
Abstract
Retinal degenerative diseases such as age-related macular degeneration, retinitis pigmentosa, and glaucoma result in permanent loss of retinal neurons and vision. Stem cell therapy could be a novel treatment strategy to restore visual function. In an ideal situation, a homogenous population of stem cell–derived retinal neurons with high purity is used for replacement therapy. Thus, it is crucial to elucidate the molecular mechanisms that regulate the development of retinal progenitor cells and subsequent generation of specific retinal neurons. Here, recent findings concerning the intrinsic and extrinsic factors that regulate retinal progenitor cell maintenance and differentiation are summarized, especially transcriptional factors and extrinsic signals. Understanding these mechanisms is indispensable because they have potential clinical applications, chiefly the generation of specific retinal cells such as retinal ganglion cells to treat glaucoma and other optic neuropathy diseases.
Introduction
A
AMD is the first leading cause of blindness in the elderly population with hallmarks of photoreceptors and retinal pigment epithelium (RPE) degeneration. Relatively speaking, testing the concept of stem cell–based therapy for RPE or photoreceptors is feasible, as these cells only need to form connections to neighboring host cells. In fact, stem cell–derived RPE has been shown to improve visual function and preserve neuronal connectivity in rodent models of retinal degeneration. 2 –9 Recently, Schwartz et al. reported that subretinal transplantation of human embryonic stem cells (ESC)–derived RPE resulted in improved visual acuity in a patient with Stargardt macular dystrophy and with dry AMD. 10
Glaucoma is a neurodegenerative disease characterized by the progressive and irreversible loss of retinal ganglion cells (RGC) and their axons. Elevated intraocular pressure (IOP) is a hallmark of glaucoma, especially in primary open angle glaucoma. Current treatment aims to lower the IOP level with pharmacological or surgical approaches. 11 However, some glaucoma patients still undergo RGC and vision loss despite a controlled IOP level. 12 Within this context, transplantation of stem cell–derived RGC is a novel strategy. 13 RGC replacement therapy is more challenging than either replacing photoreceptors or RPE. Successful therapy requires at least four sequential steps: (1) generation of homogenous stem cell–derived RGC; (2) integration into host retina and forming connections to retinal cells in the host retina; (3) robust axonal growth into the optic nerve; and (4) formation of synapses to visual targets.
Stem cells or induced pluripotent stem cells (iPSC) from various sources have all been evaluated for their viability. Both ESC and iPSC are pluripotent and can generate cell types derived from the three embryonic germ layers. In contrast, progenitor cells, fetal and adult stem cells, have limited self-renewal capacity and can only generate specific types of differentiated cells. For example, retinal progenitor cells (RPC) have the potential to differentiate into the six retinal neuronal neurons (rods, cones, bipolar cells, horizontal cells, amacrine cells, and retinal ganglion cells), and glial cells in the retina. 14 –16
Elucidation of the underlying mechanisms of differentiating RPC into RGC is crucial for designing protocols to treat optic neuropathy diseases such as glaucoma. The intrinsic and extrinsic signals related to RPC fate have been extensively reviewed. 14,16,17 This review focuses on the transcription factors and extrinsic signaling molecules that regulate RPC differentiation, especially the generation of RGC. In addition, the current findings of stem cell–based treatment will be discussed.
Factors in RPC Differentiation and RGC Generation
Retinogenesis is a spatially regulated process, beginning in the central retina and gradually progressing to the periphery. 18,19 Such temporal and spatial events require RPC proliferation and differentiation into various cell types at appropriate time points. Both intrinsic and extrinsic signals play indispensable roles in controlling these processes. 14,17 RGC is one of the “first-born” retinal neurons during retinogenesis. It is closely followed by the generation of cones, horizontal cells, and amacrine cells. After that, “late-born” retinal neurons, including bipolar cells, most rods, and Müller cells, will generate. 16,20 –22 Note that there is a considerable overlap of time in the differentiation of retinal cell types during retinogenesis. 20 –22 Intrinsic and extrinsic factors regulating retinogenesis are summarized in Table 1.
Intrinsic and extrinsic factors regulating retinogenesis
ASCL1, Achaete-scute family Homolog 1; ATOH7, Atonal BHLH Transcription Factor 7; BARHL2, BarH Like Homeobox 2; BHLH Transcription Factor 7; FGF, Fibroblast Growth Factor; FOXN4, Forkhead Box N4; NGN2, Neurogenin 2; NR2E3, Nuclear Receptor Subfamily 2, Group E, Member 3; NRL, Neural Retina Leucine Zipper; OLIG2, Oligodendrocyte Lineage Transcription Factor 2; OTX2, Orthodenticle Homeobox 2; PAX6, Paired Box Protein; POU4F2, POU Class 4 Homeobox 2; PRDM1, PR Domain Zinc Finger Protein 1; RAX, Retina And Anterior Neural Fold Homeobox 2; RPC, retinal progenitor cells; SHH, Sonic Hedgehog; SIX6, SIX Homeobox 6; SOX2, (sex determining region Y)-box 2; SUFU, Suppressor of Fused Homolog; TRβ2, Thyroid Hormone Receptor beta2; VEGF, Vascular Endothelial Growth Factor.
