
Abstract
Adeno-associated virus (AAV) has emerged as the preferred vector for targeting gene expression to the retina. Subretinally injected AAV can efficiently transduce retinal pigment epithelium and photoreceptors in primate retina. Inner and middle primate retina can be transduced by intravitreally delivered AAV, but with low efficiency. This is due to dilution of vector, potential neutralization of capsid because it is not confined to the immune-privileged retinal compartment, and the presence of the inner limiting membrane (ILM), a barrier separating the vitreous from the neural retina. We here describe a novel “subILM” injection method that addresses all three issues. Specifically, vector is placed in a surgically induced, hydrodissected space between the ILM and neural retina. In an initial experiment, we injected viscoelastic (Healon®), a substance we confirmed was biocompatible with AAV, to create a subILM bleb and subsequently injected AAV2-GFP into the bleb after irrigation with basic salt solution. For later experiments, we used a Healon–AAV mixture to place single, subILM injections. In all cases, subILM delivery of AAV was well tolerated—no inflammation or gross structural changes were observed by ophthalmological examination or optical coherence tomography. In-life fluorescence imaging revealed profound transgene expression within the area of the subILM injection bleb that persisted for the study duration. Uniform and extensive transduction of retinal ganglion cells (RGCs) was achieved in the areas beneath the subILM bleb. Transduction of Müller glia, ON bipolar cells, and photoreceptors was also observed. Robust central labeling from green fluorescent protein-expressing RGCs confirmed their continued survival, and was observed in the lateral geniculate nucleus, the superior colliculus, and the pretectum. Our results confirm that the ILM is a major barrier to transduction by AAV in primate retina and that, when it is circumvented, the efficiency and depth to which AAV2 promotes transduction of multiple retinal cell classes are greatly enhanced.
Introduction
A
Intravitreal injection is being used to target AAV to retinal ganglion cells (RGCs) of patients with Leber hereditary optic neuropathy (LHON) in ongoing clinical trials. 18 However, emerging data highlight limitations in this approach in primates, including humans. Although some RGCs can be transduced in NHPs by intravitreally injected AAV2-based vectors, the area of transduction is restricted primarily to a “ring” of RGCs around the fovea. 19,20 Even the newest AAV vectors (those rationally designed or mined via screening of complex AAV capsid libraries) delivered intravitreally in NHPs promote transduction that is primarily restricted to foveal cones, and ganglion cells/Müller cells in the foveal ring and areas surrounding major blood vessels (see Refs. 20 and 21; and S.E. Boye and P.D. Gamlin, unpublished data).
Compelling proof of concept exists for AAV-mediated transduction of other retinal cell types in rodent models of inherited retinal disease after either subretinal or intravitreal injection. 1,22 However, in NHPs, other retinal cell types identified as potential therapeutic targets, such as ON bipolar cells (ON BCs) and Müller glia, are inefficiently transduced by AAV after either intravitreal or subretinal injection, except for some small patches of cells at the site of nearby large retinal vessels. 14,20,23 –31
Overall, intravitreal injection of AAV is relatively less invasive than subretinal injection, and perhaps better suited for mediating therapy in patients with fragile, diseased retinas that are prone to further damage on surgically induced retinal detachment. However, there are three known barriers to the effective pan-retinal transduction of primate retina by intravitreally delivered AAV vectors, the first of which is the dilution effect. Intravitreally delivered vector is immediately diluted on mixing with the vitreous humor. Relative to what has been tested in rodents (1-μl injection volume into 10-μl vitreous humor volume of mouse), intravitreally delivered vector is even more diluted in primates (0.05–0.10 ml into ∼2.5 ml of vitreous humor). 32 Second, intravitreally delivered capsid is not confined to the immune-privileged retinal compartment and is thus at increased risk for neutralization. 33 The third is the physical barrier of the inner limiting membrane (ILM). 34 Primates have a more impenetrable ILM than rodents, except in and around the fovea, immediately above the optic nerve head and close to large blood vessels. 35 When the ILM is physically degraded by enzymatic digestion or lacks structural integrity because of genetic manipulation, AAV transduction in both rodents and NHP may be improved. 19,36,37 However, the long-term implications of such disruption for the structure/function of the neural retina have not been investigated. To achieve more efficient and widespread transduction of primate retina by AAV, alternative delivery strategies and surgical approaches should be considered.
Surgical manipulation of the transparent, acellular ILM is assisted by staining with dyes and is routinely performed in patients to correct macular holes, macular puckers, and myopic foveoschisis. 38 –41 We therefore sought to circumvent the barriers to transduction of NHP retina associated with intravitreal injection by using a novel surgical approach termed “subILM injection,” wherein vector is directly placed in a surgically induced, hydrodissected space between the ILM and neural retina. This approach eliminates the impact of all three barriers identified above. After such subILM injections with AAV2, we characterized the cell types transduced and the extent of transduction relative to intravitreal-delivered AAV2 in multiple NHP subjects.
Materials and Methods
AAV vectors
AAV vector plasmid containing the full-length chicken β-actin (CBA) promoter driving green fluorescent protein (GFP) was packaged in AAV serotype 2. A separate self-complementary vector plasmid containing a truncated CBA (smCBA) promoter driving mCherry reporter was packaged in AAV2. Both vectors were purified and titered according to previously published methods. 42,43 The final concentration of AAV2-CBA-GFP was 1.66 × 1012 vector genomes (VG)/ml in balanced salt solution (BSS) (Alcon, Forth Worth, TX) supplemented with 0.014% Tween 20.
In vitro AAV + Healon transduction assay
Self-complementary (sc)AAV2-smCBA-mCherry was incubated with Healon® ophthalmic viscoelastic (10-2952-10; Abbott Medical Optics, Santa Ana, CA) at a ratio of 3:1 (AAV:Healon) for either 5 min, 15 min, or 1 hr before being applied in serum-free medium to 661W mouse cone-derived photoreceptor cells (generously provided by M. Al-Ubaidi, University of Oklahoma, Oklahoma City, OK). The self-complementary AAV construct was used to decrease the time to onset of transgene expression. Cells were incubated with scAAV2 vector at a multiplicity of infection (MOI) of 2000. One hour postinfection, 300 μl of serum-containing medium (10% fetal bovine serum [FBS], glutamine [300 mg/liter], putrescine [32 mg/liter], 40 μl of 2-mercaptoethanol, 40 μg of hydrocortisone 21-hemisuccinate and progesterone, penicillin [90 units/ml], and streptomycin [0.009 mg/ml]) was added to each well and incubated for 72 hr at 37°C in 7% CO2. Three days postinfection, cells were imaged with a fluorescence microscope (EVOS XL core imaging system; Thermo Fisher Scientific, Waltham, MA), dissociated, and their mCherry fluorescence quantified via fluorescence-activated cell sorting (FACS) as previously described. 44
Animals
All procedures performed on NHPs were approved by the Institutional Animal Care and Use Committees at the University of Alabama at Birmingham (Birmingham, AL) and were conducted in accordance with the Association for Research in Vision and Ophthalmology statement for the use of animals in ophthalmic and vision research. Three male Macaca nemestrina subjects (F91-108, 18 yr; EN-28, 7 yr; EV-44, 7 yr) were used in this study.
SubILM injection
All NHP surgical procedures were carried out under sterile conditions in a dedicated veterinary ophthalmic surgical suite. Subjects were sedated, intubated, dilated, and continuously monitored as previously described. 13 The right eye (oculus dexter, OD) of each of the subjects (F91-108, EN-28, and EV-44) was prepared with Betadine scrub and draped in sterile fashion. A standard 23-gauge three-port pars planar vitrectomy was performed to access the ILM in the right eyes. A Machemer-style infusion contact vitrectomy lens was used. Sclerotomies were performed ∼3.75 mm posterior to the limbus, and the ILM was visualized with IC-Green (Akorn, Lake Forest, IL), which was promptly flushed. All subILM injections were performed with a Duke 36-gauge, 60°-angled retinal needle (Grieshaber/Alcon, Fort Worth, TX) and 500-μl gas-tight Hamilton syringe, which was manually operated for injections in the first animal (F91-108), and by a Nanomite syringe pump (Harvard Apparatus, Holliston, MA) for injections in the other two animals. We targeted a temporal area, free of large retinal vessels, approximately 2–3 mm from the fovea, with the cannula aimed toward the fovea with the goal of placing a subILM injection that would include much of the macula. In the first animal (F91-108), we initially created spaces underneath the ILM with viscoelastic before a subsequent vector injection. We used sodium hyaluronate viscoelastic at 10 mg/ml (Healon OVD; Abbott Medical Optics). This was followed by partial irrigation with BSS before injection of ∼10 μl of AAV (∼1.66 × 1010 VG delivered). We determined later that multiple injections of the same bleb were contraindicated and that the presence of Healon did not to interfere with vector transduction (Fig. 1). Therefore for EN-28 and EV-44, Healon and AAV (1.66 × 1012 VG/ml) were injected together at a 1:1 ratio. In EN-28, the Healon–AAV mixture was delivered at a rate of 150 μl/min (2.5 μl/sec) for 3 sec, yielding a 7.5-μl bleb of which 3.75 μl was AAV (6.23 × 109 VG delivered). In EV-44, the Healon–AAV mixture was delivered at a rate of 125 μl/min (2.08 μl/sec), yielding a 13.1-μl bleb of which 6.55 μl was AAV (1.09 × 1010 VG delivered). No posterior vitreous detachment was performed in any animal. A summary of animals and experimental details can be found in Table 1. A schematic of the subILM injection procedure and images from a surgical video in one animal (EN-28) are included in Fig. 2.

