
Abstract
Application of oncolytic viruses is a valuable option to broaden the armament of anticancer therapies, as these combine specific cytotoxic effects and immune-stimulating properties. The self-replicating H-1 parvovirus (H-1PV) is a prototypical oncolytic virus that, besides targeting tumor cells, also infects endothelial cells, thus combining oncolytic and angiostatic traits. To increase its therapeutic value, H-1PV can be armed with cytokines or chemokines to enhance the immunological response. Some chemokines—more specifically, the CXCR3 ligands CXCL4L1 and CXCL10—combine immune-stimulating properties with angiostatic activity. This study explores the therapeutic value of recombinant parvoviruses carrying CXCL4L1 or CXCL10 transgenes (Chi-H1/CXCL4L1 or Chi-H1/CXCL10, respectively) to inhibit the growth of the human Kaposi sarcoma cell line KS-IMM. KS-IMM cells infected by Chi-H1/CXCL4L1 or Chi-H1/CXCL10 released the corresponding chemokine and showed reduced migratory capacity. Therefore, the antitumoral capacity of Chi-H1/CXCL4L1 or Chi-H1/CXCL10 was tested in mice. Either in vitro infected KS-IMM cells were injected or subcutaneously growing KS-IMM xenografts were treated by peritumoral injections of the different viruses. Surprisingly, the transgenes did not increase the antitumoral effect of natural H-1PV. Further experiments indicated that CXCL4L1 and CXCL10 interfered with the expression of the viral NS1 protein in KS-IMM cells. These results indicate that the outcome of parvovirus-based delivery of CXCR3 ligands might be tumor cell type dependent, and hence its application must be considered carefully.
Introduction
C
Among viruses described to display selective cytotoxicity toward tumor cells while posing a minimal treat to non-transformed cells, rodent parvoviruses have indeed demonstrated oncolytic potential in preclinical studies 1 –4 and in a Phase I clinical trial. 5 Parvoviruses cause tumor regression or eradication through various mechanisms ranging from direct oncolysis to enhancement of antitumor immune reactions. 6,7 Interestingly, the wild-type rat virus H-1PV also infects actively proliferating human endothelial cells 8 and thus may promote the collapse of the newly formed tumor vasculature without harming quiescent mature blood vessels. Furthermore, recombinant rodent parvoviruses expressing various cytokines/chemokines have proven to display improved efficiency of antitumoral activity in preclinical tumor models. 9 –12 Transduced by the minute virus of mice (MVMp), parvovirus-delivered CXCL10 previously showed preclinical therapeutic benefit in the treatment of highly malignant vascular tumors. 8 CXCL10 is a ligand of the CXCR3 chemokine receptor known to mediate angiostatic activity via its endothelial cell-expressed splice variant CXCR3B. 13 In addition, to the CXCR3A splice variant is ascribed a classic chemotactic and immunomodulatory function on activated T and natural killer (NK) cells. 14 Antitumoral efficiency of CXCL10 delivered by MVMp was also demonstrated in mouse glioblastoma. 11 Yet, in this model, the antitumor effects observed with the parvovirus expressing CXCL10 were likely due to the chemotactic activity of the chemokine rather than its angiostatic properties. In contrast, treatment of human pancreatic cancer cells with an H-1PV-based vector expressing CXCL10 did not suppress their growth. 12 Therefore, the antitumoral activity of viral-encoded CXCL10 remains controversial and might be tumor model dependent. Further research is needed to clarify the potential benefit of viral delivery of recombinant CXCL10.
On the other hand, the antitumoral potential of other CXCR3 ligands delivered by recombinant viruses has also been explored by various groups using alternative viral vectors. In this respect, CXCL4-based virotherapy has shown angiostatic and antitumoral efficiency both in vitro and in vivo. 15 –17 The antitumoral activity of the non-allelic variant of CXCL4, namely CXCL4L1, repeatedly shown to be a more potent angiostatic chemokine, 18,19 had, however, not been previously evaluated within the context of virotherapy. It was recently demonstrated that recombinant H-1PV viruses were able to drive in vitro production of the recombinant angiostatic chemokine CXCL4L1 similar to CXCL10. 7
In the present study, the properties of these H-1PV-based recombinant parvoviruses (renamed here Chi-H1/CXCL4L1 and Chi-H1/CXCL10) were further evaluated in vitro. Their potential to infect transformed human cells of endothelial origin, namely Kaposi sarcoma cells (KS-IMM), and ensuing expression of recombinant chemokines was confirmed. The inhibitory activity of Chi-H1/CXCL4L1 on the migration of the endothelial Kaposi sarcoma cells was illustrated. Angiostatic activity of the different parvoviruses was further demonstrated in vivo in Kaposi sarcoma-bearing mice. However, additional benefit of the inserted transgenes on disease progression remains unclear in this particular tumor model. Indeed, the data suggest that the CXCR3 ligands might interfere with the parvovirus cycle, thereby reducing rather than enhancing the overall antitumoral effect. Although the future of anticancer therapy definitely lies with combination strategies, it seems that these particular angiostatic chemokines are not compatible with parvoviral suppressive activity on vascular KS-IMM tumors. The present results indicate that tumoral expression of CXCR3 may represent a molecular determinant for the efficiency of infection by recombinant parvoviruses carrying CXCR3 ligands.
Materials and Methods
Cell culture
A Kaposi sarcoma cell line (KS-IMM) 20 was kindly provided by A. Vecchi (Instituto Clinico Humanitas, Rozzano, Italy). Like the transformed human 293T/17 cells (ATCC, Manassas, VA), KS-IMM were cultured in Dulbecco's modified Eagle's medium (DMEM; Sigma–Aldrich, Taufkirchen, Germany) supplemented with 2 mM of L-glutamine, 100 IU/mL of penicillin, 100 μg/mL of streptomycin (Life Technologies, Carlsbad, CA), and 10% (v/v) heat-inactivated fetal calf serum (hi-FCS). The SV40-transformed human newborn kidney NB-324K cells 21 were grown in Eagle's minimal essential medium (MEM; Sigma–Aldrich) supplemented with 2 mM of L-glutamine, 100 IU/mL of penicillin, 100 μg/mL of streptomycin, and 5% (v/v) hi-FCS.
Production, purification, and titration of recombinant virus
The H-1PV-based parvoviral vectors—namely, Chi-H1/Δ800, Chi-H1/CXCL4L1, and Chi-H1/CXCL10—were obtained as previously published. 7,12 The enhanced green fluorescent protein eGFP cDNA was cloned as a Sac1-Not1 fragment into the multiple cloning site of pChi-H1/Δ800 creating pChi-H1/eGFP. 12 Recombinant virus stocks were produced and purified as described previously. 12 Virus stocks were titrated by infected cell hybridization assays and expressed as replication units (RU) per mL of virus suspension. 9 For further experiments, multiplicities of infection (MOI) are expressed as RU/cell.
