
Abstract
The bone marrow (BM) microenvironment, heavily composed of osteoblasts, plays a key role during the normal development of hematopoiesis. Endogenous miR-22 has an important function in the hematopoietic development and osteoblastic differentiation. It is unclear whether miR-22 in osteoblasts from the BM microenvironment also has an important function in the development of hematopoiesis. This study found that the capacity of hTERT-transduced fetal bone marrow osteoblasts (FBMOB-hTERT) cells to expand human cord blood (CB) CD34+ cells and maintain the multipotency of CB CD34+ cells is decreased upon ectopic expression of miR-22. Further experiments revealed that with the existence of CB CD34+ cells, the expression of β-catenin in FBMOB-hTERT cells is decreased upon ectopic expression of miR-22. The reduced ability of FBMOB-hTERT cells to expand human CB CD34+ cells and maintain the multipotency of CB CD34+ cells upon ectopic miR-22 was partly rescued by overexpression of β-catenin. The study indicated that the ability of osteoblasts to expand human CB CD34+ cells and maintain the multipotency of CB CD34+ cells is decreased upon ectopic expression of miR-22. The decreased expression of β-catenin is, at least partly, responsible for the reduced ability of osteoblasts for expanding and supporting CB CD34+ cells upon ectopic expression of miR-22.
Introduction
H
MicroRNAs (miRNAs) are a class of short, non-coding RNAs, each about 21–23 nucleotides in length. They can post-transcriptionally regulate the expression of mRNA. 11 Some miRNA genes are located near translocation breakpoints and are present in loci targeted for deletion in human leukemia, which suggests that miRNAs may have important functions in hematopoietic malignancies. 12 Furthermore, several groups presented the evidence suggesting that miRNAs play a key function during the normal hematopoietic development, including expansion, cell cycle, and self-renewal. 13,14 The relationship between the expression of miRNAs and the hematopoietic multipotent capacity has been studied by many research groups. 15 –18 MiR-22 is expressed and plays a key role in hematopoiesis. 19 –21 MiRNA expression profiling from cord blood (CB)-derived CD34 cells revealed that miR-22 has a strong positive correlation with the appearance of erythroid surface antigens and hemoglobin synthesis. 19 Expression signatures of intragenic miRNAs and their corresponding host genes in myeloid leukemia cells demonstrated that the expression signature of miR-22 is associated with the developmental lineages and specific leukemia subtypes. 20 Song et al. showed that, as a potent proto-oncogene, the aberrations in the miR-22-TET2 regulatory network are common in hematopoietic malignancies. 21 In contrast to the oncogenic role of miR-22 reported by Song et al., Jiang et al. showed that miR-22 is an essential antitumor gatekeeper in de novo acute myeloid leukemia (AML) where it is significantly downregulated. 22 Although different opinions exist about the function of miR-22, all studies indicate that miR-22 plays a key role in the development of hematopoiesis. The expression of miR-22 is also involved in the process during which the primary bone marrow stromal cells (BMSCs) differentiate into osteoblasts, 23,24 the key component of the hematopoietic microenvironment. The altered condition of the hematopoietic niche may correlate with the changed production of the hematopoietic supporting molecules and, as a result, changed support of hematopoiesis. Collectively, endogenous miR-22 has an important function in hematopoietic development and osteoblastic differentiation. However, it is not clear whether miR-22 from the BM microenvironment, especially in osteoblasts, is a key molecule for normal hematopoiesis.
One immortalized clone with the characteristic of osteoblasts, designated as hTERT-transduced fetal bone marrow osteoblasts (FBMOB-hTERT), has been reported previously. FBMOB-hTERT cells were derived from human fetal BMSCs with retroviral vectors containing the human telomerase catalytic subunit (hTERT) gene. 25 It was found that FBMOB-hTERT cells can support the expansion of CB CD34+ cells and maintain their multipotency. 25 In this study, the expression of miR-22 was found to be significantly decreased in FBMOB-hTERT cells and BMSCs that were subjected to osteoblastic differentiation (BMSCs-OD). In vitro and in vivo results indicate that the ability of FBMOB-hTERT cells to expand human CB CD34+ cells and maintain their multipotency was inhibited upon ectopic expression of miR-22. After interaction with CB CD34+ cells, the expression of β-catenin was further found to be significantly decreased. The reduced ability of FBMOB-hTERT cells to expand human CB CD34+ cells and maintain their multipotency due to the ectopic expression of miR-22 can be, at least partly, rescued by overexpression of β-catenin. These results suggest that the decreased expression of β-catenin is partly responsible for the reduced ability of FBMOB-hTERT cells for expanding and supporting CB CD34+ cells due to the ectopic expression of miR-22.
