
Abstract
Parkinson's disease (PD) is the second most frequent neurodegenerative disease after Alzheimer's disease, which is characterized by a low level of dopamine being expressing in the striatum and a deterioration of dopaminergic neurons (DAn) in the substantia nigra pars compacta. Generation of PD-derived DAn, including differentiation of human embryonic stem cells, human neural stem cells, human-induced pluripotent stem cells, and direct reprogramming, provides an ideal tool to model PD, creating the possibility of mimicking key essential pathological processes and charactering single-cell changes in vitro. Furthermore, thanks to the understanding of molecular neuropathogenesis of PD and new advances in stem-cell technology, it is anticipated that optimal functionally transplanted DAn with targeted correction and transgene-free insertion will be generated for use in cell transplantation. This review elucidates stem-cell technology for modeling PD and offering desired safe cell resources for cell transplantation therapy.
Introduction
T
In terms of the ability of stem-cell systems to model PD and uncover the pathogenic mechanisms that underline clearer reasons for PD neurodegeneration (gene–environment interaction), it is helpful to open new avenues for regenerative medicine. There are two limitations of cell transplantation therapy. The first is that traditional reprogrammed dopamine neurons are usually achieved by transfecting the transcription factors, which result in transgene integration, spontaneous insertional mutagenesis, 21 and resultant tumorigenesis. 22 The second is that the transplanted patient-derived dopamine neurons may carry transcriptomic or epigentic parental disease-causing memory, and the graft cells will show this tendency to the original disease phenotypes. New advances in stem-cell technology herald the dawn of solutions for these problems. On one hand, the discovery of small-molecule-driven reprogramming opens the door to producing safe, stable, and high-quality transplanted cell sources. Recent research has reported that the efficiency and feasibility of non-viral/non-integration systems had been greatly improved by using an additional combination of small molecules and neurotrophic factors working in signaling pathways in the neurogenesis of the dopaminic neuron. Furthermore, the latest developments in chemical reprogramming have successfully established xeno-free systems to generate functional dopamine neurons with cocktails of small molecules alone. On the other hand, to understand the inherent genetic defects of the neurodegenerative cells, scientists have turned to a correction at the genetic level. This goal has been realized with an increasingly convenient and strong genome editing technique. Cell sources derived from disease phenotypes have the potential to be expediently rescued by targeted correction of the identified gene mutations using today's powerful genome editing tools. With all of these developments in stem-cell technology, there is hope for the autologous transplantation of a patient's derived induced DAn (iDAn) without parental disease-causing memory and the generation of cell sources under xeno-free conditions.
Stem-Cell Technology as an Ideal Tool for Modeling PD
PD-related genetic disorders
Environmental factors have been considered the primary reasons in the pathogenesis of PD. MPTP and its hydroxylated derivatives, a particular nigrostriatal neurotoxin, has provided powerful evidence to support the theory of environmental risks, which develops a relatively selective destruction of DAn in the substantia nigra 23 and causes similar symptoms with PD in both animals and humans. 24 In the last decades, new discoveries in the neurogenetics of PD have suggested that genetic factors may play equally important roles in the nosetiology of PD. Several studies have demonstrated that major familial PD results from mutations in certain single genes 25,26 and monogenic forms, including autosomal dominant and recessive disorders, which may account for around 5–10% of all PD cases. 27 The G2019s mutation in the Leucine-Rich Repeat Kinase (LRRK2/PARK8 locus) 28,29 and mutations in the alpha-synuclein (SNCA/PARK1 locus) gene, 30 which generate intraneuronal inclusions containing α-synuclein protein known as Lewy bodies 31 and carry progressive degeneration of the neuron in the substantia nigra and then trigger clinical motor symptom, 32,33 are two of the most common autosomal dominant missense genes. Other autosomal dominant mutations, including PAPK3, PAPK13, UCH-L1, among others, are relatively rare. Autosomal recessive mutations in genes PTEN-induced putative kinase 1 (PINK1), Parkin, DJ-1 (Daisuke-Junko-1), ATP13A2, PLA2G6, FBXO7, DNAJC6, and SYNJ1 usually cause early onset and lesser clinical manifestation. For example, PD patients with a Parkin/PAPK2 mutation generally present with early-onset parkinsonism, slow disease progression, and a better response to L-DOPA. 34 In addition, the majority of PD cases are sporadic, known as idiopathic PD, possibly caused by a complex interaction among genetically susceptibility variants and environmental factors, which some geneticists and epidemiologists have explored as the G × E model in the pathogenesis of sporadic PD. 35 A large proportion of these sporadic forms have been identified in the monogenic form linked to familial PD. In other words, those genes responsible for monogenic aberrations are also susceptibility factors, 27 such as polymorphism variants in Mendelian genes, such as α-synuclein (SNCA), LRRK2, Parkin, PINK1, and DJ-1, and heterozygous mutations in susceptibility genes, such as microtubule-associated protein tau and glucocerebrosidase beta acid (GBA), identified by genome-wide association studies (GWAS). 36 The key feature of modeling PD using stem cells is the repertoire of mutations in the induced pluripotent stem cells (iPSCs) and other cell models. Generation of a patient's specific derived cell models carrying disease-causing mutations gives the opportunity of exploring the cellular impact of those mutations of which inherent mechanisms respond to environmental damage. Both familial PD and sporadic PD have been studied in this way. The primary validated genetic susceptibility factors that have been modeled in iPSCs or its derived-cells include LRRK2(G2019S), 32,37 –39 SCNA (A53T α-synuclein), 40,41 SNCA triplication, 42 and P1NK1, 43,44 PARK2, 45 and GBA mutations. 46 These are summarized in the Table 1. Furthermore, an increasing number of mutations identified by large gene detection projects, such as large-scale GWAS research, are expected to be replicated using those stem-cell models in the future.
