
Abstract
The liver is a major off-target organ in gene therapy approaches for cardiac and musculoskeletal disorders. Intravenous administration of most of the naturally occurring adeno-associated virus (AAV) strains invariably results in vector genome sequestration within the liver. In the current study, we compared the muscle tropism and transduction efficiency of a liver de-targeted AAV variant to AAV9 following systemic administration in newborn rhesus monkeys. In vivo bioluminescence imaging was performed to monitor transgene expression (firefly luciferase) post administration. Results indicated comparable and sustained levels of systemic firefly luciferase gene expression in skeletal muscle over a period of two years. Quantitation of vector biodistribution in harvested tissues post-administration revealed widespread recovery of vector genomes delivered by AAV9 but markedly decreased levels in major systemic organs from the AAV variant. These studies validate the translational potential and safety of liver de-targeted AAV strains for gene therapy of muscle-related diseases.
Introduction
R
From a cardiovascular perspective, therapeutic gene transfer for induction of angiogenesis to revascularize ischemic tissue, correction of familial hypercholesterolemia, arrhythmias, cardiomyopathies, and gene therapy for heart failure have all been evaluated. 9 Notably, success in the treatment of heart failure has been reported in a phase 2 study (CUPID trial) targeting the sarco(endo)plasmic reticulum Ca(2+) ATPase (SERCA2a), 10 although later studies showed no difference in primary endpoints. The trial established safety following intracoronary delivery of AAV1/SERCA2a vectors in advanced heart failure patients. Other routes of AAV vector administration for achieving global cardiac expression, such as direct transepicardial or transendocardial injection after left thoracotomy as well as venous infusion, have been assessed in preclinical settings. 11 –13 It is noteworthy to mention that the CUPID trial validated the use of a catheter-based AAV vector delivery system, which allowed access to the coronary artery through a femoral approach. In parallel, preclinical studies have demonstrated that the closely related strain AAV6 has demonstrated superior cardiac transduction in several large animal models including dogs, sheep, pigs, and nonhuman primates. 11,12,14,15 Further, although AAV9 vectors have been shown to be less efficient than AAV6 or AAV8 in canine models and adult rhesus monkeys, preclinical studies have demonstrated effective cardiac gene transfer following intracoronary administration in porcine models of heart failure. 16
One unaddressed clinical challenge pertaining to the application of natural AAV isolates for systemic gene expression in muscle is the high transduction efficiency in off-target organs. Although off-target gene expression can be mitigated to a certain extent using tissue-specific promoters and/or microRNA–based strategies, 17 –19 it should be noted that undesirably high levels of vector genome copies are still recovered from multiple tissues following systemic injection. 17 –19 Thus, from a safety perspective, the continued development of optimal AAV vectors for achieving efficient and global muscle gene expression with minimal off-target effects is essential. In the current study, we compared the ability of a novel laboratory-derived AAV2 based variant, AAV2i8, 20 with AAV9 to promote robust, selective, and long-term transgene expression in cardiac and musculoskeletal tissues following a single intravenous (IV) administration in newborn rhesus monkeys.
Materials and Methods
Vector production
Recombinant AAV2i8 previously described 20 and AAV9 vectors packaging the chicken beta actin promoter driving a firefly luciferase cassette were produced at the Vector Core at the University of North Carolina at Chapel Hill (UNC Vector Core) using previously described methods. 21 Vector genome titers were determined by quantitative PCR according to established methods.
Rhesus monkeys
All animal procedures conformed to the requirements of the Animal Welfare Act and protocols were approved prior to implementation by the Institutional Animal Care and Use Committee at the University of California, Davis. Normally cycling, adult female rhesus monkeys (Macaca mulatta) (n = 6) with a history of prior pregnancy were bred and identified as pregnant according to established methods. 22 Activities related to animal care (diet, housing) were performed per California National Primate Research Center standard operating procedures. Newborns were delivered by cesarean section at term (160 ± 2 days of gestation) according to standardized protocols 23 . Cord blood samples (12–15 mL) were collected at birth (complete blood counts [CBCs], clinical chemistry panels), and animals were raised in the nursery for postnatal studies.
