
Abstract
Mucopolysaccharidosis type II (MPS II) is a rare X-linked genetic disorder caused by deficiency of the lysosomal enzyme iduronate-2-sulfatase (IDS), leading to impaired catabolism of ubiquitous polysaccharides and abnormal accumulation of these undegraded substrates in the lysosome. Like many lysosomal storage diseases, MPS II is characterized by both somatic and central nervous system (CNS) involvement. Intravenous enzyme replacement therapy can improve somatic manifestations of MPS II, but systemic IDS does not cross the blood–brain barrier and therefore cannot address CNS disease. In this study, an adeno-associated virus serotype 9 vector carrying the IDS gene was injected into the cerebrospinal fluid (CSF) of IDS deficient mice, a model of MPS II. Treated mice exhibited dose-dependent IDS expression and resolution of brain storage lesions, as well as improvement in long-term memory in a novel object recognition test. These findings suggest that delivery of adeno-associated virus vectors into CSF could serve as a platform for efficient, long-term enzyme delivery to the CNS, potentially addressing this critical unmet need for patients with MPS II and many related lysosomal enzyme deficiencies.
Introduction
M
MPS II encompasses a broad spectrum of clinical presentations; in the most severe cases, patients present in early childhood with marked developmental delay and somatic disease and typically do not survive past the second decade. In the attenuated form of the disease, patients may not present until late in childhood, have little or no cognitive dysfunction, and have a normal life expectancy. Definitive correlations between genotype and phenotype have not been possible due to the high frequency of private mutations 2 ; however, among characterized mutations there appears to be a clear relationship between residual enzyme activity and clinical course. 3 These studies suggest that even 2% of normal IDS activity can substantially improve patient outcomes, a finding which is encouraging for therapeutic strategies aimed at augmenting IDS activity.
A recombinant form of IDS (Elaprase, Shire) has been approved in the United States and European Union for the treatment of MPS II. As with many lysosomal enzymes, the presence of terminal mannose-6-phosphate residues on glycosylated IDS allows for internalization of the enzyme by cells expressing the mannose-6-phosphate receptor. 4,5 Given as a weekly infusion, recombinant IDS improves organomegaly, mobility, and respiratory function. The primary limitation of enzyme replacement therapy is that the enzyme does not cross the blood–brain barrier, and therefore does not address the central nervous system (CNS) features of the disease. Clinical trials are ongoing to evaluate delivery of the enzyme into the cerebrospinal fluid (CSF) as a means of bypassing the blood–brain barrier. 6,7 This approach has shown promise in preclinical studies, although chronic direct CNS administration may not be feasible in clinical practice. 6
The small amount of enzyme required to attenuate the disease phenotype, as well as the ability of cells to internalize secreted IDS, suggests that correcting the genetic defect in even a small subset of cells could lead to widespread improvement in CNS disease. Gene transfer to small regions of the brain using adeno-associated virus (AAV) vectors has previously proven safe and feasible in humans. 8 –10 Recent studies using intrathecal (IT) delivery of more efficient, second-generation AAV vectors have demonstrated widespread gene transfer throughout the CNS of large animals after a single, minimally invasive injection, 11 –14 making this approach particularly suitable for clinical applications. In large animal models of lysosomal storage diseases, such as MPS I and MPS VII, IT AAV delivery has resulted in global correction of brain storage lesions. 15 –17
In this study we applied this approach to a murine model of MPS II. Delivery of an AAV serotype 9 (AAV9) vector carrying the human IDS gene into the CSF via intracerebroventricular (ICV) injection resulted in dose-dependent resolution of brain storage lesions, as well as evidence of functional improvements. AAV9 delivery into the CSF appears to be a promising approach for the treatment of CNS disease in MPS II.