Transduction efficiency of self-complementary AAV2-smCBA-mCherry in 661W cone photoreceptor cells. Vector was preincubated with Healon® at a 3:1 (AAV:Healon) ratio for either 5 min, 15 min, or 1 hr and then injected at a multiplicity of infection (MOI) of 2000. Controls included uninfected cells and cells infected with vector alone. mCherry expression was calculated by fluorescence-activated cell sorting (FACS) by multiplying the percentage of positive cells by the mean fluorescence intensity in each sample. Error bars represent ±1 standard deviation.
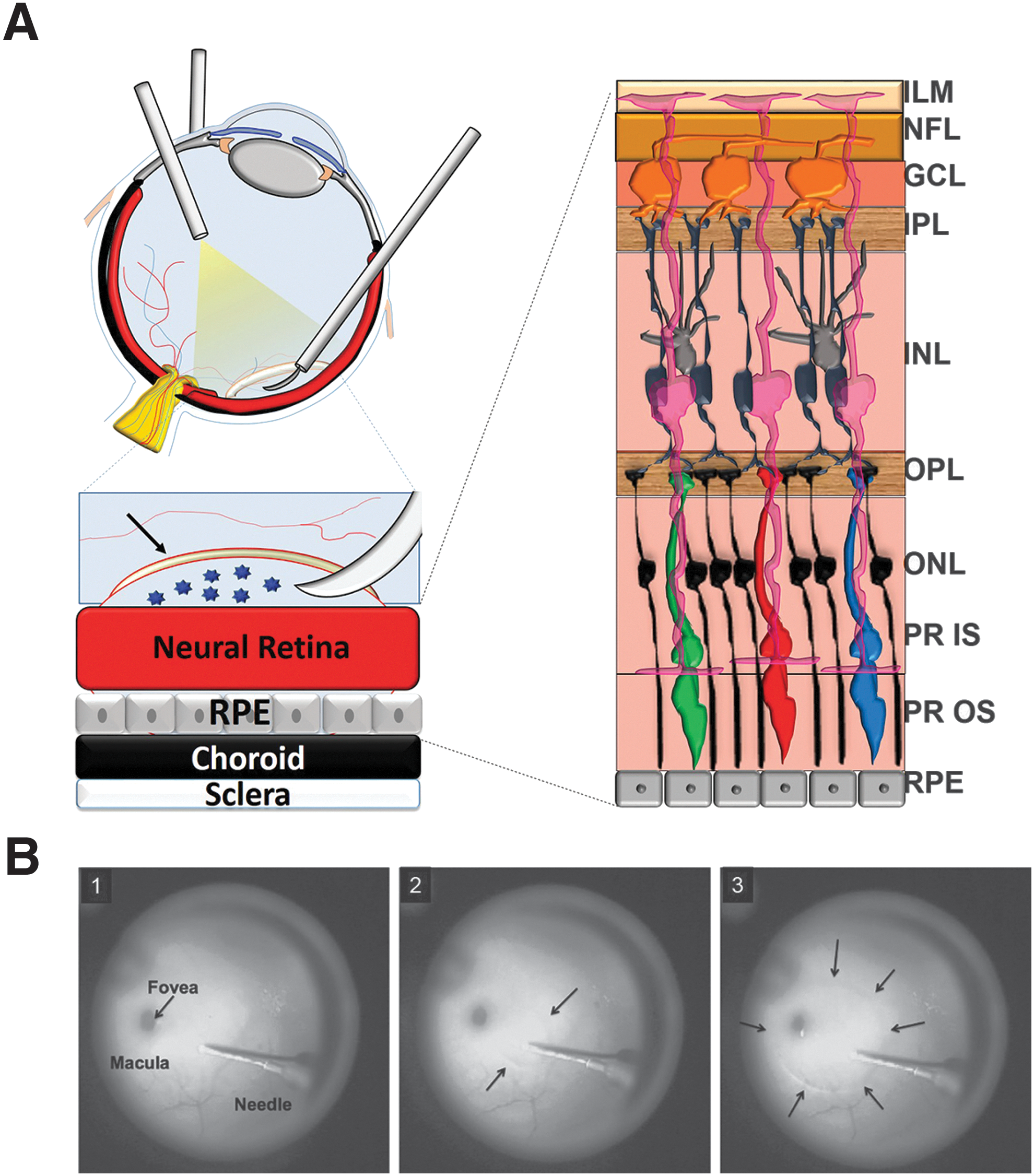
SubILM injection of AAV vector.
Animal and injection details
All subjects were male.
Vector reflux noted during surgery.
AAV, adeno-associated virus; BSS, balanced salt solution; ILM, inner limiting membrane; OD, oculus dexter (right eye); OS, oculus sinister (left eye); Tx, treatment; VG, vector genomes.
Intravitreal injections
Approximately 25 μl of AAV2-CBA-GFP containing 4.15 × 1010 VG (1.66 × 1012 VG/ml) was delivered intravitreally, using a 1-cm3 tuberculin syringe with 28-gauage, 0.5-inch needle to the left eye (OS) of subjects EN-28 and EV-44. The OS of subject F91-108 remained uninjected.
In vivo imaging
All NHP subjects were sedated, situated, and imaged with a Spectralis HRA+OCT (Heidelberg Engineering, Heidelberg, Germany) scanning laser ophthalmoscope as previously described. 13 Images were obtained with the 30-degree objective, using infrared (820 nm) and autofluorescence (488 nm) modes with and without optical coherence tomography (OCT). Volume OCT scans were obtained in the macular region of all treated eyes. F91-108 was imaged at approximately 3, 9, 10, 13, and 16 months postinjection. EN-28 was imaged at approximately 2 weeks, 1 month, 6 months, and 10 months postinjection. EV-44 was imaged at approximately 2.5 weeks, 1 month, 3 months, 6 months, and 15 months postinjection. Additional in-life images were captured with a Kowa RC-XV fundus camera with Sony digital imaging capability at the following time points: F91-108 (approximately 5, 7, and 9 months postinjection), EN-28 (10 days before surgery, and approximately 2 months postinjection), and EV-44 (approximately 2 months postinjection). A complete summary of in vivo imaging details can be found in Table 2.
In vivo imaging details
Spectralis images presented in text.
Fundus images captured.
Tissue preparation
NHP subjects F91-108, EN-28, and EV-44 were euthanized, perfused, and their eyes enucleated at approximately 16 months (F91-108), 10 months (EN-28), and 3.5 years (EV-44) postinjection. Anesthesia was initiated with ketamine followed by terminal anesthesia with pentobarbital (200 mg/kg body weight). Each animal was then perfused through the aorta with 2 liters of 1% sodium nitrite–0.9% sodium chloride, and fixed with 4 liters of 4% paraformaldehyde in 0.1 M phosphate buffer. F91-108 and EN-28 were enucleated, and their brains removed and stereotaxically blocked in the coronal plane at the level of the brainstem. Brains were placed in 30% sucrose in 0.1 M phosphate buffer for 3–5 days, and were then sectioned at 40 μm on a freezing, sliding microtome (AO 860). All sections from the level of the optic chiasm to the posterior superior colliculus were collected individually into 0.1 M phosphate buffer in 24-well tissue culture plates for later processing. Retinas from all eyes were processed as previously described. 13 Briefly, the anterior portion of the globe, lens, and vitreous were removed. Retinas were divided into five, ∼5 × 8 mm blocks for ease of sectioning. Tissue blocks were embedded/cryopreserved and their orientation within the blocks recorded. Blocks were sectioned with a Leica 3050 S cryostat (Leica Microsystems, Wetzlar, Germany) as described previously. 13 All retinal sections were cut at 10 μm and detailed notes retained during sectioning, keeping track of every section pulled, such that reasonable orientation and location within the block could later be achieved.
Immunohistochemistry and microscopy: retina
Cryosections of retinas from subjects F91-108 and EN-28 were stained and imaged as previously described. 13 Briefly, sections were rinsed with 1 × phosphate-buffered saline (PBS), incubated in 0.5% Triton X-100, and blocked in 10% goat serum–1 × PBS. Retinal sections were then incubated with primary antibodies raised against glutamine synthetase (rabbit polyclonal, diluted 1:500) (ab73593; Abcam, Cambridge, UK) and glial fibrillary acidic protein (GFAP) (chicken polyclonal, diluted 1:1000) (CPCA-GFAP; EnCor, Alachua, FL), in a solution containing 1% NHP serum–1 × PBS for ∼12 hr at 4°C. We relied on native GFP to visualize AAV-mediated transgene expression as, in our hands, immunostaining of GFP in cryosections of NHP retina diminishes the signal. Samples were then rinsed in 1 × PBS and incubated for 1 hr at room temperature with IgG secondary antibodies: goat anti-chicken (Alexa Fluor 647 conjugate) or goat anti-rabbit (Alexa Fluor 594 conjugate) (Life Technologies/Thermo Fisher Scientific). All were diluted 1:500 in a mixture of 1 × PBS containing 3% NHP serum. To eliminate autofluorescence that is characteristically found in NHP retinas (especially the retinal pigment epithelium), autofluorescence eliminator reagent (Cat. No. 2160; EMD Millipore, Billerica, MA) was applied to all retinal sections. After counterstaining with 4′,6′-diamino-2-phenylindole (DAPI) and rinsing with 1 × PBS, an aqueous-based anti-fade medium was applied (Prolong Gold; Life Technologies/Thermo Fisher Scientific) and the slides were coverslipped.
