
Abstract
Ischemia–reperfusion (I/R) injury remains the primary contributor to delayed graft function in kidney transplantation. The beneficial application of manganese superoxide dismutase (sod), delivered by a BacMam vector, against renal I/R injury has not been evaluated previously. Therefore, this study overexpressed sod-2 in proximal tubular epithelial (HK-2) cells and porcine kidney organs during simulated renal I/R injury. Incubation of HK-2 cells with antimycin A and 2-deoxyglucose resulted in a significant decrease in intracellular adenosine triphosphate (ATP) levels; following reperfusion, ATP levels significantly increased over time in cells overexpressing sod-2. In addition, lactate dehydrogenase (LDH) release declined over 72 h in BacMam-transduced injured cells. Ex vivo delivery of sod-2 significantly increased ATP levels in organs after 24 h of cold perfusion. In vitro and ex vivo results suggested that BacMam transduction successfully delivered sod-2, which reduced injury associated with I/R, by improving ATP cell content and decreasing LDH release with a subsequent increase in kidney tissue viability. These data provide further evidence for the potential application of BacMam as a gene delivery system for attenuating injury after cold preservation.
Introduction
I
I/R injury remains a major cause of acute kidney injury (AKI), resulting in high levels of morbidity and mortality, 4 and it is a major determinant of graft survival in kidney transplantation. 5 Two types of ischemia may occur during renal transplantation: warm and cold ischemia. Warm ischemia is further divided into donor warm ischemia (either after circulatory death or after brain death) and recipient warm ischemia. Cold ischemia occurs when kidneys collected from living donors are directly placed in cold preservation solution. The work described in this study sought to examine the possibility of reducing injury associated with cold preservation.
Emerging strategies have tried to prevent renal I/R injury by preserving ATP levels in an attempt to improve short- and long-term outcomes following kidney transplantation. 6 –9 In vitro models are generally used to study molecular mechanisms, specific pathways involved in cellular damage, or death caused by I/R injury events. They provide evidence for further in vivo analysis. However, as cell-based experimental models mimic only some of the features of tissue ischemia and I/R injury, they cannot reproduce the in vivo tissue environment completely. 10 The major areas of in vitro I/R injury investigation have included overexpression of protective genes, enhancing enzyme activity, and suppressing the expression of genes that activate the cell death pathway. Because low blood supply during I/R injury leads to the deprival of ATP concentration, most cell-based models of I/R have focused on inhibiting the production or synthesis of ATP through chemical, 11 enzymatic, 12 or anaerobic chamber 13 approaches. 14 The present study used a chemical approach to induce I/R injury in an immortalized proximal tubule epithelial cell line (HK-2) from normal adult human kidney.
Mitochondria are the main cellular sites of radical oxygen species production. Consequently, they represent an early target of oxygen injury via enhanced production of ROS, followed by mitochondrial dysfunction. 15 Manganese superoxide dismutase (SOD-2) is the major antioxidant in the mitochondria. It localizes in the mitochondrial matrix 16 and exists as a homotetramer containing one Mn (Manganese) atom per subunit. 8 Its main function is to catalyze the dismutation of the superoxide anion (O2 •–) producing hydrogen peroxide (H2O2). 8 Using a rodent model of renal I/R injury, studies have shown that endogenous SOD-2 was rapidly inactivated early during renal transplantation, suggesting that loss of SOD-2 activity may have played an important role in inducing further injury 17 –19 (e.g., SOD-2 deficiency in mice leads to prenatal lethality). 15 Furthermore, a number of reports have shown that high levels of SOD-2 play a key role in attenuating injury or death under conditions of enhanced oxidative stress. 20 –23
To date, >70% of clinical trials and gene therapy studies have been carried out using mammalian virus vectors such as adenovirus. 24,25 BacMam vectors (baculovirus-based vectors for expression in mammalian cells) provide an attractive alternative allowing scalable virus production. 26
A number of studies have reported that the mode and kinetics of BacMam entry into mammalian cells is identical to that found in insect cells. 27,28 Baculovirus enters insect cells by receptor-mediated endocytosis, followed by pH-dependent fusion of the envelope with the endosome. 27,28 BacMam vectors also enter mammalian cells via endocytosis and are released into the cytoplasm by acid-induced fusion of the envelope with the endosomal membrane. Studies have suggested that the rod-shaped nucleocapsid, containing the virus genome, is then transported into the nucleus through a nuclear pore. 29 The prototype baculovirus used for BacMam studies is Autographa californica multiple nucleopolyhedrovirus (AcMNPV). 22 High-levels of expression have been observed in transduced vertebrate cells, and these vectors have low cytotoxicity, high insert size capacity, and a relatively low immune-response profile compared with mammalian virus vectors. 23,26,30 –33 A number of studies have reported that baculovirus stimulates an immune response in mammalian cells. It was demonstrated that baculovirus possess an adjuvant activity that promotes humoral and cellular immune responses against co-administered antigens, maturation of dendritic cells (DCs), and production of pro-inflammatory cytokines. 34 –38 Indeed, previous studies have suggested that BacMam can regulate the cellular immune response by upregulating a number of cytokines and other immune-response genes after in vitro transduction into HEK293 cells. However, it should be noted that no apparent cellular injury or reduction in cell viability was observed after in vitro and ex vivo solid organ transduction. 39 Therefore, adaptive and innate immunity should be taken into consideration when designing the BacMam gene delivery approach.
The purpose of this study was to investigate BacMam as a novel gene delivery vector for the prevention of acute cell damage caused by ischemia. In particular, experiments were designed to examine the potential application of BacMam in delivering sod-2 using an in vitro model that closely mimicked renal I/R injury. The study also evaluated whether ex vivo BacMam delivery of sod-2 in porcine kidneys might improve I/R induced renal injury during cold preservation.