Materials and Methods
Vectors
The human IDUA and IDS cDNAs were cloned into an expression construct containing a chicken beta actin promoter, cytomegalovirus enhancer, intron, and rabbit beta globin polyadenylation sequence. The expression constructs were flanked by AAV2 inverted terminal repeats. AAV9 vectors were generated from these constructs by triple transfection of HEK 293 cells and iodixanol purification as previously described. 18
Animal procedures
All animal protocols were approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania. IDS knockout mice were obtained from Jackson Laboratory (Stock no: 024744) and bred in house. Wild type C57BL/6 males from the colony served as controls. At 2–3 months of age, animals were anesthetized with isoflurane and injected ICV with 5 μL vector diluted in sterile phosphate buffered saline. CSF was collected at the time of necropsy by suboccipital puncture with a 32-gauge needle connected to polyethylene tubing. Terminal serum samples were collected by cardiac puncture. Animals were euthanized by exsanguination under ketamine/xylazine anesthesia. Death was confirmed by cervical dislocation. The brain, heart, lungs, liver, and spleen were collected on dry ice. For histology experiments, the brain was divided into an anterior half which was fixed for LIMP2 immunohistochemistry, and a posterior half fixed for GM3 immunohistochemistry. Two cohorts of mice were used for behavior experiments. An initial cohort of wild type and IDS knockout mice was tested in a battery of procedures to determine whether gene deletion affected learning and memory. A second cohort of mice treated with low, medium, or high vector doses of AAV9.CB.hIDS (3 × 108, 3 × 109, or 3 × 1010 GC, respectively) was used to investigate the feasibility of ICV delivery as a strategy to rescue behavior deficits.
Behavior procedures: All behavior procedures were performed by operators blinded to genotype and treatment group.
Open field activity
Spontaneous activity in an open field was measured with a Photobeam Activity System—Open Field (San Diego Instruments, San Diego, CA). Mice were individually placed in the arena for a single 10 min trial. Horizontal and vertical beam breaks were collected to assess general locomotion and rearing activity.
Y maze
Short term memory was assessed with a standard Y-shaped maze (San Diego Instruments). The sequence and number of arm entries was recorded during an 8 min trial. A spontaneous alternation (SA) was defined as sequential entry into all three arms of the maze without immediately returning to a previously entered arm. Total arm entries (AE) were collected as a measure of motor activity. The percent spontaneous alternation was calculated as %SA = [SA/(AE−2) × 100].
Contextual fear conditioning
Conditioning experiments were performed as described by Abel et al. 18 On the training day, mice were allowed to explore the unique conditioning chamber (Med Associates) for 300 s. A nonsignaled, 1.5 mA continuous footshock was delivered between 248 and 250 s. After an additional 30 s in the chamber, the mice were returned to their home cage. Twenty-four hours later, recall of spatial context was assessed for 5 consecutive minutes in the same chamber where training occurred. Memory was assessed with software used to score freezing behavior (Freezescan; CleverSystems). The percent freezing in the 2.5 min prestimulus epoch of the training session is compared to the percent freezing upon reexposure to the chamber. An increase in freezing indicates that learning has occurred.
Novel object recognition
The experimental apparatus consisted of a grey rectangular arena (60 × 50 × 26 cm) on a white floor and two unique objects: 3.8 × 3.8 × 15 cm metal bars and 3.2 cm dia. × 15 cm polyvinyl chloride pipes. Prior to exposure to the apparatus, mice were handled 1–2 min/day for 5 days. During a 5-day habituation phase, mice were allowed to explore an empty arena for 5 min/day. During the training phase, mice explored two of the same objects for 15 min to establish familiarization. In the recall phase 24 h later, mice were returned to the arena with one now-familiar object and a novel object. Mice will preferentially explore the novel object. A reduced preference for novelty suggests a failure to recall the familiar object and thus a learning deficit. All sessions were recorded, and time spent exploring objects was scored with an open source image analysis program. 19
Enzyme and GAG assays
GAG, Hex, and IDUA assays were performed as previously described.