Materials and Methods
Cell cultures
Human CB CD34+ cells were collected, primarily using the MACS CD34+ Progenitor Cell Isolation Kit (Miltenyi Biotech, Inc., Bergisch Gladbach, Germany) as described previously. 25,26 FBMOB-hTERT, 25 cryopreserved primary BMSCs, 25 Phoenix, and 293T packaging cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco™; Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (FBS; Gibco™) at 37°C. Phoenix and 293T packaging cells were from American Type Culture Collection (ATCC, Manassas, VA). For osteoblastic differentiation, 8.0 × 105 primary BMSCs per well were cultured in six-well plates supplemented with 50 μg/mL ascorbic acid 2-phosphate, 10–8 M dexamethasone, and 10 mM glycerol 2-phosphate (Sigma-Aldrich, St. Louis, MO). After being cultured in differentiation medium for 3 days, the BMSCs that were subjected to osteoblastic differentiation (BMSCs-OD) were harvested for further experiments. This study was approved by the Ethics Committee of Tianjin Central Hospital. Prior to the study, subjects were informed of the objectives, requirements, and procedures of the experiments. All subjects gave informed written consent to participate in the study.
Cloning of human pre-miR-22 and β-catenin gene, derivative construction, and retroviral infection
The expression vector of pre-miR-22 (i.e., pCMV-pre-miR-22) was carried out by polymerase chain reaction (PCR) using the primers provided in Table 1. Next, the full-length of human pre-miR-22 was subsequently cloned into the retroviral vector. The Phoenix packaging cell line was co-transfected with retroviral plasmid and the helper plasmids in Lipofectanine 2000 (Invitrogen, Carlsbad, CA) to generate the viral supernatants. The expression vector of β-catenin (i.e., pLPC-β-catenin) was carried out by PCR using the primers provided in Table 1 and then subsequently cloned into the retroviral vector. The 293T packaging cell line was co-transfected with retroviral plasmid and the viral packaging plasmids containing gag-pol and env genes in Lipofectanine 2000 (Invitrogen). The viral supernatants were collected at 48 or 72 h after transfection to infect the indicated cells directly.
Primers used in this study
When the infected cells were incubated for about 5 h at 37°C, the infection medium was replaced with the fresh culture medium. After growing for about 24 h, puromycin (4 μg/mL) was added to the culture medium to select the infected cells. After 2–3 weeks of culture, the clones that are resistant to puromycin could grow out. The puromycin-resistant pool cells were expanded in fresh dishes for further experiments.
RNA extraction and quantitative real-time PCR
The total RNA was extracted with the total RNA kit I (Omega Bio-Tek, Inc., Norcross, GA). The first-strand cDNA was synthesized by the M-MLV First Strand cDNA Synthesis Kit (Omega Bio-Tek, Inc.) or the miRcute miRNA First-Strand cDNA Synthesis Kit (TIANGEN Biotech, Beijing, China). Quantitative real-time PCR (qRT-PCR) analysis was performed with the Super-Real PreMix Plus (SYBR Green) kit (TIANGEN Biotech) through ABI7500 (Applied Biosystems, Carlsbad, CA). U6 snRNA (for miRNA) or glyceraldehyde-3-phosphate dehydrogenase (GAPDH; for mRNA) was used as the endogenous control. All samples were done in triplicate. The relative expression was determined using the ΔΔCT method.
Antibodies and Western blotting
The indicated cells were washed twice with phosphate-buffered saline (PBS) and resuspended in RIPA buffer with protease inhibitor. The suspended substances were allowed to lyse on ice for 30 min. After lysis, the suspended substances were centrifuged at 15,000 g for 15 min at 4°C and then the supernatants were collected. The concentrations of protein were determined with a bicinchoninic acid protein assay kit (Pierce, Waltham, MA). Samples were analyzed by Western blotting using standard procedures. The membranes were probed with the antibodies below: GAPDH (Santa Cruz) and β-catenin (Cell Signaling Technology, Danvers, MA). The results were visualized with an enhanced chemiluminescence kit according to the manufacturer's instructions.
Co-culture of stromal cells with human CB CD34+ cells
The co-culture assay was performed as described previously. 25,26 Briefly, after being irradiated at a dose of 40 Gy, 5.0 × 105 indicated FBMOB-hTERT cells or control cells per well were seeded in 24-well plates overnight. 2.0 × 104 CB CD34+ cells per well were cultured in Iscove's modified Dulbecco's medium (IMDM; BioWhittaker™; Lonza, Basel, Switzerland) supplemented with 10% FBS, 10–4 M of 2-mercaptoethanol, 2 mM of L-glutamine, 5 mg/mL of insulin, 100 IU/mL of penicillin, and 100 mg/mL of streptomycin as well as a cytokine cocktail. The cytokine cocktail contained Flt ligand (10 ng/mL), SCF (10 ng/mL), thrombopoietin (10 ng/mL), and interleukin (IL)-6 (10 ng/mL; PeproTech, Inc., Rocky Hill, NJ). After 14 days of culture, the adherent hematopoietic cells that were loosely attached to the indicated FBMOB-hTERT cells or control cells and non-adherent hematopoietic cells were gently harvested by pipetting, counted, and analyzed for the expression of CD34 and CD38 with flow cytometry.
