
Abstract
MicroRNAs (miRNAs) are important post-transcriptional regulators that control cancer development and progression. However, the application of miRNA therapy in cancer has been hampered by a lack of an efficient and targeted delivery system. In our previous studies, an oriP promoter–based minicircle system successfully mediated targeted foreign gene expression in EBNA1-positive nasopharyngeal carcinoma (NPC). However, it remains to be evaluated whether this system can be applied for tumor miRNA therapy. miR-31-5p, a tumor suppressive miRNA involved in the tumorigenesis of EBV-positive NPC, was selected as the therapeutic miRNA to be transferred. In this work, we constructed a novel EBNA1-specific miRNA expression system, minicircle-oriP-miR-31. The results indicated that mc-oriP-miR-31 mediated selective miR-31-5p expression in EBNA1-positive NPC cells. Both the proliferation and migration of EBNA1-positive NPC cell lines were inhibited by mc-oriP-miR-31 treatment in vitro. Furthermore, mc-oriP-miR-31 treatment inhibited xenograft growth and lung metastasis in vivo. We also identified WDR5 as a novel miR-31-5p target. Knockdown of WDR5 inhibited NPC cell proliferation and migration and was associated with downregulation of Notch1. Reintroduction of WDR5 partially abrogated the suppressive effects induced by miR-31-5p.
In conclusion, we demonstrate for the first time that targeted expression of miR-31-5p using a nonviral minicircle vector can serve as a novel approach for tumor miRNA therapy. Moreover, WDR5 may be a promising therapeutic target for NPC treatment.
Introduction
M
Minicircles are minimized plasmids devoid of bacterial backbone. 11,12 They carry only the expression cassette of a therapeutic gene. 11,12 Minicircles are superior to conventional plasmids in the following aspects: transfer efficiency, biosafety, and bioavailability. 11,12 Although this novel nonviral system could mediate highly efficient expression of protein-coding genes in various cell types, 11 –21 there is only one report thus far on the feasibility of a minicircle-based miRNA expression system. 19 Hu et al. constructed a minicircle system expressing the miRNA-210 precursor and evaluated its biological effects in a murine model of myocardial infarction. 19 The results of that study indicated that minicircle-mediated miRNA-210 overexpression could improve angiogenesis and cardiac function. 19 However, it remains unknown whether a minicircle-based miRNA expression system could be applied in tumor treatment.
We demonstrated for the first time the feasibility of minicircle vectors in antitumor gene therapy. 16 Our studies revealed that the minicircle is capable of mediating persistently high expression of a therapeutic gene in human nasopharyngeal carcinoma (NPC). 16 Nearly 100% of anaplastic or poorly differentiated NPC express Epstein-Barr virus (EBV) proteins. 22 Accordingly, we designed an Epstein-Barr nuclear antigen-1 (EBNA1)–targeted minicircle system, minicircle-oriP (mc-oriP), in which foreign gene expression is driven by the oriP promoter. 18,23 The binding of the EBV protein EBNA1 to the family of repeats (FR) sequence in the oriP promoter drives expression of the therapeutic gene. 18,23 Our results showed that minicircle-oriP was capable of mediating highly efficient and targeted foreign gene expression and that it holds great potential for antitumor gene therapy for NPC. 18
The study of Cheung et al. proved that miR-31-5p functions as an important tumor suppressive miRNA in NPC. 24 Both homozygous deletion and hypermethylation of the promoter contributed to the downregulation of this miRNA. 24 This finding makes miR-31-5p an attractive target for NPC miRNA therapy. However, it should be noted that miR-31-5p functions in a context-dependent manner. 24 –29 Indeed, it could serve as a tumor suppressive miRNA or an oncogenic miRNA in different cell types. 25 Considering the complex functional roles of miR-31-5p, the optimal approach is to specifically express therapeutic miR-31-5p in tumors to avoid unexpected side effects of systemic expression. The specific downregulation of miR-31-5p in EBV-positive NPC offers the possibility of selectively expressing miR-31-5p using the EBNA1-induced oriP promoter.
Cheung et al. demonstrated that miR-31-5p decreased NPC cell growth by directly regulating FIH1 and MCM2. 24 Because miRNAs can exert their functions through multiple targets, whether there were other targets involved in the biological effects of miR-31-5p in NPC, especially on migration, remains to be investigated. Therefore, we predicted miR-31-5p targets using TargetScan, miRanda, and PicTar algorithms. The prediction results suggested the presence of a putative binding site of miR-31-5p in the WD repeat domain 5 (WDR5) 3′ UTR. WDR5 is a core member of the H3K4 mixed lineage leukemia (MLL) methyltransferase complexes. 30 Until now, the direct miRNA regulation of WDR5 has not been reported. The potential roles of WDR5 in NPC also remain unclear.
In this work, we demonstrate that minicircle-oriP-miR-31 can serve as a novel targeted miRNA therapy approach for nasopharyngeal carcinoma. WDR5 is a novel miR-31-5p target, the upregulation of which in response to miR-31-5p reduction promotes the proliferation and migration of NPC. Our results thus provide new perspectives for developing an effective miRNA therapy for nasopharyngeal carcinoma.
Materials and Methods
Cell lines and patient tissue specimens
C666-1, an undifferentiated EBV-positive NPC cell line, was a generous gift from Dr. Saiwah Tsao (University of Hong Kong, Hong Kong, P.R. China). 18 The poorly differentiated EBV-negative NPC cell lines 5-8F and 6-10B were maintained in our laboratory. HEK293T (human embryonic kidney cell line), HepG2 (human hepatocellular carcinoma cell line) and MCF-7 (human mammary adenocarcinoma cell line) cells were purchased from ATCC. Cells were grown in RPMI 1640 or DMEM medium (Invitrogen) supplemented with 10% fetal bovine serum (FBS) (Gibco), penicillin (100 units/mL) and streptomycin (100 μg/mL) at 37°C in a 5% CO2 atmosphere. The cell lines were routinely confirmed to be mycoplasma-free using a PCR-based detection kit.
Ten primary NPC tissues and six normal nasopharynx biopsies were collected from Cancer Center, Sun Yat-sen University (Guangzhou, P.R. China) with the approval of ethical review committees. Informed consent was obtained from patients.
Construction of recombinant parent p2ΦC31 plasmids expressing miR-31-5p
Our laboratory constructed pSP72-oriP-polyA, a pSP72-based vector carrying the oriP-CMV promoter and poly A sequence.