16
IDS activity was measured by incubating 10 μL sample with 20 μL of 1.25 mM 4-methylumbelliferyl (MU) α-
Histology
The brain was divided into an anterior half, which was fixed for LIMP2 immunohistochemistry, and a posterior half fixed for GM3 immunohistochemistry. LIMP2 and GM3 immunohistochemistry were performed as previously described. 16 The number of cells staining positive for LIMP2 and GM3 was quantified in four brain sections from each animal by a blinded reviewer.
Vector biodistribution
Tissues for vector biodistribution analysis were quickly dissected and frozen on dry ice. Samples were stored at −80°C until the time of analysis. DNA was isolated from tissues using the QIAmp DNA Mini Kit and vector genomes quantified by TaqMan PCR as previously described. 20
ELISA for anti-hIDS antibodies
Polystyrene ELISA plates were coated overnight with recombinant human IDS (R&D Systems) 5 μg/mL in PBS titrated to pH 5.8. Plates were washed and blocked 1 h in 2% bovine serum albumin in neutral PBS. Plates were then incubated with serum samples diluted 1:1000 in PBS. Bound antibody was detected with HRP conjugated goat anti-mouse antibody (Abcam) diluted 1:10,000 in PBS with 2% BSA. The assay was developed using tetramethylbenzidine substrate and stopped with 2 N sulfuric acid before measuring absorbance at 450 nm. Titers were determined from a standard curve generated by serial dilution of a positive serum sample arbitrarily assigned a titer of 1:10,000.
Statistics
Tissue GAG content, Hex activity, and brain storage lesions in treated and untreated mice were compared using a one-way ANOVA followed by Dunnett's multiple comparisons test. Open field and Y maze date were analyzed with Students t-test. A two-way ANOVA and Dunnett's post-hoc analysis was applied to the fear conditioning data to assess trial and genotype effects. For the novel object recognition test, time exploring the novel object versus familiar object was compared using a t-test for each group, followed by a Bonferroni correction for multiple comparisons.
Results
The MPS II mouse model was generated by homologous recombination–mediated disruption of exons 4 and 5 of the IDS gene. The model exhibits no detectable enzyme activity and develops histological storage lesions similar to those found in MPS II patients. IDSy/− mice also exhibit some clinical features of the disease, including skeletal deformities. The neurobehavioral phenotype of the mice has not been extensively evaluated, although some studies have indicated potential abnormalities. 21
Male IDSy/− mice between 2 and 3 months of age were treated with an ICV injection of an AAV9 vector expressing human IDS from a chicken beta actin promoter with a cytomegalovirus enhancer (AAV9.CB.hIDS). While a variety of potential approaches could be used clinically to deliver vector into the CSF (e.g., lumbar puncture, C1–C2 puncture, or suboccipital puncture), the ICV approach was used for the present study due to the greater feasibility and reproducibility of this method in mice. In an initial cohort, mice were treated with one of three vector doses (3 × 108, 3 × 109, or 3 × 1010 genome copies [GC], n = 8 per group) and sacrificed 3 weeks after vector administration for assessment of vector biodistribution and IDS expression. Untreated IDSy/− mice and wild type male littermates served as controls. Analysis of vector biodistribution in the high-dose group demonstrated efficient brain targeting, with an average of one vector genome per host diploid genome (Fig. 1). Consistent with previous studies of AAV delivery into CSF, there was also vector escape to the periphery and efficient hepatic targeting, with more than one vector genome per host diploid genome (Fig. 1).
12,15,22
Brain tissue, CSF, and serum all exhibited dose-dependent increases in IDS activity, which approached or exceeded wild type levels in the high-dose group (Fig. 2). Antibodies to human IDS were detected in serum of several animals in the mid- and high-dose cohorts, although this did not appear to significantly affect circulating enzyme activity (Supplementary Fig. S1; Supplementary Data are available online at

Vector biodistribution following adeno-associated virus serotype 9 (AAV9) delivery into cerebrospinal fluid. Mucopolysaccharidosis type II (MPS II) mice were treated with an intracerebroventricular (ICV) injection of 3 × 1010 genome copies (GC) of an AAV9 vector. Three weeks after injection, the animals were sacrificed and vector genomes were quantified in tissue samples by Taqman PCR.