Long-term culture initiating cells assay
Long-term culture initiating cells (LTC-IC) were analyzed as described previously. 25,26 Briefly, the indicated and nearly confluent FBMOB-hTERT cells or control cells plated in six-well plates were irradiated at a dose of 40 Gy. Next, 1.0 × 104 CB CD34+ cells were plated onto the indicated monolayer in LTC medium (Myelo-Cult; STEMCELL Technologies, Inc., Vancouver, Canada) along with 10–6 M of hydrocortisone sodium hemisuccinate (Sigma-Aldrich). The LTC medium consisted of horse serum, FBS, 2-mercaptoethanol, and α-MEM. The fresh medium was added weekly to the cultured cells and half of the old medium was replaced. During 5–8 weeks of culture, both non-adherent and adherent cells were collected weekly and then cultured in the complete methylcellulose medium at 37°C with 5% CO2. After 14–16 days of culture, the LTC-IC activities were assayed by counting the colonies with >50 cells. According to the manufacturer's instructions (STEMCELL Technologies, Inc.), the frequency of LTC-IC was calculated by counting the number of colony-forming units (CFU-C), colony-forming unit-mix (CFU-Mix), and colony-forming unit-granulocyte-macrophages (CFU-GM).
Non-obese diabetic/severe combined immunodeficient disease repopulating cell assay
Non-obese diabetic/severe combined immunodeficient disease (NOD/SCID) repopulating cell (SRC) assay was carried out as described previously. 25,26 Briefly, after being co-cultured with the infected and irradiated FBMOB-hTERT cells or control cells for 4 weeks, all the CB hematopoietic cells were harvested as described above and injected intravenously (i.v.) into 8-week-old, sublethally irradiated (3.5 Gy) NOD/SCID mice. The mice that were transplanted with freshly isolated CD34+ cells were used as positive controls. After transplantation for 12 weeks, the BM mononuclear cells were collected from the recipient mice. The human cell reconstitution was assessed by staining the BM mononuclear cells using human CD45 and CD34 markers. Mouse IgG1 was used as the isotype control. The expression of genomic human 17α-satellite gene in the BM mononuclear cells was analyzed by PCR as a second test for the presence of human cells in the transplanted mice. All the animals were cared for and handled with the approval of Institutional Authority for Laboratory Animal Care of Tianjin Central Hospital.
Statistical analysis
The results are expressed as the mean ± standard deviation (SD) of three independent experiments. A two-tailed Student's t-test was used to compare the statistical difference.
Results
Expression of miR-22 in primary BMSCs, BMSCs-OD, and FBMOB-hTERT cells
To find out the function of osteoblastic miR-22 on hematopoiesis, the expression level of miR-22 was first analyzed in the primary BMSCs, BMSCs-OD, and FBMOB-hTERT cells. qRT-PCR revealed that miR-22 is endogenously expressed in these three cells. However, a significantly reduced level of miR-22 expression was observed in BMSCs-OD and FBMOB-hTERT cells (Fig. 1A) compared with that in primary BMSCs. The expression of miR-22 in CB CD34+ cells, which is also endogenously expressed, was also analyzed (data not shown). This is consistent with previous reports. 13,19 The significantly reduced expression of miR-22 in BMSCs-OD and FBMOB-hTERT cells suggests that osteoblastic miR-22 may have a key function in the normal development of hematopoiesis.

The endogenous expression of miR-22 in primary bone marrow stromal cells (BMSCs), BMSCs subjected to osteoblastic differentiation (BMSCs-OD), and hTERT-transduced fetal bone marrow osteoblasts (FBMOB-hTERT). (
Function of FBMOB-hTERT on CB CD34+ cells upon ectopic expression of miR-22 in vitro and in vivo
To find out the function of FBMOB-hTERT cells on expanding the CB CD34+ cells upon ectopic expression of miR-22, the overexpression model of miR-22 was established with FBMOB-hTERT cells through retroviral expression of pre-miR-22 (22/OE). The expression of miR-22 in the FBMOB-hTERT cells infected with the indicated virus was examined by qRT-PCR. The significantly upregulated expression level of miR-22 in 22/OE cells was observed compared with that in control cells, which were infected with retrovirus containing the empty intron sequence (Fig. 1B).