Intrinsic factors
The ability of RPC to differentiate into various retinal cell types is fundamentally an intrinsic property. 14,23 –25 The microenvironment appears to have little effect on the RPC differentiation because progenitors do not change their developmental course with environmental changes. 23,25,26 Transcription factors are the key intrinsic regulators of the RPC fate. For example, PAX6, RAX, and SIX6 promote RPC proliferation, 16 while SOX2, SUFU, and PAX6 are critical for the maintenance of RPC multipotentiality. 17,27 –29 Inactivation of PAX6 in RPC results in the exclusive generation of amacrine cells. 27 Sox2-deficient RPC lose their ability to produce neuronal cells, 28 and Sufu-deficient RPC adopt an amacrine or horizontal cell fate. 17
A complex, hierarchical gene regulatory network, which controls RPC differentiation and photoreceptor cell development, has been extensively studied (Fig. 1). Six transcription-regulatory factors are needed to determine rod or cone cell fate: retinoid-related orphan nuclear receptor (RORβ), orthodenticle homolog 2 (OTX2), neural retina leucine zipper protein (NRL), cone-rod homeobox protein (CRX), photoreceptor-specific nuclear receptor NR2E3, and thyroid hormone receptor β2 (TRβ2). 30 NRL is the dominant transcription factor that directs photoreceptor differentiation into rod cells. 31,32 On the other hand, retinas of CRX knockout mice lack photoreceptor outer segments, and the photoreceptors will rapidly degenerate after birth. 18 NRL interacts with CRX to control the expression of rod-specific genes 33 through a cis-regulatory module. 34 OTX2, which is important in determining the RPC fate of photoreceptor cells, 35 regulates the expression of several genes, including CRX, NRL, and NR2E3. 36 Knockout of OTX2 leads to a nearly complete loss of rods and cones. 35
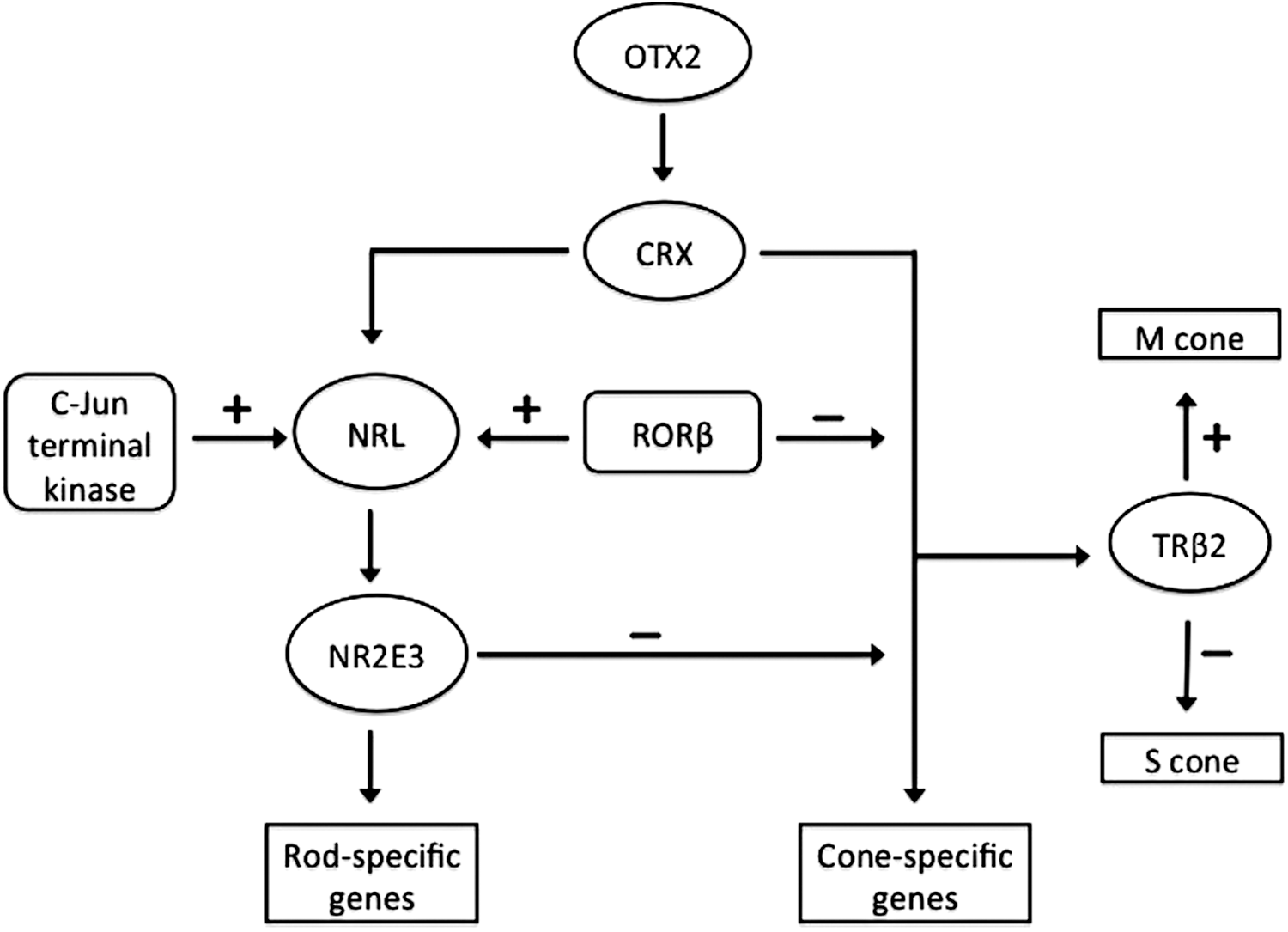
The regulatory networks in retinal progenitor cell (RPC) differentiation into photoreceptors. OTX2 is a key factor that determines the photoreceptor lineage. OTX2 controls neural retina leucine zipper protein (NRL) and cone-rod homeobox protein (CRX) in the transcriptional hierarchy in photoreceptor differentiation. NRL regulates its target NR2E3, which promotes photoreceptors to the rod cell fate and inhibits cone cell fate. CRX is also critical in regulation of rod and cone photoreceptor differentiation. NRL interacts with CRX to induce the expression of rod-specific genes. The progenitor cells adopt cone cell fate under the control of CRX and retinoid-related orphan nuclear receptor. Thyroid hormone receptor β2 promotes M opsin expression and inhibits S opsin expression in response to thyroid hormone. In addition, several signals such as c-Jun N-terminal kinase can regulate photoreceptor differentiations during retinal development through modulation of transcriptional activity of NRL.
Nuclear receptors with ligands such as retinoic acid and thyroid hormone, and orphan receptors with unknown ligands interact with other transcription factors to regulate photoreceptor differentiation and development. 37 The orphan nuclear receptor NR2E3, a transcriptional target of NRL, suppresses the development of cone cells and promotes differentiation of RPC into rod cells. 34,38 An orphan nuclear receptor (RORβ) is necessary for both rod cell differentiation and the development of functional cone cells. 30,39 TRβ2, a ligand-regulated nuclear receptor, is heavily involved in the cone opsin patterning response to thyroid hormone. 30,40,41
