human myofibers showed higher levels of INSR mRNA when compared with undifferentiated human myoblasts. A mouse myoblast cell line (C2C12) was also assessed, and differentiated C2C12 cells showed a 10-fold increase over undifferentiated C2C12 cells (Supplementary Fig. S1; supplementary data are available online at

Comparative analysis of INSR gene expression. Shown are the RT-PCR results for INSR gene expression of
Optimal dose of insulin improves AAV transduction of cultured liver cells and myofibers
An assessment of the effects of insulin on AAV2-CMV-LacZ transduction of cultured cells was performed. AAV2-CMV-LacZ infection occurred for 8 hr, and insulin was coadministered with the virus but continued for 24 hr. Transduction was assessed by LacZ staining, which determines the levels of β-galactosidase protein encoded by the lacZ coding region packaged in AAV2 (Supplementary Fig. S2: representative images of LacZ staining). An insulin dose–response study was performed first to determine whether there was an optimal dose to improve gene transduction. Liver cell lines (HuH7 and Hep3B) and differentiated myofibers (differentiated human myofibers and differentiated mouse C2C12 cells) showed the best improvement in AAV2-CMV-LacZ gene transfer at the 5-μg/ml dose of insulin (Fig. 2). Other doses that showed improvement in transduction in most of the cells were 1 and 25 μg/ml. No dose of insulin was able to achieve a significant increase in transduction in A549 human lung cells. An interesting result revealed that the highest dose tested (25 μg/ml) was not the most effective and in one instance (C2C12 fibers) it did not result in a significant improvement in transduction. Additional cell lines (HepG2 human liver cells and HEK293 human kidney cells) were tested. The 5- and 25-μg/ml doses were optimal in HepG2 cells, but only the 5-μg/ml dose significantly improved gene transfer in HEK293 cells (data not shown). To rule out that the presence of protein, and not necessarily insulin, altered AAV transduction, BSA at 5 μg/ml was coadministered instead of insulin. No improvement in lacZ transgene product was observed (data not shown).
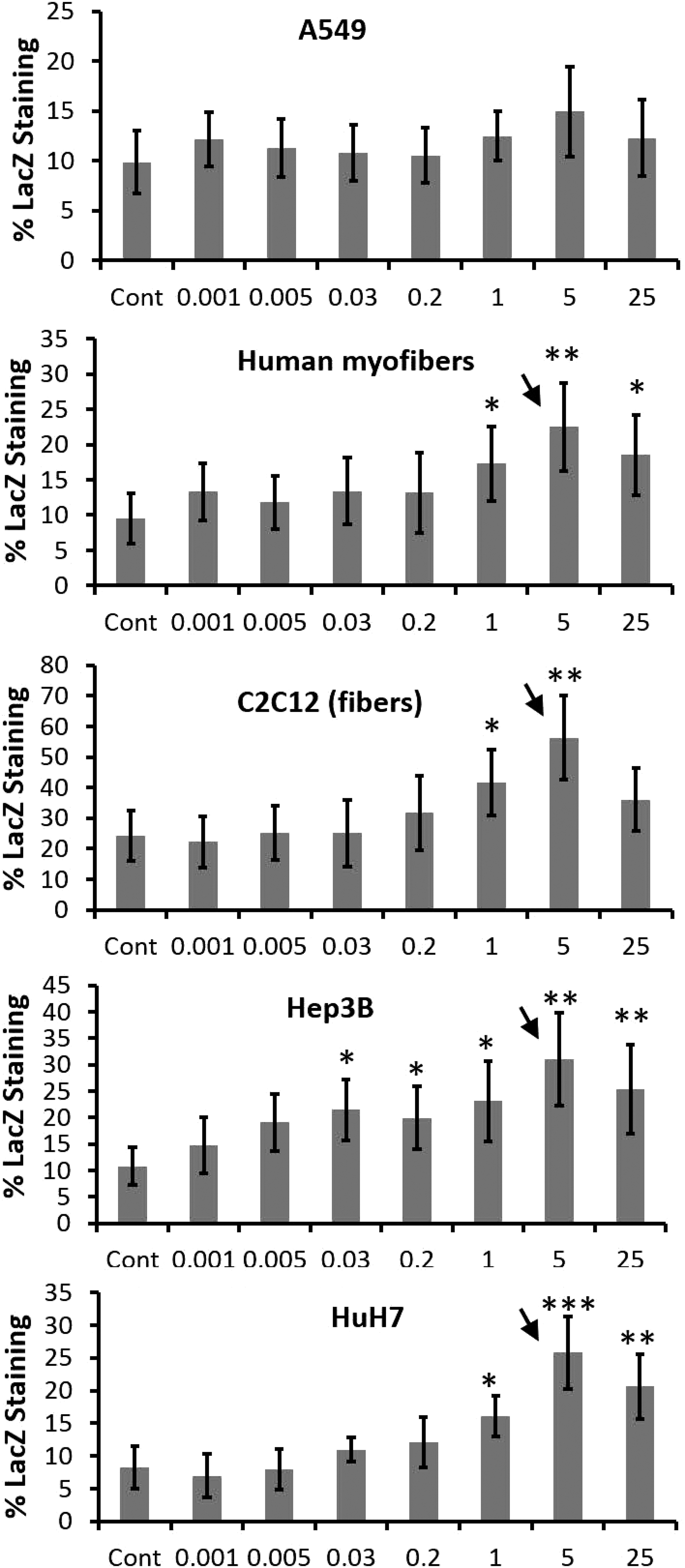
Insulin dose responses on AAV transduction. Insulin treatment for 24 hr at doses ranging from 0.001 to 25 μg/ml was used to determine whether there is an optimal dose to improve AAV transduction in cultured cells: human lung cells (A549), human liver cell lines (Hep3B and HuH7), human differentiated myofibers, and mouse differentiated myofibers (C2C12). Cells were infected with AAV2-CMV-LacZ at an MOI of 104 in triplicate for 8 hr and insulin was coadministered. Controls included virus infection for 8 hr with no insulin treatment. Transduction was determined by LacZ staining. *p < 0.05, **p < 0.01, and ***p < 0.001 (Student t-test). Arrows indicate the optimal dose, using two-way ANOVA. Data represent means ± SD.