The synopsis of well-established genetic mutations of PD
PD, Parkinson's disease; iPSC, induced pluripotent stem cells; DAn, dopaminergic neurons; ERK, extracellular-signal-regulated kinase 1/2; ER, endoplasmic reticulum; PGC-1α, peroxisome proliferator-activated receptor-γ coactivator-α (an important regulator of mitochondrial biogenesis); mitochondrial dysfunction, includes disorders of respiration and intraneuronal movement of mitochondria, and proton leakage; mROS, mitochondrial reactive oxygen species; NECAB2, neuronal calcium-binding protein 2.
Stem-cell technology reveals clear genetic factors interacting with environmental risks
To date, the underlying etiology for most cases of PD is unclear. The genetic disorders and the environmental risks leading to cell metabolism damage linked with cell pathways are taken together to account for this pathogenesis mechanism, such as oxidative stress–induced neurotoxicity via the intrinsic GTPase kinase activity pathways 47,48 and nitrosative stress–induced mitochondrial impairment via the MEF2-PGC1α pathways 40 in gene mutant–associated PD patient-derived cells. New advances in stem-cell technology—the acquirement of pure PD patient's homogeneous generation of iDAn (human embryonic stem cell [hESc]-derived DAn, 47,48 iPSc-derived DAn, 37,49,50 and directly reprogrammed DAn 51 )—provide an opportunity for the cellular events under genetically defined condition to be investigated in a human context. Compared with animal models, stem-cell models have achieved the following breakthroughs in mimicking the microenvironment of the PD cell in vitro. First, increasing evidence has been discovered to support the fact that patient-specific derived cells still retain an epigenetic memory 52,53 (e.g., the topological memory 54 ) and keep expressing the carrying transcriptomic disease-causing mutation memory. 55,56 Those PD-derived cell models show key intrinsic features with molecular alterations. Second, the isogenic cells from healthy patients as a genetically matched control were obtained from patient-specific cells via a genome-editing technique, providing a method for single-factor analysis to study observed PD phenotypes caused by a certain PD-causing mutation. Third, based on the generation of pure populations of iDAn with genetic variants, scientists could model monofactorial environmental stress–related neurodegenerative states by imposing artificial additional stress factors on pathological processes of PD, such as reactive oxygen species (ROS), 37,39 excitotoxicity, 57 and mitochondrial toxins, 58 and then analyze the aggregation and acceleration effects of environmental risks.
The molecular pathogenesis of the majority sporadic PD implied by stem cell models can be summarized as follows: environmental factors influence the progress of PD by molecular pathways in the susceptibility gene variants associated cell model. MEF2-PGC1α, among others, was discovered as a molecular protective pathway to demonstrate this. The normal activity of myocyte enhancer factor 2C (MEF2C) could excite transcription of peroxisome proliferator-activated receptor-g coactivator-1α (PGC1α; a neuroprotective factor), and environmental nitrosative/oxidative stress resulting in S-nitrosylation of MEF2C (an inhibitor) leads to mitochondrial dysfunction and increased apoptosis in a patient's specific iDAn model (A53T α-syn mutant cells). Surprisingly, this damage could be eliminated in a patient's isogenic mutation-corrected controls. 40 To sum up, stem-cell models provide possibilities to investigate the final effects of genetic background variations linked with environmental risks. The relationship between genetic susceptibility factors and PD-related environmental factors are listed in Table 2.
The relationship of PD-related genetic susceptibility factors and environmental factors
Peroxide, 6-OHDA, MG-132 induced oxidative stress injury,
Recombinant G2019S LRRK2 kinase activity.
GW5074 is the LRRK2 kinase inhibitor (protective agent).
Transcriptional dysregulation of CPNE8, CADPS2, and UHRF2.
Mutant gene: (1) LRRK2-G2019S; (2) SCNA(A53T); (3) P1NK1; (4) PAPK2; (5) GBA.
ROS, reactive oxygen species; Nrf2, Nkinase 1/2 nuclear factor erythroid 2-related factor 2; NECAB2, neuronal calcium-binding protein 2.
Environmental stress could either enhance (“↑”) the neuro-inhibitory pathway or inhibit (“↓”) the neuroprotective cell pathway in the cell model with susceptibility gene variants, resulting in corresponding neurodegenerative damage. However, that neurodegenerative damage eliminates responding to the same environmental stress after correcting the targeted gene mutation in an isogenic mutation corrected cell model. This relationship diagram reflects that environmental factors influence the progress of PD by molecular pathway in the susceptibility gene variants associated cell model.