The binding of transcriptional regulatory factors is followed by activation of a set of downstream genes that control the differentiation and specification of RPC. NRL is required for the determination of rod cell photoreceptor differentiation. 31,32 CRX, OTX2, and RORβ promote NRL transcription by direct binding to a site within the NRL promoter. 42,43 Approximately 300 direct NRL target genes have been identified, some of which are critical for photoreceptor development. 34 Signaling molecules can also modulate the expression or activity of transcription factors by post-translational modifications such as phosphorylation and sumoylation. For example, c-Jun N-terminal kinase 1 (JNK1) phosphorylation of NRL at serine 50 enhances the NRL transcription during retinal development. 44 Sumoylation of NRL at Lys-20 promotes transcription of its downstream target genes. 45
Besides the expression of these transcription factors, differentiation of RPC into specific retinal cell types is also just as regulated by suppressing the formation of other retinal cell types simultaneously. For example, NR2E3 promotes the development of rod photoreceptors and also inhibits cone photoreceptor formation. PR domain zinc finger protein 1 (PRDM1) promotes photoreceptor production and suppresses bipolar cell fate in photoreceptor precursors. 46 ATOH7 and POU4F2 are required for RGC fate determination, but they also suppress non-RGC differentiation and specification. 47,48 OTX2 is required for photoreceptor generation and diminishes the generation of amacrine cells from RPC. 35 Other transcription factors are also critical for subtype specification of retinal neurons. TRβ2 promotes M cone formation and inhibits S cone formation. BARHL2 plays a significant role in amacrine cell determination. 49
Extrinsic factors
Extrinsic signaling pathways, including WNT, NOTCH, Sonic hedgehog (Shh), and growth factors, modulate the RPC fate during retinal development. WNT activation can promote or inhibit RPC differentiation. 16 Activation of the Notch-Delta signaling pathway hinders neuronal differentiation, promotes RPC maintenance and proliferation, 50 and accelerates the cell cycle. 51 Vascular endothelial growth factor (VEGF) can increase or inhibit RGC production. 52 The extrinsic signaling pathways that regulate retinal development have been discussed in several recent reviews. 14,16,50,51,53,54
It is suggested that RPC could process signals from both extrinsic factors and intrinsic factors in order to differentiate into specific retinal cell types. 14 Multiple extrinsic signaling pathways can regulate the expression of transcription factors. For example, NRL is regulated by fibroblast growth factor (FGF) and Shh 55 –57 ; ATOH7 is regulated by FGF and Shh. 58,59 Activation of the hedgehog pathway is mediated by three Gli transcription factors, resulting in the modulation of 30 target genes. 60 Multiple signaling pathways can converge to a single transcription factor, as shown by the activation of the Notch effector gene HES1 by the Shh and VEGF pathways to regulate RGC differentiation. 52,61 A number of other transcription factors can activate extrinsic signaling pathways that regulate RPC fates. Forkhead box N4 (FOXN4) suppresses the photoreceptor cell fate of early RPC by inducing DII4 expression with subsequent activation of the DII4-Notch signaling pathway. 62
Although complex signaling pathways are known to be involved in RPC differentiation, the mechanisms responsible for determination of particular cell fates remain unclear. The next section summarizes the recent advances of mechanisms regulating the lineage specification and differentiation of RPC into retinal neurons, notably RGC.