Insulin treatment to improve lacZ gene transfer after AAV2-CMV-LacZ infection of cultured cells was repeated using only the most effective doses of insulin (5 and 25 μg/ml) (Supplementary Fig. S3). In the liver cell lines (HuH7, HepG2, and Hep3B) and differentiated myofibers (human and mouse C2C12) there was a 2.4- to 3.3-fold increase in LacZ staining at the 5-μg/ml dose, whereas the 25-μg/ml dose was effective in most but not all cell lines tested. In fact, in C2C12 differentiated myofibers there was a significant decrease in AAV transduction at the 25-μg/ml dose compared with the 5-μg/ml dose. Similar results occurred in HEK293 human kidney cells. The most effective doses of insulin were not able to improve AAV2-CMV-LacZ gene transfer to A549 human lung cells.
Insulin augmentation therapies improve AAV transduction of liver and skeletal muscle in mice
To determine whether increasing insulin will improve AAV transduction in murine liver and skeletal muscle two insulin augmentation therapies were used. Insulin was augmented exogenously by means of a subcutaneous insulin pellet that released insulin for up to 28 days or by giving the mice a high-carbohydrate diet (HCD) for 28 days. The insulin augmentation protocol began just before intramuscular administration of AAV1-CMV-schFIX or liver-directed delivery of AAV8-CMV-schFIX (via splenic capsule injection) and continued for 28 days (see Fig. 3A for an overview of the animal protocols). All mice were fasted for 1 hr before starting the insulin augmentation protocol in an effort to normalize insulin, glucose, and cortisol levels and to ensure that all mice ate once beginning the insulin augmentation protocol. Because of any potential sex-based variations in insulin, both male and female C57BL/6J mice were used. Female and male mice for all groups were studied once they reached the proper weight to ensure that severe hypoglycemia would not occur when animals were given the insulin pellet. Blood was collected during the brief interval of AAV administration (T0–240 min), long term during the insulin augmentation protocol (24 hr–day 28), and after the insulin therapy ended (day 42) to measure glucose, cortisol, and insulin levels. AAV transduction was assessed by specifically measuring the level of circulating human factor IX (hFIX) in blood. Both insulin augmentation protocols were able to improve AAV transduction to liver and skeletal muscle as seen by the 3.0- to 4.5-fold increase in hFIX levels (Fig. 3B–E). The hFIX levels remained significantly elevated after insulin augmentation therapy ended, indicating that the improved AAV transduction was stable and does not require sustained insulin therapy.