Stem-Cell Technology Offers Desired Sources for Cell Therapy
To date, none of therapeutic methods for PD have been able to stop or slow down the degeneration of DAn in SNpc or its symptoms, including rigidity, motor function impairment (bradykinesia and freezing), and resting tremor. Currently, the drugs used to control symptoms, such as levodopa (a dopamine precursor) and MAO-B (a dopamine receptor agonists), are still the primary treatment for PD. However, the side effects of these drugs, including it wearing off, dyskinesia, and motor fluctuations, appear after long-term use. Furthermore, the aim of these drugs is not to reverse the loss of DAn in SNpc. A new approach to treat the loss of dopamine neurons is anticipated using cell-transplantation therapy. However, there is a long way to go before it is possible to generate optimal transplanted DAn with targeted correction of gene mutations but without transgene insertion, as described in Fig. 1.
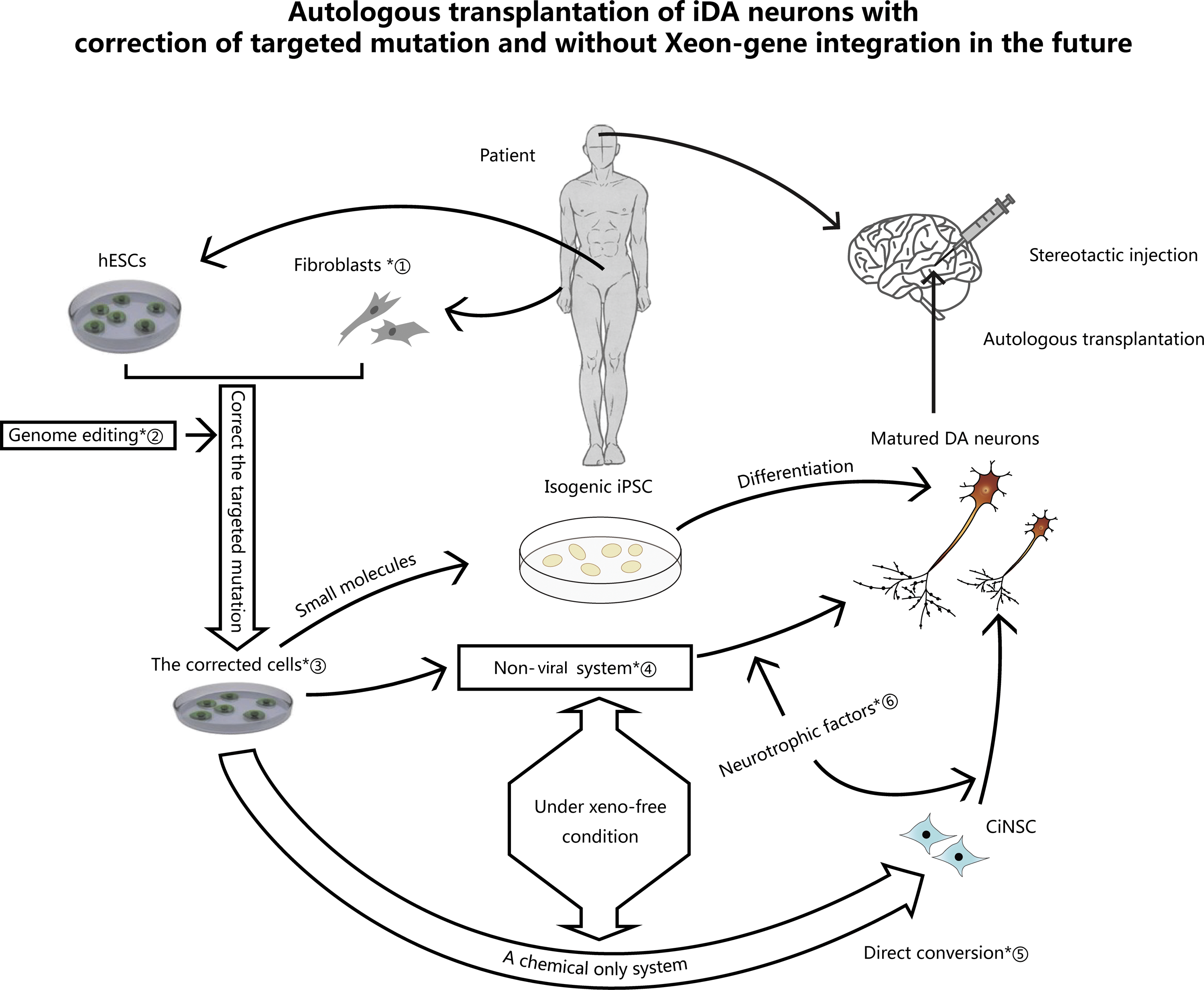
Autologous transplantation of induced dopaminergic neurons (iDAn) with correction of targeted mutation and without Xeno-gene integration. Four steps are required to achieve this. First, a supply system for autologous isogenic cells needs to be established. At present, autologous isogenic cells are mainly derived from patients' specific somatic cells via iPSCs or direct conversion. However, the transplanted cells that are derived from human embryonic pluripotent stem cells or other adult pluripotent stem cells are usually allogeneic. If the construction of an individual's specific human embryonic stem cell (hESC) line is achieved in the future, autologous cell therapy will be more easily accessible. Second, the targeted mutation corrected cells must be obtained. The realization of this goal must rely on strong genome editing techniques. Third, iDAn must be generated under xeno-free conditions. This will depend on the generation of ciDAn with use of small molecules and neurotrophic factors. Fourth step, precise stereotactic cell transplant surgery is required. ①, patient's specific fibroblasts; ②, genome editing techniques, including gDAN-NaAgo, CRISPR/Cas9, and ZFN/TALEN endonuclease-mediated genome editing; ③, the rescued cells without parental Parkinson's disease (PD)-causing mutation memory; ④, non-viral system with small molecules and neurotrophic factors in neurogenesis of DAn; ⑤, the small molecular cocktails of A-83-01, thiazovivin, purmorphamine, and VPA; ⑥, neurotrophic factors in neurogenesis of DAn, including SHH, FGF8, GDNF, NRTN, BDNF, TGF-3β, and RA.