Materials and Methods
Materials
Antimycin A and 2-deoxy-D-glucose (2-deoxyglucose) were purchased from Sigma–Aldrich. Phosphate-buffered saline (PBS) was supplied by GIBCO (Life Technologies Ltd.). Fetal bovine serum (FBS) was supplied by Lonza Group Ltd. and was heat inactivated at 56°C for 30 min.
Cell culture
Insect cells were maintained at 28°C in Erlermeyer flasks (Corning) and glass Spinner flasks, respectively. Sf-9 cells were passaged before reaching their stationary phase using Sf9-S2 medium (PAA Laboratories Ltd.); Sf-21 cells were maintained in TC100 (GIBCO, Life Technologies Ltd.) supplemented with 10% FBS. HK-2 cells were purchased from American Type Culture Collection and cultured in Keratinocyte serum-free medium (KSFM; GIBCO, Life Technologies Ltd.) supplemented with 2% antibiotics (100 IU/mL of penicillin and 100 μg/mL of streptomycin) at 37°C 5% CO2.
Plasmid construction and generation of BacMam vector
The pCI-neo plasmid, carrying the human cytomegalovirus immediate-early enhancer (CMV-IE) promoter, was kindly provided by Prof Bermudez-Diaz (Oxford Brookes University, Oxford, United Kingdom). The pMA_SOD-2 plasmid was obtained from GeneArt® (Life Technologies Ltd.). The sod-2 gene was synthesized based on the sequence of GenBank (accession number: Y00985). The baculovirus-based pBAC-1 plasmid was purchased from Novagen (EMD Biosciences, Inc.).
To generate a pBAC-1 plasmid containing a CMV-IE promoter, the CMV-IE region was excised from pCI-neo with restriction endonucleases BglII and XhoI. The CMV-IE promoter fragment was then ligated into the linearized pBAC-1 DNA, generating a pBAC-1_CMV transfer plasmid. Polymerase chain reaction (PCR) primers were designed for sod-2; (1) forward primer 5′-AATTGAATTCGCACCATGTTGAGCCGGG-3′, and (2) reverse primer 3′-GTACCGAACGTTTTTCATTAGATCTTAAT-5′. The EcoRI (GAATTC) restriction site was added to the 5′ end of the forward primer, while the XbaI (AGATCT) and terminator codon (TAA) were added to the 5′ end of the reverse primer. The sod-2 fragment was amplified from the pMA_SOD-2 plasmid by PCR using forward and reverse primers. The amplified sod-2 fragment and pBAC-1_CMV plasmid were digested simultaneously with EcoRI and XbaI and then ligated to create pBAC-1_SOD-2.
BacMam viruses were generated by homologous recombination between flashBAC ULTRA™ DNA (Oxford Expression Technologies Ltd.) and pBAC-1_SOD-2 plasmid DNA, according to the manufacturer's instructions. The virus containing supernatant, AcCMV_SOD-2, was then harvested 5 days post transfection and stored at 4°C. FBS (5% v/v) was added to the inoculum to improve stability during storage. High titer BacMam stock was obtained following further amplification of the viral stock in Sf-9 cells. AcCMV_SOD-2 virus was titrated by plaque assay. 40
Porcine kidney
Experiments were performed in accordance with the principles of the Oxford Brookes University Ethics Committee. Porcine kidneys were obtained from a local abattoir. Organs were collected within 10 min after slaughter (22-week-old large white pigs) and flushed in situ with soltran kidney perfusion fluid (0.86% w/v potassium citrate, 0.82% w/v sodium citrate, 3.38% w/v mannitol, and 1% w/v magnesium sulfate; Baxter). Kidneys were immediately transported in a biohazard bag on ice (for approximately 45 min) to the laboratory for analysis.
In vitro viral delivery
HK-2 cells were seeded at a concentration of 0.3 × 106 cells/mL and incubated overnight at 37°C in 5% CO2. The following day, cells were transduced with BacMam viruses at a multiplicity of infection (MOI) of 150 pfu/cell and incubated at 37°C for 1 h in 5% CO2. The virus inoculum was then replaced with growth culture medium, and cells were returned to 37°C in 5% CO2 for the required time.
In vitro I/R injury model
HK-2 cells were incubated in PBS containing 10 μM of antimycin A (a complex III inhibitor of mitochondrial electron transport) and 10 mM of 2-deoxyglucose (a nonmetabolizable isomer of L-glucose and a glycolysis inhibitor) for 60 min to induce in vitro ischemia injury. 41 Reperfusion was achieved in HK-2 cells by washing with PBS and then culturing in complete growth medium.
Ex vivo viral delivery
Flushed kidney organs were cannulated and perfused through the renal artery with 200 mL of cold Belzer UW solution (Bridge to Life [Europe] Ltd.), with or without BacMam. BacMam was concentrated using low-speed centrifugation, and the pellet was dissolved in a physiological saline solution. During transduction, the 200 mL of Belzer UW solution was collected and recirculated using a low-speed peristaltic pump (10 rpm; Watson Marlow 505 S, Gemini BV). Kidneys were transduced for 24 h. Tissue samples were collected using a 10 mL syringe at 0, 4, and 24 h post transduction (hpt), and cells were immediately dissociated using collagenase (Sigma–Aldrich).
Measurement of cellular injury (ATP assay)
Intracellular ATP levels were measured using a CellTitre-Glo® Luminescent Cell Viability Assay (Promega). Initially, the assay buffer and the lyophilized substrate were mixed together and left to equilibrate at room temperature for 20 min. During the equilibration time, cells to be analyzed were harvested by centrifugation and re-suspended in 100 μL of 1 × PBS. Subsequently, 100 μL of the assay reagent was added into each tube and then mixed to release intracellular ATP into solution. After 5 min incubation, luminescence was read on a GloMax® 20/20 Luminometer (Promega). Data were normalized against the live cell number obtained using a trypan blue exclusion assay.