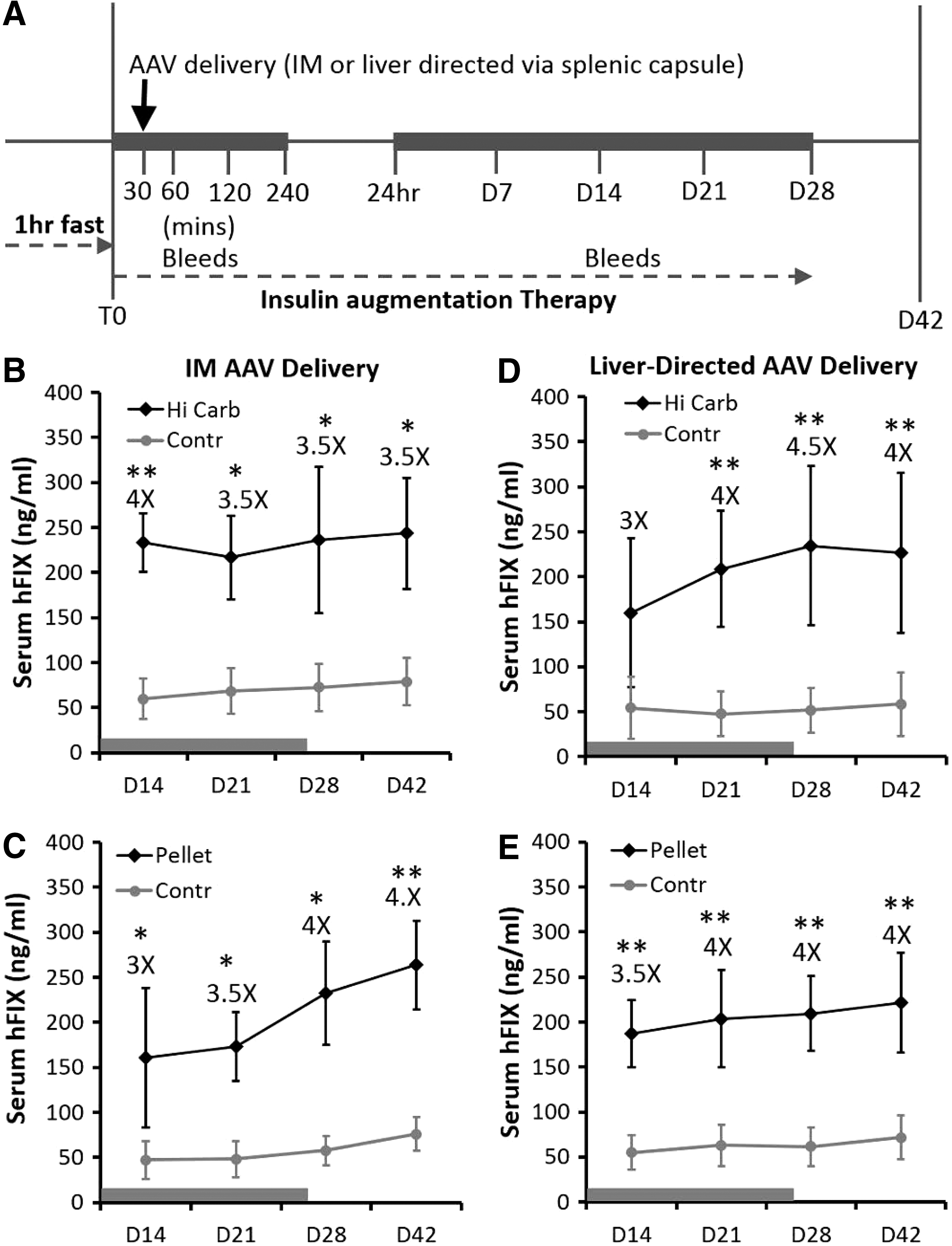
Insulin augmentation therapy to improve AAV transduction of skeletal muscle and liver in mice.
Insulin levels are elevated by augmentation therapies during and after the window of AAV administration
Insulin levels in blood samples were elevated during the window of AAV administration (Fig. 4A and C) and were sustained long term during the entire insulin therapy period (Fig. 4B and D). The data indicate that there was a peak in insulin levels just before (+30 min), during (+60 min), and just after (+120 min) AAV administration (Fig. 4A and C). The insulin levels began dropping within 24 hr (Fig. 4A and C) and stabilized during the entire insulin augmentation therapy, but the mice were still in a significant hyperinsulinemic state (day 7–day 28; Fig. 4B and D). After the end of the insulin treatment period the levels returned to normal (day 42; Fig. 4B and D).