Evolutionary secure system to optimize transplanted DAn
From the initial allogeneic cell to the later autologous cell with exogenous genes transfected, to the revised homogenous expression of exo-gene insertion, and finally to the completely isogenic cell under xeno-free conditions, there journey to generate secure sources for cell transplantation is a long one (Fig. 2). Considering the host of problems and limitations for allogeneic cell sources, including immunological rejection, ethical issues, limited cell sources, and verified complications, there are many reasons to facilitate the exploration of better autologous cell sources. Cell reprogramming was rising for disease modeling and treatment. Initially, researchers used viral vectors to deliver transcription factors, despite the efficiency and feasibility issues with this technique, which brought exogenous transgene integration and insertional mutagenesis. To solve this problem, Park et al. established a homogenous system to express exogenous genes using a drug inducer. 59 The integration/transgene-free system, such as episomal vectors (naked plasmid DNA, cationic lipids, or polymers), 60 microRNA, 61 and proteins, 62 was considered to be inefficient and unstable in the past, but the use of small molecules and neurotrophic factors greatly improved the efficacy and stability (Fig. 2, part III). Today, functional neurons can be produced via a chemical-only system (ciN), and how to realize functional integration of iDAn generated from the small molecules only system (ciDAn) in preclinical research may be the better way to reach this goal in the future (Fig. 2, part IV).

The progress of transplanted cells and expected sources. The process for the development of cell sources for transplantation and the future perspective for cell therapy is detailed. The noted time following the text box represents the starting time point for the use of the cell sources. The figure is divided into three parts: implemented clinical trials, preclinical animal experiments, and feasible cell sources in the future. The longitudinal upward time axis corresponds to a gradual process of safe transplantation as follows: I. Allogeneic cells → II. Autologous cells with exogenous genes transfected → III. Autologous cells with homogenous expression of exo-gene insertion → IV. Completely isogenic cells under xeno-free conditions. I. Initially, functional allogeneic cells were the first sources to be used in cell-replacement surgery. There were two types of allogeneic cells for PD: one extracted from the brain or other tissues, the other type derived from hESCs/hiPSCs or other pluripotent stem cells. On one type, the considered dopaminergic cells extracted from the donor's tissue were directly transplanted into the substantia nigra pars compacta of PD patients. This is the only model of cell therapy that has been carried out in clinical trials. This model was described in the lower part (Implemented clinical trials) of Fig. 2. On other type, the use of cells sources derived from hESCs has still stayed in the stage of animal experiment, the clinical trial of hESC-derived dopaminergic cells for PD is expected in the near future. This model was described in the middle part(Preclinical animal experiments) of Fig. 2. II. As shown in middle part of Fig. 2, there are two forms of cell sources in autologous cells with exogenous genes transfected: iPSC-derived dopaminergic cells and dopamine neurons derived from various somatic cells, for example fibroblasts, astrocytes, and glias. This model was described in the middle part (Preclinical animal experiments) of Fig. 2. III. To improve the viral induced exo-gene expressing system, an adult-induced system of homogenous expression of exo-gene has been proposed. This model was described in the middle part (Preclinical animal experiments) of Fig. 2. In addition, non-viral or non-integration system of reprogramming with neurotrophic factors and small molecules has been developed. The preclinical assessment is expected to be carried out as the next step. This model was described Fig. 1. IV. In the future, the goal of safe transplantation is to obtain completely isogenic cell sources via a chemical-only system. It is predicted that scientists could generate these cell sources in three ways: ciPSC, CiNSC and direct chemical conversion. This model was described in the upper part of Fig. 1.