The ability of FBMOB-hTERT cells to expand CB CD34+ cells upon ectopic expression of miR-22 was first examined in vitro. 2.0 × 104 CB CD34+ cells were co-cultured with the irradiated 22/OE or control cells. All the hematopoietic cells were gently harvested after 14 days of co-culture. The numbers of total cells, CD34+ cells, CD34+CD38− cells, and CD34+CD38+ cells were analyzed and counted by flow cytometry. The ability of FBMOB-hTERT cells to expand CB CD34+ cells (16.92- vs. 23.35-fold) or CD34+CD38− cells (13.21- vs. 17.28-fold) was significantly decreased upon the ectopic expression of miR-22 (Table 2). The expansion results indicated that miR-22 from FBMOB-hTERT cells can inhibit the ability of FBMOB-hTERT cells to expand CB CD34+ or CD34+CD38− cells in vitro.
Ex vivo expansion of CB CD34 + cells over 14 days
After being co-cultured with the irradiated 22/OE cells or control cells for 14 days in vitro, the hematopoietic cells were collected and stained with CD34 and CD38 antibodies, and then analyzed by flow cytometry. The values mean the fold increase compared with the initial number of cells (2.0 × 104/well). The results are shown as mean ± standard deviation (SD) of six separate experiments.
p < 0.05 vs. control cells (n = 6).
The ability of miR-22 in FBMOB-hTERT cells to support the self-renewal and maintain the multipotent differentiation of CB CD34+ cells was further investigated by a LTC-IC assay. After being co-cultured with the irradiated 22/OE or control cells in LTC-IC medium for 5–8 weeks, all the hematopoietic cells were gently harvested and then subjected to a CFU assay. The numbers of the colonies with >50 cells were counted after being cultured in the complete methylcellulose medium for 14–16 days. As indicated in Fig. 2A, after being co-cultured with the irradiated 22/OE or control cells for 7 or 8 weeks, both the total number of CFUs and the number of CFU-Mix from the CB CD34+ cells co-cultured with 22/OE cells were significantly decreased compared with those from the CB CD34+ cells co-cultured with control cells. The number of CFU-GM from the CB CD34+ cells co-cultured with 22/OE cells became significantly decreased compared with those from the CB CD34+ cells co-cultured with control cells only after being co-cultured for 8 weeks. The results indicated that miR-22 from FBMOB-hTERT cells can inhibit the ability of FBMOB-hTERT cells to support the self-renewal and maintain the multipotent differentiation of CB CD34+ cells in vitro.
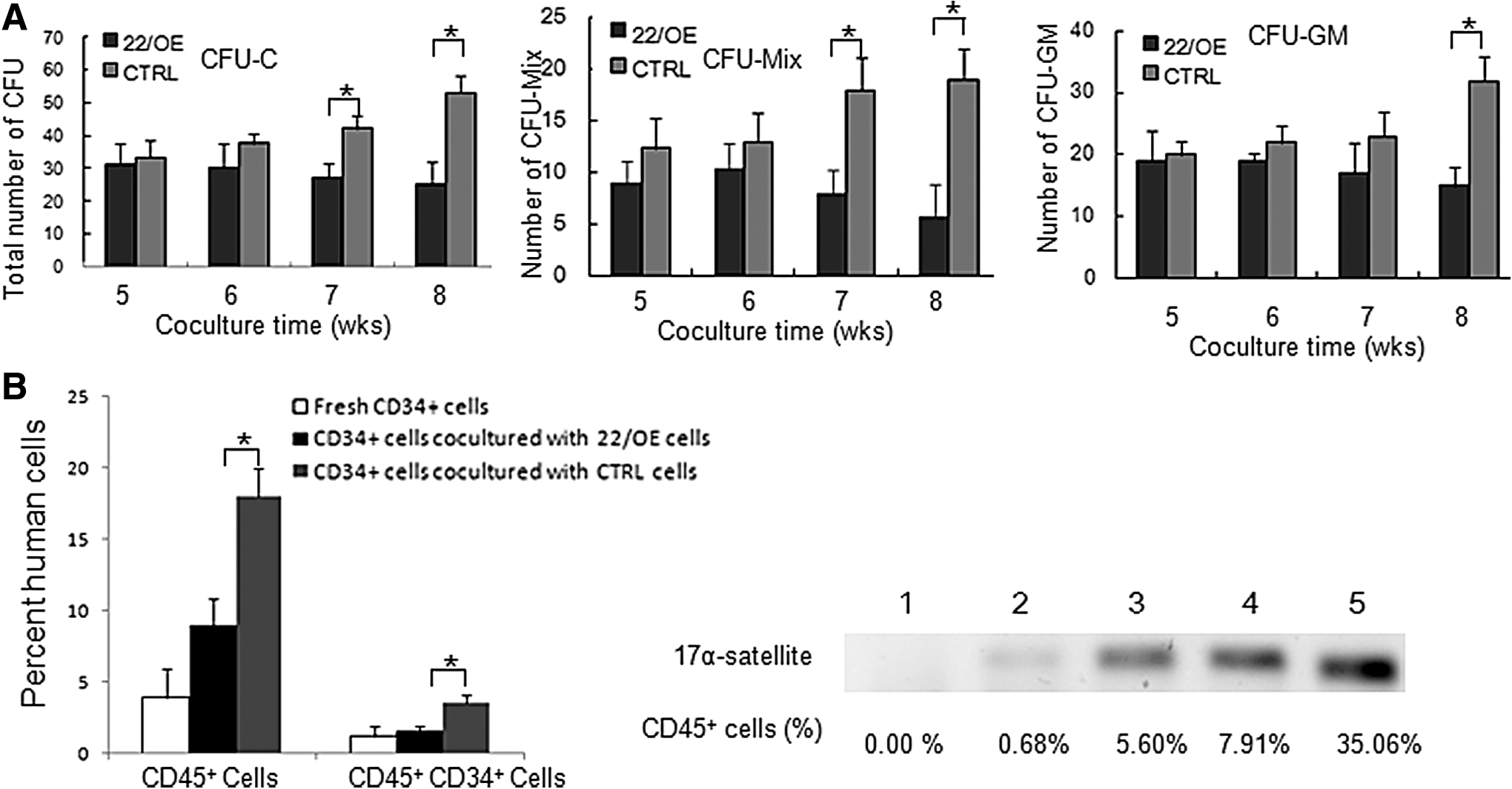
The function of FBMOB-hTERT cells upon ectopic miR-22 on CB CD34+ cells. (
To confirm the expansion and LTC-IC results in vitro further, the ability of engraftment of CB CD34+ cells after being co-cultured with 22/OE cells or control cells was investigated by using NOD/SCID mice. After being co-cultured with irradiated 22/OE cells or control cells for 4 weeks, all the hematopoietic cells were gently harvested and transplanted into the sublethally irradiated NOD/SCID mice. Human cell reconstitution was analyzed on the 12th week post transplant. The BM mononuclear cells were collected from the transplanted mice and analyzed with flow cytometry. The human 17α-satellite gene