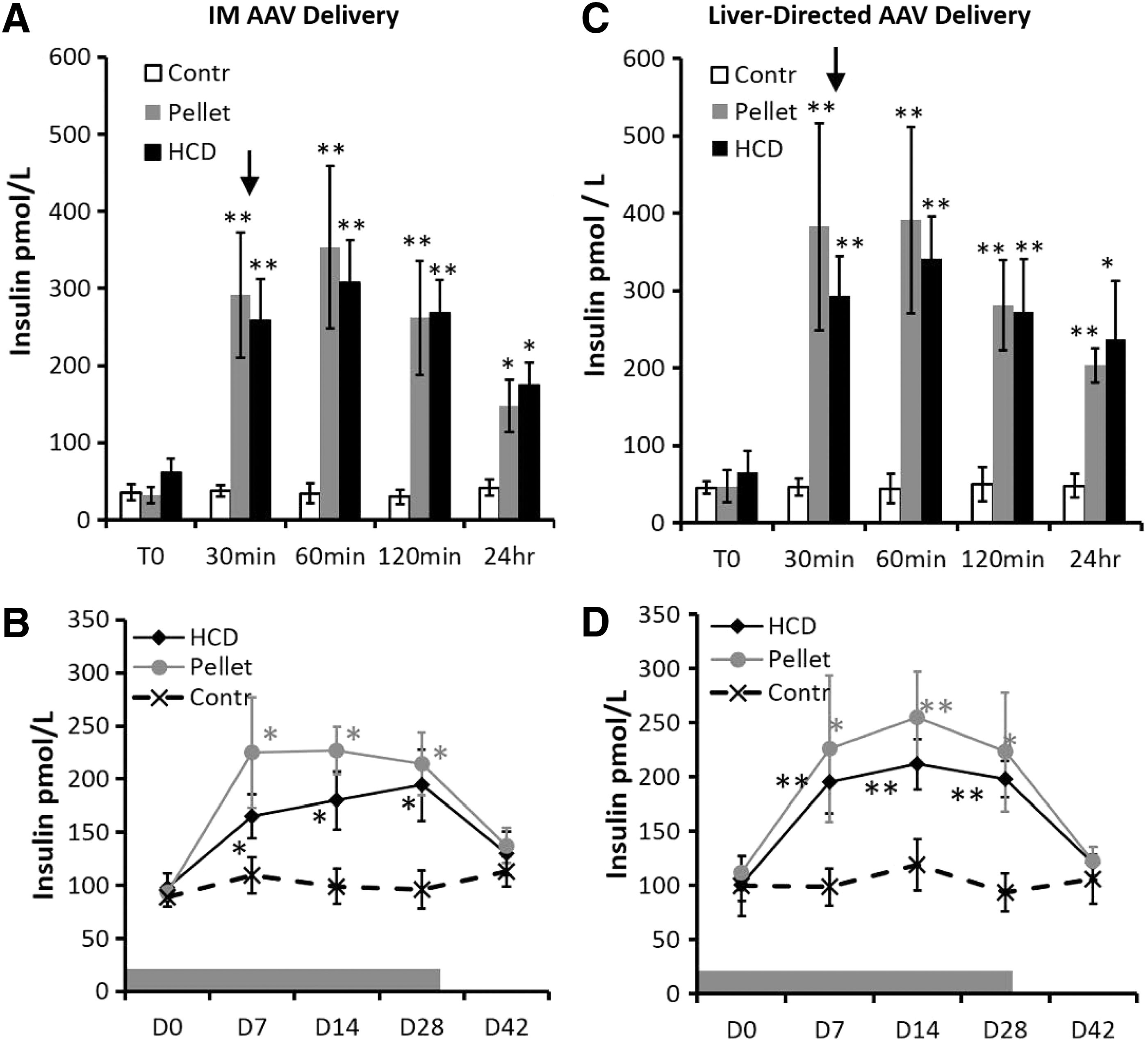
Insulin levels in mice during and after insulin augmentation therapy. Shown are insulin levels from two 28-day insulin augmentation therapies based on a subcutaneous insulin pellet or HCD during the short-term interval of AAV administration
Glucose levels demonstrate that insulin activity is elevated during AAV administration but is not sustained long term
Glucose uptake will result in a reduction in blood glucose, which is the normal response to elevated levels of insulin activity. Yet, an elevated insulin level does not always indicate an increase in insulin activity. Insulin resistance is a prime example of this situation. Measuring glucose during and after AAV administration provides evidence to support whether acute or long-term hyperinsulinemia results in insulin resistance. Glucose levels during the interval of AAV administration (Fig. 5A and C) show that blood glucose was significantly variable over time (T0–240 min) when using insulin augmentation therapy, indicating a short-term increase in insulin activity that paralleled the delivery of AAV and early cellular transduction events. This was not seen in the control mice. Long-term glucose levels stabilized and did not show any significant variations over time and were not elevated or decreased when compared with controls (Fig. 5B and D). This demonstrates that even though insulin levels were altered long term it did not result in a long-term increase in insulin activity. A repeated measures ANOVA (one-way ANOVA within individual groups but not between groups) was used to determine whether glucose levels were significantly altered over time within the experimental and control groups in Fig. 5A–D (during AAV administration). A t-test was performed on data presented in Fig. 5B and D (long term) to determine whether there were any differences in glucose levels between the control and treated mice.