Measurement of cytotoxicity (LDH assay)
Cell viability was assessed by measuring the release of LDH into the culture media using CytoTox-ONE Homogeneous Membrane Integrity Assay (Promega). The CytoTox-ONE™ assay was carried out according to the manufacturer's guidelines. Briefly, mammalian cells to be evaluated were seeded into individual wells of a 96-well plate (Corning), treated with the appropriate chemical, and incubated at 37°C for the required time. Lysis solution (2 μL; supplied by the manufacturer) was added to the untreated cells for maximal LDH release. Untreated cells were used as negative control, and a blank well served as a vehicle control. The cells were removed at 37°C and equilibrated at 22°C for 20 min. CytoTox-ONE reagent was then added in equal volume to the cell culture medium present in each well and shaken gently for 30 sec. Cells were incubated at 22°C for 10 min, and the reaction was stopped by adding 50 μL of stopping solution to each well. Fluorescence was recorded at an excitation wavelength of 560 nm and an emission wavelength of 590 nm using an Infinite® 200pro microplate reader (Tecan Group Ltd.). Cytotoxicity values were obtained using the following calculation:
Western blot analysis
Proteins separated on 12% SDS-PAGE gels were transferred onto a nitrocellulose membrane (GE Healthcare) and analyzed by Western blot. Protein gels were incubated in transfer buffer (48 mM of Tris, 39 mM of glycine, 20% methanol, and 1.3 mM of SDS) for 5 min, together with the nitrocellulose membrane and filter papers. Following the transfer, membranes were incubated in 5% powdered milk dissolved in PBST (1 × PBS containing 0.1% [v/v] of Tween-20) and agitated at room temperature for 1 h to prevent non-specific binding. Subsequently, membranes were treated with SOD-2 primary antibody for 60 min. Membranes were washed three times with PBST and incubated with a secondary antibody conjugated with alkaline phosphatase for 60 min. To remove any background, membranes were washed three times with PBST. Bound proteins were visualized using alkaline phosphatase buffer (100 mM of Tris-HCl [pH 9.5], 100 mM of NaCl, and 5 mM of MgCl) containing 66 μL of Nitro Blue tetrazolium (NBT) and 33 μL of 5-Bromo-4-Chloro-3′-Indolyphosphate p-Toluidine (BCIP). The reaction was stopped by washing the membrane with in distilled water, and any excess liquid was removed using filter paper. Band densitometry was carried out using GelQuantNET software (BiochemLabSolutions).
Confocal microscopy
Deparaffinized and hydrated kidney sections were permeabilized with 1 × PBS containing 0.1% (v/v) Triton X-100. Tissue sections were blocked in 1 × PBS-BSA prior to being incubated in the appropriate primary antibodies for 60 min at room temperature. Slides were washed three times in 1 × PBS and incubated with the indicated secondary antibodies for 60 min at room temperature. Finally, any unbound antibody was removed using PBS, and sections were mounted with Vectashield (Vector Laboratories), a mounting media containing the nuclear label 4′,6-diamidino-2-phenylindole (DAPI). A clear nail varnish was used to seal the mounted samples before being stored in the dark at 4°C.
Confocal microscopy was performed on fixed tissue sections using a Zeiss LSM 510 Meta confocal system using a × 63 oil immersion lens. Multitrack analysis was carried out in order to analyze green fluorescence (488 nm excitation, band pass filter 505–530 nm), red and blue (DAPI) fluorescence (543 nm excitation, long pass filter 560 nm for red, and 405 excitation, band pass filter 420–480 nm for blue). Image data were analyzed using LSM Image browser software.
Statistical analysis
All data were analyzed using GraphPad Prism for Windows v5 (GraphPad Software, Inc.) with results displayed as the mean ± standard error of the mean. Statistical analysis was performed using both one- and two-way analysis of variance with a Bonferroni post test that compared replicative mean values, unless otherwise stated. A p-value of <0.05 was considered statistically significant.
Results
BacMam-mediated sod-2 expression in HK-2 cells
Initial studies were performed to evaluate the relative BacMam transduction efficiency and the optimal transgene expression levels in human proximal tubule cells. HK-2 cells were transduced using different MOI with AcCMV_SOD-2 BacMam over a period of 96 h. Results indicated that the viral dose played an important role in efficient transduction of HK-2 cells. MOI 150 resulted in minimal cytotoxicity and was therefore chosen as optimal for HK-2 cells (data not shown). Preliminary experiments showed that SOD-2 gradually increased from 0 to 96 h following transduction with BacMam, as confirmed by Western blot analysis (Fig. 1). This was further confirmed by densitometry analysis, which indicated that SOD-2 expression increased from <1% to 11% within 24 h and subsequently to 23%, 32%, and 34% at 48, 72, and 96 hpt, respectively (Fig. 1). Because SOD-2 was optimally expressed between 72 and 96 hpt, the 72 hpt time point was chosen to assess the effect of BacMam-mediated gene delivery in an in vitro I/R injury model in HK-2 cells.