NHP retinas were imaged with a spinning disk confocal microscope (Nikon Eclipse TE2000 microscope equipped with a PerkinElmer UltraVIEW modular laser system and Hamamatsu O-RCA-R2 camera) as previously described. 13 All settings (exposure, gain, laser power) were kept constant across samples at each respective magnification ( ×10, ×20, or ×40). All image analysis was performed with Volocity 6.3.1 software and snapshots were exported in TIFF format.
Immunohistochemistry and microscopy: brain
All procedures were performed at room temperature unless otherwise stated. Free-floating brain sections were rinsed in PBS and then incubated in 1% hydrogen peroxide in PBS for 15 min to quench endogenous peroxidase activity. Sections were then blocked in streptavidin and biotin blocking solution (Vector Laboratories, Burlingame, CA) for 30 min each. Sections were rinsed in PBS and then blocked in 2% normal goat and normal monkey serum and 0.5% Triton X-100 in PBS for 1 hr. The sections were then incubated overnight at 4°C with a primary antibody raised against GFP (chicken IgY, diluted 1:1000) (600-901-215; Rockland, Limerick, PA) in a PBS mixture containing 2% normal goat and normal monkey serum, and 0.5% Triton X-100. Sections were then rinsed in PBS and incubated for 4 hr in biotinylated goat anti-chicken IgG (diluted 1:1000; Rockland) in PBS plus 0.5% Triton X-100. After rinsing, sections were incubated for 2 hr in streptavidin–horseradish peroxidase (HRP) (diluted 1:1000; Rockland) in PBS plus 0.5% Triton X-100. They were then rinsed and reacted with Pierce metal-enhanced DAB (3′,3′-diaminobenzidine) solution (Thermo Fisher Scientific) for 12 min. Sections were rinsed, mounted on gel-subbed slides, and allowed to dry. They were then defatted in xylene, and coverslipped with Permount.
NHP brain sections were imaged with a Zeiss Axioplan 2 fluorescence imaging system, and images were captured to computer with an AxioCam camera controlled from AxioVision. Images of the optic nerve, pretectum, superior colliculus, and magnocellular layers of the dorsal lateral geniculate nucleus (LGN) were captured to computer as single images. Images of the parvocellular layers of the LGN were generated in Photoshop as stitched montages of two separate images. Contrast and brightness of images were adjusted in Photoshop for optimal clarity.
Neutralizing antibody assays
ARPE-19 cells (purchased from the American Type Culture Collection [ATCC], Manassas, VA) were maintained and used for detection of neutralizing antibodies both before (with the exception of F91-108) and after injection as previously described with minor modifications. 13 Self-complementary AAV2-smCBA-mCherry vector (MOI, 5000) was diluted in serum-free DMEM/F-12 (1:1) modified medium and incubated with serial 1:4 dilutions (from 1:10 to 1:2560) of heat-inactivated serum samples in DMEM/F-12 (1:1) modified medium for 1 hr at 37°C. The serum–vector mixture was then used to infect ARPE-19 cells seeded in 96-well plates containing 3 × 104 cells per well for 1 hr. Three days postinfection, cells were imaged under a fluorescence microscope (Olympus IX 70 inverted fluorescence microscope equipped with a QImaging Retiga 4000R camera with RGB-HM-5 color filter and QImaging QCapture Pro 6.0 software; QImaging, Surrey, BC, Canada), dissociated, and counted/analyzed, using a BD LSR II flow cytometer equipped with BD FACSDIVA 6.2 software (BD Biosciences, San Jose, CA) as previously described. 13 The neutralizing antibody (nAb) titer was reported as the highest serum dilution that inhibited scAAV2-smCBA-mCherry transduction (mCherry expression) by ≥50%, relative to naive mouse serum.
Results
Healon does not negatively impact AAV transduction
Separation of the ILM from the underlying neural retina is improved during subILM surgery by including an ophthalmic viscoelastic (Healon). Our initial subILM surgery technique involved injection of Healon under the ILM, followed by irrigation with BSS and, ultimately, injection of AAV. Using this protocol, we reasoned that the AAV vector would interact with Healon in the bleb at a maximum ratio of 1 part Healon to 3 parts AAV. Therefore, before conducting in vivo experiments, we asked whether Healon substantially affected vector transduction by quantifying mCherry fluorescence in 661W mouse cone-derived photoreceptor cells that were infected with AAV2-smCBA-mCherry alone or with vector that was preincubated with Healon at a 3:1 ratio for 5 min, 15 min, or 1 hr. FACS analysis revealed that incubation of AAV2 with Healon for any of these time periods led to increased transduction of 661W cells (Fig. 1), with approximately 2- and 3-fold improvements observed after incubation in Healon for 15 min and 1 hr, respectively.
SubILM injections
In F91-108, we performed a core vitrectomy with partial removal of the posterior hyaloid face. However, we were unable to create a posterior vitreous detachment. Because of the amount of time spent (more than 30 min) trying to elevate and fully remove the posterior hyaloid face, the corneal epithelium became hazy during the surgery (similar to what occurs in humans). This quickly resolved after surgery but was a factor that led to some enlargement of the entry site of the bleb during virus injection as, by that time, the surgeon's visualization had deteriorated. After vitrectomy, three distinct subILM blebs were created with Healon in the OD of this subject. The first two blebs were small and the third, with the cleanest entry site, was the largest. We irrigated some Healon out of the largest bleb with BSS, and then injected AAV into this bleb. During the injection, the entry site was somewhat enlarged, which likely led to some virus leakage into the vitreous. There was also a transient compression of underlying retina during the surgery that is not seen in humans during this procedure and was unexpected given the thin nature of the ILM in this animal. On the basis of our surgical experience with this animal, we modified our procedures in subsequent animals to (1) not produce a complete posterior vitreous detachment, (2) premix Healon with vector to allow for only one injection procedure at each subILM site, and (3) use a microcontroller syringe pump to precisely control flow during the injection.
In EN-28, we performed a core vitrectomy with partial removal of the posterior hyaloid face. A single subILM bleb was created in the OD after injection of Healon:AAV (1:1). The injection occurred without incident and no reflux was observed. To view the vitrectomy, IC-Green injection and removal, and subILM injection, see Supplementary Videos S1, S2, and S3, respectively (supplementary data are available online at
In EV-44, we performed a core vitrectomy with partial removal of the posterior hyaloid face. A single subILM bleb was created in the OD of animal EV-44 after injection of 13 μl of Healon:AAV (1:1). Significant reflux of injection solution out of the bleb was noted during this surgery.
In general, all subILM blebs expanded somewhat after injections were completed, and it was assumed this was related to mild compression of the underlying retina and gradual detachment of the ILM as the retina returned to normal thickness over time.
In-life imaging reveals extensive transduction by subILM delivered AAV2 and long-term persistence of gene expression
To assess the pattern of AAV2-CBA-mediated GFP expression in subILM- and intravitreally injected subjects in life, scanning laser ophthalmoscopy was performed with a 488-nm autofluorescence filter. Imaging time points varied among subjects and are summarized in Table 2. Subject F91-108 was imaged on seven separate occasions between 3 months postinjection and the date of sacrifice (∼16 months postinjection). Vector-mediated GFP expression persisted for the duration of the in-life period after subILM injection, with areas of robust expression observed around the injection sites, the foveal rim, and the foveal pit (Fig. 3). GFP expression at the site of the two attempted subILM Healon injections that were not subsequently injected with vector could be attributed to ILM damage and subsequent leakage of vector into those sites from the vitreous. Expression within axons emanating from RGCs at the injection site and foveal rim were easily visualized at all time points (Fig. 3 and Supplementary Fig. S1). Transduction of peripheral RGCs was also observed (Supplementary Fig. S1). Moving from the temporal subILM injection site directly through the fovea and continuing nasal, GFP expression was apparent across ∼2600 μm of F91-108's retina (barring the nonfluorescent foveal rim). No diminution of this signal was apparent between 3 and 16 months postinjection. Cross-sectional OCT scans in the subILM-injected eye were generally unremarkable with only minimal cellular disorganization at the injection site (Fig. 3 and Supplementary Fig. S1). As expected, no GFP expression was observed in the uninjected OS of F91-108, and cross-sectional OCT scans through the fovea revealed no structural abnormalities.