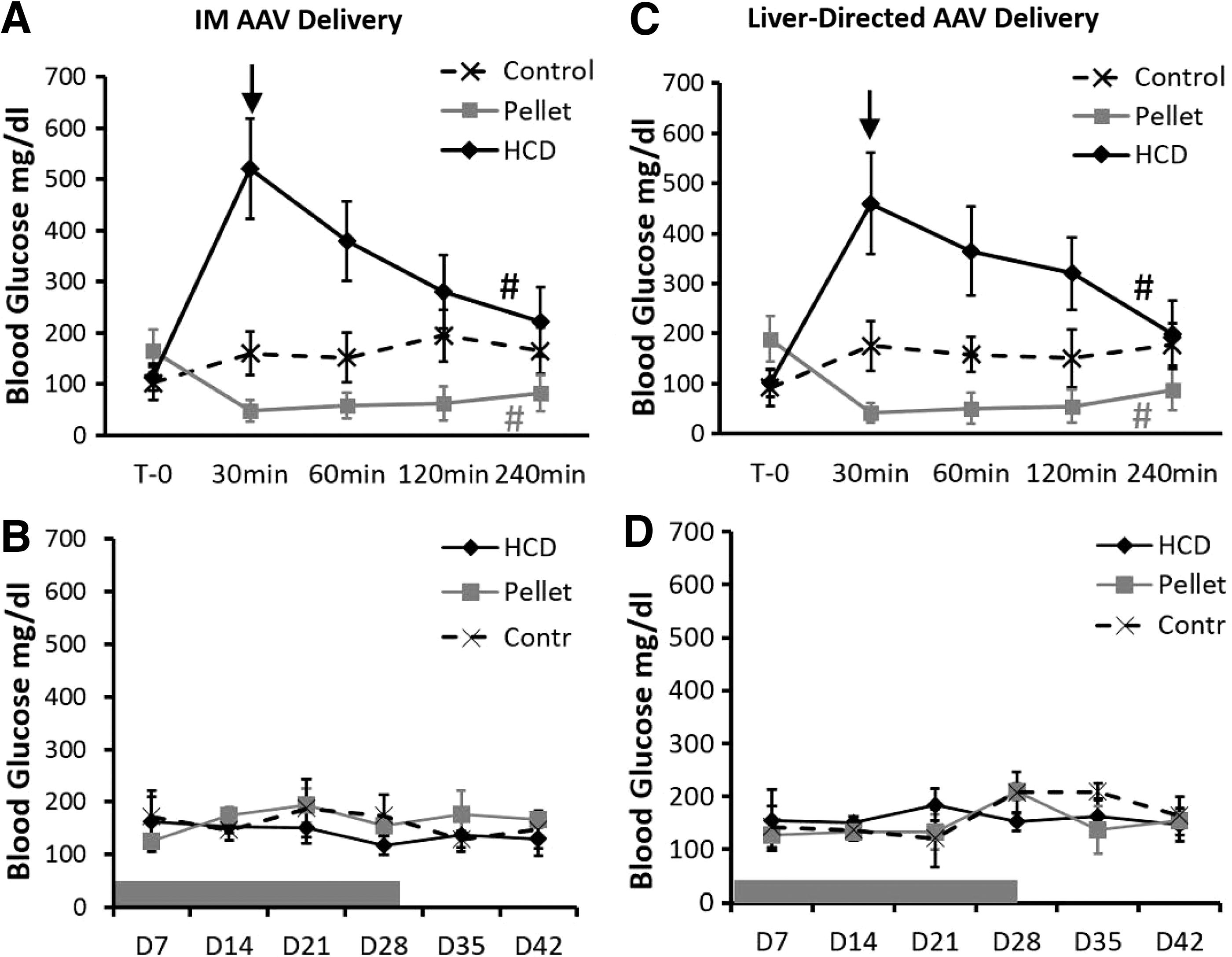
Blood glucose levels in mice during and after insulin augmentation therapy. Shown are blood glucose levels resulting from two 28-day insulin augmentation therapies using a subcutaneous insulin pellet or HCD during the short-term interval of AAV administration
Cortisol levels are elevated after, but not during, the interval of AAV administration
Cortisol experiments were performed to determine whether there is a physiological response to hyperinsulinemia, due to the insulin augmentation therapies, that could blunt the effects of insulin therapy on AAV cellular transduction. Assessing cortisol levels during and after AAV administration was important in determining whether physiological events that could limit insulin activity emerged during the early or late phase of cellular transduction or both. Cortisol levels in blood samples were assessed during the acute phase of AAV administration (Supplementary Fig. S4A and C) and long term (Supplementary Fig. S4B and D). The results show that cortisol levels were not elevated during the early phase of cellular transduction immediately after AAV administration (2 hr after the start of insulin augmentation and 1.5 hr after AAV administration). The levels of cortisol did increase within 24 hr after the insulin augmentation protocol began, and the levels remained elevated throughout the insulin augmentation protocol. Cortisol levels returned to normal after the insulin therapy ended.