In vitro virus infection
Adherent cells were inoculated with virus 1 day after seeding. Growth medium was replaced with virus suspension diluted in DMEM without further supplements. Cells were exposed to this inoculum for 1 h, incubated at 37°C, while plates were gently rocked every 10 min. Mock-treated cultures were simultaneously incubated with DMEM. Afterwards, the inoculum was removed, and fresh growth medium was added to the cells. The fresh medium could also be supplemented with purified recombinant human chemokines, either CXCL4L1 (produced by recombinant baculovirus-infected insect cells and purified to homogeneity as previously described) 22 or CXCL10 (PeproTech, Rocky Hill, CT), in order to study the effect of chemokine stimulation on Chi-H1/Δ800 virus infection.
Immunoblotting
KS-IMM cells (2 × 105) were mock-treated or infected with Chi-H1/Δ800, Chi-H1/CXCL4L1, or Chi-H1/CXCL10 at an MOI of 12 RU/cell as described above, and harvested 2 days post infection (p.i.). After cell lysis in RIPA buffer (150 mM of NaCl, 10 mM of Tris, 1 mM of EDTA, 1% NP-40, 0.5% Na deoxycholate, 0.1% SDS, complete mini protease inhibitor [Roche, Mannheim, Germany], pH 7.5) and protein quantification, 20 μg of total proteins was separated by SDS-10% polyacrylamide gel electrophoresis (SDS-PAGE) and electro-transferred to Protran nitrocellulose membranes (GE Healthcare, Dornstadt, Germany). The membranes were incubated with rabbit polyclonal antiserum directed against NS1 (MK3, kindly provided by N. Salomé, INSERM U1109, Strasbourg, France), or mouse monoclonal anti GAPDH antibody (Santa Cruz Biotechnology, Heidelberg, Germany), and the appropriate peroxidase-coupled goat antibody (Dianova, Hamburg, Germany). Immunoreactive proteins were revealed by enhanced chemiluminescence (Western-Lightning Plus-ECL, PerkinElmer, Rodgau, Germany). The intensity of the signals was quantified using a Chemo Cam Imager 3.2 (Intas, Göttingen, Germany). Alternatively, the membranes were exposed to high-performance chemiluminescence films (Amersham Hyperfilm™ ECL; GE Healthcare).
Quantification of transgene expression
Levels of secreted cytokines were measured in the cell culture supernatants of recombinant H-1PV-infected NB-324K and KS-IMM cells using specific enzyme-linked immunosorbent assays for CXCL10 or CXCL4L1. 7 eGFP expression was visualized 2 days p.i. using a fluorescence microscope (BZ-9000; Keyence, Neu-Isenburg, Germany).
Wound healing assay
KS-IMM cells were seeded in a 24-well plate (1 × 105 cells/well). The following day, confluent monolayers were scarred reproducibly (drawing a straight line with a 1 mL pipette tip). After washing twice with phosphate-buffered saline (PBS), the cells were inoculated for 2 h with 200 μL of virus suspension at a MOI of 12 or mock (buffer)-treated. Complete growth medium was then added for 2 days. Mock-treated cultures were either incubated with complete medium or medium supplemented with 50 ng of purified recombinant CXCL4L1. Two days p.i., cultures were fixed with CH3OH for 3 min and stained with hematoxylin and eosin, both for 2 min. Wound healing was monitored microscopically at a 10-fold magnification.
Tumor model
Female nude Balb/c mice (Janvier Labs, St Berthevin, France) were housed under pathogen-free conditions in individually ventilated cages within the central animal facility. At the age of 7–8 weeks, the mice were implanted subcutaneously into the right flank with 3 × 106 KS-IMM cells suspended in ice-cold calcium-, magnesium-, and glucose-containing Dulbecco's PBS mixed with liquid Matrigel (BD Biosciences, Heidelberg, Germany) to a final volume of 200 μL. KS-IMM cells were treated in vitro before implantation with buffer (mock-treated) or infected with recombinant H-1PV with a MOI of 12 RU/cell. Alternatively, established tumors were injected peritumorally on days 4, 7, and 11 post implantation with buffer (mock) or recombinant H-1PV (7 × 106 RU/injection) as a single treatment or in combination with peritumoral injection of 50 ng of purified recombinant CXCL4L1 protein. Tumor growth was monitored externally using an electronic digital caliper (Farnell, Aschheim, Germany) three times a week. Tumor volume was calculated according to the formula: V = 1/2 × l × w 2 (mm3). Animals were sacrificed for ethical reasons when the tumor volume reached 1,500 mm3. All animal experimental procedures were approved by the responsible animal protection officer at the German Cancer Research Center and the regional council, according to the German Protection Law.
Gene expression studies
Total RNA was isolated from resected tumors using ceramic bead dissociation (Precellys; Bertin Technologies, Rockville, MD) and Trizol Isolation Reagent (Invitrogen, Merelbeke, Belgium) according to the manufacturer's instructions. RNA (1 μg) was treated with DNAse I (Invitrogen) and reverse transcribed (1 h at 37°C) in a reaction mix containing 200 IU of RNA-dependent DNA polymerase M-MLV RT (Promega, Madison, WI), 500 μM of dNTPs (Invitrogen), 20 IU of RNasin, and 500 ng of random primers (both Promega) in a total reaction volume of 25 μL. Relative changes in gene expression within the tumor and tumor microenvironment were evaluated by quantitative polymerase chain reaction (qPCR), as previously described. 23 Primers and probes used are listed in Table 1 (Integrated DNA Technologies, Haasrode, Belgium). Obtained CT were processed following the 2−ΔΔCT method, with 18S ribosomal 1 RNA serving as an internal control. 24
Quantitative polymerase chain reaction assays: primer and probe sequences for murine endothelial markers
All primer/probe sets were supplied as PrimeTime quantitative polymerase chain reaction assays by Integrated DNA Technologies.
Statistical analysis
The Mann–Whitney test was used as a non-parametric test to compare data from two independent groups, and was performed using Statistica 12. Data represent the mean ± standard error of the mean (SEM), unless stated otherwise. A p-value of <0.05 was considered to indicate a statistically significant difference. The number of independent experiments was indicated by n.
Results
Recombinant Chi-H1 viruses infect KS-IMM cells successfully and thereby instigate vector-encoded recombinant protein expression
To confirm successful transduction by recombinant virus stocks upon infection, KS-IMM cells were infected in vitro with Chi-H1/CXCL4L1 and Chi-H1/CXCL10 at different MOI, and production of the secreted chemokines was monitored with time in the cell culture medium (Fig. 1). Concentrations of CXCL4L1 and CXCL10 peaked on the second day (MOI of 12 RU/cell), as measured in daily refreshed culture medium (Fig. 1A and C). In comparison, transgene expression was also evaluated in vector-infected NB-324K cells, the reference human cell line that is fully permissive to H-1PV infection. Although NB-324K cells efficiently expressed viral vector-encoded chemokines at low MOI (1 RU/cell), KS-IMM did release similar chemokine levels of CXCL10, though at a higher MOI (12 RU/cell; Fig. 1C). However, CXCL4L1 release by KS-IMM cells infected at a MOI of 12 RU/cell with Chi-H1/CXCL4L1 was reproducibly lower than after infection of NB-324K cells with this virus (Fig. 1A). Similar observations were made when evaluating accumulation of recombinant chemokines produced over 3 days (Fig. 1B and D). No CXCL4L1 or CXCL10 was detected in the culture medium of cells that were mock (buffer)-treated or inoculated with the control virus Chi-H1/Δ800. It is worth mentioning that the cumulated levels of CXCL10 secreted by Chi-H1/CXCL10-infected KS-IMM cells were often slightly higher than the sum of the daily production levels. Since no increase of CXCL10 was observed after infection of these cells with control (empty) vector, the levels of CXCL10 are likely resulting from some autocrine CXCL10 production by Chi-H1/CXCL10-infected KS-IMM cells.