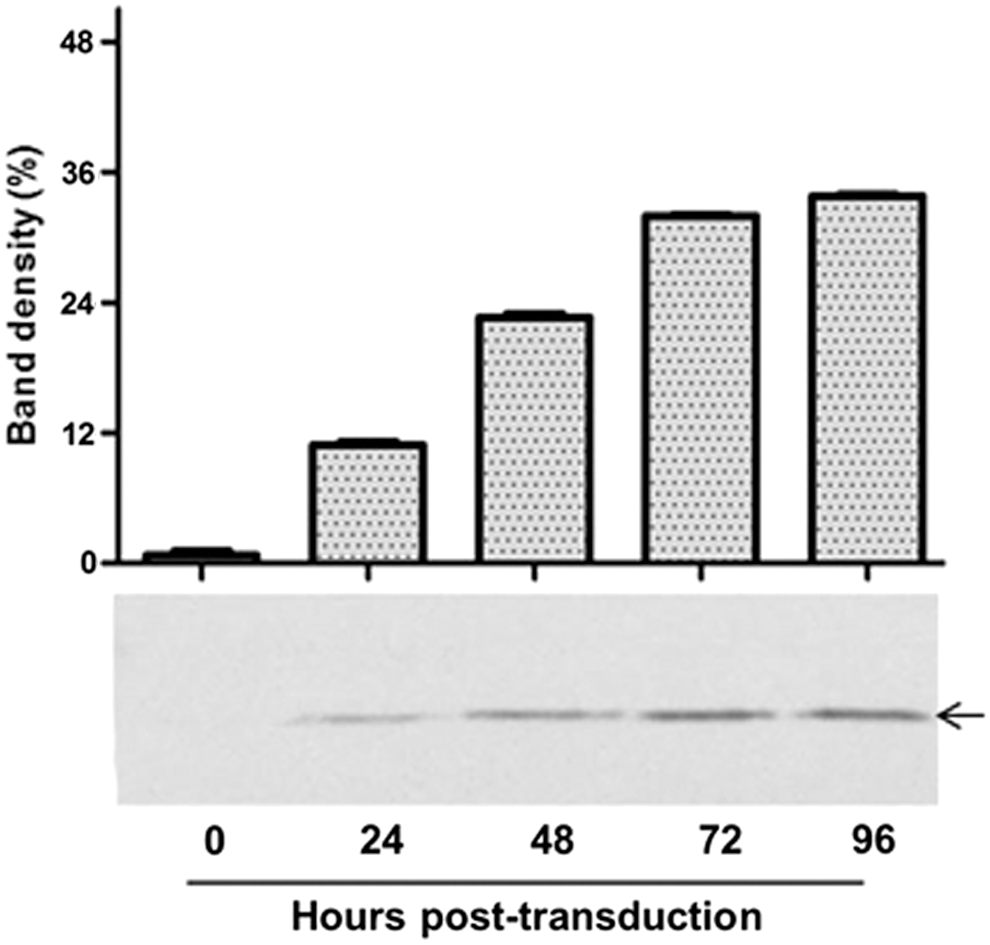
MnSOD expression in HK-2 cells. BacMam virus containing MnSOD gene under control of the cytomegalovirus immediate early (CMV-IE) promoter, was used to transduce HK-2 cells at a multiplicity of infection (MOI) of 150. Transduced HK-2 cells were harvested at 0, 24, 48, 72, and 96 h, and cell lysates were Western blotted onto nitrocellulose membrane using anti-MnSOD antibody. Lanes 0–96 represent time post transduction. The arrow refers to MnSOD proteins at approximately 23 kDa.
Antimycin A and 2-Deoxyglucose/ATP-depletion injury in HK-2 cells
To investigate the effects of I/R on renal cell injury, HK-2 cells were treated with antimycin A (10 μM) and 2-deoxyglucose (10 mM) to mimic the in vivo I/R response. As shown in Fig. 2, exposure of HK-2 cells to chemicals caused an approximately twofold decrease in intracellular ATP levels within 15 min of their addition compared with non-treated cells (filled squares vs. filled circles). This decrease in ATP levels was further induced in a time-dependent manner; the cellular ATP content decreased by four-, nine-, and sixfold following 30, 45, and 60 min of chemical treatment, respectively (p < 0.001, n = 3).
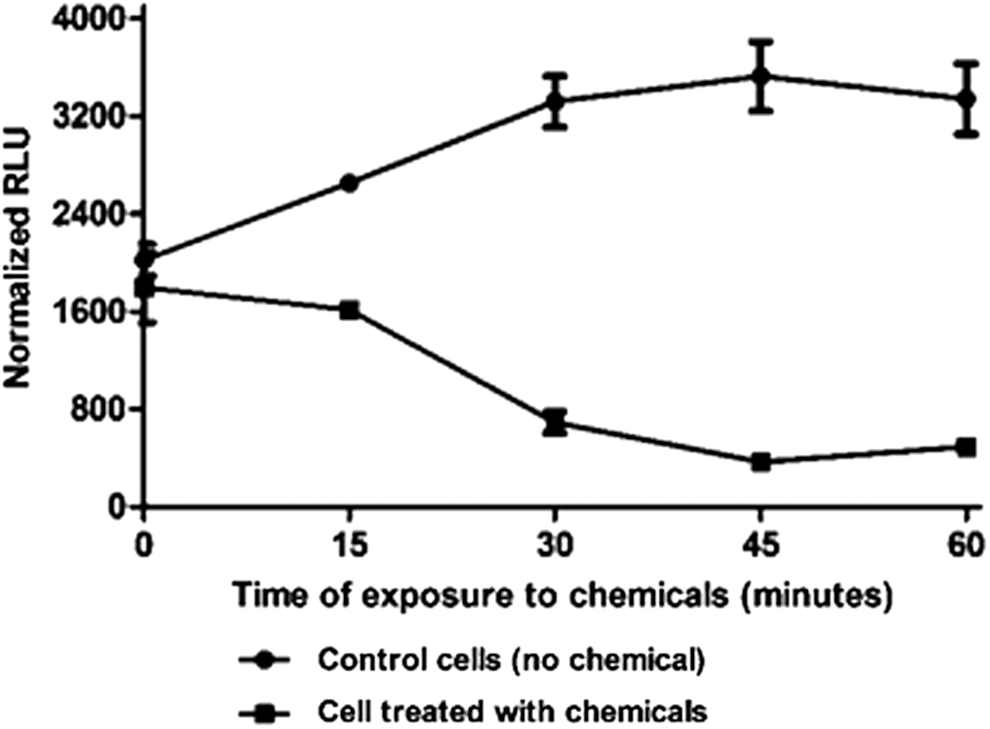
Time course of adenosine triphosphate (ATP) decay during simulated ischemia in HK-2 cells. Control cells were incubated in Keratinocyte serum-free medium. HK-2 cells were exposed to antimycin A (10 μM) and 2-deoxy-D-glucose (10 mM). ATP levels were determined every 15 min using a CellTitre-Glo® assay (Promega). Error bars represent the mean ± standard error of the mean (SEM; n = 3). A p-value of <0.001 indicated a significant difference between control and injured cells (two-way analysis of variance).