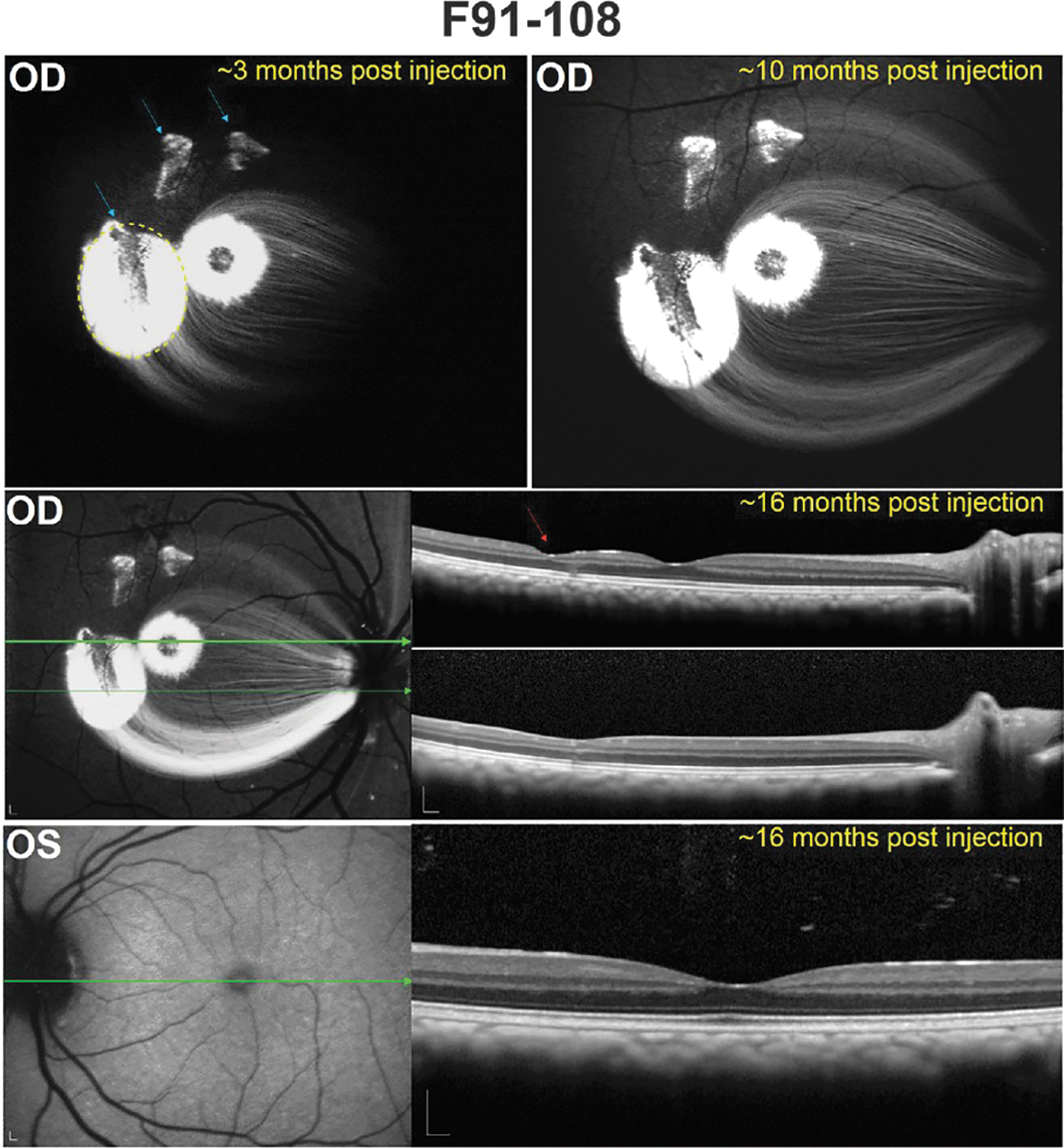
Fluorescence fundus images and corresponding optical coherence tomography (OCT) scans in the retina of F91-108. A representative image from the earliest postinjection time point (∼3 months) reveals the location of the three subILM blebs (blue arrows) in the OD (oculus dexter, right eye), only the largest of which was injected with AAV. The extent of this subILM bleb created during surgery is denoted by the dashed yellow line. Green fluorescent protein (GFP) expression, observed at the initial smaller injection sites, may have resulted from ILM damage and subsequent leakage of vector into those regions from the vitreous. There was no apparent diminution of GFP signal in this eye until at least ∼16 months after subILM injection. OCT scans through the injection site of the largest bleb show minimal cellular displacement (red arrow), and there were no apparent structural abnormalities within the fovea. As expected, a representative image of the uninjected OS (oculus sinister, left eye) obtained during the final imaging session reveals no GFP expression or retinal disorganization. The green arrows correspond to the location in the retina through which an OCT scan was collected (OCT scans are shown on the right in each instance). Scale bars: 200 μm.
Subject EN-28 was imaged 10 days before subILM and intravitreal injection and at five time points ranging from 13 days to 10 months postinjection. Scanning laser ophthalmoscope (SLO) images were acquired both in the infrared (IR) and blue reflectance mode and the 488 autofluorescence mode to visualize vector-mediated GFP expression. As shown in Fig. 4, there was robust fluorescence around the injection site except for an ∼650-μm diameter, central macular region that was devoid of fluorescence, barring a hint of fluorescence directly inside the foveal pit. The width of this nonfluorescent “spot” is consistent with the ganglion-free region of the retina, as well as the everted and mostly nontransduced inner nuclear layer (INL), and is accentuated by macular pigment that preferentially absorbs short-wavelength (blue) light. 45 Cross-sectional OCT scans taken 13 days postinjection revealed apparent detachment of the ILM from the underlying macular retina (white arrow in Fig. 4), an observation that was restricted to this subject alone. Slight displacement of cells within the INL at the foveal rim was also noted. Minimal cellular disorganization was seen at the injection site (red arrow in Fig. 4). Notably, the dark, narrow track that appears in en face SLO images, possibly an injection track, did not match the much more limited entry site made by the needle in the surgical video (Supplementary Video S1). Indeed, OCT scans confirm a lack of injection-related disorganization anywhere other than the perceived needle entry point (Fig. 4). By 6 weeks postinjection, slight tangential traction of the ILM is apparent as a lamellar macular defect, possibly as a result of a mildly fibrotic ILM, and some INL displacement is apparent (Supplementary Fig. S2). Overall, robust GFP expression was observed around the injection site, within the foveal rim and in RGC axons, and no obvious diminution in GFP signal was observed over time (Fig. 4). Moving from the temporal subILM injection site directly through the fovea and continuing nasal, GFP expression was apparent across ∼2800 μm of EN-28's retina (barring the nonfluorescent foveal rim). Two weeks after intravitreal injection, no AAV-mediated GFP expression was evident in EN-28's intravitreally injected OS (Fig. 4). Four weeks later, however, minimal GFP expression was observed in the foveal rim and sparse RGC axons of this eye (Supplementary Fig. S2) that persisted to at least 10 months postinjection (Fig. 4). GFP expression spanned a relatively small region (∼800 μm) of the central retina (barring the nonfluorescent foveal rim). No structural abnormalities were observed in the intravitreally injected eye, as evidenced by OCT scans encompassing the region of expression (Fig. 4 and Supplementary Fig. S2).
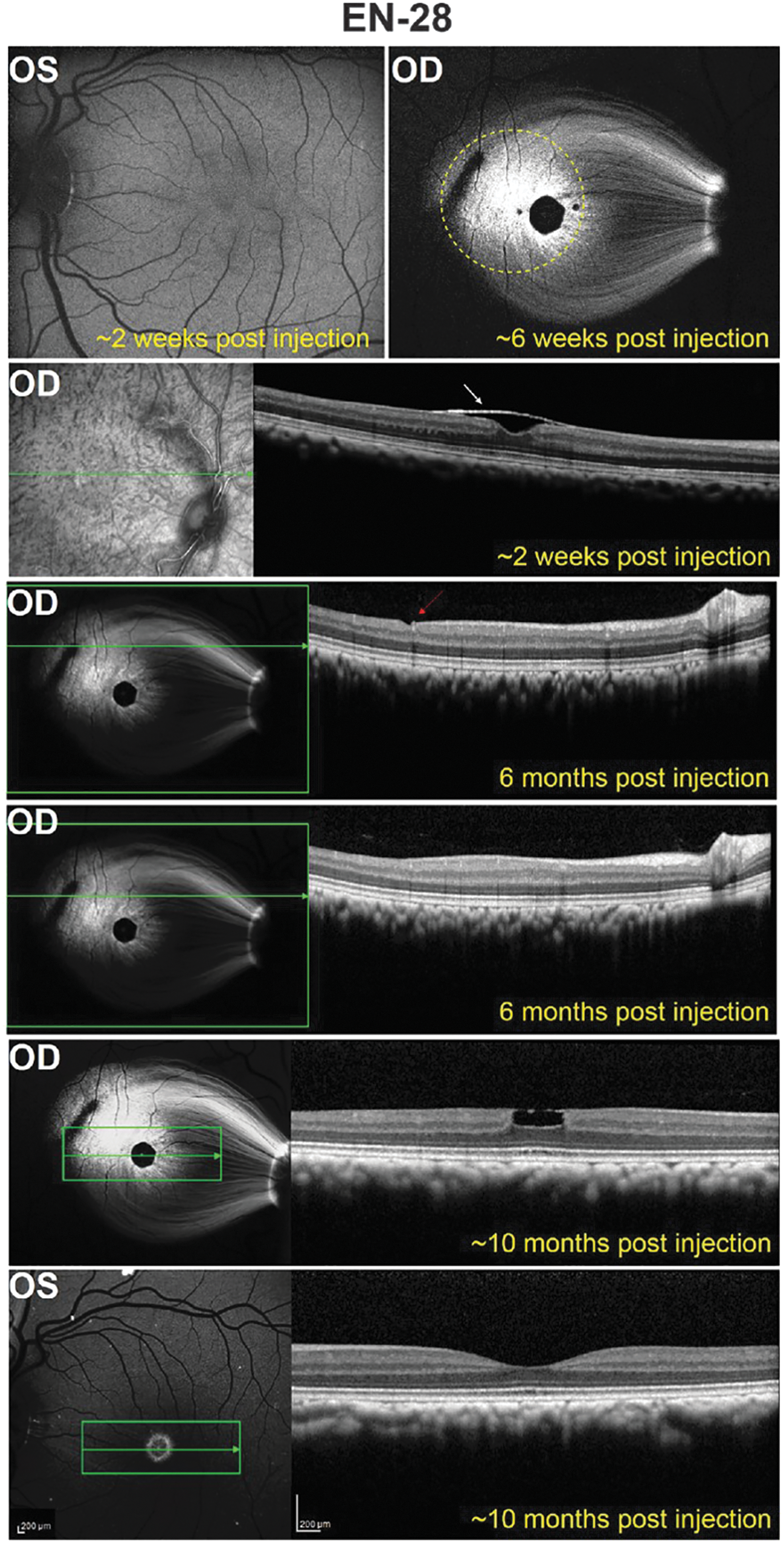
Fluorescence fundus images and corresponding OCT scans in the retina of EN-28. A representative image from ∼6 weeks postinjection shows GFP expression in a single subILM bleb in the OD. The extent of the subILM bleb created during surgery is denoted by the dashed yellow line. An OCT scan at the level indicated by the green line in the adjacent red-free image suggests that the ILM remained detached at this time point. Six months after subILM injection, an OCT scan through the injection site shows minimal cellular displacement only at the needle entry site (red arrow) and not beyond, as shown in the panel below. About 10 months after subILM injection, there was no reduction in GFP expression. However, there is evidence of a lamellar macular defect, presumably due to slight tangential traction from the ILM/epiretinal membrane. A representative fundus image and OCT scan of the intravitreally injected OS obtained at ∼10 months postinjection reveals GFP expression in the foveal ring and pit and no structural disorganization. The green arrows correspond to the location in the retina through which an OCT scan was collected (OCT scans are shown on the right in each instance). Scale bars: 200 μm.