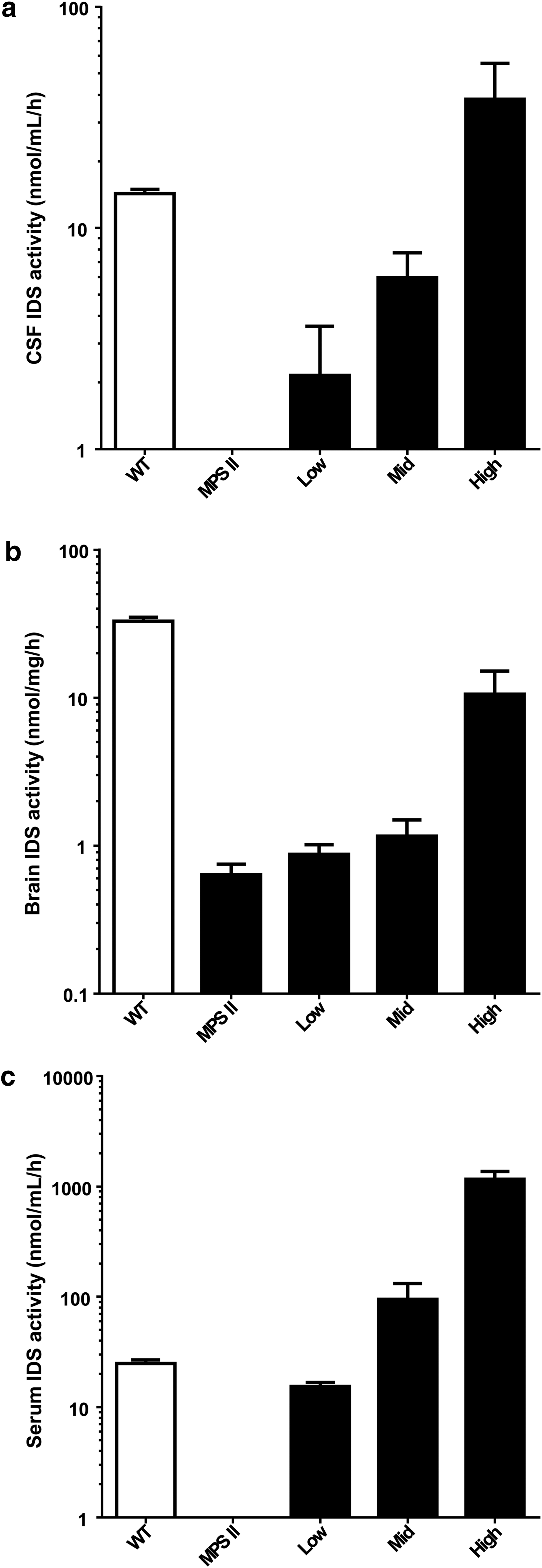
Iduronate-2-sulfatase (IDS) expression in central nervous system and blood of MPS II mice after ICV vector treatment. MPS II mice were treated at 2–3 months of age with an ICV injection of AAV9 vector expressing human IDS from a chicken beta actin promoter with a cytomegalovirus enhancer (AAV9.CB.hIDS) at one of three doses: 3 × 108 GC (low), 3 × 109 GC (mid), or 3 × 1010 GC (high). Animals were sacrificed 3 weeks after injection. IDS activity was measured in cerebrospinal fluid
In order to evaluate the therapeutic potential of ICV AAV9–mediated delivery of the IDS gene, an additional cohort of IDSy/− mice was treated with equivalent vector doses, then evaluated at later time points to assess the impact of gene transfer on disease progression. Two months after vector administration the mice were subjected to behavioral and neurocognitive tests. Three months after treatment the animals were sacrificed and tissues harvested for histological and biochemical assessment of disease activity.