in the BM mononuclear cells was also examined by PCR. The mice transplanted with the fresh CB CD34+ cells were used as controls. As shown in Fig. 2B, left panel, the percentage of human CD45+ cells or CD45+CD34+ cells in BM mononuclear cells from recipient NOD/SCID mice transplanted with CB CD34+ cells co-cultured with 22/OE cells was significantly reduced compared with that from recipient NOD/SCID mice transplanted with CB CD34+ cells co-cultured with control cells. Even when the percentage of human CD45+ cells was 0.68%, human-specific 17α-satellite gene could also be detected in BM mononuclear cells from the recipient mice by PCR (Fig. 2B, right panel), which further confirmed the human cell reconstitution in the transplanted mice. These in vivo results are consistent with and were confirmed by the in vitro expansion data. Collectively, all the in vitro and in vivo data indicated that miR-22 in FBMOB-hTERT cells inhibit the ability of FBMOB-hTERT cells to expand human CB CD34+ cells and maintain the multipotency of CB CD34+ cells.
miR-22 inhibits the expression of β-catenin after being co-cultured with CB CD34+ cells
To find out the possible mechanisms through which miR-22 inhibits the ability of FBMOB-hTERT cells to expand human CB CD34+ cells and maintain the multipotency of CB CD34+ cells, the expression of several supporting molecules from hematopoietic niche including SCF, TGF-β, Ang-1, and β-catenin were investigated in 22/OE or control cells after being co-cultured with CB CD34+ cells. After interaction with or without 1.0 × 104 CB CD34+ cells for 24 h, the loosely adherent and non-adherent hematopoietic cells were washed off. The 22/OE cells or control cells were harvested and analyzed by qRT-PCR or Western-blotting to examine the expression of the indicated hematopoietic supporting molecules. After being co-cultured with CB CD34+ cells, the expression of β-catenin in FBMOB-hTERT cells was significantly reduced upon ectopic expression of miR-22 compared with that in control cells (Fig. 3A). After being co-cultured with CB CD34+ cells, the expression level of SCF in 22/OE cells was also significantly reduced compared with that in control cells, but it was weaker than β-catenin. After being co-cultured with CB CD34+ cells, the expression level of TGF-β and Ang-1 in 22/OE cells had a decreasing trend, but there was no statistical significance. Next, the protein level of β-catenin was examined by Western-blotting. As shown in Fig. 3B, the protein levels of β-catenin were significantly reduced in FBMOB-hTERT cells upon ectopic expression of miR-22 after interaction with CB CD34+ cells compared with those in control cells. Without being co-cultured with CB CD34+ cells, the expression levels of these molecules in FBMOB-hTERT cells upon ectopic expression of miR-22 had a decreasing trend, but there was no statistical significance (Fig. 3C). These data indicated that during the co-culturing with CB CD34+ cells, the expression of hematopoietic supporting molecules from the FBMOB-hTERT cells was changed upon ectopic expression of miR-22, which may explain the reduced ability of FBMOB-hTERT cells to support hematopoiesis upon the ectopic expression of miR-22.