Vector administration and sample collection
A total volume of 2 mL of the AAV vector supernatant (3 × 1012 genome copies/kg; AAV2/9, n = 2 one male and one female; AAV2i8, n = 4 three males and one female) was injected IV via a peripheral vessel (upper limb) on the day of birth. Postinjection, animals were nursery reared up through 3 months postnatal age and then housed in a juvenile housing environment. Animals were paired with another study animal at the appropriate age for the duration of the study. Infant health, food intake, and body weights were recorded daily in the nursery and then on a routine basis thereafter. Blood samples (3–6 mL, dependent on age) were collected monthly from a peripheral vessel to monitor CBCs and clinical chemistry panels.
Bioluminescence imaging
Bioluminescence imaging (BLI) for firefly luciferase expression was performed beginning at 2 weeks then at 1 and 2 months postnatal age, then approximately every 3–6 months thereafter, and immediately after an IV injection of D-luciferin (100 mg/kg), using a IVIS imaging system with Living Image software (Perkin Elmer, Waltham, MA) according to established protocols. 24 Ventral, dorsal, and right and left lateral images of each animal were acquired at each imaging session in a light-tight chamber, and whole body images were obtained with quantification. Bioluminescent and photographic images were superimposed, using Living Image 3.0 software. Regions of interest were defined by selecting areas showing bioluminescence. Total photons per second per square centimeter (p/s) detected in regions of interest were recorded and compared across animals and time points.
Tissue harvest
Animals were euthanized with an overdose of pentobarbital after an IV injection of telazol. Tissue harvests were performed according to established protocols at approximately 2 years of age. 23 All organs were removed and imaged for firefly luciferase expression immediately after tissue harvest including the brain (cerebral hemispheres, cerebellum), lung lobes (all lobes), all major muscle groups (e.g., upper and lower limbs, thoracic, intercostals, abdominal), trachea, esophagus, heart, pericardium, thymus, spleen, liver (all lobes), axillary/inguinal/mesenteric lymph nodes, pancreas, right and left adrenals and kidneys, reproductive organs including right and left gonads, gastrointestinal tract (stomach, duodenum, jejunum, ileum, colon), muscular component of the diaphragm, omentum, and peritoneum (muscle wall). Bone marrow was also collected and cryopreserved. All tissues were collected for quantitative PCR and were placed in 1.5 mL microcentrifuge tubes and immediately frozen in liquid nitrogen and stored at ≤−80°C until assay. In addition, tissue sections from all of the above were placed in Optimal Cutting Temperature (OCT) embedding compound and then quick frozen over liquid nitrogen or fixed in 10% buffered formalin and embedded in paraffin. Representative 5–6 μm sections were obtained from all tissues and stained with hematoxylin and eosin for routine histopathology. 25
Vector genome biodistribution
Genomic DNA was isolated from all tissues using the Gentra Puregene Tissue kit (Qiagen, Valencia, CA) as recommended by the manufacturer. 26 The primers for AAV were as follows: forward 5′-sequence-3′, reverse 5′-sequence-3′, and probe 5′-6FAM-sequence-TAMSp-3′. The ɛ-globin system was utilized as an internal control for DNA isolation and PCR reactions. The primer sequences for ɛ-globin were as follows: forward 5′-TGGCAA GGAGTTCACCCCT-3′; reverse 5′-AATGGCGACA GCAGACACC-3′; and probe 5′-6FAM-TGCAGGCT GCCTGGCAGAAGC-TAMSp-3′. Real-time quantitative PCR analysis was carried out in 96-well optical plates using the 7900 ABI Sequence Detection System (Applied Biosystems, Foster City, CA) and the TaqMan Universal PCR Master Mix (Applied Biosystems) according to the manufacturer's protocols. PCR reactions contained 1 × TaqMan universal master mix with 400 nM of forward and reverse primers and 100 nM probe in a 25-μL reaction volume. The PCR protocol consisted of one cycle of 2 min at 50°C, 15 min at 95°C, followed by 40 cycles at 15 s at 95°C, and 60 s at 60°C.
Results
Outcome
All animals (n = 6) displayed normal growth trajectories when compared to historical and concurrent controls (data not shown). All CBCs and chemistry panels were within normal limits with no abnormalities detected (not shown). Animals remained robust and healthy throughout the study period.