Protective effect of sod-2 following ATP depletion-recovery in HK-2 cells
To investigate whether BacMam-delivered sod-2 would protect against renal injury, HK-2 cells were transduced for 72 h with AcCMV_SOD-2 followed by exposure to antimycin A and 2-deoxyglucose. 41 As shown in Fig. 3, HK-2 cells transduced with AcCMV_SOD-2, 72 h prior to I/R injury, exhibited a time-dependent increase in their intracellular ATP content (filled triangles) compared with cells transduced with an empty BacMam containing no gene (AcCMV_NULL; filled square). BacMam-mediated sod-2 delivery showed an improved recovery of the cellular ATP content in the first 24 h of reperfusion (60% increases) compared with AcCMV_NULL transduced/injured cells (40%). ATP levels were significantly increased at 48 and 72 h after reperfusion in AcCMV_SOD-2 transduced/injured cells when compared with control cells and AcCMV_NULL transduced/injured cells (p < 0.001, n = 3).
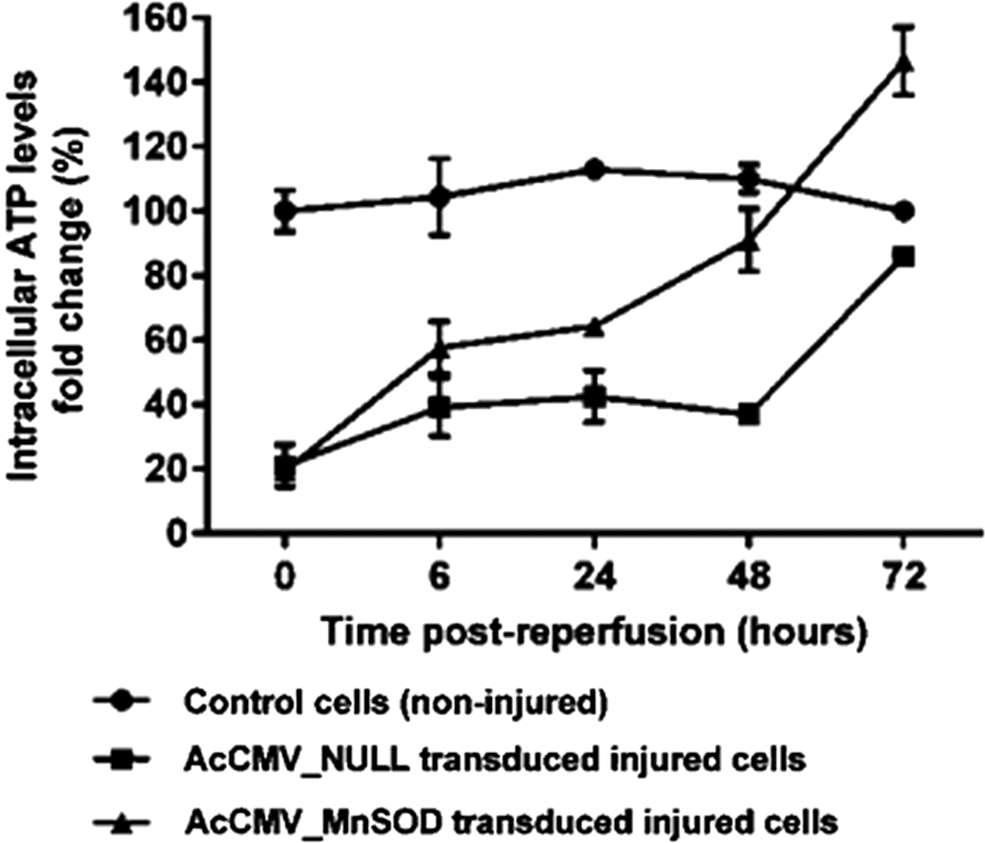
Intracellular ATP levels in ischemia–reperfusion (I/R) injured HK-2 cells. HK-2 cells were transduced with either AcCMV_NULL (filled squares) or AcCMV_MnSOD (filled triangles) at a MOI of 150. At 72 h post transduction, transduced HK-2 cells were subjected to 60 min of simulated ischemia. Intracellular ATP levels were quantified over time. Error bars represent the mean ± SEM (n = 3). A p-value of <0.001 indicated a significant difference between control cells (non-injured) and AcCMV_MnSOD transduced injured cells.
Cytotoxicity was measured by the amount of LDH released from the cells into the media following the ATP depletion-recovery period. As shown in Fig. 4, following 60 min of ischemia and 5 min of reperfusion, a significant increase in LDH release was detected (p < 0.05, n = 3) in AcCMV_NULL and AcCMV_SOD-2 transduced/injured cells (filled squares and triangles, respectively) compared with control cells (non-injured; filled circles). Prolonging the reperfusion phase, it was observed that SOD-2 overexpression in HK-2 cells resulted in 80%, 75%, and 70% reduction in LDH release following 24, 48, and 72 h of reperfusion, respectively, compared with AcCMV_NULL transduced/injured cells (Fig. 4). However, this time-dependent reduction in LDH release was only partial, since cytotoxicity in AcCMV_SOD-2 transduced injured cells, at 72 h after reperfusion, was still significantly higher compared with control cells (non-injured).