18
The oriP-CMV promoter sequence in this vector was subcloned from
The engineered pre-miRNA double-stranded oligo fragment (mature has-miR-31-5p sequence flanked on either side with sequences from murine miR-155, Supplementary Fig. S1; Supplementary Data are available online at
Plasmid pSP72-CMV-miR-31 was obtained by substituting the oriP-CMV sequence of pSP72-oriP-miR-31 with the CMV promoter. The parent plasmid p2ΦC31-CMV-miR-31 was then constructed by inserting the SalI-CMV-miR-31-polyA-ApaI fragment into the XhoI-Apa I sites of p2ΦC31, and pSP72-CMV-miR-NC and p2ΦC31-CMV-miR-NC were constructed as negative controls. The plasmids were confirmed by sequencing.
Production of minicircles expressing miR-31-5p
Minicircle-oriP-miR-31 and minicircle-CMV-miR-31 were produced as described. 12,16,18
Cell transfection
Transfections were conducted using Lipofectamine 2000 (Life Technologies). MicroRNA inhibitor and hsa-miR-31-5p and negative control inhibitor (abbreviated as NC I) were obtained from GenePharma (Shanghai, China). Small interfering RNA (siRNA) for Notch1 was obtained from Santa Cruz Biotechnology. Small interfering RNAs for EBNA1 were synthesized as described. 31 Small interfering (si) RNAs of WDR5 (sense: 5′- GCUGGGAAUAUCCGAUGUATT; antisense: 5′-UACAUCGGAUAUUCCCAGCTT-3′) and WDR5-2 (sense: 5′-GAAUGAGAAAUACUGCAUATT-3′; antisense: 5′-UAUGCAGUAUUUCUCAUUCTT-3′) were synthesized by Invitrogen. Plasmid pcDNA-WDR5, a vector encoding the full-length coding sequence of WDR5 (without the 3′ UTR), was constructed by our laboratory. For transient transfection, cells were plated in 24-well plates. Twenty-four hours later, the cells were transfected with 50 nM of oligonucleotides or 0.5 μg of DNA vector (plasmid/minicircle). For plasmid and minicircle co-transfection, 0.25 μg plasmid plus 0.25 μg minicircle vector were used.
Construction of stable cell lines
Lentivirus particles for the stable overexpression of hsa-miR-31-5p and negative control (abbreviated as LV-NC) were purchased from GenePharma Genolution. Cell lines stably expressing miR-31-5p were selected with 2 μg/mL puromycin for 2 weeks. pLNCX2/EBNA1 was a generous gift from Prof. Musheng Zeng (Cancer Center, Sun Yat-sen University, Guangzhou, P.R. China). Cell lines stably expressing EBNA1 (5-8F/EBNA1 vs. 5-8F/CTRL) were constructed by selection with 0.5 mg/mL G418 for 14 days.
RNA purification and qRT-PCR analysis
Total RNA was isolated from tissues and cultured cells using Trizol (Eastwin, Beijing, China). Reverse transcription was performed using Moloney Murine Leukemia Virus Reverse Transcriptase (Promega). Primers were designed for WDR5 (forward, 5′-GCCTACACCTGTGAAGCCAAAC-3′; reverse, 5′-GAATCTGACGACCAGGCTACAT-3′) and JAG2 (forward, 5′-GACGGCTTTGAGTGCATCTG-3′; reverse, 5′-GCATGGCTTCCCTTCACACT-3′). The primers for Notch1, Notch3, JAG1, and DLL4 were synthesized according to the sequences, as described by Man et al. 32 β-actin was used as an internal control for mRNA. 16 Quantitative RT-PCR analysis was conducted with SYBR green master mix (Invitrogen). A Bulge-Loop™ miRNA qRT-PCR Primer Set was purchased from Ribobio to quantify hsa-miR-31-5p expression. U6 was employed as the internal control for miRNA.
WST analysis
Cell viability was measured by WST analysis as described. 16,18
Antibodies
The primary antibodies (Santa Cruz, CA) used included anti-WDR5, anti-EBNA1, anti-Notch1, and anti-β-actin. β-actin was employed as a loading control.
In vitro migration assay
Cell migratory capability was determined using 8-μm-pore transwell chambers (BD Biosciences). A 5-8F cell suspension (2.5 × 104) in 1640 medium without serum was pipetted into the upper insert. RPMI 1640 medium containing 10% FBS was added in the bottom chamber. After 16 hours, cells on the lower surface of the insert were stained with crystal violet. For the C666-1 cell line, migration assay was conducted as described by Shairet al. 33 A cell suspension (1 × 105) in 1% FBS 1640 medium was added to the upper insert of a 24-well chamber precoated with 20 μg/mL fibronectin (BD Biosciences). Fibronectin (50 μg/mL) was used as a chemoattractant. Cells on the lower surface of the insert were stained 24 hours later. All of the experiments were conducted in triplicate. Under a microscope, five independent fields were counted.
Animal model of NPC tumor growth and lung metastasis
Animal experiments were conducted according to established guidelines, with approval of the Institutional Animal Care and Usage Committee (Sun Yat-sen University Cancer Center). Four- to five-week-old male BALB/c nude mice were obtained from Slaccas Experimental Animals Co. Ltd.. To determine the effects of minicircle-mediated miR-31-5p expression on the growth of NPC xenografts, mice were inoculated with 1 × 106 5-8F/EBNA1 cells subcutaneously into the right flanks. When tumors of approximately 100 mm3 had formed, the mice were randomly assigned (n = 5; 4 groups). Tumor tissues were injected with minicircles (15 μg minicircle vector +60 μL Lipofectamine 2000 per week for 2 weeks). The length and width of tumors were measured at the indicated time points. At the end point, the primary tumors were removed and weighed.
To evaluate the effects of minicircle-mediated miR-31-5p overexpression on the experimental lung metastases of NPC xenografts, 5-8F/EBNA1 cell line was transfected with mc-CMV-miR-31 or mc-oriP-miR-31, respectively; mc-CMV-NC or mc-oriP-NC was used for the control group. Mice (n = 7) were inoculated in the tail vein with 1 × 106 minicircle-treated 5-8F/EBNA1 cells, 24 hours after transfection. Metastasis was determined by measuring the lung weights and counting the number of lung metastatic nodules 18 days after injection.
To produce subcutaneous tumors or experimental lung metastases with 5-8F cells stably overexpressing miR-31-5p or nonspecific miRNA, 1 × 106 5-8F/LV-NC or 5-8F/LV-miR31 cells were injected into the right flanks (n = 5) or tail veins (n = 7) of nude mice. The animals were euthanized 16 or 18 days after inoculation, and their primary tumors or lungs were removed and weighed.