ciDAn: a desired safe source for cell transplantation in the future
Enormous effort is being made to achieve preclinical efficacy and potency of functional ciDAn, and huge rewards are being reaped. In 2009, a chemical platform was introduced that dramatically enhanced the efficacy of iPSC generation. This platform focused on three pathways: the improvement of mesenchymal to epithelial transition (MET), such as inhibition of TGFβ pathway, the inhibition of MEK-ERK pathway, and the enhancement of cell survival. It was found that the combination of SB43125 (a TGF-β pathway AL5K inhibitor) and PD0325901 (a MEK inhibitor) worked at the three pathways, providing a basis for a non-viral, advancing, secure system for reprogramming. 63 Since then, various small molecules have been explored to replace exogenous transcription factors. Hou et al. tried various chemical agents to produce ciPSCs, include four essential small molecules: Forskolin (a PKA agonist), Chir99021 (a WNT pathway regulator), DENep (a histone methylation modulator), Repsox (a TGF-β pathway AL5K inhibitor), and other nonessential molecules. 64 SOX2 is the most common Yamanaka factor that might be replaced by small molecule compounds. Staerk et al. found that Repsox 616452, LY-364947, Dasatinib, iPYrazine (iPY), PP1 might be a replacement for SOX2. 65 In addition, Moon et al. demonstrated the targeted Sonic hedgehog signaling pathway factors Purmorphamine, Oxysterol, and Shh might work instead of reprogramming factors Sox2, Klf4, and C-Myc. 66 In 2015, Li et al. achieved a chemical lineage reprogramming for functional neurons derived from mouse fibroblasts with small chemical cocktails of ISX9 (an neurogenesis inducer), I-BET151 (a BET family protein inhibitor), CHIR99021, and Forskolin. 67 In the same year, Hu et al. directly converted human fibroblasts into cells possessing a neuronal gene expression and electrophysiological properties with only a small chemical cocktail of VCRF (V, valproic acid; C, CHIR99021; R, Repsox; F, Forskolin). 68 The latest development of chemical reprogramming reported a key finding that chemical-induced neural stem cells (ciNSC) could be directly reprogrammed from mouse fibroblasts using a combination of small molecules of A-83-01, thiazovivin, purmorphamine, and VPA, bringing a new avenue for generation of iDAn differentiated from ciNSC. 69 To date, the functional integration of ciDAn has not been reported, but capitalizing on the developments of chemical reprogramming, DAn derived from ciPSCs or ciNSCs or directly converted from somatic cells by a chemical-only system for PD therapy holds great promise for the future.
Genome editing: NaAgo—a powerful tool to correct PD-causing mutation
Despite the high cost and poor efficiency of early endonuclease-mediated genome editing including the ZFN (Zinc finger nuclease)/TALEN (transcription activator-like effector nuclease)-mediated, the RNA-guided CRISPR/Cas9 (the clustered, regularly interspaced, short palindromic repeat [CRISPR] technology) produced a revolution in the field with a highly improved efficiency and specificity. 70 Today, it has become the most commonly adopted genome editing tool, as its nucleases can easily, rapidly, and efficiently alter endogenous genes. 71 The latest DNA-guided NaAgo system constituted a major breakthrough, overcoming the restriction of tolerable temperature to applying DNA guides, and due to the more complicated structure of DNA guides, it shows a greater tolerance to mismatches, a broader targeting range, and a higher efficiency and precision than RNA-guided “CRISPR/Cas9.” 72 In particular, compared with CRISPR/Cas9, single-stranded DNA (ssDNA) in the gDNA-NaAgo system is overwhelmingly simple to be designed and synthesized for its concentration, and the NaAgo DNA guides show very low selectivity and high efficiency in targeting ssDNA without the need for protospacer-adjacent motif (PAM). 72 While the Cas9-sgRNA system used the special design PAM presenting oligonucleotides (PAMmers) to bind and catalyze DNA substrates or cut RNA targets exclusively, and the need for PAM make the CRISPR/Cas9 system enjoying a stronger stability and specificity than NaAgo. 73 In addition, the simplicity and high efficiency of CRISPR/Cas9 has been improving in recent years, 74 and in the continued quest for better specificity when performing genome editing. 75,76 It is believed that the CRISPR/Cas9 system and gDNA-NaAgo system will lead the way in the future.
Clinical perspective: safe and targeted corrected iDAn for the Precision Medicine Initiative
The Precision Medicine Initiative (PMI) that was announced by President Obama 77 proposed the concept of combining an individual patient's information, including biomarkers, genomics, and environmental factors, with electronic health records data to provide better clinical care and suitable treatment for each individual. Currently, the PMI primary supports the research of cardiovascular disease and cancers, 78,79 but its long-term goal of benefiting all areas of health is underway. 80 As elucidated in this review, gene–environment interaction is extremely important in the pathogenesis in the majority of patients with PD, and the specific pathogeny of each individual is distinct for different genetic factors with different environments. Accordingly, precision medicine shows its unique meaningfulness for clinical assessment and therapy for this type of disease. With the achievement of the feasibility of producing unlimited amounts of safe and corrected functional patient-specific iDAn, the anticipation of precise regenerative medicine for cell transplantation of each individual can be a reality.