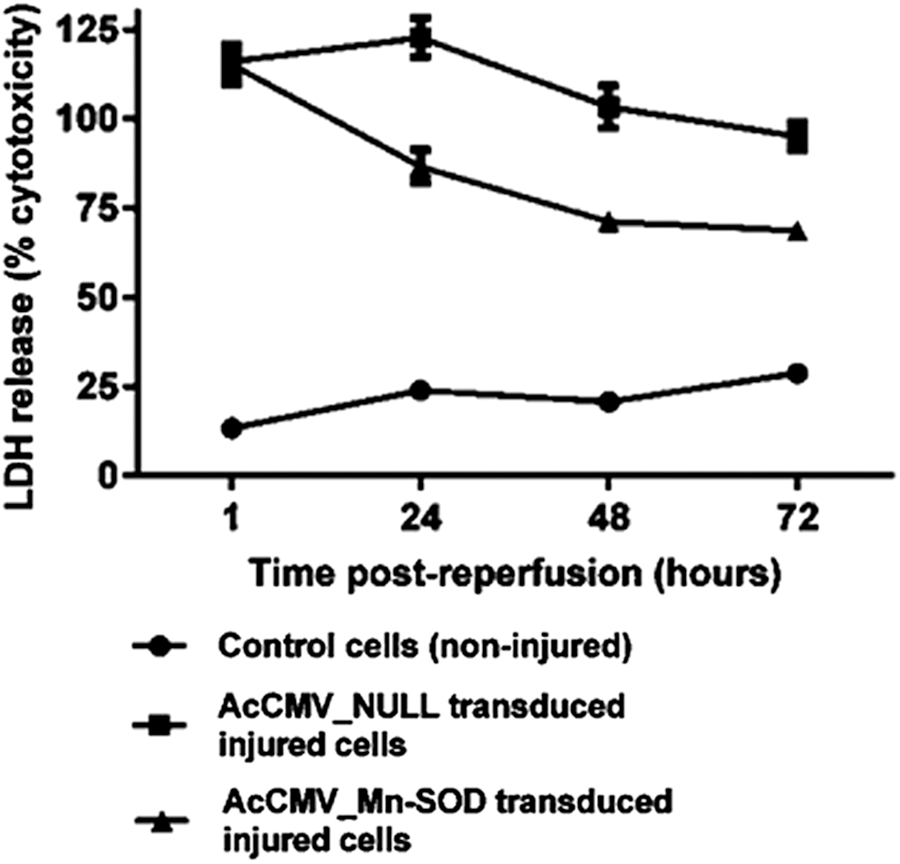
Effect of AcCMV_MnSOD on the release of lactate dehydrogenase (LDH) from injured HK-2 cells. Cells were transduced with either AcCMV_NULL (filled squares) or AcCMV_MnSOD (filled triangles) at a MOI of 150 for 72 h before being subjected to 60 min of simulated ischemia following reperfusion. AcCMV_MnSOD causes a time-dependent reduction of LDH release caused by antimycin A and 2-deoxyglucose. Data are expressed as the mean ± SEM (n = 3). A p-value of <0.001 indicated a significant difference between control cells (filled circles) and AcCMV_MnSOD transduced injured cells (filled triangles).
Effects of ex vivo BacMam-mediated sod-2 delivery against I/R-induced renal injury during cold preservation
Initially, the feasibility was demonstrated of ex vivo BacMam gene transfer into a whole porcine kidney during cold preservation using a fluorescent marker. Immuno-detection showed Enhanced Green Fluorescent Protein (EGFP) specific staining in AcCMV_EGFP transduced kidney tissues but not in control kidneys (Fig. 5).
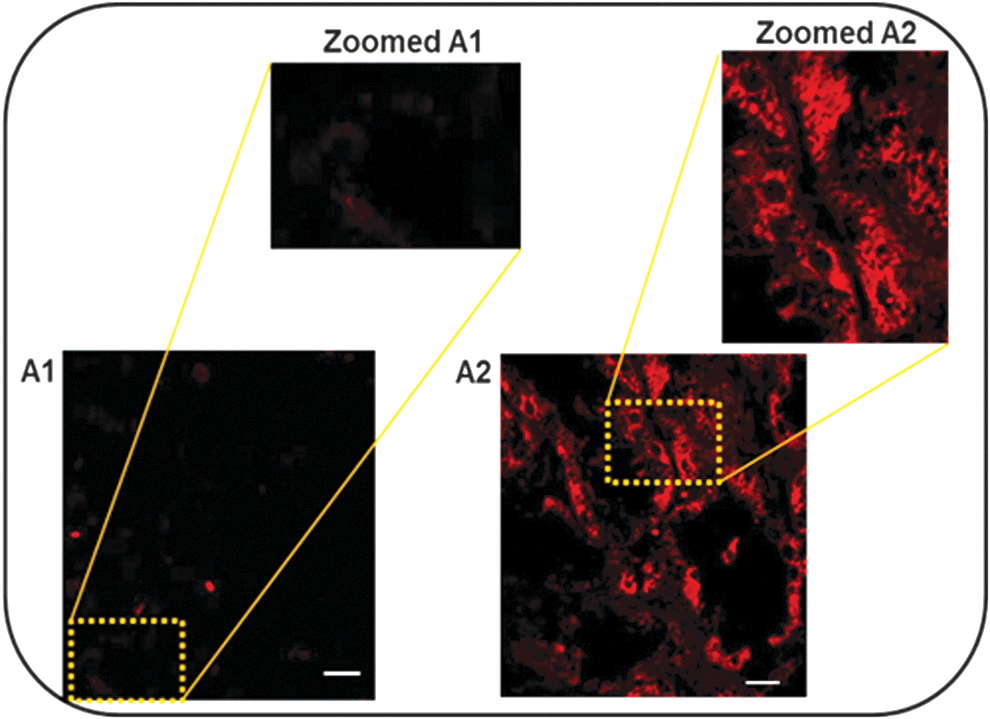
Egfp expression in sections of control and BacMam transduced kidneys. Kidneys were incubated in BELZER UW® solution supplemented with or without AcCMV_EGFP virus and incubated at 4°C for 24 h. (A1) Control and (A2) BacMam transduced kidneys. The kidney sections were fixed with paraformaldehyde, immunofluorescent stained for egfp primary antibody, and then labeled red with Alexa Fluor®-568 conjugated phalloidin secondary antibody. Tissues were analyzed by confocal microscopy using an oil immersion × 63 objective. Scale bar = 10 μm.