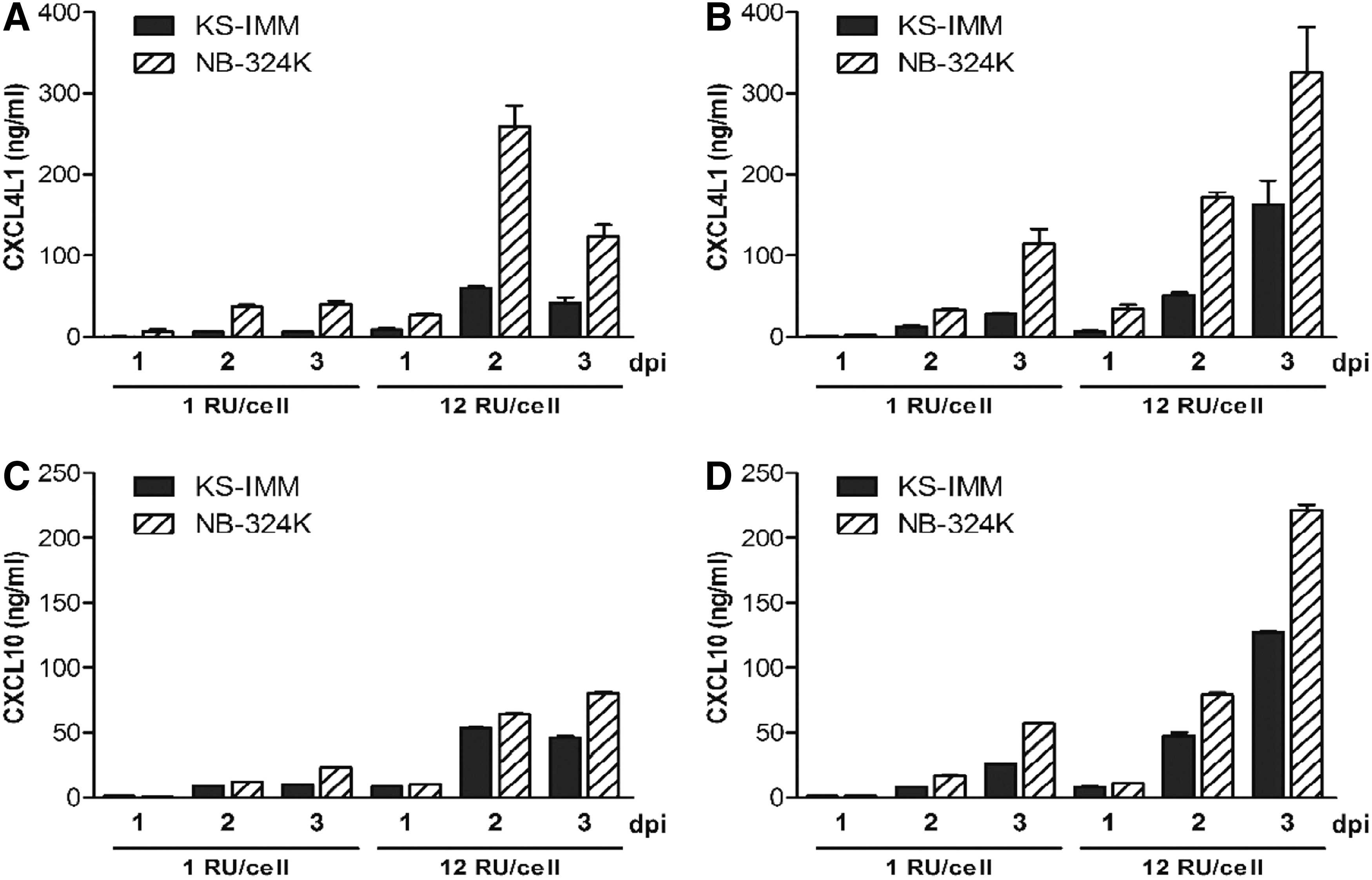
Expression of recombinant chemokines by KS-IMM or NB-324K cells after Chi-H1/CXCL4L1 or Chi-H1/CXCL10 virus inoculation. KS-IMM and NB-324K cells were infected with Chi-H1/CXCL4L1 (
On the other hand, cell-dependent susceptibility to viral infection and subsequent transgene expression was also validated after infection with Chi-H1/eGFP, a H-1PV-based vector that transduces a reporter gene, namely the green fluorescent protein. As shown in Fig. 2, the transduction efficacy of the vector is much lower in KS-IMM cells compared with the NB-324K cells, which were tested in parallel, as can be deduced from the low number of green fluorescent cells at a MOI of 3, and even more evidenced at the higher MOI of 12. Indeed, virus infection at a MOI of 3, 6, or 12 RU/cell resulted in 37%, 86%, and 84% of transduced NB-324K cells and 3%, 4%, and 5% of transduced KS-IMM cells, respectively. Since KS-IMM cultures were continuously growing, irrespective of whether they were mock-treated or virus-infected, the lower transduction efficiency in KS-IMM cells compared with NB-324K cells is most likely due to the resistance of KS-IMM cells to parvovirus infection rather than higher toxicity of the virus in these cells.

Expression of enhanced green fluorescent protein (eGFP) by Chi-H1/eGFP-infected KS-IMM or NB-324K cells. KS-IMM and NB-324K cells were infected with recombinant Chi-H1/eGFP parvoviruses at different MOI (expressed as RU per cell). Mock-treated cells were not exposed to the recombinant virus, only to buffer. The expression of recombinant GFP was visualized 2 d.p.i. Pictures were taken at 63 × magnification.
Chi-H1/CXCL4L1 inhibits KS-IMM wound healing
In order to confirm the angiostatic ability of virus-encoded CXCL4L1 on the transformed endothelial KS-IMM cells, similar to the activity previously attributed to the purified recombinant chemokine 18,25 or conditioned medium, 7 in vitro healing of a KS-IMM monolayer was evaluated. Figure 3 shows residual scarring 2 days after creation of an initial wound and virus inoculation, and thus represents the inhibitory effect of infection on the ability of cultured KS-IMM to restore the monolayer efficiently. As confirmed in three independent experiments, testing all conditions in duplicate, the virus itself mildly inhibited full recovery of the confluent monolayer, as shown upon Chi-H1/Δ800 infection. More importantly, both recombinant viruses Chi-H1/CXCL4L1 and Chi-H1/CXCL10 enhanced this anti-migratory activity. Inhibition of wound healing as seen after inoculation with Chi-H1/CXCL4L1 is similar to the inhibitory effect observed with purified recombinant CXCL4L1 (50 ng/mL).