Consistent with high levels of serum enzyme activity, GAG storage was reduced in the liver, and there was a nonsignificant trend toward normalization of GAG storage in the heart (Fig. 3A, B). In addition, activity of the lysosomal enzyme hexosaminidase (Hex), which is upregulated in the setting of GAG storage, was normalized in both tissues at all doses (Fig. 3C, D).
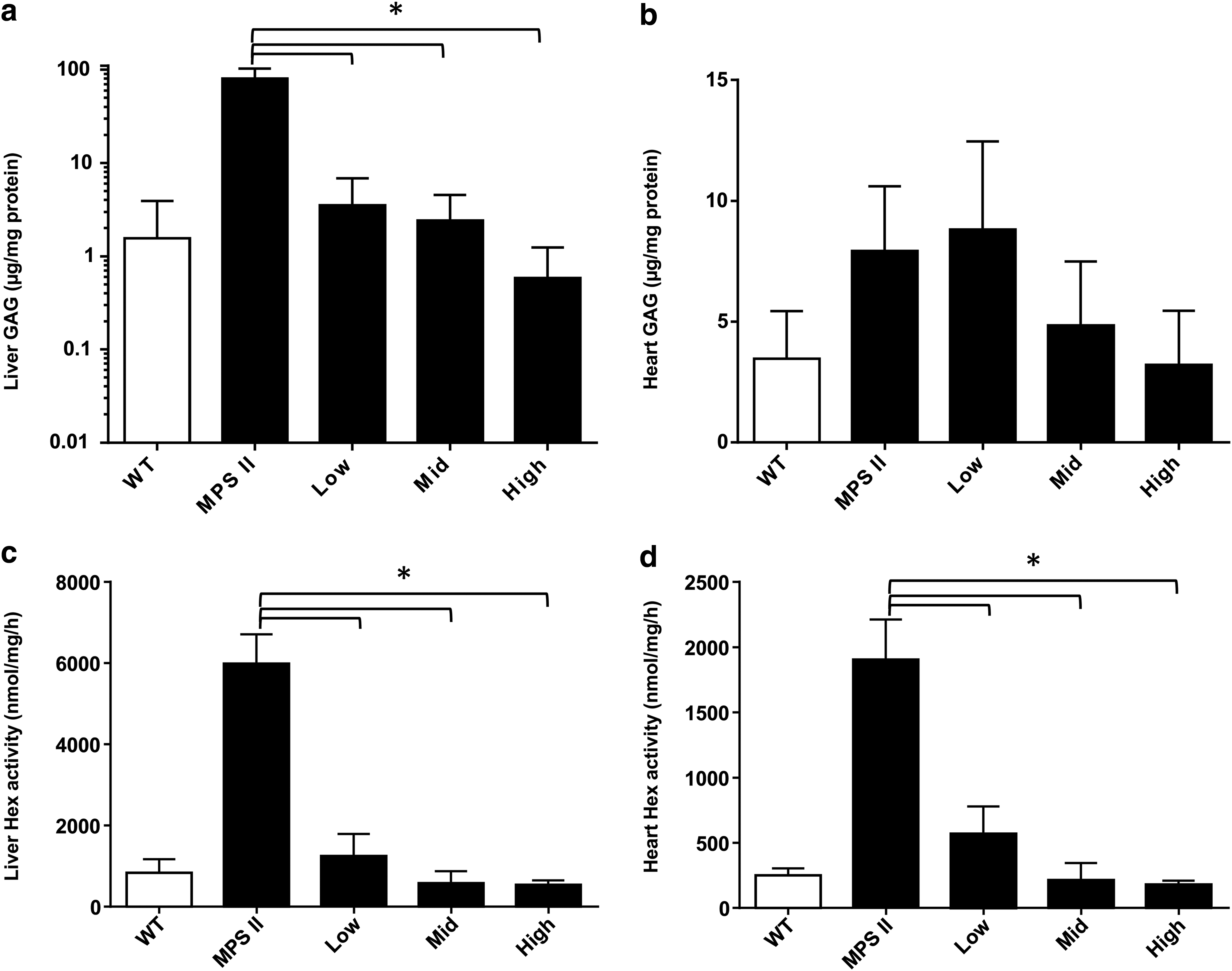
Correction of peripheral organ glycosaminoglycan storage after ICV vector delivery in MPS II mice. MPS II mice were treated at 2–3 months of age with an ICV injection of AAV9.CB.hIDS at one of three doses: 3 × 108 GC (low), 3 × 109 GC (mid), or 3 × 1010 GC (high). Animals were sacrificed 3 months after injection. Glycosaminoglycan content was measured in liver
The brains of untreated IDSy/− mice showed clear histological evidence of lysosomal storage in neurons, including accumulation of the lysosomal membrane protein LIMP2, as well as secondary storage of gangliosides including GM3 (Fig. 4A). Treated mice demonstrated dose-dependent decreases in neuronal storage lesions evident by both LIMP2 and GM3 staining (Fig. 4B, C).

Dose-dependent resolution of brain storage lesions in MPS II mice. MPS II mice were treated at 2–3 months of age with an ICV injection of AAV9.CB.hIDS at one of three doses: 3 × 108 GC (low), 3 × 109 GC (mid), or 3 × 1010 GC (high). Animals were sacrificed 3 months after injection, and brains were stained for the lysosomal membrane protein LIMP2 and the ganglioside GM3. Representative cortical brain sections are shown
Behavioral testing was performed when the mice reached 4–5 months of age, 2 months after vector administration. A comprehensive battery of tests was performed to evaluate general behavior as well as short and long-term memory. IDSy/− mice showed normal exploratory activity in an open field arena (Supplementary Fig. S2 A–C). Spontaneous alternations