Six fresh porcine kidneys (three left and three right) were obtained from a local abattoir, flushed in situ, placed in a plastic bag surrounded by ice packs, and immediately transported to the laboratory for analysis. Kidneys were divided into three experimental groups: group I (perfused control kidneys, no virus), group II (AcCMV_EGFP transduced and perfused kidneys), and group III (AcCMV_SOD-2 transduced and perfused kidneys). Each group contained a paired kidney (one left and one right) excised from the same pig. During the 24 h period of perfusion/transduction, triplicate tissue samples were collected from the left kidneys of each experimental group, weighed, and enzymatically dissociated into single cells using collagenase for intracellular ATP analysis. As shown in Fig. 6, ATP levels detected 0 h post perfusion (hpp) in AcCMV_EGFP and AcCMV_SOD-2 transduced kidneys (0.4 and 0.3 nM, respectively) were not significantly different (p > 0.05, n = 3) compared to ATP concentrations detected in the non-transduced control group (0.3 nM). Significantly increased ATP levels were detected at 4 hpp in AcCMV_SOD-2 perfused/transduced kidney (0.7 nM) compared with those detected in non-treated transduced kidney and AcCMV_EGFP transduced/perfused kidney, 0.25 and 0.24 nM (p < 0.001, n = 3), respectively. ATP levels in AcCMV_EGFP transduced kidney were not significantly different (p > 0.05, n = 3) from the non-transduced control group (Fig. 6, light gray bars vs. black bars).
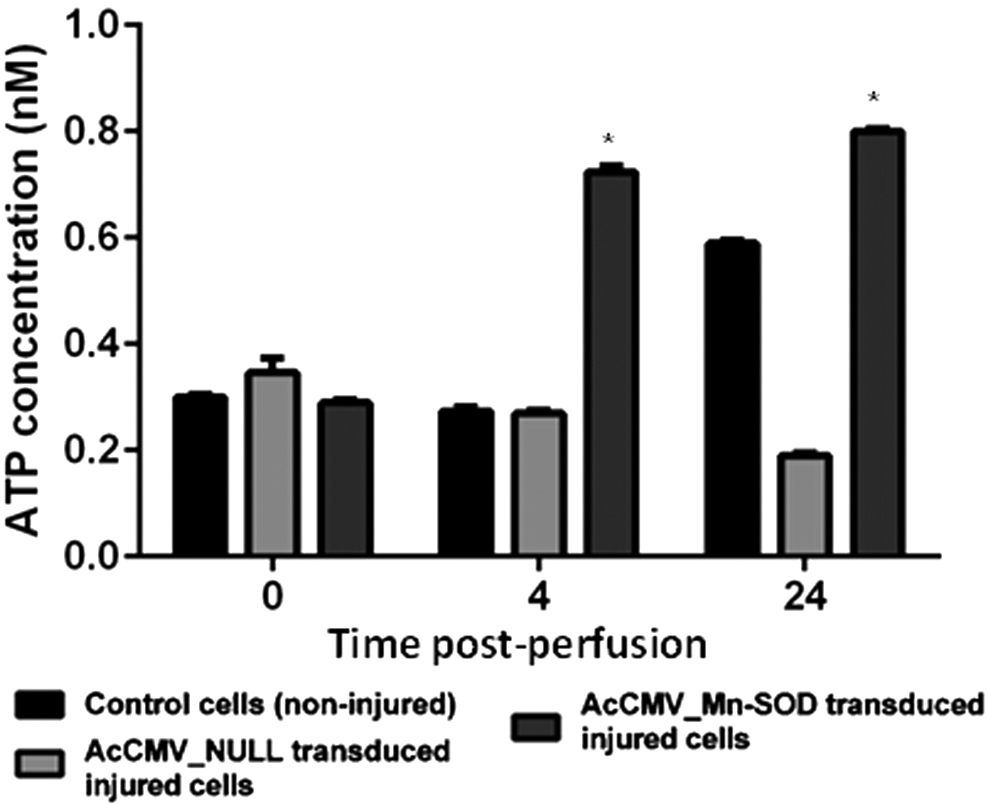
Intracellular ATP levels in non-transduced and BacMam perfused/transduced porcine kidneys. Tissue kidney samples were collected over time from control groups (no virus; black bars), AcCMV_EGFP (light gray bars) and AcCMV_MnSOD (dark gray bars) transduced kidney groups. Kidney samples were enzymatically dissociated in single cells, and the ATP level was measured over time. Error bars represent mean ± SEM (n = 3). A p-value of <0.001 indicated a significant difference between the ATP levels of perfused control experimental group and AcCMV_MnSOD perfused/transduced kidneys.
Furthermore, a marked and significant increase (p < 0.001, n = 3) in intracellular ATP concentration was detected at 24 hpp in AcCMV_SOD-2 transduced/perfused kidney tissue (0.8 nM) compared with ATP levels observed in the experimental control group 0.5 nM (Fig. 6, dark gray bars vs. black bars). In contrast, the intracellular ATP levels significantly decreased (p < 0.001, n = 3) in AcCMV_EGFP transduced/perfused kidney (0.2 nM) when compared with both AcCMV_SOD-2 and control experimental groups (Fig. 6, light gray bars vs. dark gray bars).