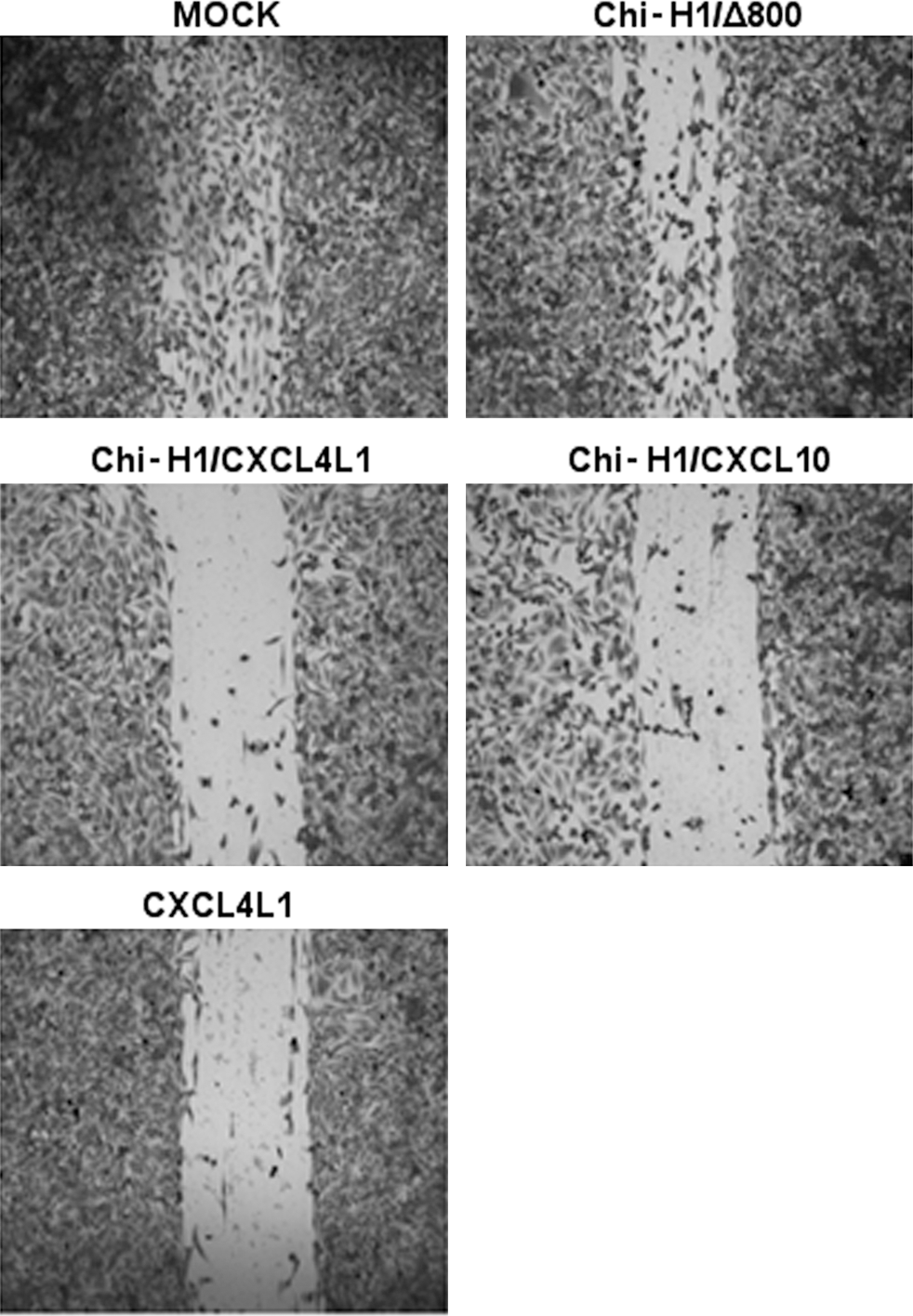
Migration of parvovirus-infected KS-IMM in a wound-healing assay. The ability of KS-IMM monolayers to restore a wound within 2 days was evaluated under different conditions. Directly following scarring of the monolayer, KS-IMM cells were either treated with buffer (mock) or inoculated with Chi-H1/CXCL4L1 or Chi-H1/CXCL10. Alternatively, mock-treated monolayers were incubated with 50 ng of purified recombinant CXCL4L1. Pictures were taken at 10 × magnification. One representative experiment out of three is shown.
Virus-encoded chemokines prove incapable of efficiently improving the antitumoral activity of H-1PV on KS-IMM tumors
The oncolytic activity of H-1PV combined with the angiostatic activity of the viral vectors could be useful as an alternative, innovative cancer therapy. Considering the angiostatic potential of the encoded chemokines, a tumor model of endothelial origin was opted for, namely KS-IMM, shown to be susceptible in vitro to rodent parvovirus infection when high doses of viruses are used. 7 The overall tumor progression was indeed inhibited by in vitro treatment with the virus Chi-H1/Δ800 (Fig. 4A, ▴symbols) compared with the mock-treated group (Fig. 4, × symbols), and, as a consequence, a prolonged survival time (53 vs. 36 days) of the animals was observed in Chi-H1/Δ800-treated group (Fig. 4B). An in vivo experiment was also performed with the Δ800 recombinant virus, alongside wild-type parvovirus (data not shown) and it was observed that the Δ800 is as effective as the wild-type virus in inhibiting KS-IMM tumor growth. When the treatment with Chi-H1/CXCL4L1 or Chi-H1/CXCL10 viruses was applied in established tumors (Fig. 4C and D), no significant difference was observed in tumor growth between mock-treated and vector-treated tumors, except at early stages of tumor development, as shown in Fig. 5. Figure 5 shows the relative increase in tumor size at day 11 p.i. compared with day 4 p.i. A slightly, though significantly reduced, tumor growth was observed when tumors were inoculated with Chi-H1/Δ800 compared with mock treatment (Fig. 5; 169 ± 46% growth, n = 8 reduced to 65 ± 13% growth, n = 6, p = 0.045). However, no added preclinical benefit, on either tumor growth or survival, was observed after treatment of KS-IMM tumors with Chi-H1/CXCL4L1 or Chi-H1/CXCL10 compared to the vector Chi-H1/Δ800 (Figs. 4 and 5). Expression of the transgene was verified by qPCR 3 days after peritumoral injection of the recombinant viruses. CXCL4L1 and CXCL10 mRNA transcripts were detected in tumors treated with the corresponding recombinant viruses (data not shown). Mock-treated tumors or Δ800-treated tumors did not express CXCL4L1 or CXCL10 mRNA. This demonstrates that the implanted tumor cells are infected by the recombinant viruses after peritumoral injection. Surprisingly, production of chemokines rather seemed to moderate the antitumoral benefit of the virus. This was corroborated by including treatment of tumor-bearing mice with the Chi-H1/Δ800 virus in the presence of purified recombinant CXCL4L1 protein (Figs. 4E and F and 5). In contrast, sole administration of purified recombinant CXCL4L1 did show significant antitumoral activity in the studied tumor model (Fig. 5; 24 ± 15% growth, n = 6, p = 0.016). These apparent discrepancies in antitumoral activity (i.e., on KS-IMM tumor growth) between virus-transduced and the solely administered protein CXCL4L1 prompted an investigation as to whether the vector-transduced CXCL4L1 and CXCL10 could inhibit tumor neo-angiogenesis driven by murine cells. Therefore, the expression of markers specific for murine endothelial cells was analyzed in tumor samples.