MiR-22 inhibits β-catenin expression after being co-cultured with CB CD34+ cells. (
Function of 22/OE on CB CD34+ cells was partly rescued upon ectopic β-catenin
Since the expression of β-catenin in FBMOB-hTERT cells was significantly reduced upon ectopic expression of miR-22 during the co-culturing with CB CD34+ cells, this study further investigated whether the reduced production of β-catenin is responsible for the inhibited hematopoietic supporting ability of FBMOB-hTERT cells upon ectopic expression of miR-22. The overexpression model of β-catenin was obtained with 22/OE cells through infection of the virus stably expressing β-catenin, which were designated β-catenin/OE cells. 22/OE cells infected with virus containing the empty intron sequence were used as control cells. Western blotting was performed to confirm the expression level of β-catenin protein in the β-catenin/OE and control cells (Fig. 4A).
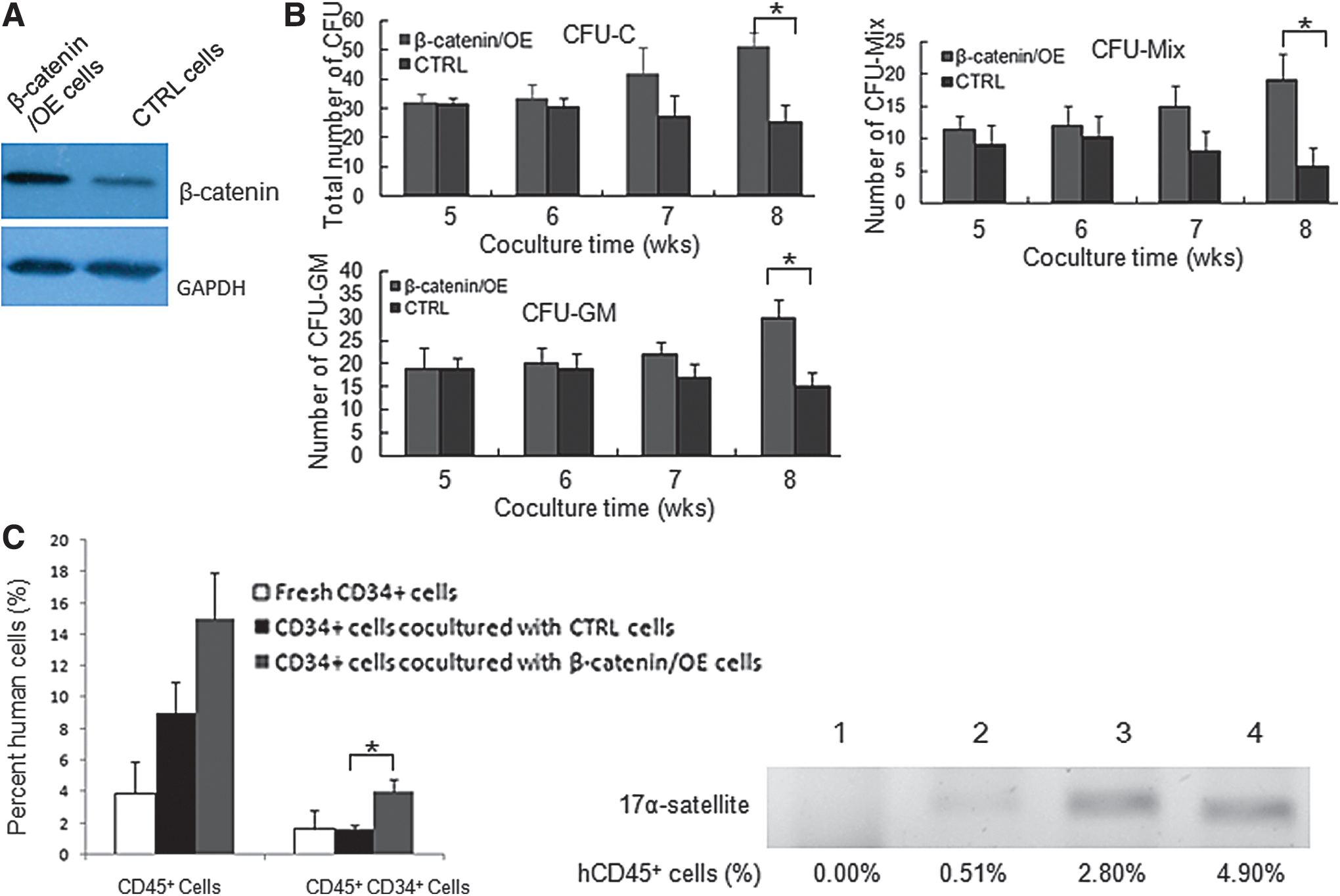
The function of 22/OE cells on CB CD34+ cells was partly rescued upon ectopic β-catenin. (
Using the obtained cells mentioned above, the expansion abilities CB CD34+ cells were examined upon ectopic expression of β-catenin. In vitro co-culture experiments (Table 3) indicated that the inhibited hematopoietic expanding ability of FBMOB-hTERT cells upon ectopic miR-22 was rescued, at least partly, upon ectopic expression of β-catenin (CD34+ cells: 21.99- vs. 17.01-fold; CD34+CD38− cells: 16.01- vs. 12.98-folds). The inhibited long-term hematopoietic supporting ability of FBMOB-hTERT cells upon ectopic expression of miR-22 was also rescued, at least partly, upon ectopic expression of β-catenin according to the LTC-IC assay. After being co-cultured for 8 weeks, the total number of CFUs, the number of CFU-Mix, and the number of CFU-GM (Fig. 4B) from CB CD34+ cells co-cultured with β-catenin/OE cells became significantly higher than those from CB CD34+ cells co-cultured with control cells. Consistent with the results in vitro, the engraftment ability of CB CD34+ cells co-cultured with 22/OE cells was also partly rescued upon ectopic expression of β-catenin. As shown in Fig. 4C, the percentage of human CD45+CD34+ cells in BM mononuclear cells from NOD/SCID mice transplanted with CB CD34+ cells co-cultured with 22/OE cells became significantly increased upon ectopic expression of β-catenin compared with that from the control group. Compared with that from the control group, the percentage of human CD45+ cells in BM mononuclear cells from NOD/SCID mice transplanted with CB CD34+ cells co-cultured with β-catenin/OE cells had a tendency to increase, but there was no significant difference. The expression of human-specific 17α-satellite gene in BM mononuclear cells from recipient mice was also examined by PCR to confirm the human cell reconstitution in transplanted mice. All these in vitro and in vivo data indicated that the decreased expression of β-catenin is, at least partly, responsible for the inhibited ability of FBMOB-hTERT cells to expand and support CB CD34+ cells upon ectopic expression of miR-22.