Transcription Factors that Determine RGC Differentiation and Cell Fate
Most transcription factors that regulate RPC heterogeneity, RGC differentiation, and specification 63 belong to the basic helix-loop-helix (bHLH), foxhead, and homeodomain families, 59,64 and regulate a wide range of genes that direct specification of each retinal cell type.
Transcription factors that determine RPC heterogeneity
Early lineage analyses have shown that the cells of neural retina develop from a pool of multipotent progenitors. 65,66 Gene expression profiling has revealed that RPC are heterogeneous with a mixture of subpopulations. 67 The roles of transcription factors in modulating the development of RPC subpopulations toward specific retinal cell fates are diverse. 14,67 ATOH7 influences the generation of post-mitotic RGC, but it has only been expressed in a subset of RPC. 67 –69 OLIG2-expressing RPC tend to become rod photoreceptors and amacrine cells, 70 while ASCL1 restricts RPC to become RGC. 13 Pax6-positive RPC will commit to RGC and amacrine cell fates. 71 The available evidence suggests that the competencies to generate particular retinal cell types are determined by transcription factors expressed within the RPC population.
Transcription factors for RGC competence
The competence of RPC to generate RGC is determined by various transcription factors. PAX6 and SOX2 are required for the specification of retinal neurons; inactivation of either gene in RPC results in failure to generate RGC and other retinal neuronal cells. 27,28,72 Knockout of either PAX6 or SOX2 results in a loss of ATOH (also called Math5) expression. 27,28,59,64,73 ATOH7 is a proneuronal bHLH transcription factor expressed in a subpopulation of RPC that commit to RGC fate, 13,59,64 suggesting that it is necessary for RGC differentiation. However, not all ATOH7-expressing RPC differentiate into RGC. 74 Some of these cells differentiate into other retinal cell types. 74,75 ATOH7-deficient mice exhibit an almost but not complete RGC loss, 48,76 suggesting that some ATOH7-independent specification programs determine generation of RGC.
Neurogenic differentiation 1 (NEUROD1) is an ATOH7-independent transcription factor that plays a role in RGC specification. 77,78 Replacement of the ATOH7 gene with NEUROD1 (another bHLH gene) reduces the loss of RGC seen in ATOH7-deficient mice. 77 The ATOH7-independent program for initiating RGC development can be suppressed by REST/NRSF, which is a transcriptional repressor of neuronal differentiation. 79