BLI reveals systemic and long-term gene expression
Based on previous mouse studies characterizing chimeric AAV2 capsid (AAV2i8), 20 studies demonstrated a dramatic de-targeting of liver transduction and a shift in biodistribution of vector transduction primarily to skeletal muscle and cardiac tissue illustrating a sharp contrast in vector tropism when compared with AAV2-based vectors which are almost exclusively liver tropic (Fig. 1a). 20 To ensure these original observations were not species specific, we demonstrated AAV2i8 transduction in the rhesus monkey model when compared to the well-characterized AAV9 vector known for systemic transduction in all species tested. Rhesus monkeys were injected IV with AAV9 or previously described chimeric AAV2-based capsid AAV2i820 vectors packaging a firefly luciferase reporter transgene driven by the chicken beta actin promoter. Transgene expression levels were comparable (within 2-fold) at early time intervals (up to 3 months) post-administration (Fig. 1b). At later time intervals (6 months to 1 year), AAV2i8 vectors transduced the thoracic and abdominal regions with higher efficiency compared to AAV9. Transgene expression levels assessed by BLI persisted for the duration of analysis. Semiquantitation of bioluminescence at different time intervals revealed that AAV9 mediated rapid onset of transgene expression compared to AAV2i8 (Fig. 2). In addition, the BLI signal output persisted at high levels in all animals. Robust and selective muscle gene expression was observed across the time points and in several harvested tissues. Specifically, firefly luciferase activity was observed in the intercostal muscles (1.98 × 1011 p/s), quadriceps (1.57 × 1011 p/s), abdominal muscles (8.33 × 1010 p/s), muscular component of the diaphragm (1.28 × 1011 p/s), and heart (2.06 × 1010 p/s), but was barely detectable over background levels in the lung or liver lobes with AAV2i8. Taken together, these results suggest that AAV2i8 and AAV9 can mediate robust, systemic, and persistent transgene expression following IV administration in rhesus monkeys at birth.