Luciferase reporter assay
A WDR5 mRNA 3′ UTR fragment (NM_017588.2, region 1210-2989) was subcloned into the pGL3-control vector (Xba I site, Promega), resulting in a reporter vector named pGL3-WDR5 WT. A mutant variant of the WDR5 mRNA 3′ UTR was placed into pGL3-WDR5 MUT. HEK293T cells were cotransfected with pGL3-WDR5-WT (240 ng) or pGL3-WDR5-MUT (240 ng), and pGL4.73 vector (10 ng; Promega) using Lipofectamine 2000. Where indicated, mc-CMV-miR-31 (250 ng) or miR-31-5p inhibitor (50 nM) was cotransfected. Forty-eight hours after transfection, cells were harvested. Dual-Luciferase Reporter Assay System (Promega) was used to detect luciferase activities. For each experiment, wells were transfected in triplicate, and each well was assayed in triplicate.
Statistical analysis
Results are expressed as the means ± standard errors from three independent experiments. Data were assessed using Student's two-tailed t-tests. Spearman's correlation analysis was employed to assess the correlation of miR-31-5p with WDR5 expression. Statistical analyses were carried out using SPSS 17.0 software (SPSS, Inc.). p Values less than 0.05 were considered statistically significant.
Results
mc-oriP-miR-31 mediates selective expression of miR-31-5p in EBNA1-positive cells
To mediate the selective expression of miR-31-5p in EBNA1-positive cells, an engineered pre-miR-31 expression cassette under the control of the oriP promoter was constructed (Fig. 1A; Supplementary Fig. S1). An engineered pre-miR-31 expression cassette driven by the CMV promoter was used as a control group (Fig. 1A; Supplementary Fig. S1).
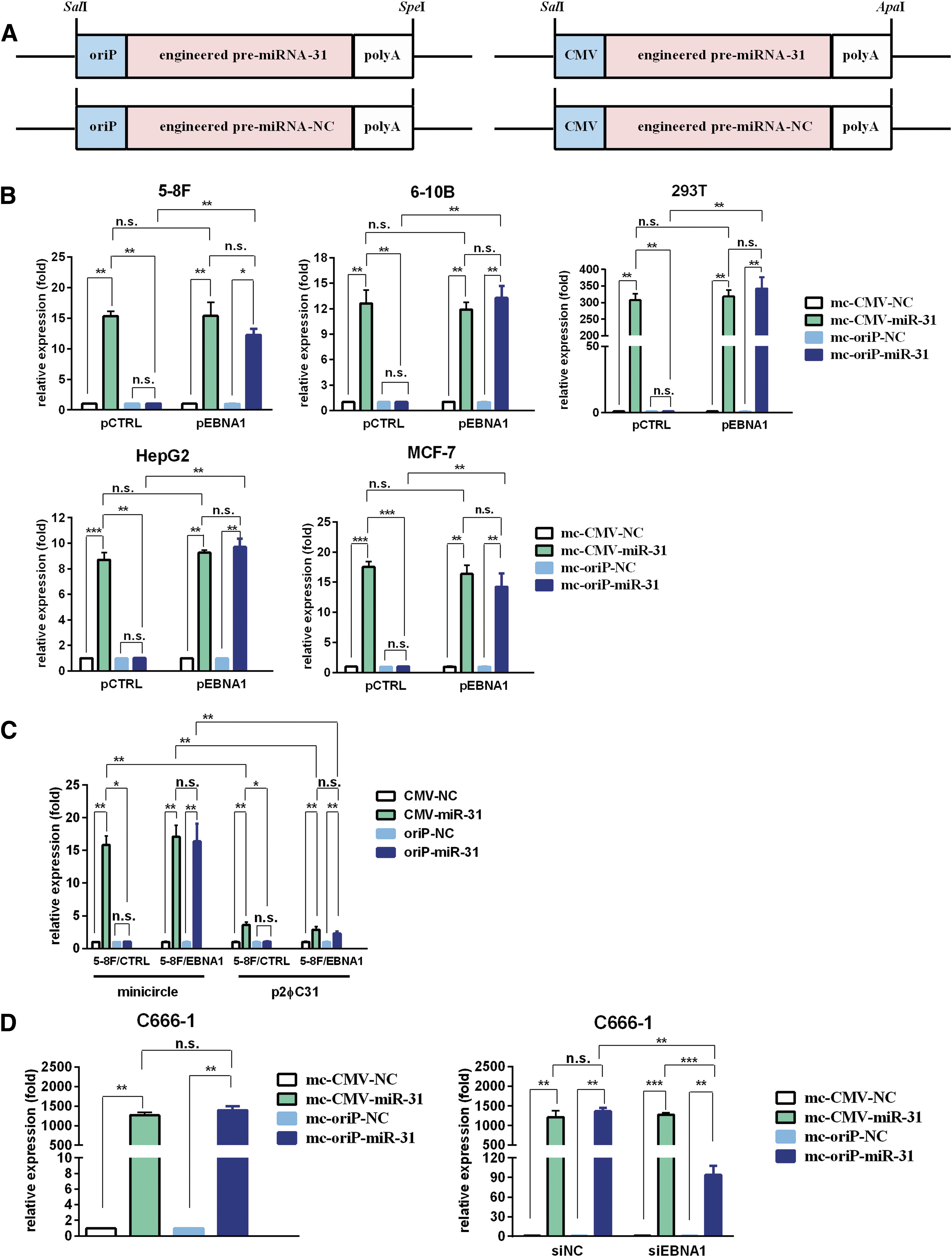
mc-oriP-miR-31 mediates selective miR-31-5p expression in EBNA1-positive cells.
To determine miRNA expression in the mc-oriP-miR-31 or mc-CMV-miR-31 group in the presence or absence of EBNA1, a panel of EBV-negative cell lines (5-8F, 6-10B, 293T, HepG2, and MCF-7) was co-transfected with minicircle vector and an EBNA1 expression plasmid. Ectopic expression of EBNA1 was confirmed by qRT-PCR (Supplementary Fig. S2A). As shown in Fig. 1B, ectopic EBNA1 expression significantly increased the miR-31-5p levels in the mc-oriP-miR-31 group (p < 0.05; Fig. 1B). No changes in miR-31-5p expression in the mc-CMV-miR-31 treatment group were observed (p > 0.05; Fig. 1B). In the presence of EBNA1, miR-31-5p levels in the mc-oriP-miR-31 group were comparable to those in the mc-CMV-miR-31 transfected group (p > 0.05; Fig. 1B), which suggested that oriP promoter could efficiently mediate selective foreign miRNA expression in EBNA1-positive cell lines.