A number of transcription factors contribute to RPC competence and RGC fate determination (Fig. 2), including ASCL1 (MASH1), NGN2 (NUEROG2), and OLIG2, which are expressed in RPC in every possible combination. 13 RPC that express NGN2 can differentiate into all the retinal cell types. 13,80 The transcription factor ASCL1 works similarly as NGN2 in RPC differentiation, but the RPC rarely commit to RGC fate. 13 ASCL1-deficient mice generate normal RGC number, but ASCL1 expression does not alter RGC development, and only a small number of the descendants of ASCL1-expressing cells express ATOH7, which is required for RGC competence. This suggests that ASCL1 restricts RPC competence in a specific cell lineage only. RGC development in this cell lineage expressing other transcription factors is not affected. 13
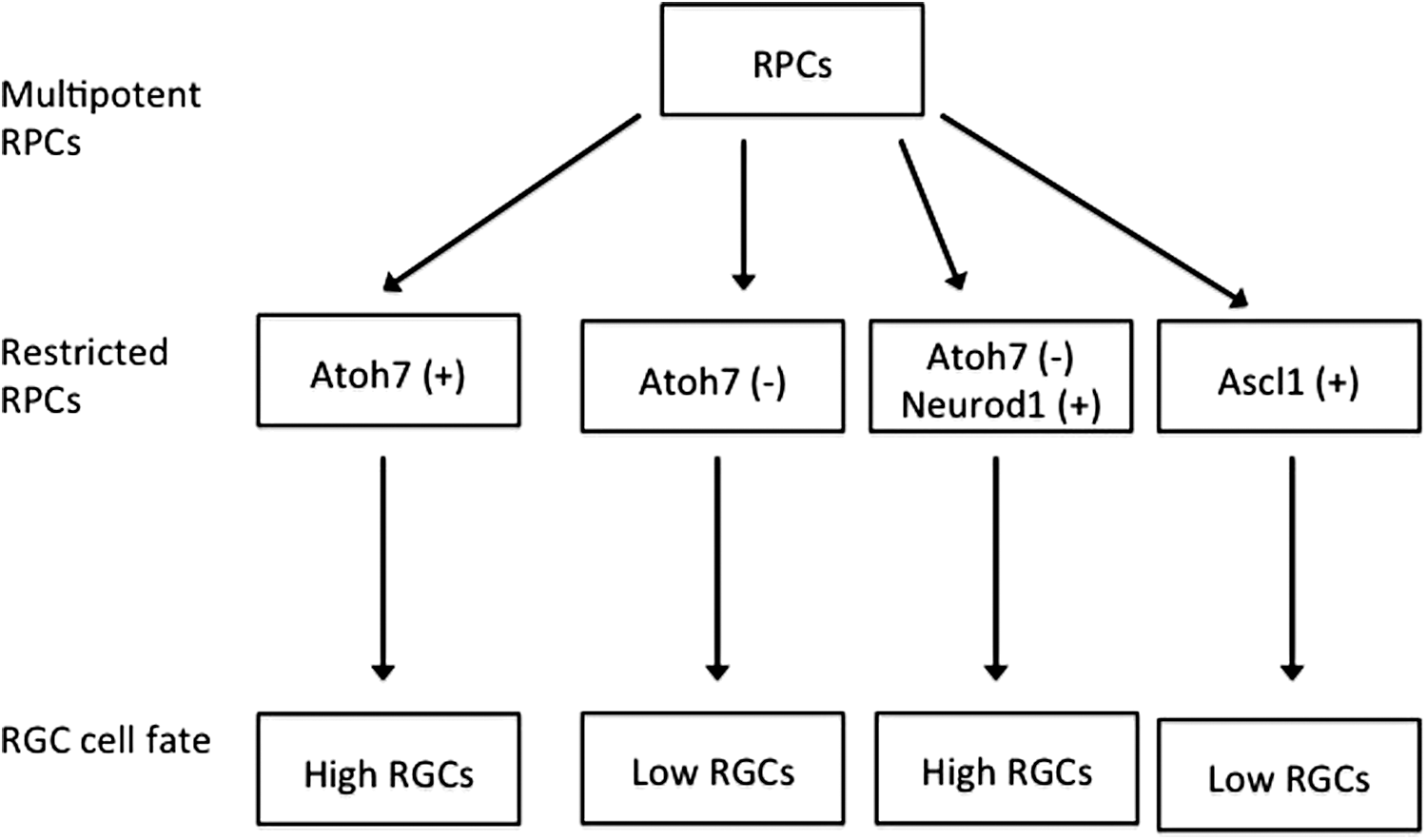
Transcription factors determine retinal ganglion cell (RGC) fate. ATHO7 is required for RGC development, but not all ATOH7-expressing RPC differentiate into RGC. Knockout of ATOH7 results in a severe but not complete loss of RGC. NEUROD1, an ATOH7 independent transcription factor, can partially rescue the severe loss of RGC. ASCL1-expressing RPC generate fewer RGC.
The molecular basis of temporal patterns in the RPC competence to particular retinal cell types has also been explored. The transcriptional regulator IKAROS (IKAROS family zinc finger 1) is responsible for generating early-born neurons, including RGC. 81 Inactivation of IKAROS decreases the number of early-born neurons (RGC, amacrine cells, and horizontal cells) but does not affect the late-born neurons. Misexpression of IKAROS in RPC results in promotion of early-born retinal neurons. 81 Therefore, timing of IKAROS inactivation during RGC specification is important for the generation of late-born neurons.
miRNA might be responsible for IKAROS inactivation. Dozens of miRNAs are expressed in the developing retina and are coupled to developmental timing and cell fate. 82,83 In the absence of the miRNA processing enzyme Dicer, generation of early-born retinal neurons (RGC and horizontal cells) continues beyond the normal competence window, suggesting that miRNA plays a key role in early progenitor competence. 84 Since misexpression of IKAROS promotes generation of early-born retinal cells during RGC specification, IKAROS is possibly one of the targets on which miRNA acts.
Influential Factors in RGC Development
Microarray analysis shows that ATOH7 regulates a number of potential downstream genes, 85 two of which, POU4F2 and ISL1, encode the POU-domain factor POU4F2 (also called Brn3b) and the LIM-homeodomain factor ISL1, and they are positively regulated by ATOH7. Transcription factors POU4F2 and ISL1 are critical to determine the RGC fate and differentiation.