Subject EV-44 was imaged at five time points ranging from 17 days to 15 months postinjection. GFP expression was evident in the subILM-injected OD by 2 weeks postinjection and persisted for at least 15 months (Fig. 5 and Supplementary Fig. S3). Lower relative GFP expression was noted in EV-44's OD relative to that seen in other subILM-injected eyes, a result likely owing to vector reflux during the injection procedure. Representative OCT scans revealed minimal INL loss and cellular displacement at the injection site (Fig. 5). AAV-mediated GFP expression in the foveal rim of the intravitreally injected OS of EV-44 was not evident at 2 weeks (Fig. 5) but was observed 1 month postinjection (Supplementary Fig. S3). OCT scans of EV-44 OS were also generally unremarkable.
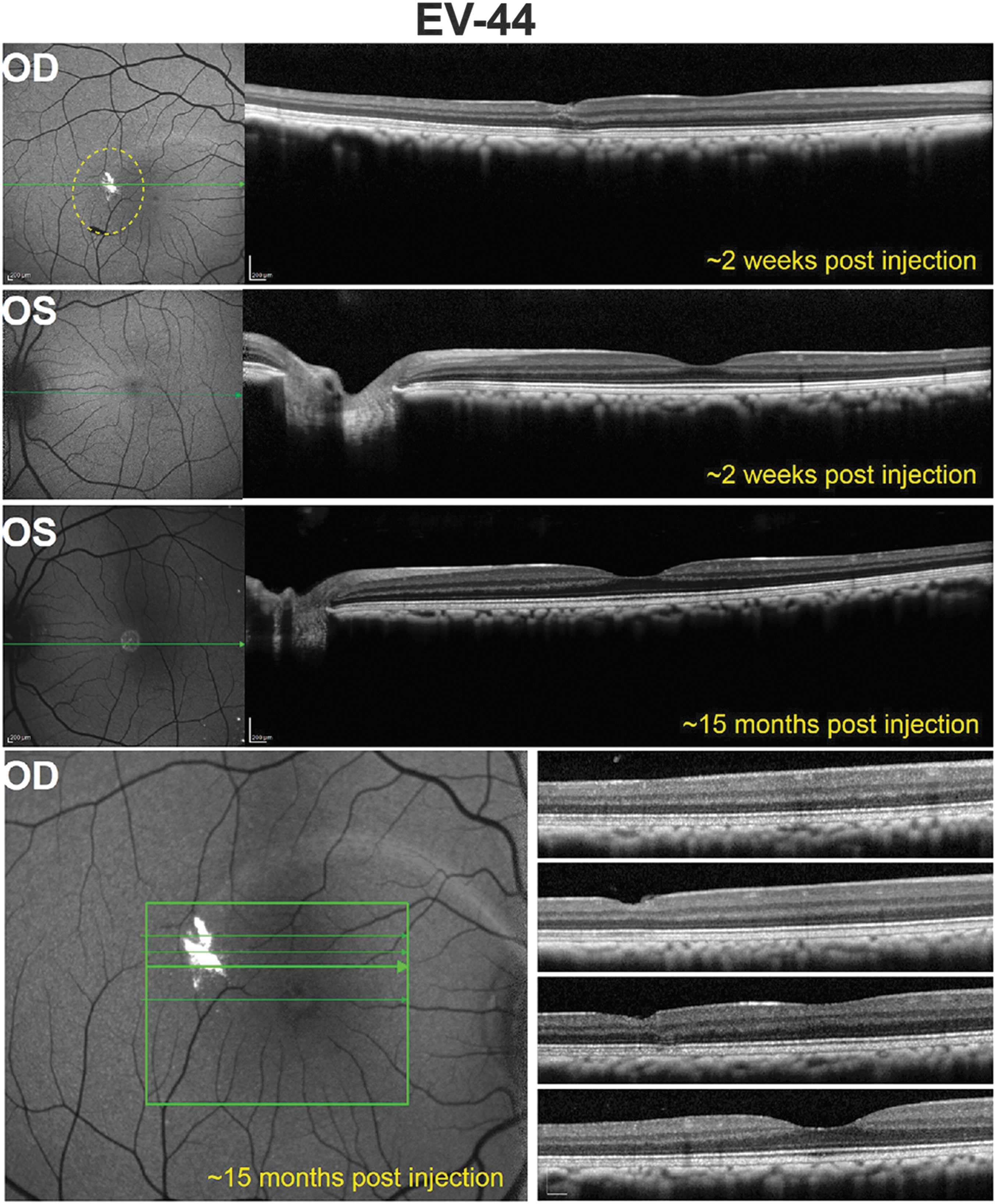
Fluorescence fundus images and corresponding OCT scans in the retina of EV-44. A representative image from the earliest postinjection time point (∼2 weeks) reveals GFP expression in a single, small subILM bleb in the OD. The extent of the subILM bleb created during surgery is denoted by the dashed yellow line. OCT scans through this region show minimal cellular displacement, and no structural abnormalities were noted elsewhere. There was no apparent diminution of GFP signal for at least ∼15 months in the subILM-injected OD. GFP expression was not detectable in a representative fundus image of the intravitreally injected OS obtained ∼2 weeks postinjection, but was apparent ∼15 months postinjection. OCT scans showed no structural abnormalities in the OS. The green arrows correspond to the location in the retina through which an OCT scan was collected (OCT scans are shown on the right in each instance). Scale bars: 200 μm.
SubILM-delivered AAV2 leads to transduction of RGCs, Müller cells, ON bipolar cells, and photoreceptors
AAV2-CBA-mediated GFP expression was next evaluated in frozen retinal cross-sections from the subILM-injected eyes of subjects F91-108 and EN-28. Both the lateral spread described previously and the depth of transduction within the fovea/macula were clear in low-magnification ( × 10) images of both subjects (Fig. 6). In F91-108 OD, most, if not all RGCs within the foveal ring were GFP positive. Expression was also seen in cell bodies within the inner and outer nuclear layers. At higher magnification (Fig. 6, right-hand images), GFP-positive foveal cone cell bodies and inner/outer segments and Müller glia were apparent in this subject (Fig. 6). Notably, the foveal ring and pit were located outside the subILM injection bleb, which, as seen in Fig. 2, was located temporal to F91-108's fovea. In subject EN-28, dense GFP expression was also observed in RGCs within the foveal ring and some expression, albeit less robust than that seen in F91-108, was observed in middle retinal cells that physically resembled Müller glia (Fig. 6). DAPI staining revealed slight displacement of INL cell bodies in the foveal pit, confirming in-life observations made by OCT.