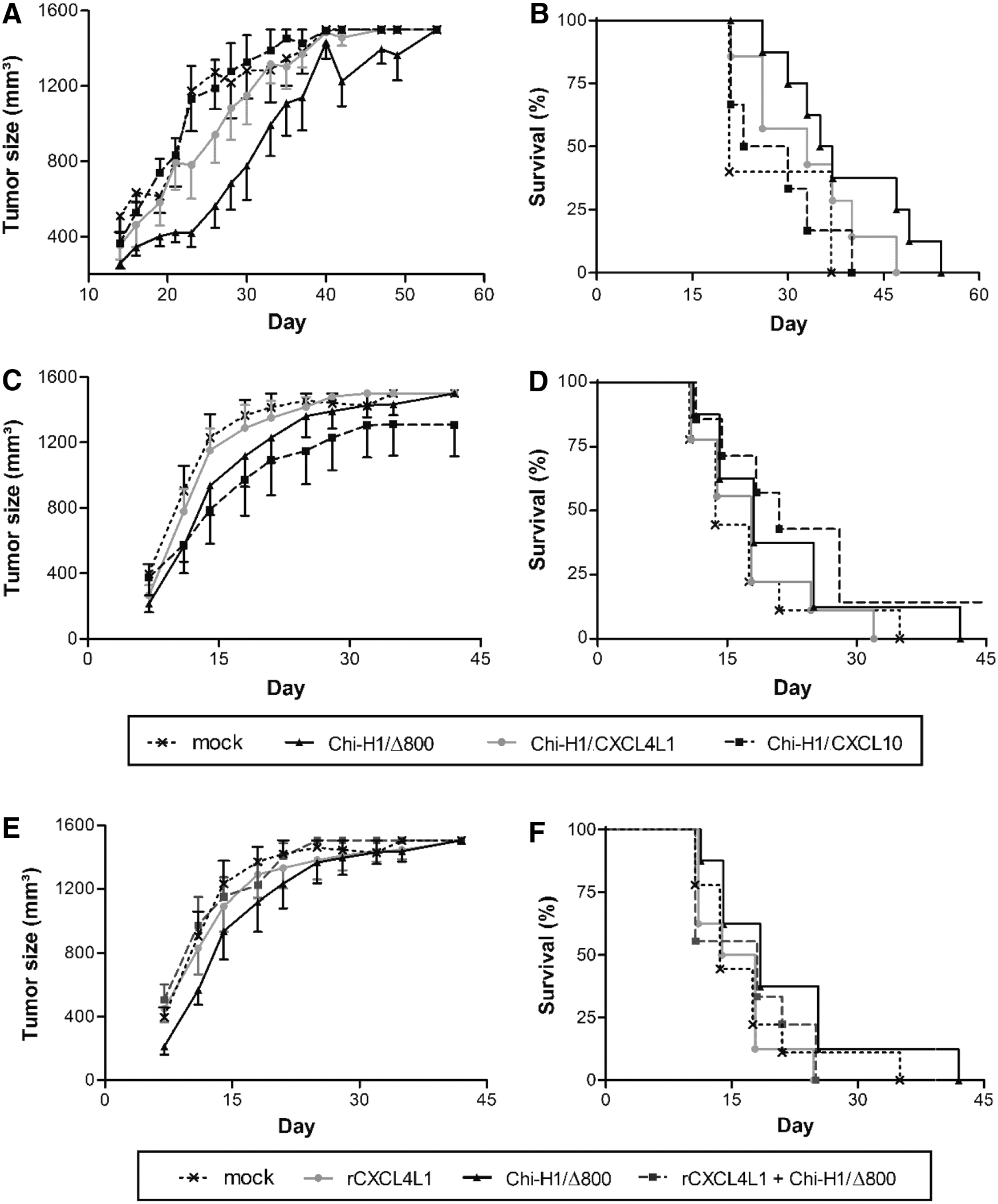
Tumor growth kinetics and survival of mice after in vitro infection or peritumoral injection with recombinant parvoviruses. KS-IMM were treated with Chi-H1/Δ800, Chi-H1/CXCL4L1, or Chi-H1/CXCL10, either pre (
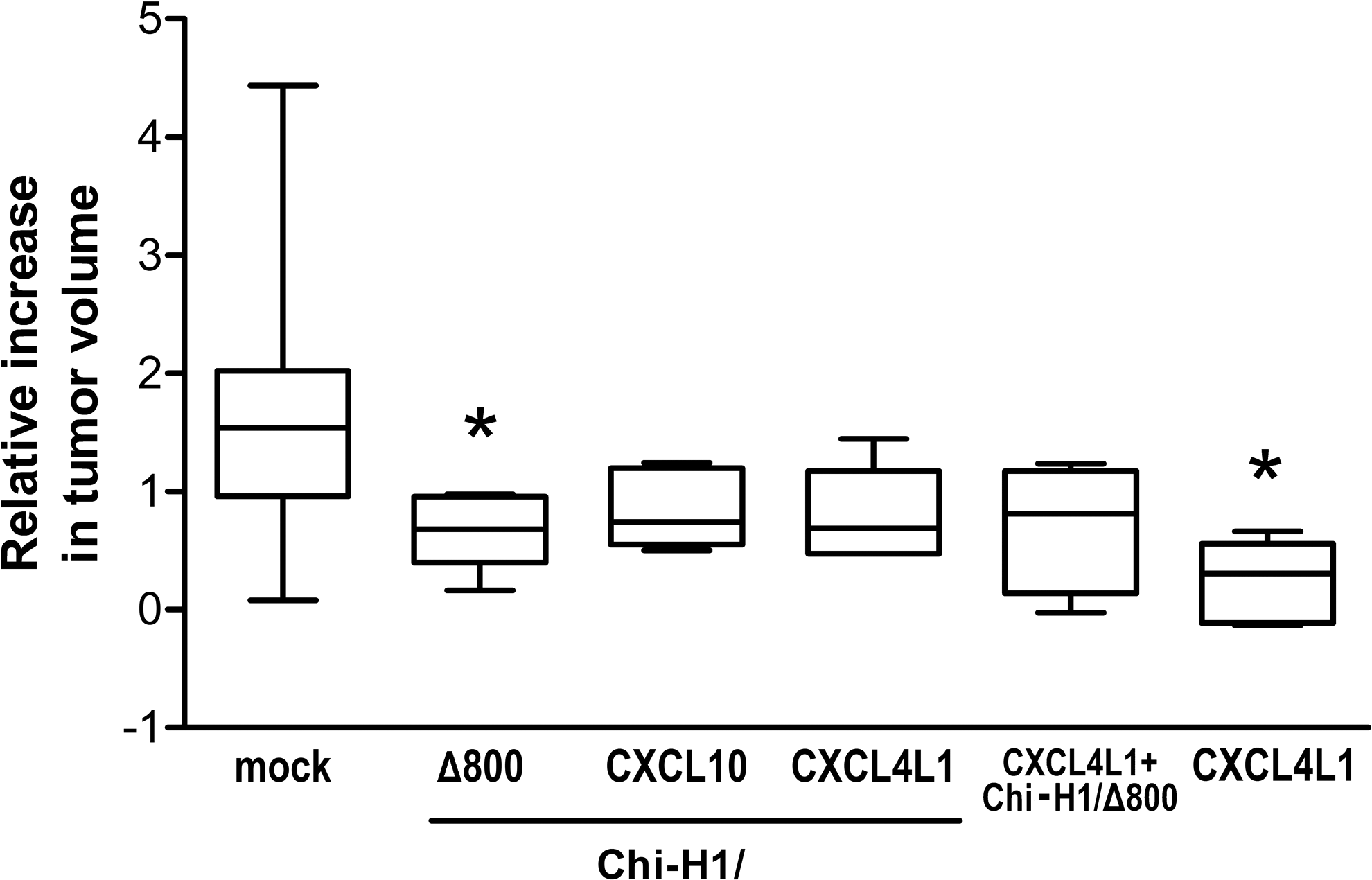
Relative incremental tumor change in peritumorally in vivo treated KS-IMM tumors. Subcutaneously growing KS-IMM tumors were treated with Chi-H1/Δ800, Chi-H1/CXCL4L1, or Chi-H1/CXCL10 on day 4, 7, and 11 post implantation (see also Fig. 4C and D). The relative increase in tumor size was calculated as (V day 11 − V day 4)/V day 4 in which the tumor volume on day 11 (V day 11) is compared against and corrected for the original tumor volume at the start of treatment, namely 4 days after implantation (V day 4). Relative changes in tumor size are depicted as a 10–90th percentile Whiskers plot, in which the horizontal line marks the median value. n = 4–8; *p < 0.05 (vs. mock).
Recombinant parvoviruses transducing CXCL4L1 or CXCL10 reduce murine expression of endothelial markers in the vicinity of KS tumors
In order to evaluate the in vivo angiostatic effect of peritumorally administered viruses, qPCR was performed on tumors resected at early stages of development, collected on day 7 or day 11 post implantation. The intratumoral expression of murine blood and lymphatic endothelial markers, PECAM1 and LYVE-1, respectively, was