Ex vivo expansion of CB CD34 + cells over 14 days
The function of β-catenin/OE cells and CTRL cells on expanding CB CD34+ cells was assayed in vitro according to the methods described above.
p < 0.05 vs. control cells (n = 6).
Discussion
This study found that the expression level of miR-22 became significantly downregulated in the FBMOT-hTERT cells that have the characteristics of osteoblasts when compared with that in the primary BMSCs (Fig. 1A). The osteoblast is an important component of the BM microenvironment, and it plays a crucial role in regulating the development of hematopoiesis. 5,6 The FBMOB-hTERT cells, established in the lab, have the ability to expand CB CD34+ cells and support the multipotency of CB CD34+ cells in vitro and in vivo. 25 Studies by several groups have indicated that miRNAs not only affect the cellular constituents of hematopoietic cells, but also take part in the regulation of the hematopoietic microenvironment. 27 –30 Consistent with the present results, miR-22 has been shown to play a key role during the differentiation of osteoblasts. Huang et al. reported that upregulation of miR-22 can promote osteogenic differentiation and inhibit adipogenic differentiation of human adipose tissue–derived mesenchymal stem cells (MSCs). 23 However, Liang et al. showed that the expression of miR-22 was significantly downregulated during osteogenesis, which suggested that miR-22 may be a potential negative regulator of osteoblast differentiation. 24 The inconsistent results concerning the function of miR-22 on the differentiation of osteoblasts between the two groups may have resulted from the origin of the human MSCs used in their study. However, both of the studies suggested the key role of miR-22 on osteoblastic differentiation. To a certain extent, the present data are consistent with the reports from Liang et al., suggesting that miR-22 may be downregulated during the osteoblastic differentiation of primary BMSCs. The key molecules that were differently expressed during the osteoblastic differentiation of primary BMSCs may be necessary for the development of functional niche that affect HSCs and their progenitors. Therefore, in addition to the important function in osteoblastic differentiation, the significantly different expression of miR-22 between primary BMSCs and FBMOT-hTERT cells also suggested that osteoblastic miR-22 may be involved in normal hematopoiesis.