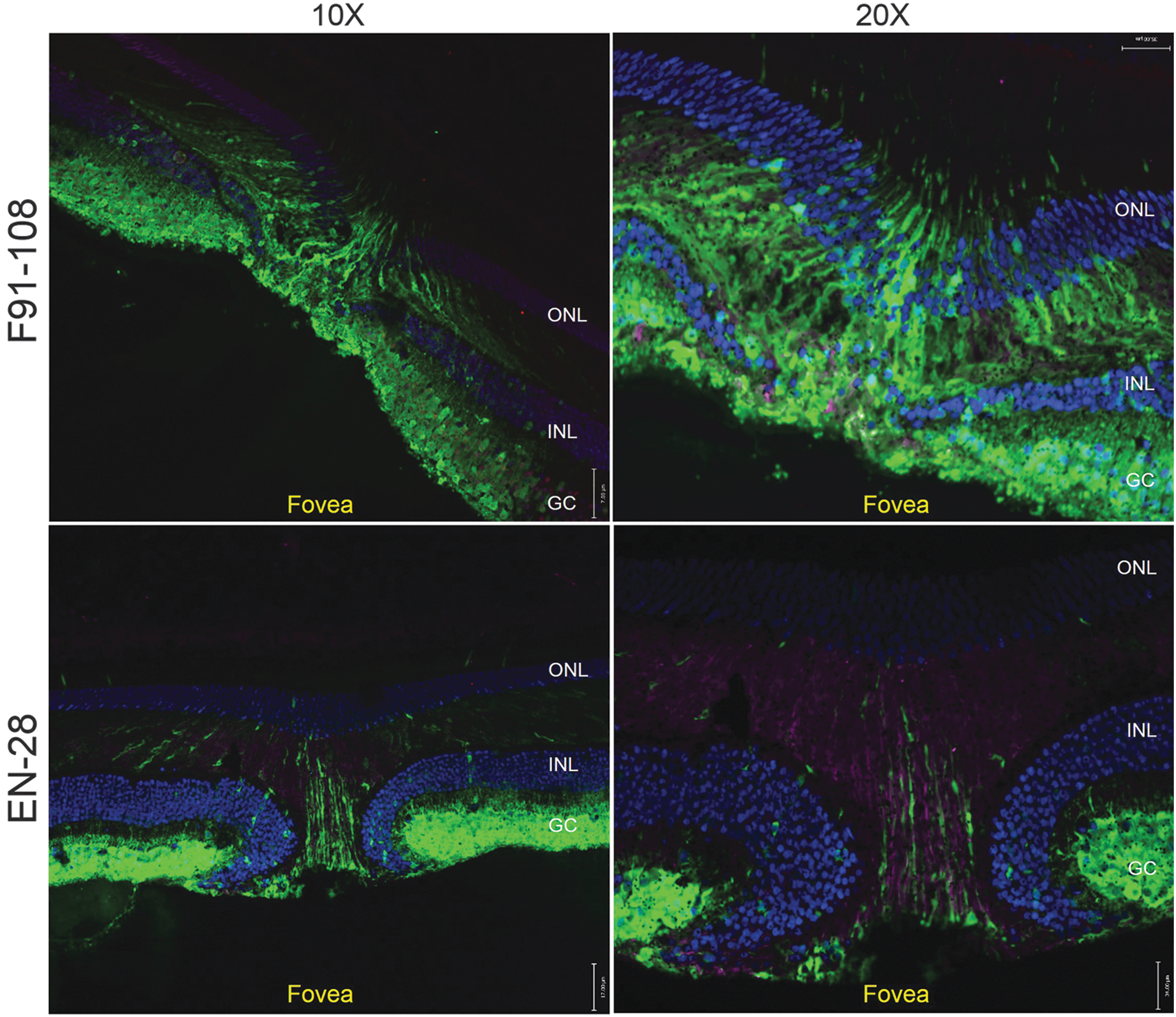
AAV2-CBA-mediated GFP expression in foveas of subILM-injected NHPs. Cross-sections of foveal pits of F91-108 OD (top) and EN-28 OD (bottom) were stained with an antibody raised against glial fibrillary acidic protein (GFAP; purple) and counterstained with 4′,6′-diamino-2-phenylindole (DAPI; blue). Confocal images taken at original magnifications of ×10 (left) and ×20 (right) revealed the extent of native GFP fluorescence (green) in retinal ganglion cells (RGCs), middle retina, and foveal cones of both animals. No reactive gliosis was observed in either retina as evidenced by a lack of GFAP staining. Scale bars at ×10 and ×20 represent 17 and 34 μm, respectively. ONL, outer nuclear layer; INL, inner nuclear layer; GC, ganglion cell layer.
Next, a detailed analysis of retinal cell types transduced by subILM-delivered AAV2 was undertaken (Fig. 7). We focused our efforts on subjects that exhibited robust in-life GFP expression (F91-109 and EN-28). With detailed documentation, we were able to localize from where in each retina our images were taken. The retinal location of each image is superimposed with the respective panel letter in associated SLO images (Fig. 7). The majority of RGCs within the main injection sites of both animals were strongly transduced (Fig. 7A, D, F, J, and K) with a depth and lateral spread greater than that seen in any previous study evaluating intravitreally injected vector in primates. 19,20 Notably, the efficiency and depth of RGC transduction near the injection site of EN-28, which fell outside the foveal ring, was equal to that achieved within the foveal rings of each animal (Fig. 7J and K). Müller glia and ON bipolar cells were strongly transduced in the fovea (Fig. 6 and Fig. 7B, C, G, and H) and, to a lesser extent, within the injection site (Fig. 7J). Transduction of outer retina was achieved predominantly in the fovea (with individual photoreceptors sparsely labeled in the injection site). Interestingly, in almost all cases where outer retinal transduction was observed, transgene expression was restricted to cones, as evidenced by the location of their cell bodies at outermost portion of the outer nuclear layer and their characteristic morphology (Fig. 6 and Fig. 7C–E and I).
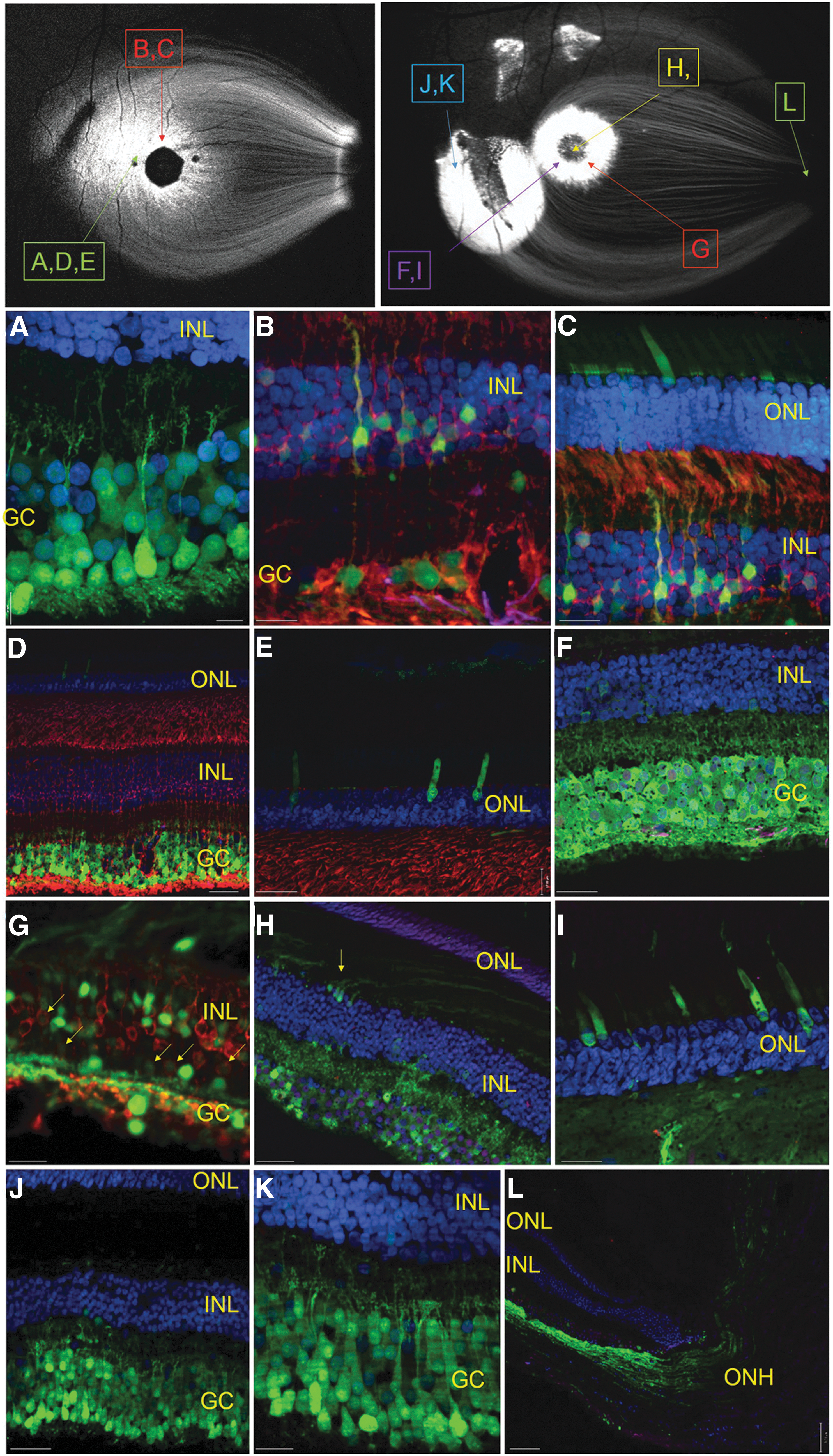
AAV2-CBA-mediated GFP expression in retinas of subILM-injected NHPs. Cross-sections of retina from the foveal ring of EN-28 OD reveal robust native GFP expression (green) in RGCs
Costaining of retinas with an antibody raised against GFAP was performed to assess whether the injection procedure, AAV capsid, or transgene expression led to activation of Müller glia. No activation was observed in the fovea of subject F91-108 (Fig. 6). Despite the displacement of inner nuclear layer cell bodies, presumably caused by macular traction during subILM surgery, no GFAP immunoreactivity was observed in EN-28 OD. To further probe whether subILM-delivered AAV promoted reactive gliosis, we compared GFAP-staining retinas from a subILM-injected subject (F91-108 OD) and an uninjected, naive control (F91-108 OS). Representative images reveal no qualitative differences in GFAP immunoreactivity between retinas of naive or subILM-injected subjects (Fig. 8) with levels akin to that previously reported in normal primate retina. 46 Staining of a retinal section taken from the injection site of F91-108 also lacked GFAP immunoreactivity (Fig. 8). Taken together, our data suggest that neither the subILM injection procedure itself nor the AAV capsid/transgene led to persistent reactive gliosis.