quantified (Fig. 6A and B). Compared with control (mock-treated) tumors, both markers were significantly downregulated upon Chi-H1/CXCL4L1 inoculation (62 ± 9%, p = 0.007, n = 7, and 46 ± 12%, p = 0.011, n = 7, for PECAM1 and LYVE-1, respectively), similar to the reduction observed after administration of purified recombinant CXCL4L1 (65 ± 13%, p = 0.030, n = 7, and 47 ± 12%, p = 0.021; n = 7, respectively). LYVE-1 expression was also reduced by Chi-H1/Δ800 (54 ± 10%, p = 0.018, n = 6) or Chi-H1/CXCL10 treatment (42 ± 6%, p = 0.003, n = 7). However, none of the recombinant viruses induced a reduction that was significantly different from the effect of Chi-H1/Δ800 on LYVE-1 expression.
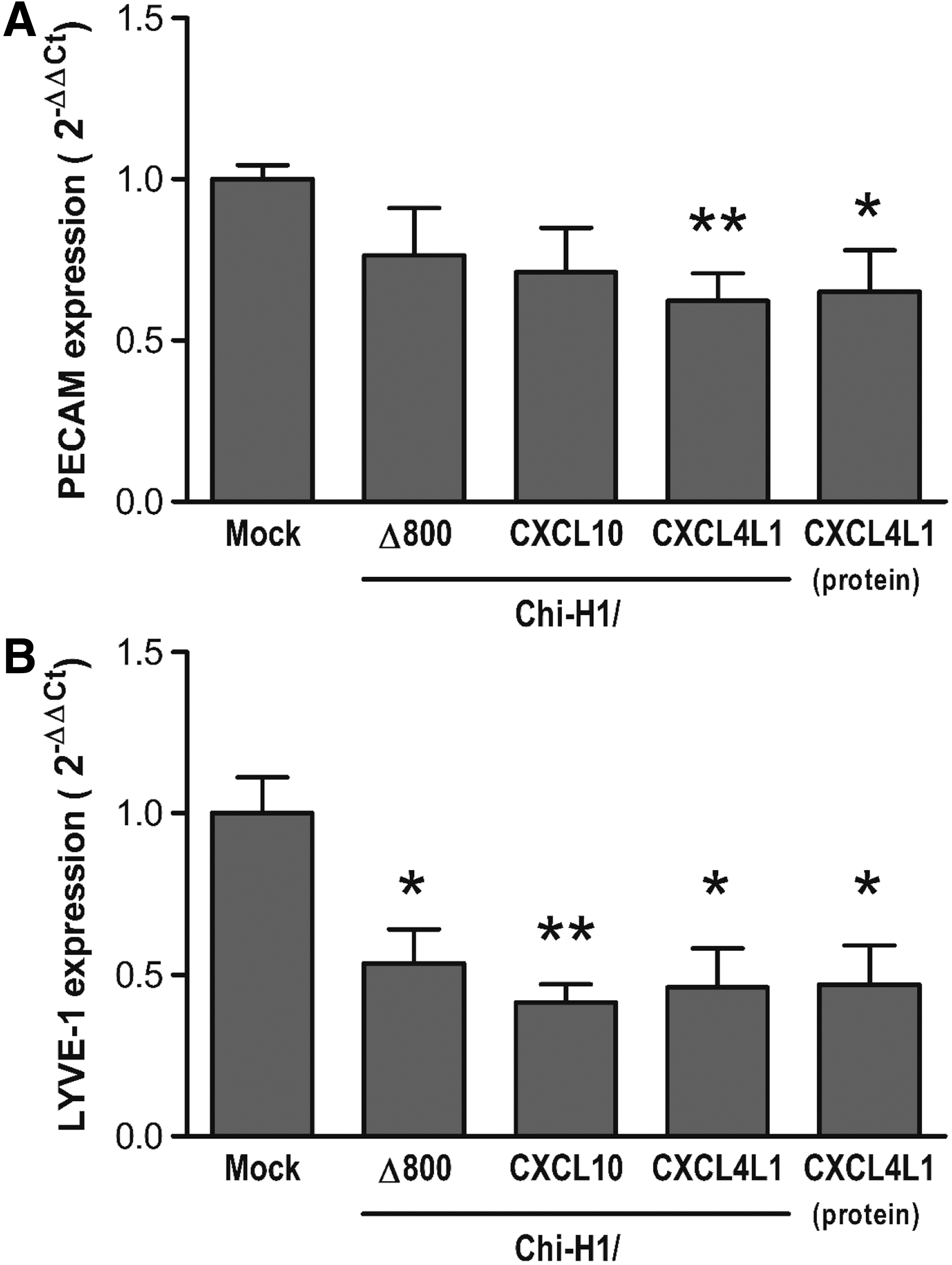
In vivo angiostatic activity of peritumorally administered recombinant parvoviruses. Relative expression of murine endothelial and angiogenic markers was evaluated by quantitative polymerase chain reaction on mRNA isolated from resected KS-IMM tumors that were peritumorally injected with Chi-H1/Δ800, Chi-H1/CXCL10, Chi-H1/CXCL4L1 (7 × 106 RU/injection), or with purified CXCL4L1 (50 ng). Tumors were collected on day 7 or day 11 post implantation (i.e., 3 days after the first peritumoral injection or 4 days after the second injection, respectively). Fold changes in mRNA levels (mean ± SEM) were calculated according to the 2−ΔΔCt method. n = 6–7; *p < 0.05; **p < 0.01 (vs. mock).