The in vitro and in vivo data indicated that ectopic miR-22 in FBMOB-hTERT cells can inhibit the ability of FBMOB-hTERT cells to expand CB CD34+ cells and maintain the multipotency of CB CD34+ cells (Fig. 2). Based on the aberrations in the miR-22-TET2 regulatory network in hematopoietic malignancies, Song et al. regarded miR-22 as a potent proto-oncogene. 21 Palacios et al. found that the activation of the phosphoinositide 3-kinase/AKT pathway by miR-22 resulted in chronic lymphocytic leukemia B-cell proliferation. 31 However, as the significantly downregulated expression level of miR-22 was found in de novo AML, Jiang et al. identified that miR-22 is an essential antitumor gatekeeper. 22 All of these data have shown that endogenous miR-22 in hematopoietic cells plays a key function in hematopoiesis. The present results suggest the inhibiting function of miR-22 from hematopoietic niche cells on hematopoiesis, which are in line with the findings from Jiang et al. to a certain extent. 22 The present data indicated that except for function of endogenous miR-22 on hematopoietic development and osteoblastic differentiation, miR-22 in osteoblasts from hematopoietic niche was also involved in the normal development of hematopoiesis.
This study further found that after being co-cultured with CB CD34+ cells, the expression of β-catenin in FBMOB-hTERT cells at the RNA and protein levels was significantly reduced upon ectopic miR-22 comparison with that in control cells (Fig. 3) Hematopoietic supporting molecules, including growth factors and membrane-bound factors, produced by the hematopoietic niche cells are important for their ability to support hematopoiesis. 32 –35 The hematopoietic supporting molecules are usually involved in the known pathways that control the normal development of hematopoiesis. In addition to the leading members of the Wnt signaling pathway, 36,37 β-catenin may also link to either E-cadherin or N-cadherin through their cytoplasmic tails. 38 –40 It is possible that through the interaction with the adherens, β-catenin influenced the hematopoiesis. It seems that the co-culture with CB CD34+ cells is important for the decreased expression of β-catenin upon ectopic expression of miR-22 because, without the existence of CB CD34+ cells, the expression level of β-catenin in FBMOB-hTERT cells upon ectopic expression of miR-22 had a trend to decrease, but there is no statistical significance compared to that in control cells (Fig. 3C). By transfecting hMSCs with miRNA mimics, Song et al. showed that miR-22 can significantly downregulate the expression of β-catenin at RNA and protein levels, even without the interaction with CB CD34+ cells. 24 Song et al. further found that β-catenin was a bona fide target for miR-22 through luciferase reporter assays in HEK293 cells. 24 Whether miR-22 can target β-catenin to inhibit its expression in FBMOB-hTERT cells upon co-cultured with CB CD34+ cells will be further explored in the authors' lab.
The in vitro and in vivo data further indicated that the inhibited ability of FBMOB-hTERT cells for expanding and supporting CB CD34+ cells upon ectopic expression of miR-22 can be, at least partly, rescued upon ectopic expression of β-catenin (Fig. 4). These results indicated that the decreased expression of β-catenin is, at least partly, responsible for the inhibited ability of FBMOB-hTERT cells to expand and support CB CD34+ cells upon the ectopic expression of miR-22. It is possible that the decreased ability of the FBMOB-hTERT cells to support hematopoiesis due to the ectopic miR-22 is caused by the disruption of adherens between FBMOB-hTERT cells and CB CD34+ cells. In addition to β-catenin, although weaker than that of β-catenin, the expression level of SCF in FBMOB-hTERT cells was also significantly reduced upon ectopic miR-22. Whether the inhibited ability of FBMOB-hTERT cells to expand and support CB CD34+ cells upon ectopic expression of miR-22 can be rescued upon ectopic SCF will be further examined in the authors' lab.
Conclusions
In conclusion, a potential function of miR-22 in osteoblasts from the BM microenvironment on the normal development of hematopoiesis was found in this study. By decreasing the expression of the hematopoietic supporting molecules, especially β-catenin, the ability of osteoblasts for expanding and supporting CB CD34+ cells can be inhibited upon ectopic expression of miR-22. For the first time, this study suggested that in addition to endogenous miR-22 functions on hematopoietic development and osteoblastic differentiation, miR-22 in osteoblasts from the BM microenvironment is also involved in regulating the development of normal hematopoiesis. Further investigation of the function of miR-22 in osteoblasts on hematopoiesis may raise the possibility that miR-22 has a wide range of functions on regulating the hematopoietic microenvironment.
Footnotes
Acknowledgment
This work was supported by Leading Academic Discipline Project of Beijing Education Bureau (BMU20110254).
Author Disclosure
No competing financial interests exist.