To confirm further the efficiency and selective expression activity of mc-oriP-miR-31 in EBNA1-positive NPC cells, 5-8F cell lines stably expressing EBNA1 (5-8F/EBNA1 vs. 5-8F/CTRL) were constructed. The stable overexpression of EBNA1 in the cell lines was validated (Supplementary Figs. S2B, S3A). No significant changes in the basal level of miR-31-5p, cell viability or migration capacity was observed between the stable cell lines and corresponding parent cell lines (Supplementary Fig. S3). The expression efficiency of minicircle-miR-31 was compared with that of p2φC31-miR-31, the parent plasmid. All of these plasmids contain an engineered pre-miR-31 expression cassette with the oriP or CMV promoter (Fig. 1A). The results suggested that the minicircle groups attained significantly higher expression of miR-31-5p than the parent plasmid p2φC31 groups (p < 0.05; Fig. 1C). The miR-31 expression levels were similar when controlled by the CMV or oriP promoter in 5-8F/EBNA1 cell lines (p > 0.05; Fig. 1C), which suggested that EBNA1-induced oriP promoter was capable of mediating as efficiently the expression of foreign miRNA as that of the CMV promoter.
For the C666-1 NPC cell line (EBNA1-positive), mc-oriP-miR-31 transfection resulted in high foreign miRNA expression that was similar to that of mc-CMV-miR-31 (p > 0.05; Fig. 1D). When C666-1 was co-transfected with siEBNA1 and the minicircle vector, decreased miR-31-5p expression in the mc-oriP-miR-31 group was observed (p < 0.05; Fig. 1D, Supplementary Fig. S2C). No significant changes in miR-31-5p levels in the mc-CMV-miR-31 transfected group were observed (p > 0.05; Fig. 1D), consistent with the selective expression activity of the oriP promoter.
Taken together, these results showed that the minicircle vector could efficiently mediate the ectopic expression of miR-31-5p in various types of cell lines. In particular, we achieved selective miR-31-5p expression in EBNA1-positive cells using oriP promoter.
mc-oriP-miR-31 treatment inhibits the proliferation and migration of EBNA1-positive nasopharyngeal carcinoma cells
In order to evaluate the effects of mc-oriP-miR-31 on proliferation in EBNA1-positive NPC cells, 5-8F cells stably expressing EBNA1 (5-8F/EBNA1 vs. 5-8F/CTRL) were transfected with mc-oriP-miR-31 or mc-CMV-miR-31, respectively. As shown in Fig. 2A, the viability of both 5-8F/EBNA1 and 5-8F/CTRL was significantly inhibited by mc-CMV-miR-31 treatment (p < 0.05), while mc-oriP-miR-31 treatment showed selective antiproliferation activity in 5-8F/EBNA1 cells. No effects on cell viability were observed for the mc-oriP-miR-31 treatment group in EBNA1-negative 5-8F/CTRL cells (p > 0.05; Fig. 2A). To confirm further the antiproliferation effects of mc-oriP-miR-31 in EBNA1-positive NPC cell lines, C666-1 was transfected with mc-oriP-miR-31 or mc-CMV-miR-31. The results indicated that both mc-oriP-miR-31 and mc-CMV-miR-31 treatment significantly reduced the viability of C666-1 (p < 0.05; Fig. 2B). No difference between the inhibition rates of these two groups was observed (p > 0.05; Fig. 2B).
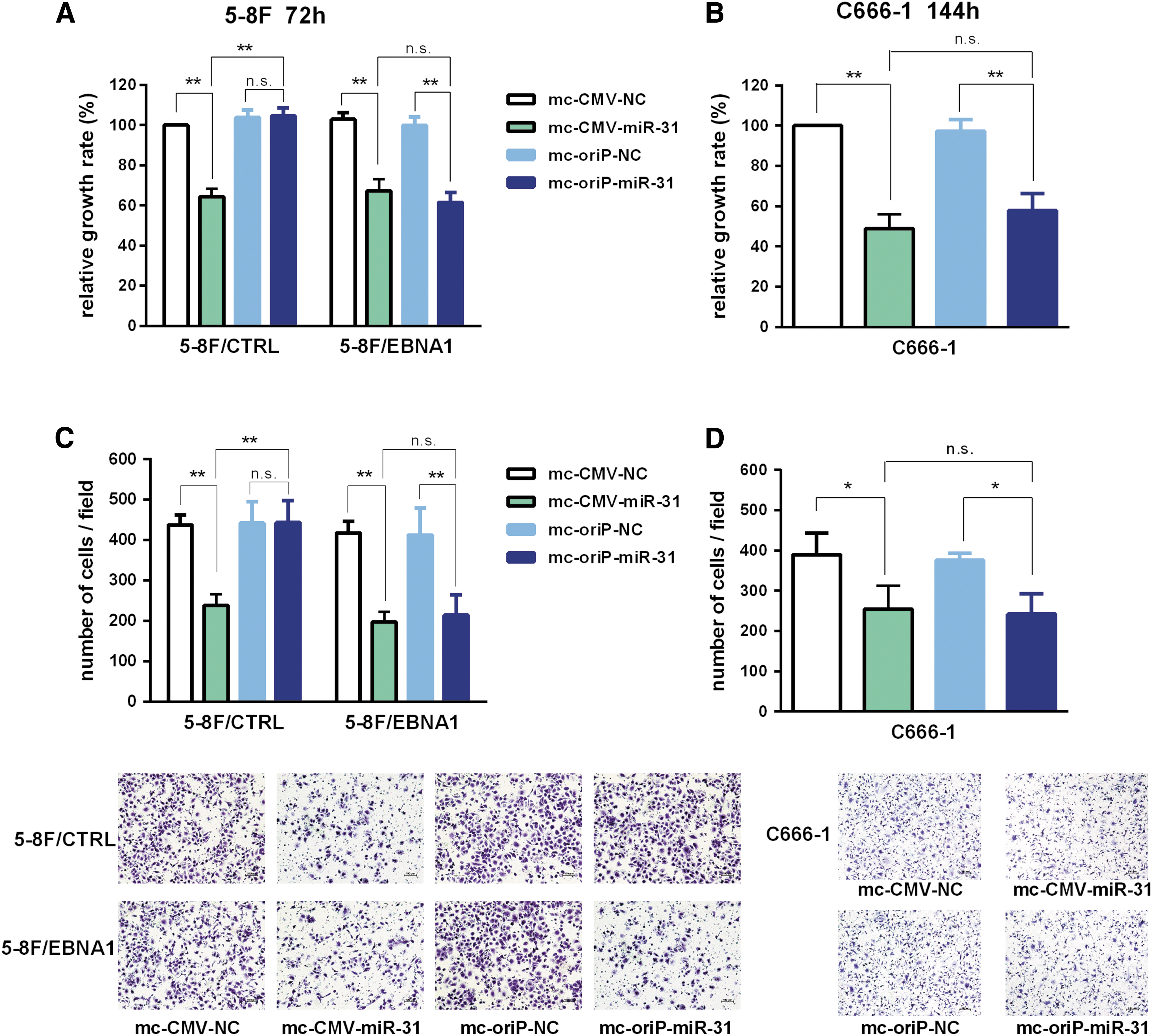
mc-oriP-miR-31 inhibits proliferation and migration of EBNA1-positive nasopharyngeal carcinoma cells in vitro.