POU4F2 is one of the three POU4F family members (Pou4f1–3 or Brn3a–c) that is expressed in RGC 86 at various stages such as in migrating RGC precursors, differentiating, and mature RGC. 87,88 POU4F2 inactivation could results in defects of RGC differentiation leading to most RGC loss, optic nerve hypoplasia, and axon growth defects. 24,88 –90 Knockout POU4F2 causes RGC precursors to adopt amacrine or horizontal cell fates and misexpression of POU4F2 suppresses non-RGC cell fates. It suggests that POU4F2 specifies RGC fate in precursors by downregulation of the non-RGC differentiation program. 47 Differential expression of POU4F2 in subgroups of RGC with distinct properties suggests that it could be involved in the regulation of specific cell functions. 62 POU4F2 regulates hundreds of downstream genes that encode transcription factors and proteins involved in neuronal integrity and synaptic vesicle function, all of which identify POU4F2 as an influential factor in RGC development, morphology, and function. 91
ISL1 is co-expressed with POU4F2 and resembles POU4F2 in the regulation of RGC development. 92,93 As with POU4F2, inactivation of the ISL1 gene also results in the loss of most RGC, axon growth defects, and optic nerve hypoplasia. Knockout ISL1 and POU4F2 leads to severe retinal abnormalities with a nearly complete RGC loss, suggesting that ISL1 and POU4F2 have redundant but distinct functions in the regulation of RGC development. 92 In fact, similar downstream targets are shared by these two transcription factors. 92,93 In addition, ISL1 is necessary for sustained expression of POU4F2, suggesting that ISL1 may positively regulate POU4F2 acting in parallel with or downstream of POU4F2, since deletion of ISL1 does not alter the expression of POU4F2 in the developing retina. 93
Extrinsic Signaling in RGC Differentiation
Extrinsic signaling molecules such as FGFs, Shh, and Delta-Notch can act either positively or negatively to regulate differentiation of RPC and RGC specification. 14,16 FGFs are involved in the development of RGC beginning in the center of the retina spreading toward the periphery. 94 The initial steps of RGC differentiation are offset by FGF3 and FGF8 from RPC, and they also regulate ATOH7 expression. 58 FGF signaling promotes ATOH7 expression by activating ATOH7 genes through a spatial regulatory sequence. 59 FGF signaling is also involved in the spreading of a RGC differentiation wave generated by Shh. 16 FGF19 activates Shh expression by regulation of PEAS and ERM genes to initiate the wave of RGC differentiation. 16,95 Additionally, FGF signaling plays a crucial role in the distribution of guidance cues of RGC axon required for normal optic tract development. 96
Shh signaling is required for RGC differentiation initiated by FGFs 16,95 and appears to function as a negative regulator of RGC development. 51 Overexpression of Shh in the chick retina reduces RGC production, while inhibition of endogenous Shh activity leads to increased production of RGC. 97 Conditional knockout of Shh or Smo (smoothened) in mice results in overproduction of RGC. 61,98 The enhanced RGC production found in SMO knockout retinas is abolished by suppressing ATOH7 gene expression, suggesting that ATOH7 mediates the Shh effects on RGC specification. 61 Shh-mediated inhibition of RGC development is also associated with its activation of the DII-NOTCH effector gene HES1, which suppresses RGC differentiation. 61,98 Finally, Shh signaling also mediates the growth and guidance of RGC axons 99 and indirectly controls RGC axon guidance cues through patterning of the optic stalk. 100
NOTCH encodes a transmembrane receptor that binds to ligands (Delta, Serrate, or Jagged) on the cell surface. Receptor-ligand binding initiates the γ-secretase-dependent cleavage of the NOTCH intracellular domain, which is translocated to the nucleus where it interacts with Rbpj transcription coactivators to activate the target HES (Hairy and Enhancer of Split) gene. This classic Delta-Notch signaling pathway inhibits the production of retinal neurons by maintaining RPC proliferation. 50 Activated NOTCH and elevated DELTA weaken RGC growth and disruption of the Delta-Notch pathway promotes RPC differentiation and hence RGC generation. 101,102 Involvement of NOTCH signaling has also been demonstrated by the autonomous suppression of RGC fate by RBPJ, a classical indicator of multiple NOTCH signals. 103 This is also in line with the result that inactivation of NOTCH pathway components, including NOTCH, RBPJ, and HES results in increased retinal neurogenesis. 102 –105 Conditional knockout of Dicer severely reduces Notch signaling 84,106 and induces the generation of RGC beyond their normal competence window. 84 However, rescue of Notch signaling has little effect on enhanced generation of RGC, suggesting that Notch pathway inactivation is not required for the initial stages of RGC differentiation. 106
Overall, the mechanisms detailed above summarize current the understanding of mechanisms regulating RGC generation. This knowledge is critical as we move forward to apply stem cell–derived RGC for clinical application.