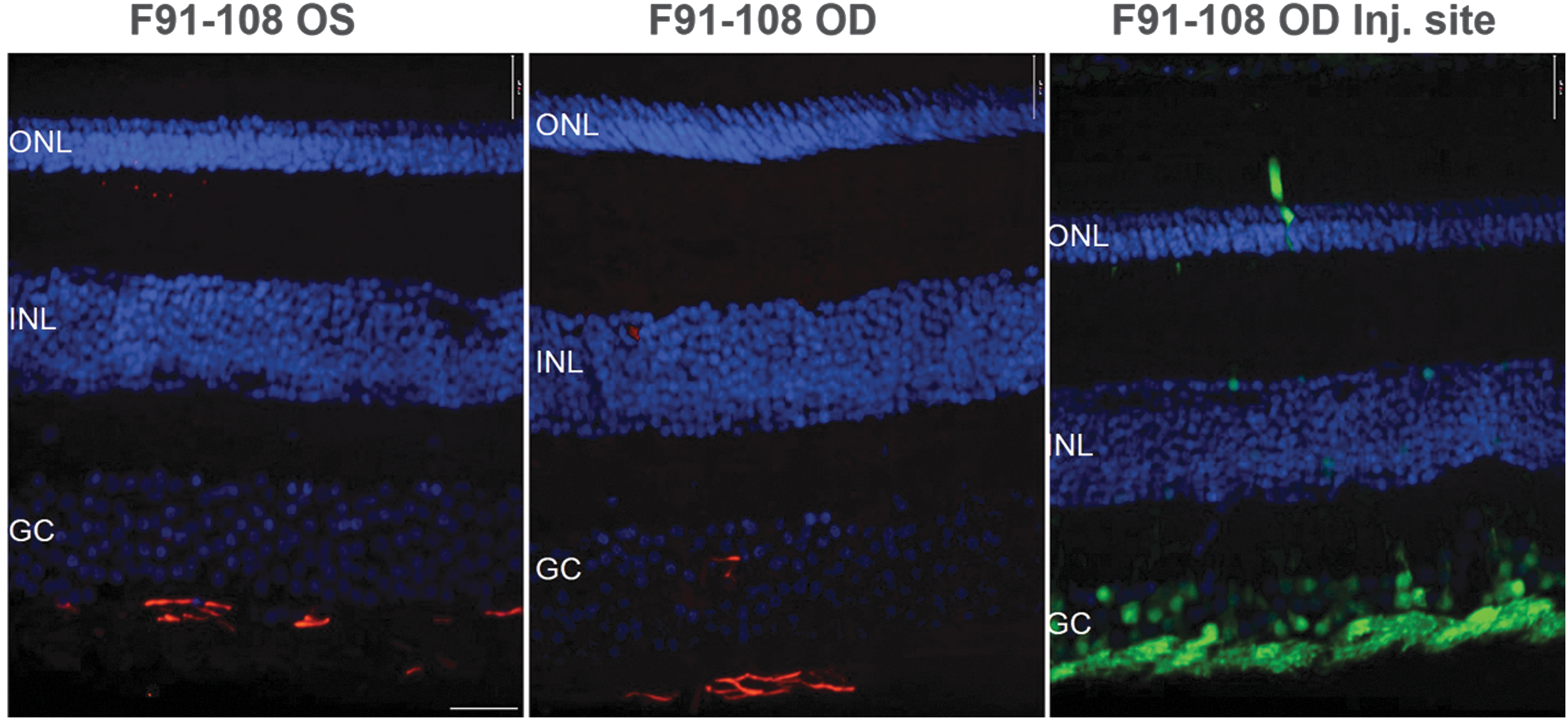
GFAP staining in subILM-injected and uninjected eyes of an NHP. Representative retinal cross-sections from F91-108 OD and OS were immunostained with an antibody raised against GFAP and counterstained with DAPI. The uninjected eye (F91-108 OS, left) had similar levels of GFAP immunoreactivity as an area outside the bleb of the subILM-injected eye (F91-108 OD, middle). There was no apparent GFAP staining immediately inside the subILM injection site of F91-108 OD (right). ONL, outer nuclear layer; INL, inner nuclear layer; GC, ganglion cell layer. Scale bar: 17 μm.
Extensive RGC projections to central targets
In both F91-108 and EN-28, without immunohistochemical staining, numerous GFP-positive RGC axons could be seen coursing through sections of the optic nerve (Fig. 9A). In addition, using an anti-GFP antibody, biotinylated secondary and streptavidin–HRP to stain sections through the various retinorecipient nuclei, we were able to visualize extremely robust central projections of GFP-expressing RGCs. Dark-field images of HRP-stained sections indicated that there were substantial RGC projections both to the core of the pretectal olivary nucleus (PON) and to the superior colliculus (Fig. 9B and C). Robust retinal projections were also observed to both the magnocellular and parvocellular layers of the dorsal lateral geniculate nucleus (dLGN) (Fig. 9D and E). A stronger ipsilateral projection to both the magnocellular and parvocellular layers of the dLGN was seen. This is consistent with subILM injections that were predominantly confined to the temporal retina. In addition, projections to the ventral lateral geniculate nucleus (vLGN) were also observed (data not shown). However, in both animals, there was negligible labeling of the suprachiasmatic nucleus (SCN) and the shell region of the PON; both of which receive substantial input from intrinsically photosensitive RGCs. 47

Fluorescence and dark-field images of GFP-positive axons and terminal fields. Representative sections from F91-108. Fluorescence image of GFP expression within retinal axons in the optic nerve
AAV2 neutralizing antibody titers increase after vector delivery
Serum from all NHP subjects was screened for the existence of neutralizing antibodies (nAbs) against AAV2 before purchase (with the exception of F91-108) and postinjection with an assay using self-complementary AAV2-smCBA-mCherry and ARPE-19 cells. FACS was done to quantify mCherry expression in cells that were infected with AAV-mCherry in the presence of serial dilutions of NHP serum. Subjects EN-28 and EV-44 exhibited mild and robust neutralization, respectively, before injection (Table 3). Strong neutralization of AAV2 was observed in postinjection serum samples from all subjects, with nAb titers of 1:2560, 1:640, and 1:2560 observed in subjects F91-108, EN-28, and EV-44, respectively. These values were obtained from serum samples collected at either 4 months (EN-28 and EV-44) or 7 months (F91-108) postinjection. Although we cannot conclude whether the nAb titer in F91-108 increased after subILM injection of AAV2-CBA-GFP (we did not collect a baseline, preinjection titer in this subject), nAb titers in subjects EN-28 and EV-44 increased 15-fold after subILM/intravitreal injection.
Neutralizing antibody titers of nonhuman primate serum pre- and postinjection
N/A, not applicable.
Discussion
Gene delivery strategies that allow efficient targeting of inner and middle retinal cells and/or enable treating photoreceptors of diseased retina without the need for invasive subretinal injection will greatly advance the retinal gene therapy field. Initial efforts to achieve this have focused on examining the transduction profile of AAV2 after noninvasive, intravitreal injection in a variety of species. A general pattern has emerged—transduction is highest in rodents, lower in larger animals such as dog, and even lower in primates (macaques). Intravitreal AAV2 promotes pan-retinal transduction of RGCs (and some Müller glia) in adult rodents. 34,48,49 In dog, AAV2 transduces RGCs and Müller glia throughout the retina with low efficiency, and with higher efficiency in regions surrounding large blood vessels. 50 In macaques, AAV2 transduction is restricted primarily to the inner retina within the foveal ring. 19 Our own experience in this study, using the same vector (AAV2-CBA-GFP) delivered by intravitreal injection, was consistent with that previously seen in macaques. 19
The ILM is a basement membrane with attached extracellular matrix that separates the vitreous from the neural retina and is a formidable barrier to AAV vector transduction. 34 Attempts to increase transduction by intravitreally delivered AAV have been made by enzymatically digesting the ILM in rodents and primates. 19,34,36 In rodents, the extent of transduction was improved after application of proteases. 34 In macaques, a similar approach involving pretreatment (5 days before vector injection) with intravitreal microplasmin, a serine protease used to degrade the vitreoretinal junction in the human eye, led to expression from AAV vectors previously unable to mediate expression when delivered by intravitreal injection alone. However, there was no increase in the area of transduction, or of the cell types expressing transgene after enzymatic digestion. Expression remained restricted to the central foveal ring. 19
Failure of first-generation AAV2 to promote widespread transduction of NHP retina after intravitreal injection prompted efforts by multiple laboratories to engineer novel AAV capsid variants through both directed evolution and rational design. Significant improvements in transduction of rodents were achieved with two variants in particular, AAV2-7m8 and AAV2(quadY-F+T-V), capable of transducing all layers of the rodent retina including photoreceptors. 20,31,49,51 Although improved in terms of magnitude of expression in primates, these variants still produced the restrictive pattern of expression limited to the foveal area and large blood vessels (see Ref. 20, and S.E. Boye and P.D. Gamlin, unpublished data).