To explore the contributions of mc-oriP-miR-31 on the migration of EBNA1-positive NPC cells, 5-8F/EBNA1 was transfected with mc-oriP-miR-31 or mc-CMV-miR-31. For the mc-CMV-miR-31 treatment group, which showed foreign miRNA overexpression despite the EBNA1 status, antimigration activity was observed in both 5-8F/EBNA1 and 5-8F/CTRL cells (p < 0.05; Fig. 2C). The mc-oriP-miR-31 vector specifically inhibited the migration of 5-8F/EBNA1 cells (p < 0.05; Fig. 2C). For the C666-1 cell line, mc-oriP-miR-31 and mc-CMV-miR-31 treatment resulted in similar migration inhibition effects (Fig. 2D).
Effects of mc-oriP-miR-31 on nasopharyngeal xenograft growth and lung metastasis in vivo
In order to evaluate further the activity of miR-31-5p on 5-8F cell lines in vivo, we constructed 5-8F stable cell lines overexpressing miR-31-5p or nonspecific miRNA, termed 5-8F/LV-miR31 or 5-8F/LV-NC (Fig. 3A). Cells were inoculated subcutaneously. Compared with those of 5-8F/LV-NC cells, the volume and weight of the tumors in the 5-8F/LV-miR31 group were significantly decreased (p < 0.05; Fig. 3A).
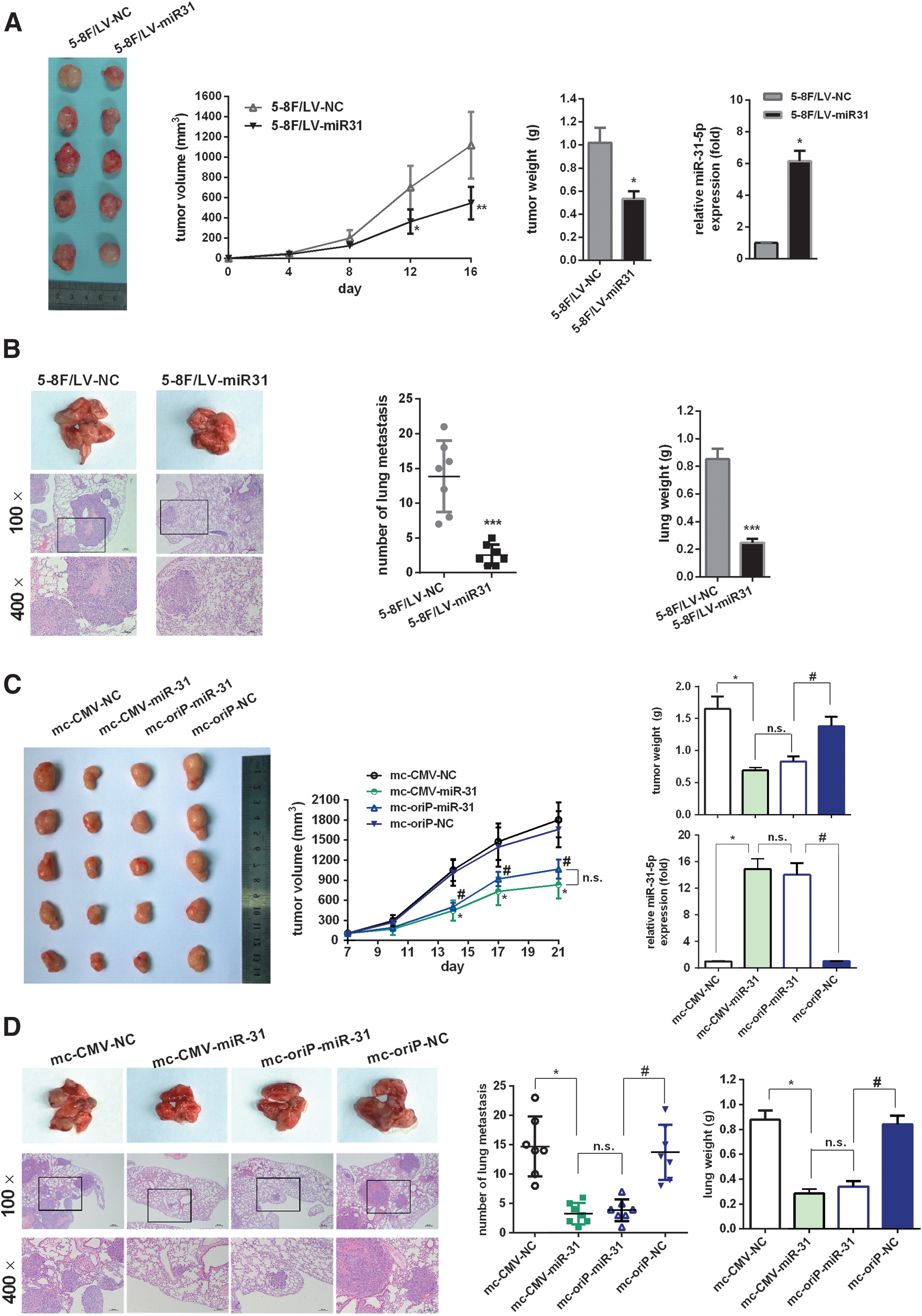
mc-oriP-miR-31 inhibits EBNA1-positive nasopharyngeal carcinoma cell growth and migration in vivo.