Historical Review of Cell Transplantation
Process of cell-replacement therapy for PD
At present, there are two types of grafts resources for PD: allogeneic and autogenous. Allografts include fetal brain or other human embryonic tissues and hESC-derived DAn (Fig. 2, part I), while autologous grafts derived from patient-specific somatic cells via in vitro iPSC reprogramming or direct lineage conversion. DAns from fetal ventral mesencephalic (FVM) tissues were the major cell replacements in the implemented clinical trials. In the late 1980s, intrastriatal transplantation of FVM DAn started in a small, independent, open-label clinical trial. 81 In the following decades, a series of clinical trials were conducted with variable results. To be specific, the early studies were quite promising, 82,83 but the later large, randomized studies revealed negative results, with complications emerging gradually. 84 Ultimately, to resolve controversies, a large-scale multicenter, collaborative study—TRANSEURO—was performed, reevaluating neural transplantation. It reported the significant side effects of FVM DAn transplantation therapy that those who had levodapa-induced dyskinesia will give rise to graft-induced dyskinesias after transplantation. After TRANSEURO, the enthusiasm of transplantation trials of allogeneic dopaminergic grafts waned (Fig. 3, part I). In addition, quite a few scientists studied the transplantation of human embryonic dopaminergic grafts. They found that the transplant goes beyond just dopamine systems. 85 However, as natural sources are finite, researchers poured their attention into the PSC-derived DAn. In 2011, considerable progress was made in directed differentiation of hESCs to specialized midbrain DAn, 86 when preclinical animal experiments manifested functional integration, targeted-specific reinnervation, and motor improvement. 87 Besides hESCs, there are some pluripotent adult stem cells in the human body. Among these, in the human brain are neural stem cells, which have the ability to self-renew and can be differentiated into neurons, astrocytes, and oligodendrocytes. 88 A large number of preclinical studies have demonstrated that various types of hNSCs involving human embryonic neural stem cells, 89 fetal neural stem cells, 90 and adult neural stem cell 91 could survive, proliferate, and migrate after being transplanted into specific areas. In addition, there is definitive immunohistochemical evidence to show that the transplanted neural stem cells could differentiate into functional neurons, and integrate into structural and functional neural circuits. 89 –91
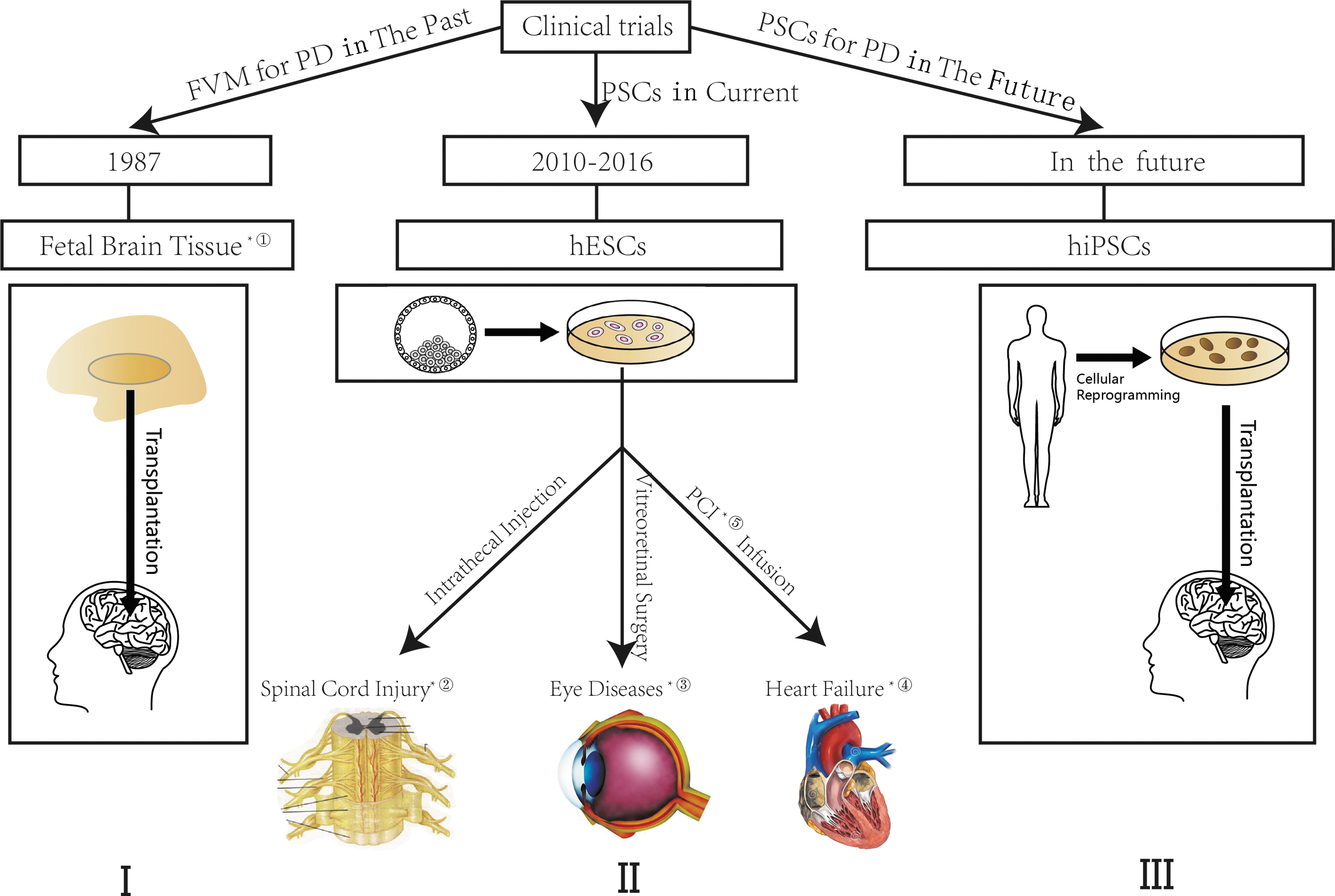
The progress of clinical trials of PSC-derived cells and its perspective for PD. ① In 1987, the first transplantation of FVM DAn started as a small, independent, open-label clinical trial. ② In 2014, the first clinical trial of hESC-derived oligodendrocyte progenitors for cell supplements to spinal cord injury has been approved. ③ Since 2010, a series of trials of hESC-derived retinal pigment epithelium (RPE) for eye diseases has been recruited, including Stargard's diseases, dry or wet age-related macular degeneration, and myopic macular degeneration. ④ In 2015, the first clinical trial of hESC-derived cardiac progenitors for severe heart failure was in recruiting. ⑤ Percutaneous coronary intervention. The figure has three parts: I. FVM for PD in the past; II. The use of PSC at present; III. The prospect of use of PSC for PD therapy in the future. I. DAn from fetal ventral mesencephalic (FVM) tissues were the major cell sources in the implemented clinical trials. This started in 1987 with a small, independent open-label clinical trial. The rise and fall of clinical trials of FVM DAn is detailed in the text. II. Human ESC-derived cell sources are the major PSC-derived cell sources in current clinical trials. Their application covers various systems: ophthalmology, cardiovascular system, nervous system, endocrine system, and others (lung, skin, etc.). The published clinical trials are described in the text, including eye disease, heart failure, spinal cord injury. III. Human iPSC-derived DAn is the most promising cell source for its isogenic nature.