CXCL4L1 and CXCL10 interfere with the expression of viral proteins in KS-IMM cells
KS-IMM cells express CXCR3, known as the unique receptor for CXCL4L1 and CXCL10. Having shown the angiostatic activities of virus-transduced CXCL4L1 and CXCL10, next the study aimed to determine why the transduced chemokines did not improve the tumor suppressive activity of the virus alone. It was hypothesized that CXCL4L1 and CXCL10 might interfere with the viral cycle, maybe in an autocrine way. To test this possibility, the expression of the viral protein NS1, a key factor for the viral cycle and the major determinant of viral cytotoxicity, was analyzed by Western blotting. KS-IMM cells (2 × 105) were infected with Chi-H1/CXCL4L1, Chi-H1/CXCL10, or Chi-H1/Δ800 at a MOI of 12 RU/cell and harvested 2 days p.i. Figure 7A shows that the accumulation of NS1 was dramatically reduced when expressed by either Chi-H1/CXCL4L1 or Chi-H1/CXCL10 (60–70% reduction) compared with NS1 accumulation after infection with the control Chi-H1/Δ800 (referred to as 100%). This was confirmed at the mRNA level by qPCR in kinetics experiments showing that at day 1 p.i., the levels of NS1 mRNA were similar in control Chi-H1/Δ800 and in cells infected with either Chi-H1/CXCL10 or (even slightly higher) with Chi-H1/CXCL4L1, while from day 2 p.i., its levels dropped dramatically to 25% of the control in CXCL10 and in CXCL4L1-infected cells (data not shown). This suggests that the transduced chemokines may inhibit some step(s) of the viral cycle, resulting in decreased accumulation of NS1. Indeed, recombinant CXCL10 and CXCL4L1 had a similar, though weaker, inhibitory activity than the recombinant viruses (Fig. 7B). Which step(s) of the viral cycle (DNA amplification, gene expression, protein stability and/or activity) is (are) affected by the chemokines remains to be determined.
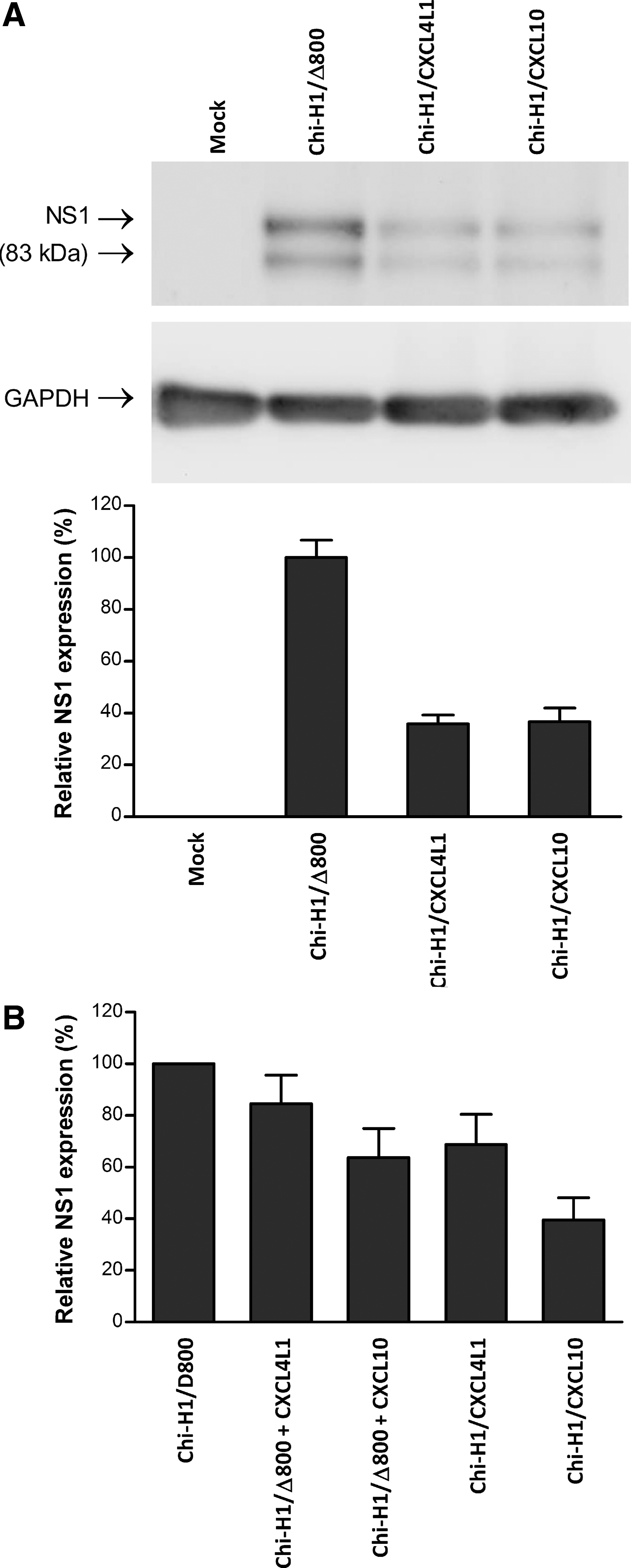
Western blot analysis of viral NS1 protein in Chi-H1/Δ800, Chi-H1/CXCL4L1, and Chi-H1/CXCL10-infected KS-IMM cells. Cell lysates from cultures were harvested 2 d.p.i. and processed for Western blotting by using primary antibody specific for NS1 and peroxidase-conjugated secondary antibodies. Immunoreactive NS1 bands were revealed through enhanced chemiluminescence. GAPDH was used as the loading control. The intensity of NS1 signals was quantified using a Chemo Cam Imager and expressed as percentage of NS1 levels after infection with the control Chi-H1/Δ800. (
Discussion
The therapeutic antitumoral potential of some viruses was recognized already in the early 1900s. 26 The use of oncolytic viruses in the clinic may not be a new concept, but major technical advances in the field of genetic engineering facilitated the development of a new generation of optimized oncolytic virotherapeutics. Recombinant oncolytic viruses armed to express factors with additional antitumoral properties represent an important step toward future multi-approach anticancer drugs attacking the tumor and tumor microenvironment on different fronts. Though virotherapy has demonstrated clinical therapeutic benefit, use of recombinant viruses could indeed more efficiently inhibit tumor progression, as suggested by multiple preclinical studies. 1,27 Previous work reported improved therapeutic efficacy with recombinant viruses engineered to instigate chemokine production. 9,12 In this study, antitumoral activity of recombinant rodent parvoviruses Chi-H1/CXCL4L1 and Chi-H1/CXCL10 in a preclinical Kaposi sarcoma model was investigated. Both chemokines have been attributed angiostatic and antitumoral activity before, mediated at least in part through their common receptor CXCR3. 25,28,29 The CXCR3 receptor distinguishes itself from most other chemokine receptors. Indeed, whereas other chemokine receptors are also subject to alternative splicing, 30,31 in some cases leading to the expression of receptors with changed ligand-binding properties, such as CCR9B, the human CXCR3B splice variant is unique. This variant loses the ability to signal through Gαi and changes toward Gαs-mediated anti-proliferative and anti-migratory signaling. 13,22 Selective expression of CXCR3B on endothelial cells accordingly contributes to the angiostatic effect of the CXCR3 ligands. It should be noted that although CXCR3 ligands had been originally described as antitumoral agents, recent studies highlighted the impact of the receptor's duality on the interpretation of its prognostic versus therapeutic value. More specifically, CXCR3 splice variant expression by the tumor cells themselves is a defining factor, balancing the pro- versus antitumoral activity of CXCR3 ligands. 