Acute insulin treatment is sufficient to improve AAV transduction in cultured cells
This study was performed to align insulin treatment with AAV cellular transduction events, that is, uptake, trafficking, and gene expression. For these studies cultured cells were infected with AAV2-CMV-LacZ for only 2 hr. For most studies insulin was coadministered with AAV2-CMV-LacZ but was extended beyond the infection time. The time periods for the various insulin coadministration treatments were 2, 8, 24, and 96 hr. A post-AAV infection insulin treatment was also given 24 hr after AAV2-CMV-LacZ infection. These data show that insulin given for 2 hr significantly improved AAV2-CMV-LacZ transduction of HuH7 cells and C2C12 differentiated myofibers and that 8-, 24-, and 96-hr treatments showed similar improvement (Fig. 6A and B). Interestingly, the post-AAV infection insulin treatment given 24 hr after infection with virus did not increase LacZ staining.
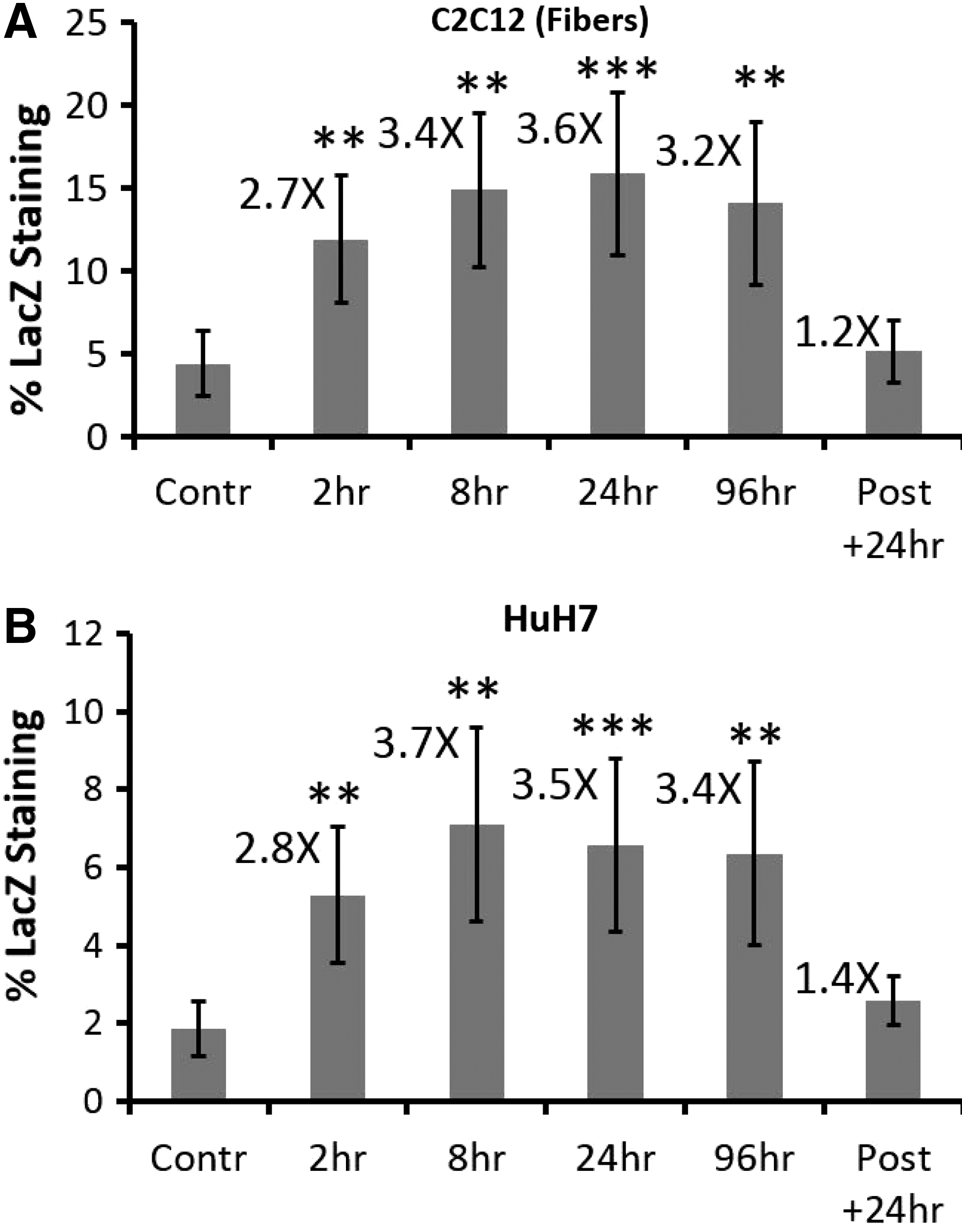
An acute 2-hr insulin treatment is sufficient to improve AAV transduction in cell culture.