Since iPSC was first described by Takahashi and Yamanaka in 2006, 92 autologous transplantation of DAn derived from a patient's specific iPSCs has attracted much attention for its easy accessibility and comparable ethicalness. A vast array of somatic cells have been successfully reprogrammed to hiPSC, for example source cells from blood cells, 93 skin fibroblasts, 94,95 and epithelial cells. 96 The potential clinical approach to test the long-term safety, efficacy, and feasibility of iPSC-derived DAn has been validated in a host of preclinical animal experiments from a rodent model 92 to a non-human primate model. 97 Compared with ESCs and iPSCs which possess teratoma formation potential with tumorigenicity, 98 the direct lineage transdifferentiation which switch one somatic type to another defined type such as DAn, skip the process of intermediates and eliminate the risk of oncogenesis when transplanted into an undifferentiated state. In 2011, Kim et al. achieved functional integration of DAn directly converted from mouse fibroblasts 99 and, in the same year, the human fibroblasts, 100 demonstrating the potential use of iDAn obtained by direct conversion. Furthermore, many terminally differentiated gliacytes spatially throughout the mesencephalon promise to be the ideal cell sources for regenerative medicine. For instance, the conversion of astrocytes and gial cells into DAn in vitro has been successfully accomplished. 101,102 Importantly, the investigation of direct lineage conversion provides the idea of in vivo reprogramming form astrocytes/gial cells as the ideal starting cells in the nervous system with natural microenvironment of midbrain (Fig. 2, part II). The progress of transplanted cells and expected sources for PD is described in Fig. 2.
Development of animal models for ex vivo therapeutics of PD
Currently, the most common model of PD replicated in the animal adult brain involves the intraperitoneal injection of MPTP and the stereotactic injection of 6-OHDA. The administration of these neurotoxins could induce specific damage of DAn to study pathophysiology, clinical features, and pathogenesis of PD. 103,104 For MPTP, its metabolic product 1-methyl-4-phenylpyridinium ion (MPP+) in vivo, which can enter DAn through the dopamine transporter (DAT) and then block mitochondrial complex I activity, deplete intracellular ATP, and enhance oxidative stress, induces the specific neurotoxicity of DAn in SNpc. 105 For 6-OHDA, which can contribute to the accumulation of ROS in the mitochondria and the inhibition of the mitochondrial respiratory enzymes, this leads to gradual neuron death and selective degeneration of DAn. 106 To model a particular state that is infinitely close to the human brain's pathological condition of PD, animal models to recapitulate transient parkinsonian-like states have been generated in various species from less developed creatures such as insecta to highly evolved species such as non-human primates. So far, the animal models used to model PD have involved the insecta of drosophila 107 ; the actinopterygii of zebrafish 108 ; the mammalian of rodents including mice 109 and rats, 110 carnivores including cats 111 and dogs, 112 artiodactyla including minipigs 113 and sheep, 114 and primates including Old Word baboons 115 and macaques, 116 and New Word squirrel monkeys 117 and marmosets. 118
Current clinical trials of PSC-derived sources and its perspective for PD
Most current clinical studies have been concentrating on eye disease because of its immunoprivileged nature. Subretinal transplantation of hESC-derived retinal pigment epithelium (RPE) in Stargardt's macular dystrophy and atrophic age-related macular degeneration (dry AMD) was the most important and the first to reach Phase I/II. The results were the first to show the safety and tolerability of hESC-derived cells in humans and possible PSC biological activity-led improvement of visual acuity. 119 –121 Human ESC-derived cells were also used to treat disorders in other systems, for example heart failure for myocardial regeneration after acute myocardial infarction, 122,123 diabetes for functional replacement of islet cells, and nervous system diseases for cell supplements to spinal cord injury. Although the first clinical trial on the use of human ESC-derived cells for spinal cord injury by Geron's did not have more data reported till now, the latest research provided evidence to support the preclinical safety for the next clinical trials in spinal cord injury (Fig. 3, part II). However, for clinical trials of iPSC, the requirement of examination is more severe because of the greater tumorigenicity risks. 124 The Labor Ministry of Japan approved the first clinical trial of iPSC-derived RPE for therapy on wet AMD in June 2013, but this was finally put on hold when a patient's specific iPSC were found to carry genetic mutations. The authors' team has also transplanted allogeneic umbilical cord blood stem cells (UCBSCs) into severe cerebral palsy (CP) patients with partial controllable adverse events. Treatment with UCBSCs has been proven to be relatively safe, 125 and further research for stem cells is ongoing. Although there has been no clinical trial for PSC-derived cells for PD, it is thought to be on the verge of coming for its beneficial effects of preclinical assessments 65,66 and the particular strength in replacing a loss of DAn. The iPSC-derived DAn is anticipated to be the most appropriate transplanted cell for PD therapy because of its ability to generate isogenic cell sources for PD patient (Fig. 3, part III).The progress of clinical trials of PSC-derived cells and its perspective for PD is described in Fig. 3.