To explore the inhibition effects of miR-31-5p on lung metastasis, 5-8F/LV-miR31 or 5-8F/LV-NC cells were injected into the lateral tail vein of mice. Micrometastasis to the lungs in the 5-8F/LV-miR31 group was reduced and of a smaller size than that observed in the 5-8F/LV-NC group (p < 0.05; Fig. 3B). The lung weights in the 5-8F/LV-miR31 group were also less than that of the 5-8F/LV-NC group (p < 0.05; Fig. 3B). Taken together, these results showed that miR-31-5p suppressed 5-8F cell xenograft growth and lung metastasis in vivo.
To evaluate the contribution of minicircle-mediated miR-31 overexpression in vivo, 5-8F/EBNA1 cells were inoculated subcutaneously. When tumors of approximately 100 mm3 had formed, mice were treated with mc-CMV-miR-31 for 2 weeks. The results suggested that the weights and sizes of tumors in the mc-CMV-miR-31 group were significantly decreased compared with the mc-CMV-NC group (p < 0.05; Fig. 3C). Moreover, treatment of EBNA1-positive NPC xenografts with mc-oriP-miR-31 achieved comparable inhibition effects to those in the mc-CMV-miR-31 group (p > 0.05; Fig. 3C). For the mc-CMV-NC and mc-oriP-NC negative control groups, there were no significant differences in tumor weight or volume (p > 0.05; Fig. 3C).
Furthermore, 5-8F/EBNA1 cells pretreated with minicircle vectors were injected into the tail veins of mice to investigate the effects of minicircle-mediated miR-31 overexpression on metastasis. The micrometastatic loci of lungs in the mc-CMV-miR-31 group were far fewer and smaller than in the mc-CMV-NC group (p < 0.05; Fig. 3D). The lung weights of the mc-CMV-miR-31 group were also less than those in the mc-CMV-NC group (p < 0.05; Fig. 3D). Moreover, mc-oriP-miR-31 showed antimetastatic effects similar to those observed in the mc-CMV-miR-31 group (p > 0.05; Fig. 3D). For the mc-CMV-NC and mc-oriP-NC negative control groups, there were no significant differences in lung weight or number of metastatic loci (p > 0.05; Fig. 3D).
In conclusion, minicircle-mediated miR-31-5p therapy inhibited NPC growth and metastasis in vivo. In the presence of EBNA1, the targeted mc-oriP-miR-31 vector could achieve comparable therapeutic effects as nontargeted mc-CMV-miR-31.
WDR5 is a direct target of miR-31-5p
We predicted the potential targets of miR-31-5p using the TargetScan, miRanda, and PicTar algorithms. The results indicated that WDR5 contains a putative miR-31-5p-binding site in its 3′ UTR. To characterize the impacts of miR-31-5p overexpression on WDR5, the 293T, 5-8F and C666-1 cell lines were transfected with mc-CMV-miR-31 or mc-CMV-NC. Ectopic expression of miR-31-5p suppressed WDR5 mRNA and protein levels compared with cells transfected with the scramble miRNA vector (p < 0.05; Fig. 4A). In 293T and 5-8F cells, we also found a significant increase in WDR5 mRNA and protein levels after transfection with the miR-31-5p inhibitor (p < 0.05; Fig. 4A). However, no differences in WDR5 mRNA or protein levels were detected in C666-1 cells transfected with the miR-31-5p inhibitor, which could be attributed to the extremely low endogenous level of miR-31-5p (p > 0.05; Fig. 4A).
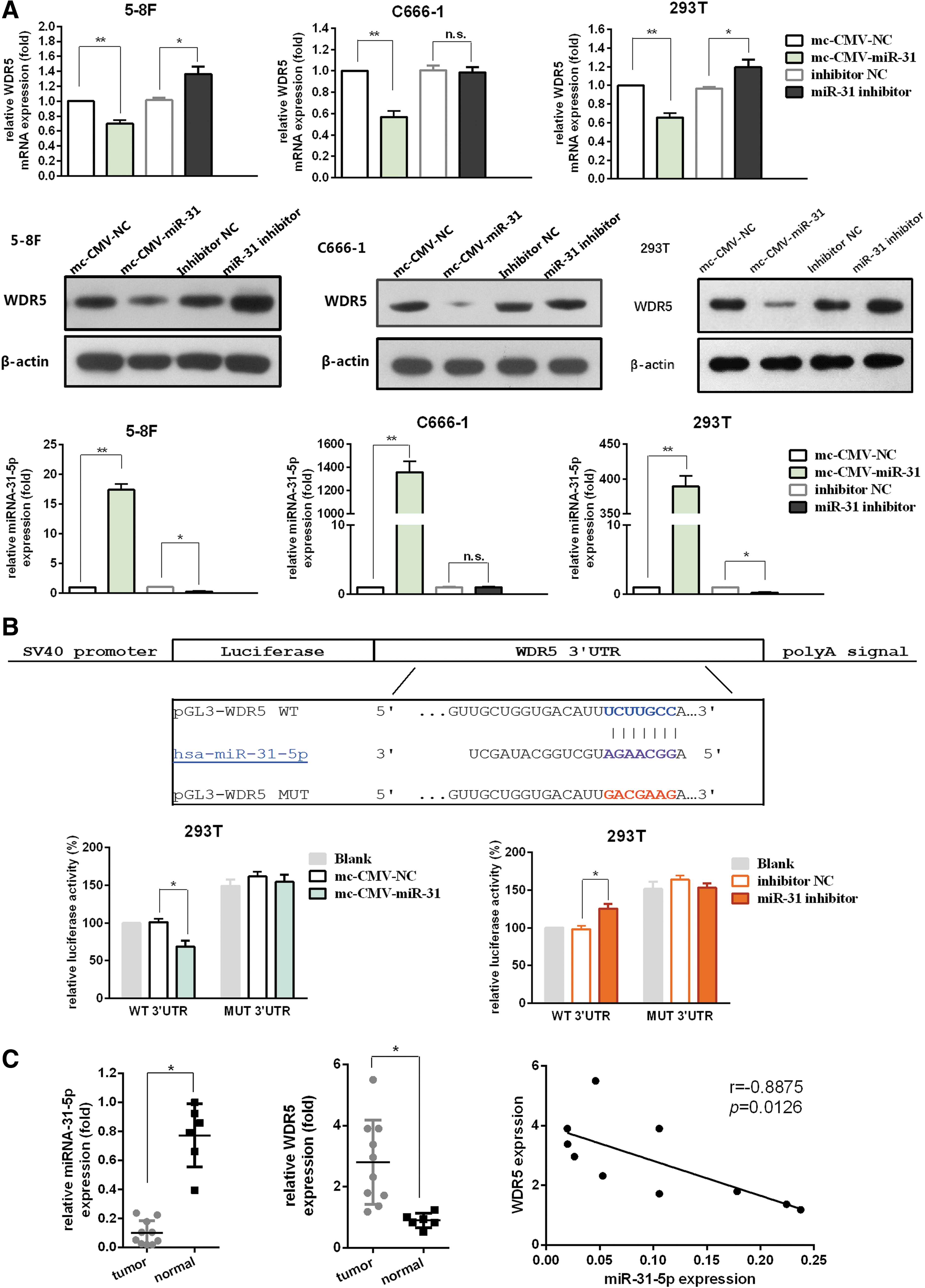
miR-31-5p directly targets WDR5.