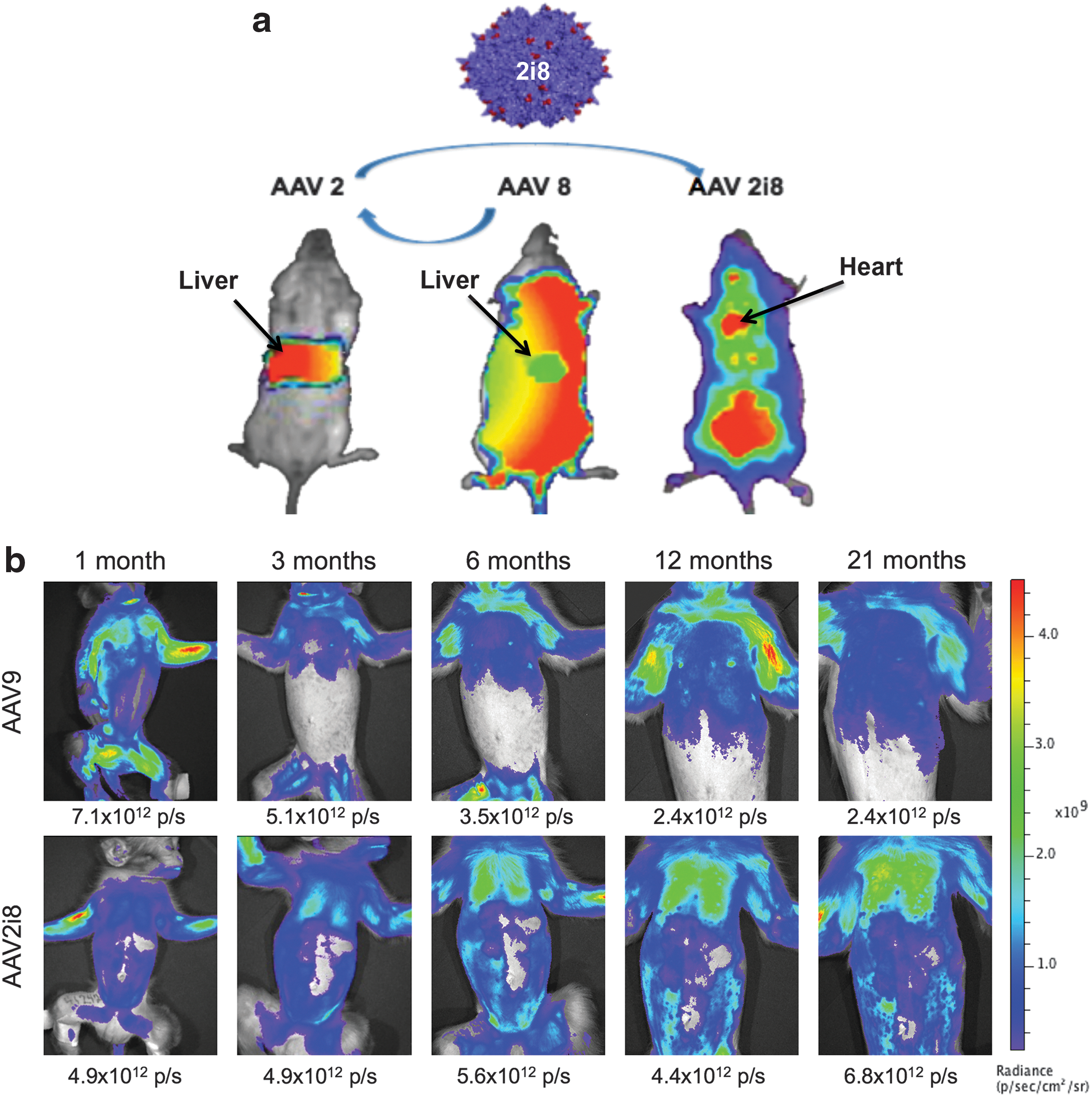

Bioluminescence (photons/sec) was quantified for individual animals at all time points. Stable levels of bioluminescence were observed through 21 months as noted. AAV9 showed high, transient expression of firefly luciferase up to 2 months postnatal age. Mean levels of bioluminescence for animals in each group (n = 2 for AAV9, n = 4 for AAV2i8) are shown (standard error bars for AAV2i8).
Biodistribution
Animals injected with AAV vectors at birth were euthanized at approximately 2 years postnatal age and tissues harvested for quantitation of transgene copies. Overall, animals administered AAV9 showed higher levels of transduction in more tissues compared to AAV2i8 (Fig. 3). For example, AAV9 showed a higher level of transgene copies in the cerebral hemispheres (4.8 × 101 copies of firefly luciferase per 5 × 104 copies of ɛ-globin) compared to AAV2i8 (1.4 copies of firefly luciferase per 5.0 × 104 copies of ɛ-globin). AAV9 also showed higher levels in the lung (2.1 × 103 copies of firefly luciferase per 5 × 104 copies of ɛ-globin), heart (8.3 × 102 copies of firefly luciferase per 5 × 104 copies of ɛ-globin), and muscle (3.8 × 103 copies of firefly luciferase per 5 × 104 copies of ɛ-globin) compared to AAV2i8 (lung: 1.4 × 102 copies of firefly luciferase per 5 × 104 copies of ɛ-globin; heart: 1.8 × 101 copies of firefly luciferase per 5 × 104 copies of ɛ-globin; muscle: 3.1 × 101 copies of firefly luciferase per 5 × 104 copies of ɛ-globin). No copies to minimal copies of the transgene were observed in the gonads with either AAV vector. AAV9 showed transduction in the liver (2.8 × 101 copies of firefly luciferase per 5 × 104 copies of ɛ-globin), but a very low level of transduction was observed with AAV2i8 (2.6 copies of firefly luciferase per 5 × 104 copies of ɛ-globin). Taken together, these results suggest that AAV2i8 appears to display a distinct biodistribution profile in nonhuman primates when compared to the natural isolate, AAV9. This observation is consistent with comparison of the parental AAV2 capsid backbone to AAV2i8 when analyzed in mice (Fig. 1a).