Discussion
The primary outcome of this study is that insulin augmentation therapy improves AAV transduction of liver and skeletal muscle but not that of cultured lung epithelial cells. Mouse studies show that insulin augmentation protocols using exogenous insulin supplementation (via insulin pellet) or increasing endogenous insulin levels (via a high-carbohydrate diet) improve AAV gene transfer in vivo to skeletal muscle, using intramuscular injection of AAV1-CMV-schFIX or liver-directed delivery of AAV8-CMV-schFIX. In addition, the mouse studies show that this improvement in transduction was sustained even after insulin therapy ended and that insulin levels returned to normal. Finally, the results demonstrate that an acute increase in insulin activity (2-hr treatment) improves AAV2-CMV-LacZ transduction of cultured liver cells and differentiated muscle fibers.
Assessment of the cell culture data shows that cultured liver cells, myofibers, and kidney cells have increased INSR gene expression levels; similar gene expression profiles were observed in mouse tissue biopsy samples. The cell lines that showed elevated levels of INSR had improved AAV transduction when treated with insulin for 24 hr. Improvement in most cell lines was seen with multiple insulin doses, with 5 μg/ml showing improvement in transduction in all cell lines expect A549 lung cells. When analyzing the liver cell lines, 5 and 25 μg/ml were optimal but Hep3B cells showed significant improvement in transduction in response to 0.03- to 25-μg/ml doses. For differentiated myofibers 5 μg/ml was the preferred dose. Interestingly, in HEK293 cells and differentiated C2C12 myofibers there was a significant decrease in AAV transduction from the 5-μg/ml dose to the 25-μg/ml dose. Concerning HEK293 cells, it is possible that these kidney cells are more efficient at clearing insulin via cellular uptake of the ligand insulin receptor because a function of the kidney is rapid removal of insulin from the circulation to ensure proper blood glucose balance. 71 The high levels of insulin at the 25-μg/ml dose could reflect the depletion of insulin receptors after binding, which would blunt the response to insulin while the insulin receptors are replenished at the surface of the cells. The same effect could be occurring in the differentiated C2C12 myofibers.
To determine whether acute insulin treatment would improve transduction, cultured liver cells and differentiated C2C12 myofibers were infected for 2 hr with AAV2-CMV-LacZ and insulin was coadministered only during that 2-hr period. This brief 2-hr coadministration of insulin resulted in a significant increase in AAV2-CMV-LacZ transduction measured 96 hr after infection, and the result of the 2-hr insulin treatment was similar to those of the 8-, 24-, and 96-hr insulin treatments. To understand the significance of the acute insulin augmentation therapy in improving AAV transduction and that the enhanced levels of transgene product were sustained in mice even after the insulin therapy is ended, it must be placed in the context of basic cellular transduction events after AAV administration. Complete AAV uptake typically occurs within the first 2 hr of cellular transduction, followed by nuclear trafficking, which usually occurs within 8 hr of AAV infection. 33,72 After nuclear trafficking, second-strand synthesis occurs and finally sustained cellular transduction is due to stable gene expression of the transgene product. 73 Although acute insulin augmentation therapy could potentially influence AAV uptake and intracellular trafficking, and improve second-strand synthesis, it is not feasible to suggest that it will impact long-term transgene expression. Moreover, because AAV cellular transduction continued to be improved in mice even after the insulin augmentation protocol was ended, this further suggests that insulin augmentation does not have an effect on transgene expression but likely influences early events such as AAV uptake, intracellular trafficking, or both. More evidence is provided from this study to support the notion that insulin augmentation does not impact gene expression. A post-AAV infection insulin treatment was performed in cultured liver cells and differentiated myofibers, to which insulin was given 24 hr after a 2-hr AAV-CMV-LacZ infection, and showed no increase in the lacZ transgene product.