To evaluate whether miR-31-5p regulates WDR5 through direct binding to its 3′ UTR, luciferase reporter vectors containing the wild-type or mutant WDR5 3′ UTR were constructed (Fig. 4B). The wild-type WDR5 3′ UTR reporter (pGL3-WDR5-WT) showed markedly reduced luciferase activity in 293T cells transfected with mc-CMV-miR-31, compared with that of the mc-CMV-NC group (p < 0.05; Fig. 4B). In contrast, no differences in luciferase activity were observed in cells co-transfected with pGL3-WDR5-MUT and mc-CMV-miR-31 compared with the mc-CMV-NC group (p > 0.05; Fig. 4B). Moreover, co-transfection with pGL3-WDR5-WT and the miR-31-5p inhibitor resulted in increased luciferase activity (p < 0.05; Fig. 4B), whereas the activity of pGL3-WDR5-MUT was unaffected after transfection with the miR-31-5p inhibitor (p > 0.05; Fig. 4B). These findings suggested that WDR5 expression was suppressed by miR-31-5p through direct targeting of the 3′ UTR.
We used qRT-PCR to examine the relative expression of miR-31-5p or WDR5 in 10 NPC tissues and 6 normal tissues. MiR-31-5p expression was significantly downregulated in NPC specimens (p < 0.05; Fig. 4C), but WDR5 expression was significantly upregulated (p < 0.05; Fig. 4C). Furthermore, an inverse correlation between the levels of miR-31-5p and WDR5 expression was observed in NPC specimens (p < 0.05; Fig. 4C).
Downregulation of WDR5 by miR-31-5p inhibits NPC cell proliferation and migration
In order to determine whether WDR5 functioned as a downstream target of miR-31-5p, WST assays, and migration assays for the 5-8F and C666-1 cell lines were conducted. First, two siRNA sequences of WDR5 were tested (data not shown). The sequence attaining stronger inhibition of WDR5 expression (Fig. 5A, F) was called siWDR5 and was chosen for use in subsequent activity experiments. The results suggested that knockdown of WDR5 could significantly inhibit cell viability (p < 0.05; Fig. 5B) and migratory ability (p < 0.05; Fig. 5C). Next, we tested whether ectopic expression of WDR5 could reverse the effects of miR-31-5p. The results indicated that overexpression of WDR5 could significantly promote cell viability and migratory ability (p < 0.05; Fig. 5D, E). Concomitant overexpression of miR-31-5p and WDR5 nullified the inhibitory effects of miR-31-5p on the viability and migration capacity of NPC cells (Fig. 5D, E). In conclusion, these results indicated that miR-31-5p could inhibit NPC cell viability and migration, partially by targeting WDR5.
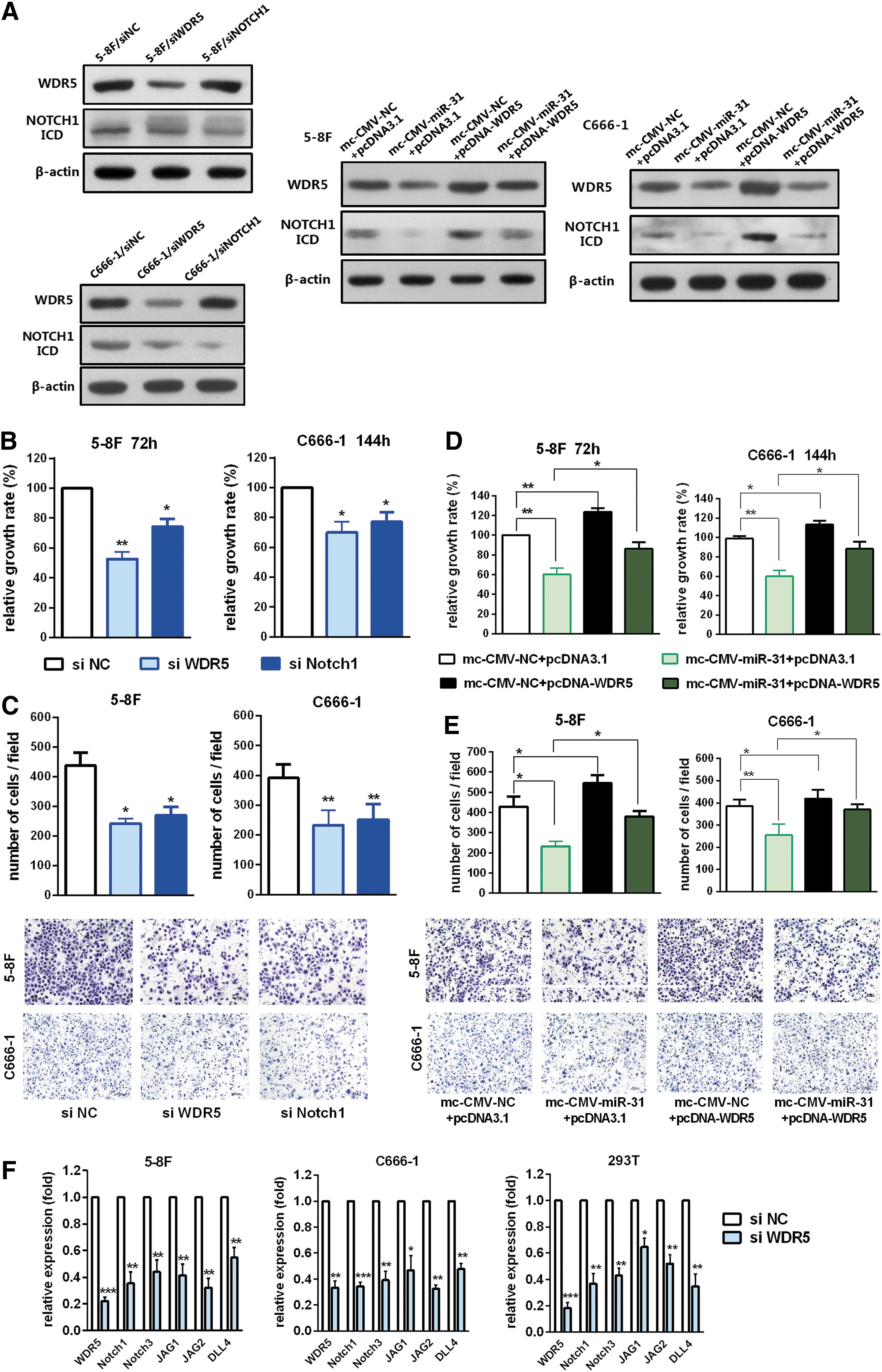
miR-31-5p inhibits NPC cell growth and migration through WDR5.