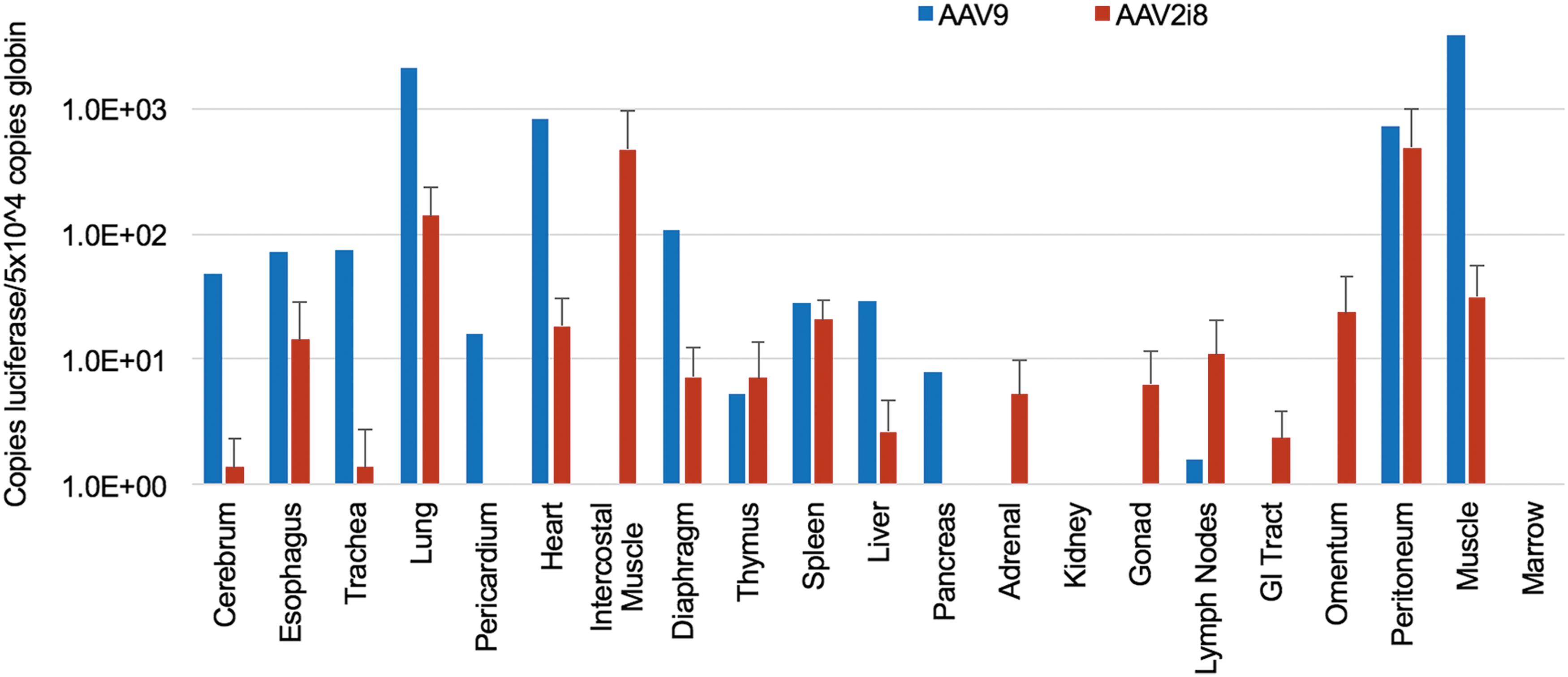
Quantitation of vector genome biodistribution with AAV9 or AAV2i8 at 2 years postnatal age. Copies of the firefly luciferase transgene per 5 × 104 copies of globin are shown. The gastrointestinal (GI) tract includes the stomach, duodenum, jejunum, ileum, and colon; lymph nodes include axillary, inguinal, and mesenteric. Mean levels of bioluminescence for animals in each group (n = 2 for AAV9, n = 4 for AAV2i8) (standard error bars for AAV2i8).
Discussion
Tissue tropisms displayed by different AAV serotypes are mediated in part by capsid interactions with host glycans. The earliest human isolate, AAV2, which has been evaluated in several clinical trials, utilizes heparan sulfate for cell surface attachment and displays a liver-tropic transduction profile (Fig. 1a). In earlier studies, several groups demonstrated that mutation of a basic cluster of amino acid residues in the major capsid protein, VP3 of AAV2, abrogated heparin binding and reduced transduction in the liver. Subsequently, we reported the design of a novel panel of liver de-targeted AAV strains derived from AAV2 by replacing key residues with corresponding residues from different natural AAV isolates. 20 These novel AAV2i mutants not only displayed decreased heparin binding and liver transduction, but also showed systemic muscle tropism. One particular engineered strain, AAV2i8, which contains six amino acids from AAV8 in the altered capsid surface loop, has shown the potential for translation in the context of therapeutic muscle gene transfer applications.