in a Y-maze were used to assess short term working memory. IDSy/− mice had similar numbers of arm entries (Supplementary Fig. S2 D) and equivalent spontaneous alternations to wild type littermates, demonstrating intact short term memory (Fig. 5A). Long-term memory was assessed using classic contextual fear conditioning and novel object recognition (NOR). In fear conditioning, the association of an aversive stimulus with a specific context evokes freezing behavior upon reexposure to that context. All mice showed an increase in the percent time freezing during the recall phase of the test demonstrating that learning occurred, however IDSy/− mice showed reduced freezing relative to wild type littermates (Fig. 5B). In treated animals, there was no clear improvement in contextual fear conditioning, although treatment effects were difficult to evaluate due to the small difference between normal and untreated IDSy/− mice (Fig. 5B). In NOR, mice are allowed to explore a pair of similar objects. Twenty-four hours after training, one now-familiar object is replaced with a novel object. Mice have an innate propensity to explore the novel object; failure to do so demonstrates a lack of recognition of the familiar object and reveals a memory deficit. Wild type mice demonstrated a preference for a novel object, but IDSy/− mice did not show a preference. Remarkably, intrathecal AAV9 gene therapy rescued the long term NOR deficits observed in the IDSy/− mice (Fig. 5C). Object discrimination appeared to be improved in all treated IDSy/− mice, with all groups exhibiting a trend toward greater percentage of time exploring a novel object compared with a familiar one. The preference for the novel object was statistically significant only in the mid-dose cohort, although the study was not sufficiently powered to compare the relative degree of rescue of behavioral deficits among dosing groups (Fig. 5).