Stem Cell–Based Therapy
Animal studies have shown that stem or progenitor cells transplants can be engrafted into injured mature retina, but they fail to differentiate fully into functional retinal neurons. 107,108 The mature retina lacks differentiation cues and/or permissive environment for the integration of stem or progenitor cells. 1,109 However, some studies have reported improvement in visual function, even though the transplanted cells integrated into the host retina to a limited extent. 110,111 Improvement of vision may be due to neuroprotection of host retinal neurons by trophic factors secreted from transplanted cells. 110 –112 However, the duration and longevity of the neurotrophic effect remains unclear. Aside from the trophic effect, integration of transplanted stem or progenitor cells may also contributes to such a phenomenon. 2,3,6 –9,14,108,113 –117 MacLaren et al. reported that transplanted photoreceptor precursors differentiated into rod cells that formed functional synapses in three mouse models of inherited retinal degeneration (rd), including retinal degeneration slow (rds), retinal degeneration fast (rd), and rhodopsin knockout (rho–/–). 3 The developmental age of engrafted cells is critical for integration and differentiation into functional photoreceptors. 3 Recently, Lamba et al. reported that the transplantation of human ESC-derived RPC partially restored visual function in Crx knockout mice, a mouse model of Leber's congenital amaurosis. 7
Although there is no evidence showing vision improvement following transplantation of RGC, transplanted cells derived from ESC and mesenchymal stem cells (MSC), and RPC can integrate into the ganglion cell layer and express RGC markers. 107 –110 Further studies need to be done to examine if the transplanted cells could extend axons entering the optic nerve.
In a preliminary clinical report, Schwartz et al. 10 reported the first subretinal transplantation study of human ESC-derived RPE in patients with advanced vision loss. One patient had Stargardt macular dystrophy and the other had dry AMD. Using the Early Treatment Diabetic Retinopathy Study Chart for reference, the AMD patient had an improvement in visual acuity from 20/500 to 20/200 with stable vision for 3 months. Mild improvement in visual function was observed in the fellow eye. Visual acuity in the patient with Stargardt macular dystrophy improved from seeing hand motions to 20/800 with stable vision for 1–3 months post-transplantation. Improvement in color vision, contrast, and dark adaptation were also detected in the treated eye, whereas no improvement was observed in the control eye. Although this seminal study confirmed the feasibility and safety of human ESC-derived cell transplantation, a larger and long-term cohort study is needed to determine the efficacy, safety, and tolerability of human ESC-derived RPE.
Unlike the generation of RPE, it is still challenging to generate a high purity of RGC for replacement study in optic neuropathy diseases such as glaucoma. It is not surprising that a paucity of clinical data is available for RGC replacement therapy. The next section focuses on the molecular events underlying RGC generation from RPC and their potential application in RGC replacement therapy.
Generation of RGC for Cell Replacement Therapy
Glaucoma is the second leading cause of blindness. The pathogenesis of glaucoma is marked by progressive degeneration of RGC and their axons 118 with elevated intraocular pressure. Initially, RGC axon damage is believed to occur in the lamina area, and subsequently these changes activate multiple pathways leading to axonal degeneration and RGC death via multiple mechanisms. 119 Generation of homogenous population of RGC is critical toward the goal of stem cell–based therapy to treat optic neuropathy diseases such as glaucoma.
A number of studies have suggested that stem cell–derived RGC can be produced by recapitulating the mechanisms responsible for RGC generation during retinogenesis. Transfection of ESC with PAX6 can induce the formation of RGC-like cells in vitro, 120 while transient expression of the intrinsic factor ATOH7 in RPC leads to upregulation of RGC markers such as POU4F2 and induction of RGC generation. 121 Overexpression of ATOH7 results in the generation of RGC-like cells from iPSC. 122 RGC generation is also enhanced in the presence of NOGGIN (DN), which inhibits Notch signaling activity and harks back to the previous discussion on extrinsic signals in RGC differentiation. Furthermore, the microenvironment of early retinal progenitors promotes differentiation of RPC into RGC. 123 Production of iPSC-derived RGC could even occur in response to stimulation within the microenvironment of early retinal histogenesis without any genetic manipulation. 124 Exogenous growth factors such as FGF2 and insulin are required for RGC generation, 4,125 and generation of FGF2-induced RGC is enhanced by Shh exposure. 4
In vivo RGC generation following stem cell transplantation has yet to be reported in animal models. Other studies have shown transplantation of ESC or iPSC would mainly differentiate into photoreceptors. 7,126 It has also been reported that transplanted cells derived from ESC and neural progenitor cells could migrate and integrate into host retina and also express RGC markers. 107 –110 In addition, Hertz et al. reported that transplantation of RPC-derived RGC could survive and integrate into the RGC layer of rats with normal or injured retinas. 127 Although iPSC-derived RGC survived after intravitreal injection into the mouse retina, these cells may not integrate into the host retina. 122
Replacement studies of photoreceptor progenitors suggest that the stage of transplanted cells is critical to the integration into the host retina. 3 So far, reports show that transplantation of neuronal precursors, RPC and RGC, integrates with the host retina to a limited extent. 4,122,128 Thus, a comprehensive study is needed to examine the outcomes of integration and axonal growth following transplantation of various stages of RGC progenitors into the retina.