Concurrent with these studies, another natural human isolate, AAV9, has shown promise for achieving robust and systemic gene expression following a single IV bolus dose. This distinct AAV strain utilizes galactose as the host glycan receptor and displays widespread transduction in several animal models including nonhuman primates. 27 –30 Therefore, in the current study, we compared the ability of AAV2i8 and AAV9 to mediate selective and efficient gene expression in muscle following IV administration in newborn rhesus monkeys. Previous studies have demonstrated that the single amino acid insertion can enhance the muscle transduction efficiency of AAV2 and related strains following direct intramuscular injection. In the current study, we observed that a single systemic dose in newborn rhesus monkeys resulted in rapid onset and persistent transgene expression in multiple tissues over a period of two years. Despite comparable transgene expression levels, we observed several notable differences in tissue biodistribution. First, we were unable to recover significant viral genome copies from major off-target organs such as the liver and lung with AAV2i8 vectors. These results are consistent with earlier studies in rodent and porcine models validating this laboratory-derived AAV strain as a highly efficient liver de-targeted vector. 20 However, it should be noted that AAV2i8 vector genomes were recovered from several other organs including the thymus, spleen, and prostate at levels similar to AAV9. Consistent with the muscle-specific firefly luciferase expression profile in the study reported here as well as earlier preclinical studies, AAV2i8 vector genome copies were recovered from several skeletal muscle subtypes. Paradoxically, despite efficient cardiac gene expression, we were unable to recover AAV2i8 vector genome copies from cardiac tissue subtypes. One potential explanation for this finding is that post-entry processing of AAV2i8 is highly efficient in cardiac and muscle tissues, resulting in robust gene expression whereas typical transduction requires 1×103 AAV genomes to obtain 1 infectious particle that can complete the infectious pathway. Recent studies in our laboratory demonstrated numerous small molecules that will enhance AAV transduction in animal models up to 100-fold, 31 strongly suggesting that improvement of current AAV vectors by capsid modification or small molecule formulation is still warranted.
Our results demonstrate that when considering AAV vectors for efficient widespread muscle delivery, the laboratory-derived AAV2i8 strain is an excellent alternative candidate vector, which when administered through the IV route in newborns, can mediate rapid onset and selective and sustained levels of transgene expression in a range of muscle groups with a liver de-target profile. With numerous AAV clinical trials using various AAV vectors (e.g., 2, 8, or 9) all eliciting an immune response in human liver, this strongly suggests that it would be advantageous to utilize natural isolates to de-target this tissue. To date, we have not identified any natural isolates that will de-target liver, but have synthetically engineered capsid vectors that reduce liver transduction significantly. It is noteworthy to mention that we have also engineered liver de-targeted vectors derived from AAV9 for systemic gene transfer applications focused on cardiovascular, pulmonary, musculoskeletal, and the central nervous system suggesting that this approach or “rational” design of AAV capsid with preferred tropism is amenable to all serotypes and provides researchers with a vector tool box that may address some of the rate limiting clinical observations currently being encountered in numerous phase 1 studies. A similar clinical paradigm can be envisioned in the treatment of neuromuscular disorders such as Duchenne muscular dystrophy or neurologic diseases wherein the route of vector administration is likely to be intrathecal, intracisternal, or intracranial.
In summary, the continued development and evaluation of novel AAV strains in conjunction with gene regulation strategies that will allow spatiotemporal control of transgene expression is likely to improve vector safety and efficacy in human clinical trials.
Footnotes
Acknowledgments
These studies were supported by the National Heart, Lung, and Blood Institute (NHLBI) Center for Fetal Monkey Gene Transfer for Heart, Lung, and Blood Diseases No. HL085794 (A.F.T.), and National Institutes of Health grant numbers HL112761 (R.J.S.) and HL089221 (A.A.), and the Primate Center base operating grant (No. OD011107).
Author Disclosure
No competing financial interests exist.