In the current study, we explored how physical circumvention of the ILM would alter the magnitude and pattern of retinal transduction relative to that seen with intravitreally injected AAV2 in macaques. To do so, we created a surgically induced, hydrodissected space between the ILM and neural retina (i.e., subILM) where vector was sequestered. Our goal was to bypass three major barriers associated with intravitreal injection—dilution of vector in the vitreous humor, exposure to neutralizing antibodies, and the ILM itself. Surgical manipulation of the ILM is routinely performed in humans to correct macular holes, macular puckers, and myopic foveoschisis. 39 –41 The ILM can be surgical minimized (i.e., “peeled”) and AAV immediately delivered to the vitreous, but this leaves the neural retina exposed to the immunological environment of the vitreous, which has just been exposed to antigen (AAV capsid). Such an approach can lead to inflammation in NHP retina. 52 For this reason we chose to pursue subILM delivery of AAV. Before initiating in vivo experiments we first sought to determine compatibility of AAV with Healon. AAVs use glycans as primary receptors for cellular recognition and internalization, and the hyaluronic acid in Healon is a glycosoaminoglycan (GAG). 53 We therefore preincubated AAV2 with Healon and then performed an in vitro transduction assay in 661W cone-derived photoreceptor cells. Results indicated Healon did not negatively impact transduction. Our first attempt at AAV delivery under the ILM involved creating a virtual space between the ILM and neural retina (in animal F91-108) via injection of Healon alone. Next, a fraction of Healon was irrigated from the subILM injection blebs with BSS, and AAV vector was subsequently delivered through the same injection site. From this initial surgery, we learned that complete posterior vitreous detachment was not required to create this virtual space and, if anything, is contraindicated because of the increased potential for corneal haze during surgery. Repeated manipulation of the subILM injection site (with Healon, BSS, and finally AAV) in F91-108 enlarged the entry points, thereby increasing the potential for vector reflux into the vitreous. Last, more control over delivery rate was desired to more accurately quantify vector dose within the subILM bleb. Despite this, in-life imaging 3 months postinjection revealed robust and extensive transduction that far surpassed what had been observed after intravitreal vector injection, especially when taking into account the limited volume and vector genomes delivered (∼1.66 × 1010 VG in ∼10 μl). Transgene expression persisted for at least 16 months with no apparent diminution. The in vitro transduction of the Healon–AAV mixture indicated that Healon can function well as a vehicle for AAV2. Therefore in the second animal (EN-28), we opted to premix Healon and AAV at a 1:1 volumetric ratio to deliver them simultaneously, thus allowing for only one subILM injection. This led to the creation of a clean subILM bleb and no reflux. We also incorporated a Nanomite injection device that allowed for accurate quantification of the injection rate. The lamellar macular defect noted in EN-28 appears to be a macular pseudohole, likely due to mild ILM/epiretinal membrane traction. 54,55 Similar defects in humans are not uncommon and rarely affect vision. They rarely progress to a true macular hole and may resolve without requiring surgery. 54,55 This was the only eye in which the fovea was directly involved at the time of injection. This preliminary finding supports further investigation into whether detachment of the ILM specifically over the fovea consistently leads to similar structural changes. Again, as in the first animal, we observed robust and extensive transgene expression from subILM vector delivery as early as 6 weeks postinjection that persisted for the duration of the in-life period (10 months). In this case we also delivered the same vector by intravitreal injection in the contralateral eye, which resulted in weak expression limited to the foveal ring, consistent with previous reports of transgene expression after intravitreal delivery of AAV2 in macaques. In the third animal (EV-44), significant reflux of Healon–vector occurred during the subILM injection. Consistent with this, transduction, although apparent at the site of injection, was clearly reduced relative to what was seen in the first two animals. Nevertheless, it persisted for at least 15 months postinjection. In addition, this animal had a relatively high preexisting AAV2 nAb titer (1:160), but we believe that this was not the basis for the poor transduction after subILM injection because the contralateral, intravitreally injected eye showed expression consistent with that observed after intravitreal injection in EN-28. At this point, we cannot determine whether a subILM injection alone (of Healon–AAV) without reflux results in an increase in nAbs because the second and third animals (EN-28 and EV-44) both received intravitreal injections of AAV2 in their left eyes, which in and of itself leads to an increase in the formation of neutralizing antibodies. 19,33 A preinjection nAb titer was not established for the first animal (F91-108) as a serum sample was not available. Further, because of the methodology employed, the entry site was enlarged and we suspect a significant fraction of the AAV solution refluxed into the vitreous. What can be concluded is that persistent transgene expression was seen after subILM delivery (as long at 16 months postinjection) despite moderate to high preexisting nAb titers in EN-28 and EV-44 (1:40 and 1:160, respectively). This is significant as the presence of nAbs to AAV often appears to result in weak, decaying, or absent transgene expression for intravitreally delivered AAV in primates. 33
Careful analysis of retinal sections taken from within the subILM bleb and foveal ring of F91-108 revealed transduction of almost all RGCs, with clearly visible GFP-positive dendrites extending into both ON and OFF layers of the inner plexiform layer. Other transduced cell types within this region included Müller glia, cone photoreceptors, and ON bipolar cells. Within the subILM injection bleb of EN-28 (located outside the foveal ring), similarly robust RGC transduction was observed. Sporadic transduction of Müller glia and cone photoreceptors was also noted in this region. The increased area and depth of transduction achieved by subILM-delivered AAV2 was for the most part restricted to the area of retina immediately within the subILM bleb except for the fovea of animal F91-108. The reason for this is not fully understood. It can be seen from the in-life image that the two large areas of expression share a “bridge.” It may have been that vector spread from the point of injection through a connecting channel to overlay the fovea.
Extensive analysis of the RGC projections to the retinorecipient nuclei in both F91-108 and EN-28 demonstrates that the GFP-expressing RGCs are fundamentally healthy and maintain robust central projections. We visualized strong retinal projections to both the magnocellular and parvocellular layers of the dorsal lateral geniculate nucleus (dLGN). Because the dLGN is the major relay of conscious visual information to the visual cortex, the proposed subILM injection approach would therefore be expected to be a good solution for studies in which optogenetic molecules such as Channelrhodopsin-2 or its derivatives are expressed in RGCs of primates including humans, especially so when a light-driven retinal prosthesis is implemented. 56 –58 In addition, we saw strong projections to both the pretectal olivary nucleus and the superior colliculus, which strongly suggests that RGCs other than midget and parasol cells are GFP-expressing and project to these additional retinorecipient target structures. 59 Therefore, if specific RGC-specific promoters can be identified in primates including humans, this approach should allow one to selectively target any of the various primate RGC classes for basic, translational, or clinical studies. 59 Interestingly, we failed to observe significant GFP-expressing axons in either the suprachiasmatic nucleus (SCN) or shell of the pretectal olivary nucleus (PON). These two regions receive substantial input from intrinsically photosensitive retinal ganglion cells (ipRGCs), but the specific class or classes of ipRGC underlying this projection in primates are currently unknown. 47 However, it is known that many outer stratifying ipRGCs (M1 homologous cells) are displaced to the inner nuclear layer, and it is likely that our subILM-delivered vector did not efficiently transduce these displaced ipRGCs. 60,61 Further, in rodents, M1 cells project to the SCN and the shell of the PON to mediate circadian photoentrainment (SCN) and the pupillary light reflex (PON). 62,63 Therefore, it is tempting to speculate that in primates, the outer stratifying ipRGCs (M1 homologous cells) account for much of the projection to the SCN and PON. Retrograde studies in future experiments will be required to confirm this hypothesis.
Our results confirm that the ILM is a major barrier to transduction by AAV and that, when it is circumvented, the efficiency and depth of transduction by AAV2 are greatly enhanced. Future studies using the subILM approach will be focused on addressing several questions. First, can capsids that mediate efficient transduction of primate photoreceptors when subretinally injected, namely AAV5 and AAV8, reach and then transduce photoreceptors if placed under the ILM? 13,15 Experiments done in rodents, where the ILM was physically digested or was ablated via genetic manipulation followed by intravitreal delivery of AAV5, resulted in efficient transduction of outer retina. 34,37 This would suggest that AAV5 may be capable of trafficking to the outer retina when placed under the ILM. However, substantial shifts in vector tropism have been observed when the biology of the retina has been substantially perturbed, as would be the case in both of the aforementioned treatments. Second, how will enhanced AAV2-based capsid mutants that promote improved transduction after intravitreal injection [AAV2-7m8 and AAV2(quadY-F+T-V)] perform when placed under the ILM? Interaction of the capsid with heparan sulfate proteoglycan (HSPG), as shown by binding affinity of the capsid to a heparin column, is a strict requirement for intravitreally mediated transduction when the ILM is intact. 64 Interestingly, these two capsid mutants retain the ability to bind heparin, but with lower affinity than wild-type AAV2. 20,64 It has been hypothesized that the improved transduction is partially due to the fact that these variants interact with the ILM via HSPG but do not become sequestered there and thus can traffic further into the retina. These two capsids have already been shown to have tropism for Müller glia and ON BCs and, therefore, they will also be interesting candidates for testing via this route. In addition to histological analyses, an investigation of the functional consequences of subILM injections is also warranted. Multifocal electroretinograms 65 will be used in future studies to evaluate ERG responses simultaneously from various regions of the retina (within and outside injection blebs). In so doing, we will determine whether any deleterious functional effects result from the subILM surgery itself.
Footnotes
Acknowledgments
This study was supported by grants from the National Institutes of Health (R01EY024280 to S.E.B.; R01EY025555 to P.D.G), core grants to the University of Florida (P30EY021721) and the University of Alabama at Birmingham (P30EY03039), a UAB CCTS Pilot Grant (5UL1 RR025777), the Eyesight Foundation of Alabama, the Foundation Fighting Blindness, the Macula Vision Research Foundation, and unrestricted funding from the Research to Prevent Blindness to the University of Florida Department of Ophthalmology. The authors thank Dr. Stephen Breaud for valuable technical assistance and the Retinal Gene Therapy Vector Core at the University of Florida for packaging AAV vectors used in this study.
Author Disclosure
W.W.H. and the University of Florida have a financial interest in the use of AAV therapies and own equity in a company (AGTC, Inc.) that might, in the future, commercialize AAV vectors for the treatment of blindness. S.E.B., J.J.A., C.D.W., S.L.B., J.J.P., M.E.C., K.J.S., C.A.G., and P.D.G. have no conflicts to report.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.