The data from this study do not identify a definitive mechanism. Yet, given that acute insulin treatment (2 hr) improved AAV transduction in cultured cells and that the results were similar to 8-, 24-, and 96-hr insulin treatments, this suggests that early transduction events (uptake and intracellular trafficking) are impacted. As previously mentioned, dynamin-dependent clathrin-mediated endocytosis is used for uptake of the liganded insulin receptor (IR), and it has previously been demonstrated that this is one of the mechanisms used by AAV2 and AAV5 to enter the cell. 35 –38 Therefore enhanced AAV uptake through the liganded IR endocytosis response is possible. In addition, IR signaling activates the phosphatidylinositol-3-kinase (PI3K) pathway. 27 Previous research has demonstrated that PI3K activity facilitates AAV trafficking to the nucleus. 74 This provides another potential link between increased insulin activity and improved AAV transduction. Phosphorylation of tyrosine residues on the AAV capsid limits AAV transduction because of subsequent ubiquitination. 75 This capsid tyrosine phosphorylation occurs through the epidermal growth factor receptor (EGFR), which is a receptor tyrosine kinase. Because the IR is also a receptor tyrosine kinase, activation of the IR could mimic EGFR but there would be a decrease in AAV transduction, not an increase. Therefore enhanced capsid tyrosine phosphorylation is unlikely. There currently is no evidence to suggest that insulin signaling through the IR influences the single-stranded D-sequence-binding protein (ssD-BP) or FK506-binding protein (FKBP52), both of which are involved in stabilizing the AAV genome by regulating second-strand synthesis of the single-stranded AAV genome. 76,77 A previous publication has shown that glucocorticoid receptor (GR) signaling impacts the state of serine/threonine phosphorylation of FKBP52 via protein phosphatase 5 (PP5) activity. 78 Yet, the data collected from this study do not support that GR signaling due to increased cortisol levels caused by prolonged hyperinsulinemia is involved. First, the use of a self-complementary hFIX coding region in this study would suggest that improvement in second-strand synthesis is not a factor. In addition, AAV transduction events were improved by insulin treatment in cultured cells and cortisol was not given.
From the data provided in this study, it is evident that using an insulin treatment to improve AAV transduction is influenced by elevated IR levels, yet other intracellular signaling factors are also likely involved. There was a range of INSR gene expression levels amongst the liver and myofiber cell lines as well as the liver and skeletal muscle mouse tissues but, in comparison, the improvement observed in AAV transduction due to insulin treatment did not vary significantly. The activity of rab5 and dynamin GTPase signaling molecules could account for this observation. It has been previously demonstrated that rab5 and dynamin are required for uptake of the liganded IR. 79 –81 In addition, rab5 plays a role in recycling the IR after uptake and is also involved in insulin signaling activity via the PI3K pathway. 79,81,82 In skeletal muscle, rab5 regulates the levels of GLUT4 but in liver it regulates glucose metabolism and glycogen production. 83,84 Interestingly, overexpression of rab5 in skeletal muscle can result in insulin resistance and a decrease in translocated GLUT4 molecules whereas rab5 expression positively regulates glycogen storage in liver. 83,84 Furthermore, rab5 expression has been shown to be dysregulated in hepatocarcinomas. 85 Therefore, even though there is almost a 3-fold increase in INSR expression between skeletal muscle and liver mouse tissue biopsy samples there is a potential for different expression patterns of rab5 and possibly dynamin between these tissues, which could be limiting the amount of improvement in AAV transduction in skeletal muscle. This same rationale could apply to the cultured cells, particularly the liver cell line, because they are derived from hepatocarcinomas.
Overall, this study clearly demonstrates that insulin therapy improves AAV transduction of skeletal muscle and liver tissue and that elevating endogenous insulin levels by diet is just as effective as exogenous insulin supplementation. The use of a long-term insulin augmentation protocol is not optimal given that the mice develop early signs of insulin resistance during prolonged hyperinsulinemia. This was indicated by the long-term increase (but not short term during the interval of AAV administration) in cortisol 24 hr after AAV administration. Also, insulin levels remained increased 24 hr after the insulin therapy was started but glucose levels remained normal and stable. However, the use of an acute insulin augmentation therapy is sufficient to achieve a long-term improvement in AAV transduction in liver cells and myofibers, and short-term hyperinsulinemia does not result in insulin resistance. Furthermore, the increase in AAV transduction by insulin augmentation is not limited to a single serotype (three serotypes were used in this study). Finally, the data show that an acute 2-hr insulin treatment has a similar improvement in gene transduction compared with 8-, 24-, and 96-hr treatments and that a post-AAV infection insulin treatment has no effect. This suggests that acute insulin therapy to improve AAV transduction likely involves early transduction events. Further studies will be required to pinpoint which early cellular transduction event(s) is/are involved. Other follow-up studies using acute insulin augmentation therapy in mice during the brief interval of administration will provide confirmation that a temporary spike in insulin is sufficient to improve AAV transduction to skeletal muscle and liver.
Footnotes
Acknowledgments
Many individuals provided valuable resources for this study. The authors thank Roland Herzog (University of Florida), who supplied the cis pITR2-CMV-sc-hFIX plasmid; Todd E. Golde (University of Florida), who provided the trans pDP helper plasmids through communications with Pedro Cruz (University of Florida); Lin Mantell (St. John's University), who supplied the A549 cell line; Woon-Kai Low (St. John's University), who provided the HepG2 cell line; and Xinggou Cheng (St. John's University), who supplied the HuH7 and Hep3B cells lines. This project was funded by St. John's University (Queens, NY).
Author Disclosure
None of the authors has anything to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.