Discussion
In order to study renal I/R injury, initially in vitro experiments were performed using proximal tubular cells. Western blot analysis confirmed BacMam transduction of HK-2 cells and demonstrated increasing SOD-2 expression over time, as indicated by densitometry analysis (Fig. 1). Although a number of studies have described the delivery of sod-2 by viral vectors, mainly using adenovirus, 42,43 this is the first study demonstrating the feasibility of BacMam to deliver sod-2 to HK-2 cells.
The in vitro I/R injury model used in this study reproduced a number of key hallmark features of ischemia, such as intracellular ATP depletion as well as reduction in tissue oxygen and glucose contents. 44 Previous studies demonstrated that the combination of antimycin A and 2-deoxyglucose produced 90% ATP depletion in HK-2 cells. 41,44,45 This is consistent with the present findings where the combination of antimycin A and 2-deoxyglucose induced cellular injury. Indeed, the intracellular ATP levels were significantly reduced within 30 min of treatment (Fig. 2). Likewise, previous studies have demonstrated that the ATP/glucose depletion model of anoxia/ischemia Belzer UW dramatically depleted intracellular ATP levels to 1–4% of control cells, 41,46 as observed in the present experiments. Kevin et al. 47 demonstrated that treatment of Madin–Darby canine kidney cells with 10 μM of antimycin A dissolved in a Dulbecco's PBS (glucose-free) caused a rapid decrease in the intracellular level of ATP; after 2 h, levels were below the limit of detection and essentially zero after 5 h.
Having established and validated a cell culture model of renal injury, next the study determined whether I/R injury could be either reduced or prevented in HK-2 cells by the delivery of sod-2 via BacMam. SOD-2 transduced-injured cells showed a trend of increasing intracellular ATP levels over time (20–140%), with ATP recovery observed at 72 h, compared with AcCMV_NULL transduced/injured cells (19–86%; Fig. 3). Since the intracellular ATP content is essential for maintaining normal cellular functions as well as cellular integrity, it should be noted that any increase, even a small one, in ATP content during I/R injury might contribute to long-term cellular recovery. Although ATP recovery was observed at 72 h post reperfusion (hpr) in AcCMV_SOD-2 transduced/injured cells, the increase in ATP levels, observed between 6 and 48 hpr, also exerted a protective effect against I/R injury. These observations were in agreement with published models of renal I/R injury in cells overexpressing SOD-2. 35 Cruthirds et al. 48 demonstrated that after stable transfection, normal renal kidney cells producing recombinant SOD-2 were protected against I/R injury induced by ATP depletion. Koo et al. 43 generated a stable alveolar epithelial cell line overexpressing SOD-2. They demonstrated that the overexpression of SOD-2 significantly protected cells within the first 48 h of hyperoxia exposure and significantly increased the number of viable cells.
Exposure of transduced HK-2 cells to I/R injury (0 hpr) resulted in an increased release of LDH compared with control cells (Fig. 4). However, time-course analysis of I/R-induced LDH release in AcCMV_SOD-2 injured HK-2 cells showed a decrease in LDH levels at 24, 48, and 72 hpr compared with AcCMV_NULL transduced/injured cells, which remained higher during the 72 h of reperfusion (Fig. 4). These findings were consistent with the ATP data (Fig. 3).
A number of reports have demonstrated that the primary cause of ischemia in the kidneys is due to clamping of the renal arteries, which may occur during the surgical procedure of renal transplantation. 49 In particular, strategies for organ preservation are fundamentally based on the prevention or suppression of the metabolic changes that occur during I/R injury. 49
EGFP expression was observed following ex vivo transduction into porcine kidneys (Fig. 5, panel A2), 50 and to the authors' knowledge, this is the first evidence of ex vivo sod-2 delivery in kidneys using BacMam. The study also tested whether augmentation of sod-2 might improve I/R induced renal injury during cold preservation. A progressive and significant increase in ATP concentration was observed, between 4 and 24 hpp, in kidneys overexpressing sod-2 but not in control kidneys (Fig. 6, dark gray bars vs. black bars). Results also showed a significant negative effect in AcCMV_EGFP transduced kidney (Fig. 6, light gray bars), with markedly reduced levels of intracellular ATP at 24 hpp compared with untreated AcCMV_SOD-2 transduced kidneys (Fig. 6). These data were in line with results of the in vitro experiments (unpublished data).
These observations are consistent with those made by Land et al. 51 that sod exerts a beneficial effect against renal I/R injury through its antioxidant action. Furthermore, this study suggests a potential protective effect of ex vivo BacMam-mediated delivery of sod-2 against long cold kidney preservation injury. However, there are clearly limitations inherent to the ex vivo cold perfusion model used in this experiment that need to be addressed in future studies. The main limitation is represented by the short life-span of the organ perfused and knowing when valid data can be extracted during this time. 52 Ex vivo cold perfusion models are also restricted by the fact that hypothermic preservation can significantly decrease the organ's metabolic functions, which can preclude full recovery. 53
In conclusion, the present findings confirm the feasibility of using BacMam as a gene delivery vector to prevent renal deterioration after tissue I/R injury in vitro as well as in ex vivo isolated perfused porcine kidney. In particular, this study showed the beneficial effect of BacMam-delivered sod-2, which protected kidney from further renal injury after ischemia.
Footnotes
Acknowledgments
The authors are grateful to Dr. Susan Brooks for helpful discussions on histopathology. We also thank Drs. John Runions and Sarah Irons for their assistance with confocal microscopy. This study was funded by Oxford Brookes University studentship and partially supported by Oxford Expression Technologies Ltd., Oxford, United Kingdom.
Author Disclosure
The authors declare no conflicting interests.