Chung et al. 34 reported that WDR5 and CBX8 acted cooperatively to regulate normal mammary stem cells and breast tumorigenesis through the regulation of Notch-network gene promoters. Notch signaling plays oncogenic roles in NPC. 32,35,36 To clarify whether WDR5 also regulates Notch-network genes at transcriptional level in the context of NPC, qRT-PCR analysis of Notch pathway genes was conducted. For 5-8F and C666-1 cells, WDR5 knockdown resulted in a decrease in the mRNA levels of multiple Notch pathway genes, including Notch1, Notch3, JAG1, JAG2 and DLL4 (Fig. 5F). Similar regulation was observed in 293T cells (Fig. 5F). Notch1 was selected as a representative molecule to evaluate the potential functional link between miR31-5p/WDR5 and the Notch pathway. The results showed that overexpression of miR31-5p downregulated Notch1 expression (Fig. 5A). In contrast, the ectopic expression of WDR5 partially reversed this regulation (Fig. 5A). Moreover, Notch1 knockdown significantly impaired the cell viability and migration capacity of NPC cells (p < 0.05; Fig. 5B, C), suggesting that loss of Notch1 recapitulated the effects of WDR5 depletion.
Discussion
The activities of miRNA are cell-type dependent. 2 Accordingly, the same miRNA can regulate different downstream genes in different cells, resulting in diverse biological effects. 2 Therefore, the delivery of therapeutic miRNAs must be targeted to specific cell types and tissues. To our knowledge, the current study is the first report on a minicircle-based tumor-specific miRNA therapy approach.
It remains to be evaluated whether minicircle vectors carrying other types of tissue-specific promoters could be used in tumor miRNA therapy. Taking hepatocellular carcinoma as an example, miR-31-5p functions as a tumor suppressive miRNA by regulating cell cycle proteins and epithelial–mesenchymal transition regulatory factors. 29 Thus, an hepatocellular carcinoma–specific promoter could be used in mediating the selective expression of miR-31-5p in this type of tumor. In addition to the applications in mediating miRNA overexpression, the minicircle-based vector may have potential use in mediating expression of miRNA inhibitors or long noncoding RNA (lncRNA).
Our results indicated that minicircle served as an efficient gene therapy vector and achieved comparable foreign miRNA expression to lentivirus vector (Fig. 3). In contrast with lentivirus vector system, minicircles could mediate sustained foreign gene expression without risk of integration into the target cell genome. Therefore, minicircle may provide a safe and efficient approach for tumor miRNA therapy.
Our study identified a novel miR-31-5p target: WDR5. It is a core subunit of the human MLL and seT1 (hCOMPASS) histone H3 Lys4 methyltransferase complexes. 30 Recently, growing recognition of the pro-tumorigenic properties of WDR5 and its value as anticancer drug target has provided an impetus for discovering small molecules that bind key interaction surfaces on WDR5, such as the MYC/RBBP5/KANSL2 binding cleft. 37 –44
In our study, the potential roles of WDR5 in NPC cell viability and migration capacity were investigated. The results indicated that downregulation of WDR5 decreased the viability and migration of NPC cells (Fig. 5). Therefore, WDR5 inhibition could serve as a novel therapeutic option for NPC, and the overexpression of WDR5 might be partially attributed to miR-31-5p silencing in NPC. However, whether there are other mechanisms for WDR5 deregulation in NPC remains unclear. For example, it would be interesting to evaluate the possible regulation of EBV latent products in relation to WDR5. In addition, the possible interactions between WDR5 and major deregulated pathways of NPC require further investigation. Although there have been no reports of the direct miRNA regulation of WDR5, the association of WDR5 with lncRNA has been demonstrated. Recently, Sun et al. reported that lncRNA GClnc1 promoted gastric carcinogenesis by acting as a modular scaffold of WDR5 and KAT2A complexes. 37 However, the potential interaction between lncRNA and WDR5 in NPC remains unknown.
The aberrant activation and important oncogenic roles of Notch signaling have been reported in NPC. 32,35,36 Specifically, Notch ligands (DLL4 or JAG1) and a Notch downstream molecule (HEY1) were shown to be upregulated in the majority of EBV-positive tumor cells and primary tumors 32,35 and activation of Notch receptors (Notch3, Notch1, and Notch4) was also demonstrated. 32,35 Interestingly, the reports of both Man et al. 32 and Chen et al. 35 revealed that the Notch signaling inhibitor DAPT, a γ-secretase inhibitor (GSI), failed to inhibit aberrant Notch3 activation in NPC cell lines. The reason for the inability of GSI to inhibit Notch3 remains unknown. One possible explanation is that Notch3 intracellular domain accumulation in NPC cells is γ-secretase independent. 32,35 This resistance of Notch3 receptor maturation to γ-secretase inhibition has restricted the application of GSI for the treatment of NPC. Our results indicated that WDR5 knockdown decreased Notch3 levels in NPC cell lines, providing an alternative pathway to overcome the GSI resistance of Notch3.
In summary, our results suggest the feasibility of mc-oriP-miR-31 as a novel miRNA therapy approach for NPC. This link between miR-31-5p and WDR5 identifies a promising therapeutic target for NPC.
Footnotes
Acknowledgments
We thank Rong Geng, Ling Zhou, and Chen Yao for their helpful assistance. This work was supported by the National Natural Science Foundation of China (Nos. 81272513 and 81272638).
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.