Improved object discrimination in vector treated MPS II mice. Untreated MPS II mice and WT male littermates underwent behavioral testing at 4–5 months of age. MPS II mice were treated at 2–3 months of age with an ICV injection of AAV9.CB.hIDS at one of three doses: 3 × 108 GC (low), 3 × 109 GC (mid) or 3 × 1010 GC (high). Y maze behavior
One drawback of evaluating AAV delivery into CSF in a murine disease model is that the extremely small mouse CNS—with a brain mass of just 0.4 g and CSF volume of 40 μL—may not accurately model the diffusion of vector and secreted enzyme in the human brain, which is 3,500-fold larger. In previous studies of the related lysosomal storage disease mucopolysaccharidosis type I (MPS I), we utilized naturally occurring large animal disease models to address this issue. 15,16 With average brain masses of 30 g and 72 g, respectively, the MPS I cat and dog provided much more realistic models to address the challenge of achieving widespread vector and enzyme delivery in the human brain. Studies carried out in these models provided critical evidence of the efficacy of AAV9 delivery into CSF for MPS I. 15,16 In order to determine whether similar efficacy would be possible using this same approach in MPS II, we performed a parallel dose-ranging study in MPS I mice to examine the relative efficiency of enzyme expression and cross-correction in MPS II. MPS I mice were treated with a vector identical to that used for IDSy/− mice, with the exception of the α-L-iduronidase (IDUA) transgene in place of IDS. As with the MPS II study, MPS I mice were treated with doses of 3 × 108, 3 × 109, or 3 × 1010 GC (n = 8 per group) at 2–3 months of age and sacrificed at day 21 posttreatment for measurement of brain enzyme activity or at 3 months posttreatment for histological analysis. There was a similar dose response in brain enzyme expression to that observed in IDSy/− mice, although expression of IDUA appeared somewhat more efficient compared with IDS, both in absolute enzyme activity and relative to wild type expression levels (Fig. 6). Correction of brain storage lesions was similar between the two disease models, although treatment appeared modestly more effective in MPS I mice at the lowest vector dose. Together these results suggest that AAV9-mediated gene transfer for MPS II yields correction of brain storage lesions nearly as efficient as that observed in MPS I, and that this approach will remain effective when scaled up from mice. However, slightly higher vector doses compared to MPS I may be necessary in MPS II in order to overcome less efficient enzyme expression in the brain.

Comparison of enzyme expression and correction of brain storage lesions in MPS I mice treated with ICV AAV9. MPS I mice were treated at 2–3 months of age with an ICV injection of AAV9.CB.hIDUA at one of three doses: 3 × 108 GC (low), 3 × 109 GC (mid), or 3 × 1010 GC (high). One cohort of animals was sacrificed at 3 weeks post vector injection, and brains were harvested for measurement of IDUA activity
Discussion
There is a critical unmet need in MPS II and many related lysosomal storage diseases for a safe and effective method to deliver the deficient enzymes to the CNS. Viral vector-mediated delivery of the gene encoding the enzyme to even a small subset of cells could establish a depot of secreted enzyme in the brain, allowing for cross-correction of cells throughout the CNS. CNS-directed gene transfer has previously been limited by the inflammatory response and restricted distribution achieved with direct brain injections of AAV vectors, and the low efficiency of CNS targeting with intravascular vector delivery. 23 –25 The development of CSF delivery approaches and more efficient neurotropic AAV vectors has now made it possible to achieve gene transfer to cells throughout the brain of large animals using a minimally invasive injection approach, potentially allowing for sustained and effective levels of transgene expression in diseases like MPS II. AAV delivery into the CSF has the additional advantage of being insensitive to preexisting antibodies against the AAV capsid, making this injection method applicable to larger patient populations than are intravascular approaches. 22,26