Transcriptional control and specific differentiation of midbrain DAn
Mesencephalic progenitors differentiate to mature DAn when they pass through regional specification, Sonic Hedgehog (SHH), fibroblast growth factor 8 (FGF8), and other downstream molecules of signaling pathways, including WNT, TGFβ/Activin, and BMP signaling pathways, promote the differentiation at an early or late stage. 126,127 Each stage is regulated by a set of transcriptional regulators described in Fig. 1. Transcriptional network composed of OCT2, Mash1, Msx1, NURR1, LMX1B, PITX3, and EN1/2 is involved in the determination of the DA phenotype in postmitotic precursors. 128 Based on these theories, efforts have been made to obtain optimized protocols in both iPSC-derived DAn and ESC differentiated DAn. For iPSC differentiation, it initially transitioned into embryonic stage in a serum-containing medium basic with fibroblast growth factor (bFGF), PA6-derived stromal cell-derived inducing activity (SDIA), and retinoic acid (RA)-free. 129,130 Finally, to differentiate specialized DAn, the intermediates need to be treated with N2 medium supplemented with SHH, FGF8, cAMP, laminin, ascorbic acid (AA), 129 and cytomegalovirus plasmid (pCMV). 131 For ES cell differentiation, neurotrophic factors working in signaling pathways of neurogenesis are indispensable. SHH/FGF8 are the crucial mesencephalon growth and development factor, facilitating the maturation of mDA progenitor cells. 132 And later, the immature cells need striatum-secreting neurotrophic factors, including neurturin and cell-derived neurotrophic factor. 133 Finally, to promote the ultimate maturation of midbrain dopamine neurons, the cultures in medium should be added with the frizzled family transmembrane receptors (Wnt1 and Wnt5α) associated ligands, 134 TGF3β, BDNF, and RA. 135 Having produced the functional iDAn and finished the preclinical evaluation, the future for the application of stem cell–induced DAn looks bright.
Challenge and related issues of the therapeutics of stem cells
This review has summarized the advantages and potentials of stem cells for clinical use. However, many questions and obstacles exist in the domain of the therapeutics of stem cells. Nowadays, the primary challenges to limit the clinical use of stem cells involve ethical questions, tumorigenesis, immune response, and toxicity to a degree. There are huge ethical issues and intense immune response in the use of ESCs. Compared with ESCs, iPSCs stand out as having fewer ethical issues and reduced immune rejection, but because of powerful pluripotency, the risk of tumor development of iPSCs may be greater than it is with other stem cells. Furthermore, iPSCs derived from autologous PD patients may carry pathogenic gene mutations that affect the prognosis for cell-replacement therapy. To obtain a higher rate of survival and integration, the details of canonical grafting procedures need to be estimated through more studies. These should include (1) the cell number and timing of transplantation, (2) the intended location and injection site of transplanted cells, (3) the status of proliferation and differentiation capacity of cells when transplant surgery is performed, and (4) the type of surgery. In short, stem-cell therapy is currently at the initial stage, and great effort is needed to move it forward.
Footnotes
Acknowledgments
This work was supported by National Natural Science Foundation of China (Grant no. 31271595 to Tongxiang Lin), Key Project of Chinese National Programs for Fundamental Research and Development (973 Programs; 2013CB9669002 to T. Lin). This work was also supported by Science and Technology Department of Guangdong Province China (2015A020212032 to T. Lin), Guangdong Provincial Hospital of Chinese Medicine (YK2013B2N08 to T. Lin), Medical Research Fund of Guangdong Province (B2014257), Science Popularization Project of Haizhu District in Guangzhou (KP2013(TJ)-24), and Scientific Research Initiation Project of Southern Medical University (PY2014N071).
Author Disclosure
This review manuscript is not submitted or being considered for publication elsewhere. All authors have seen and approved the manuscript and have contributed significantly for the paper. No financial conflict inappropriately influences or biases the content of this manuscript.