32,33 Tumoral expression of CXCR3A in the presence of CXCR3 ligands indeed has been described to fuel the tumor's metastatic potential, thereby overriding potential beneficial angiostatic or immunomodulatory effects. In contrast, a shift in splice variant expression would inhibit CXCR3 ligand-stimulated proliferation and metastatic capacity. Originating from CXCR3B-expressing endothelial cells, the transformed Kaposi sarcoma cells, as expected, express exclusively CXCR3B, while CXCR3A transcripts are undetectable by qPCR. Therefore, KS-IMM cells are presumably valid targets for CXCR3 ligand therapy. Similar to the data obtained by Lavie et al., the present results indeed suggest Chi-H1/CXCL4L1 to inhibit KS-IMM migration in comparison to Chi-H1/Δ800, which in itself has angiostatic and cytotoxic properties. Though other studies have previously demonstrated added preclinical benefit when combining oncolytic herpes and vaccinia viruses with antiangiogenic strategies, 34 parvovirus H-1PV and recombinant H-1PV-derived vector Chi-H1/Δ800 exhibit endotheliotropic and antiangiogenic traits themselves, both in vitro and in vivo. This could explain the lack of outspoken added activity of the recombinant angiostatic chemokines in vivo.
The application of CXCL10-encoding viruses has been previously evaluated in other preclinical tumor models, but its therapeutic success in these studies has been associated primarily with the immunomodulatory function of CXCL10, rather than its angiostatic activity. 11 On the other hand, Giese et al. 8 reported the angiostatic effect of a recombinant parvoviral CXCL10-transducing vector, in the absence of any changes in immune cell markers. Interestingly, LYVE-1, specifically marking lymphatic endothelial cells, was also found to be expressed at significantly lower levels following either one of the included parvovirus-based treatments. The impact of Chi-H1/Δ800, Chi-H1/CXCL4L1, and Chi-H1/CXCL10 on the lymphatic vasculature within the tumor microenvironment accordingly suggests a potential virus-induced inhibition of tumor cell dissemination. However, this effect should be indirect, since mouse cells cannot be efficiently infected with rat H-1PV- and H-1PV-based vectors. One potential candidate for mediating this indirect effect could be VEGF-A, whose expression is downregulated in KS-IMM cells after infection with parvovirus. 7 Given their lymphangiostatic effect, parvoviruses might complement currently available first-line cancer therapies, which primarily cause eradication of the primary tumor. Further evaluation of the anti-metastatic potential of the recombinant viruses in appropriate preclinical metastatic tumor models is recommended in order to assess the true therapeutic value of Chi-H1/Δ800, Chi-H1/CXCL4L1, and Chi-H1/CXCL10.
Nevertheless, some concern remains regarding the lack of added antitumoral or angiostatic efficiency after introduction of the transgenes encoding CXCL4L1 and CXCL10. In vitro evaluation of the recombinant parvoviruses suggests that the transduced CXCR3 ligands might interfere with the anti-proliferative and/or cytotoxic activity of Chi-H1/Δ800 on KS-IMM. Indeed, the presence of either CXCL4L1 or CXCL10 dramatically reduces the expression of the viral cytotoxic non-structural protein NS1 in virus-infected KS-IMM cells. It is hypothesized that CXCL4L1 and CXCL10 display antiviral activity, thereby reducing rather than enhancing the therapeutic potential of the rodent parvovirus. However, the molecular mechanism responsible for the chemokines' effect on NS1 expression remains enigmatic. Accordingly, it is not yet clear if this obstacle is characteristic of the Kaposi sarcoma model. Interference of the CXCR3 ligands with viral cytotoxicity may also be specific to their interaction with the rodent parvovirus. These results indicate that CXCR3 expression might be used as a criterion to identify tumors that may not be appropriate for a parvovirus-based delivery of CXCR3 ligands.
Many promising studies have illustrated the potential therapeutic advantages of virotherapy in the treatment of cancer. Furthermore, introduction of the genetic manipulation of oncolytic viruses leads to entirely new, seemingly endless possibilities. This study, however, highlights remaining limitations. Setup of new therapeutic regimes and combination therapies requires careful consideration. Continuing study of the basic biology of all players involved should go hand-in-hand with preclinical and clinical research. Only fundamental understanding of the complex pleiotropic functions and working mechanisms of both the viral vectors at hand and the considered transgenes can truly allow innovation and lead the way for the development of promising new anticancer strategies.
Footnotes
Acknowledgments
This work was supported by the Fund for Scientific Research of Flanders (FWO-Vlaanderen project G.0764.14, G.0773.13 and G0D2517N), the Interuniversity Attraction Poles Programme initiated by the Belgian Science Policy Office (I.A.P. project P7/40), and the Concerted Research Actions of the Regional Government of Flanders (GOA13/014).
Author Disclosure
No competing financial interests exist.