To this day, cell therapy studies for retinal degenerative diseases using stem cell–derived RGC are lacking. Mellough et al. found that NSC-like cells could integrate into ganglion cell layer of neonatal retina with RGC depletion. 128 Nevertheless, most integrated cells do not express RGC markers.
In addition to integration into the host retina and reconnecting to retinal circuity, the effect of stem cell–based therapy may be also be mediated by a neuroprotective effect. Recently, Satarian et al. reported that the transplantation of human iPSC-derived anterior-specific neural progenitors integrated into the RGC layer and that these progenitors express neuronal markers in an animal model of optic nerve crush injury. However, the improvement effect was mainly facilitated by the production of neurotrophic factors (NFs). 129
Deprivation of NFs has been implicated in the pathophysiology of neuronal loss such as glaucoma. 130 –132 Purified or recombinant NFs have been extensively examined for their role in the neuroprotection of RGC. 129,133 –135 Transplantation studies of stem cells have shown that enhanced neuronal survival was observed in the degenerative retinas via production of NFs. 5,108,134,136 –138
Neuronal progenitors and MSC are known to secrete a wide variety of NFs such as ciliary neurotrophic factor (CNTF), brain-derived neurotrophic factor (BDNF), and fibroblast growth factor (FGF). 108,129,134,138 However, the direct role of NFs in RGC protection remains ambiguous.
Several in vitro and in vivo studies have investigated the neuroprotective effect of NFs, including BDNF and CNTF, on RGC. BDNF released from engineered MSC improved RGC survival in vitro and also in a chronic hypertensive rat model. 139 These studies posit that MSC can be an effective vehicle for delivering NFs. Fang et al. reported that elevated expression of BDNF and CNTF following transplantation of iPSC without c-Myc in rats with retinal ischemia and reperfusion injury augmented the beneficial effect of cell replacement therapy. 135 Nevertheless, the roles of NFs in stem cell–based therapy for RGC loss have yet to be further elucidated.
In summary, the more the underlying mechanisms to generate RGC during retinal development or from stem cells are understood, the more stem cell–based therapy will become feasible to treat optic neuropathic diseases such as glaucoma. However, to reach the goal, a few hurdles need to be overcome. First, a high purity of RGC at an appropriate stage needs to be generated from ESC or iPSC. Second, the best stage of disease needs to be identified to receive cell transplants for robust integration, functional connections to host retinal neurons, and axon extension toward the optic nerve. Third, a robust growth of RGC axons toward the brain is needed, and the visual targets need to be reinnervated. Although these obstacles are challenging, overcoming them is not beyond the realm of possibility. Recently, Benowitz's group showed that administration of Zymosan and cAMP analogue in PTEN knockout mice is sufficient to induce robust optic nerve regeneration, reinnervate to the visual targets, and regain partial recovery of visual functions. 140 It provides hope for the success of stem cell–based therapy for optic neuropathy diseases such as glaucoma in the future.
Safety and Ethics of Stem Cell Therapy
Using ESC from the human fetus to treat neurodegenerative diseases has raised ethical concerns. 111,141,142 After the development of inducing somatic cells to become pluripotent cells called iPSC, such concerns have been largely avoided.
Stem cell–based therapies are also privy to safety issues such as the tumorigenic potential of undifferentiated stem cells, especially ESC. 111,143 Although some studies have reported that teratoma has been detected following transplantation of rodent 143 –145 or human 143 ESC-derived grafts, other studies have shown human 10,146 –149 or mouse 126,150 ESC-derived transplants did not form teratoma. Such teratoma/tumor formation may be due to the purity of the transplanted cells. Therefore, it is crucial that selection be stringent in order to eliminate any undifferentiated pluripotent stem cells.
Perspectives
The role of intrinsic and extrinsic factors in the generation of RGC has been discussed. Based on this knowledge in the field, we have a better idea of how to differentiate stem or progenitor cells into RGC. Thus, it is anticipated that the field of regenerative medicine will move on to formulate an approach to generate stem cell–derived RGC. With a homogenous population of RGC, it will be possible to examine the ability of integration and axonal growth in the host retina. That said, further investigation should explore the mechanisms responsible for integration and axonal growth of the stem cell–derived RGC in the retina.
Footnotes
Acknowledgments
The study was supported by grants from National Natural Science Foundation of China (grant no. 30901645) and the Youth Innovation Fund of the Third Military Medical University (grant no. 2009XQN29).
Author Disclosure
No competing financial interests exist.