In the present study, ICV delivery of an AAV9 vector carrying the human IDS gene in MPS II mice resulted in therapeutic levels of expression in the CNS and resolution of brain storage lesions. Functional improvement was indicated by greater novel object discrimination in treated mice compared with untreated controls. Neurocognitive deficits have not been previously well characterized in IDSy/− mice. Our findings demonstrate that IDSy/− mice have normal exploratory activity in the open field as well as similar arm entries in the Y-maze relative to wild type littermates. These locomotor assessments are critical in considering other types of behavior that require normal exploration. We found that IDSy/− mice have intact short term working memory demonstrated by similar spontaneous alternations in the Y-maze compared with controls. In contrast, Higuchi et al. 21 reported reduced spontaneous alternations and increased arm entries in 32 week old IDSy/− mice. The differing Y-maze results may be due to the different age at testing and underscore the progressive nature of MPS II. We assessed the impact of IDS deficiency on two forms of long term memory. We identified a mild deficit in contextual fear conditioning in the IDSy/− mouse. Although IDSy/− mice were able to recall the context in which they received an aversive stimulus, they showed a significantly reduced freezing response compared to wild type littermates. IDSy/− mice that received IT delivery of AAV9 showed freezing responses similar to uninjected mice. Therefore, no recovery was detected. A long term memory deficit was also found in novel object recognition. In NOR, IDSy/− mice showed no preference for a novel object relative to a familiar object. Vector treated mice demonstrated recovery of the NOR deficit seen in untreated IDSy/− mice. Object discrimination was statistically significant only in the mid-dose cohort, although with 8 animals per group, the study was not well powered to evaluate the dose response of behavioral endpoints. The ability of IT delivery of AAV9 to differentially affect recovery of two types of long term memory may be due to different neural substrates required for each task. Contextual fear conditioning is a hippocampal-dependent task as opposed to NOR which requires the perientorhinal cortex. 27,28 Thus, future studies are warranted to discern the neuroanatomical loci of inclusions and how the extent of biochemical rescue at each locus impacts recovery of particular cognitive functions.
Comparison of the efficiency of ICV AAV9 delivery in mouse models of MPS II and MPS I indicated that enzyme expression and tissue correction were similar between the two diseases, providing encouraging evidence that AAV delivery into CSF for MPS II will be efficient enough to yield widespread disease correction in the context of much larger brain sizes and CSF volumes, as has previously been shown in large animal models of MPS I. 15,16 Interestingly, expression of the deficient enzyme was somewhat less efficient in MPS II compared with MPS I. This could simply be a product of the expression efficiency of these particular vectors, although the control elements in the expression constructs were identical. Some studies have suggested that expression of sulfatases such as IDS can be limited by the availability of the sulfatase modifying factor, SUMF1, which is required for posttranslational modification of IDS. 29 However, our pilot experiments evaluating co-expression of IDS and SUMF1 have not demonstrated more active IDS expression (unpublished results). Regardless, correction of storage lesions was nearly as efficient in MPS II mice as in MPS I mice, indicating that gene transfer should be similarly effective for both MPS I and MPS II patients, though in the latter case modestly higher vector doses may be necessary for optimal outcomes.
In addition to CNS gene transfer, there was significant liver transduction and peripheral enzyme expression in treated MPS II mice, indicating possible systemic benefits of AAV9 delivery into CSF. This is consistent with studies of CSF AAV delivery in a variety of other species. 11,12,15 –17,22,26 Notably, liver transduction after AAV injection into CSF varies substantially among species, so it is not yet clear whether humans would exhibit significant peripheral expression.
This study demonstrates that AAV9 delivery into the CSF can achieve effective IDS gene transfer to the brain and resolve CNS manifestation of MPS II, supporting the advancement of this approach into the clinic.
Footnotes
Acknowledgments
The authors would like to acknowledge the support of the University of Pennsylvania Vector Core (Philadelphia, PA) and the University of Pennsylvania Neurobehavior Testing Core (Philadelphia, PA). The study was funded by a grant from REGENXBIO (J.M.W.).
Author Disclosure
J.M. Wilson is an advisor to REGENXBIO, Dimension Therapeutics, and Solid Gene Therapy and is a founder of, holds equity in, and has a sponsored research agreement with REGENXBIO and Dimension Therapeutics; in addition, he is a consultant to several biopharmaceutical companies and is an inventor on patents licensed to various biopharmaceutical companies. The remaining authors have nothing to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.